
Oral Administration of Propolis and Lysozyme Combination Improves Feline Oral Health and Modulates Systemic Inflammatory and Oxidative Responses

 , , , , ,
, , , , ,
Abstract
1. Introduction
2. Materials and Methods
2.1. Experimental Animals and Experimental Design
2.2. Measurement of VSC Concentration
2.3. Cat Supragingival Flora Collection
2.4. Oral Symptoms Indices
2.5. 16S rRNA Sequencing of Supragingival Samples
2.6. Blood Collection and Testing
2.7. Statistical Analysis
3. Results
3.1. Baseline Characteristics of Participating Cats
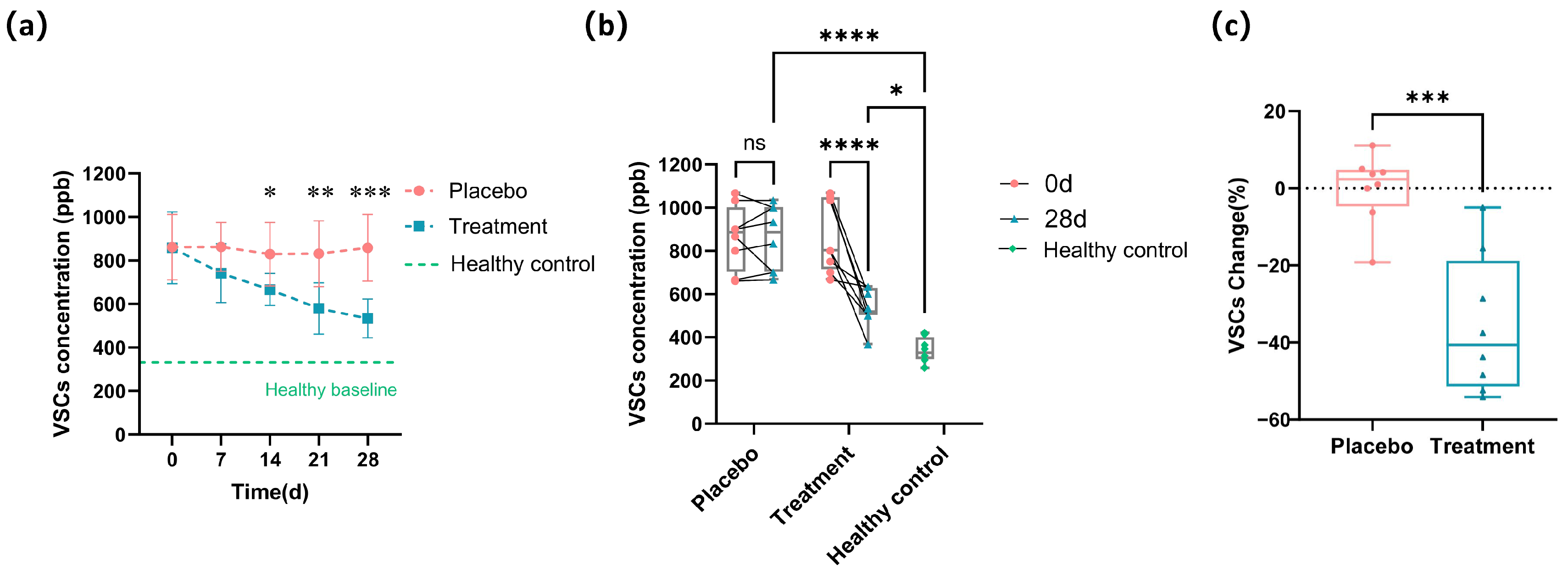
3.2. Improvement of VSCs in Cats with Halitosis by PL Intervention
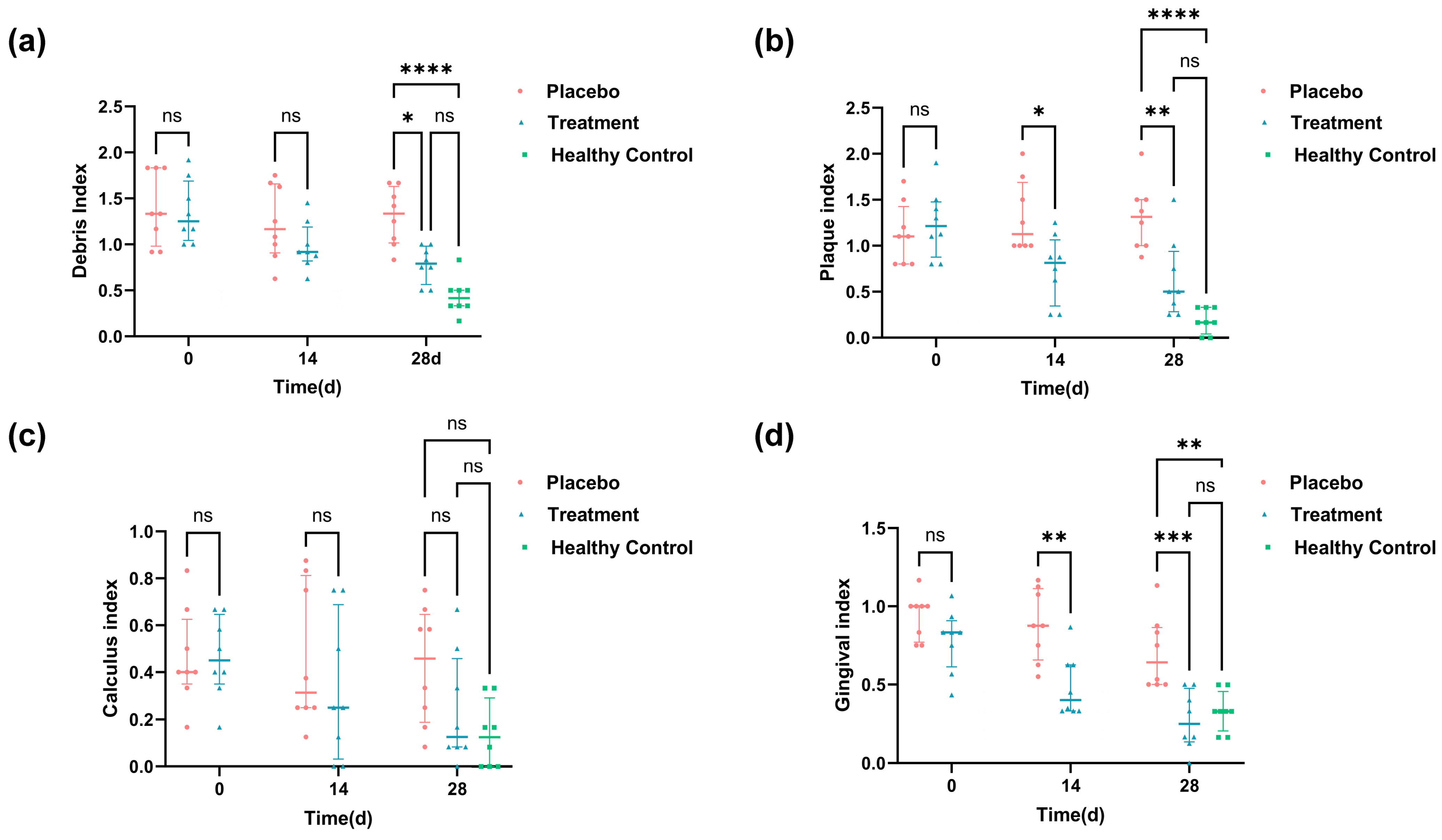
3.3. Oral Symptom Improvement in Response to PL Supplementations
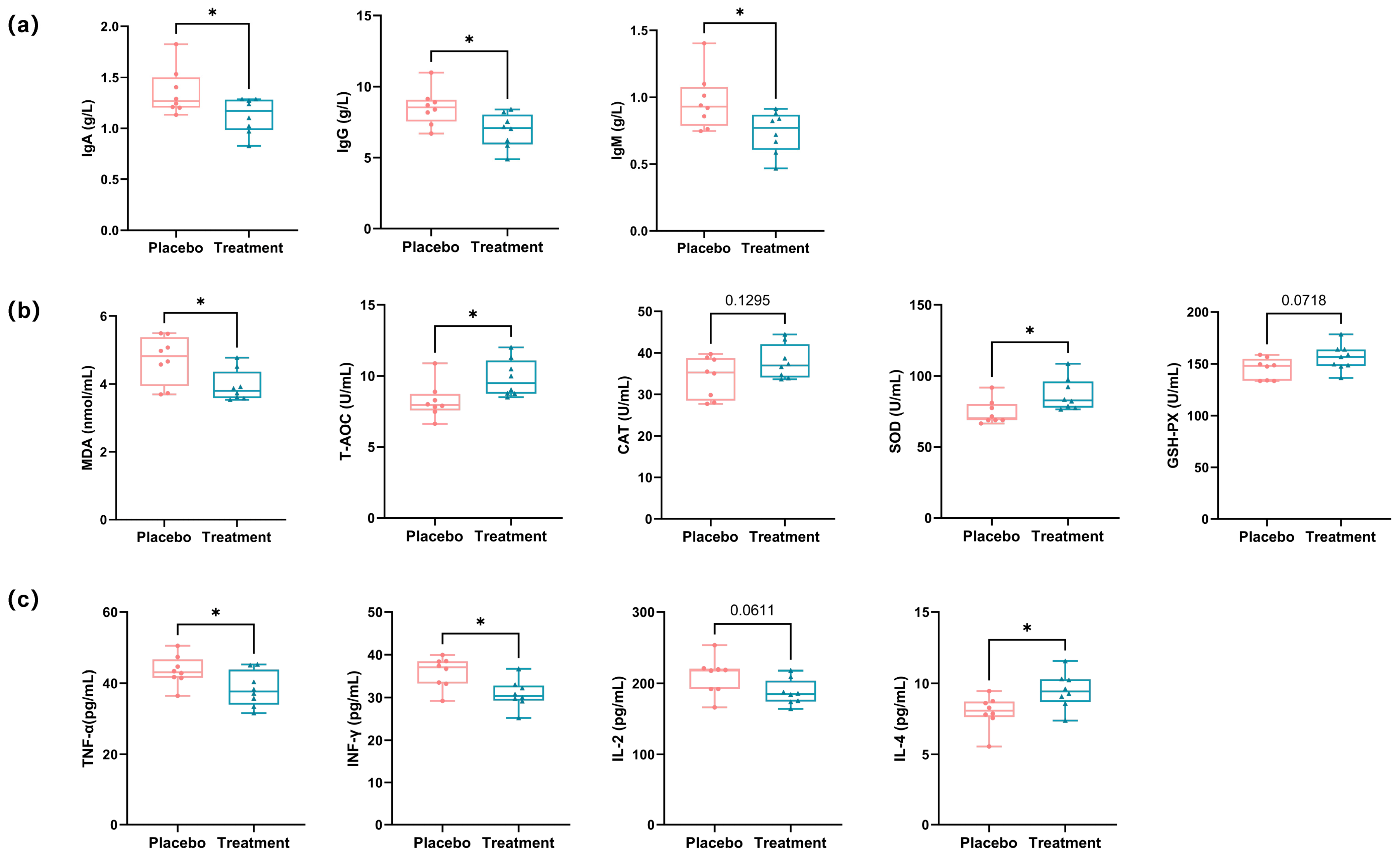
3.4. Effects of Propolis and Lysozyme Ingestion on Immune Responses, Oxidative Stress Levels, and Inflammatory Markers in Domestic Cats
3.5. Taxonomic Composition of Cat Gingival Microbiome
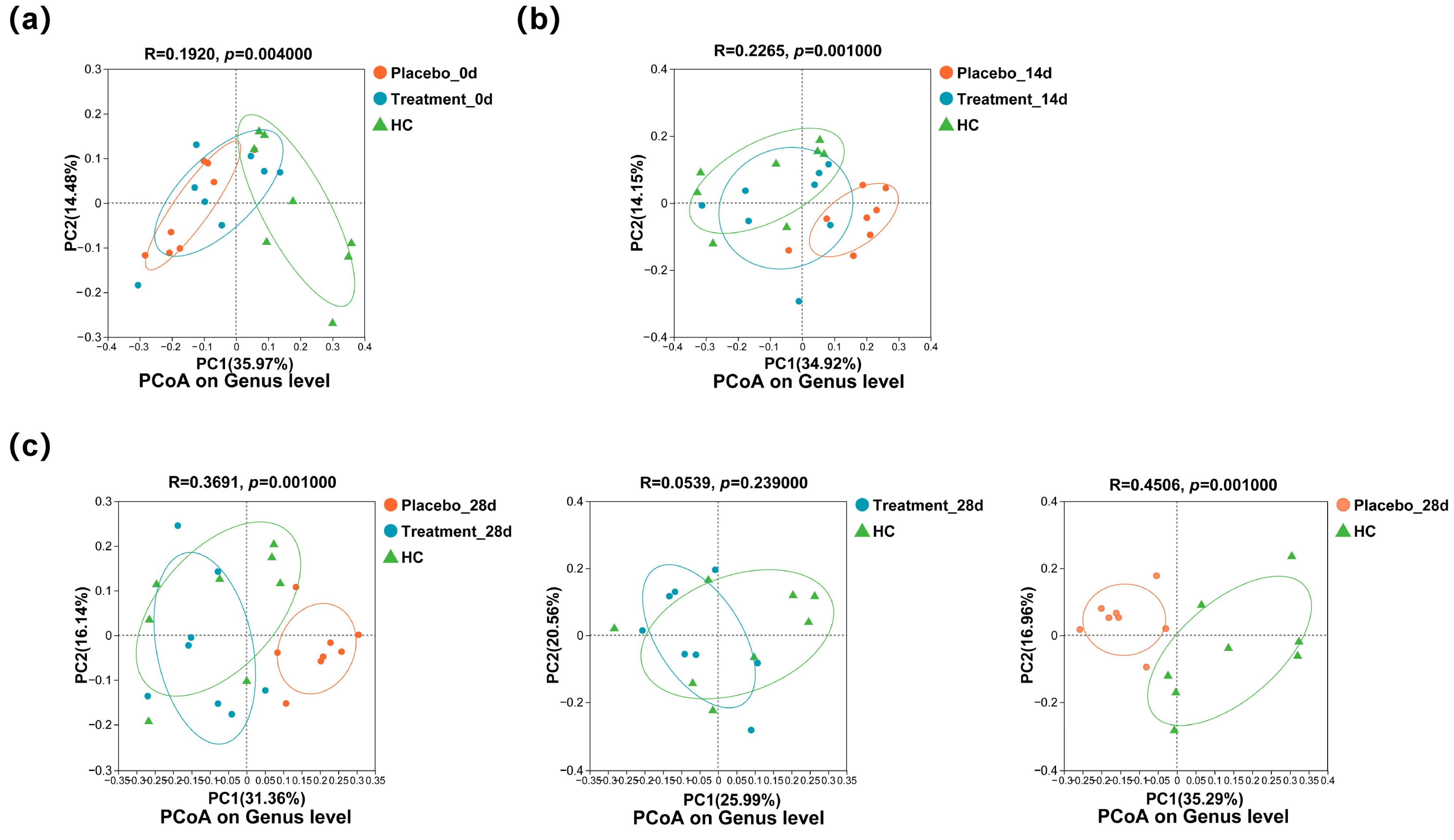
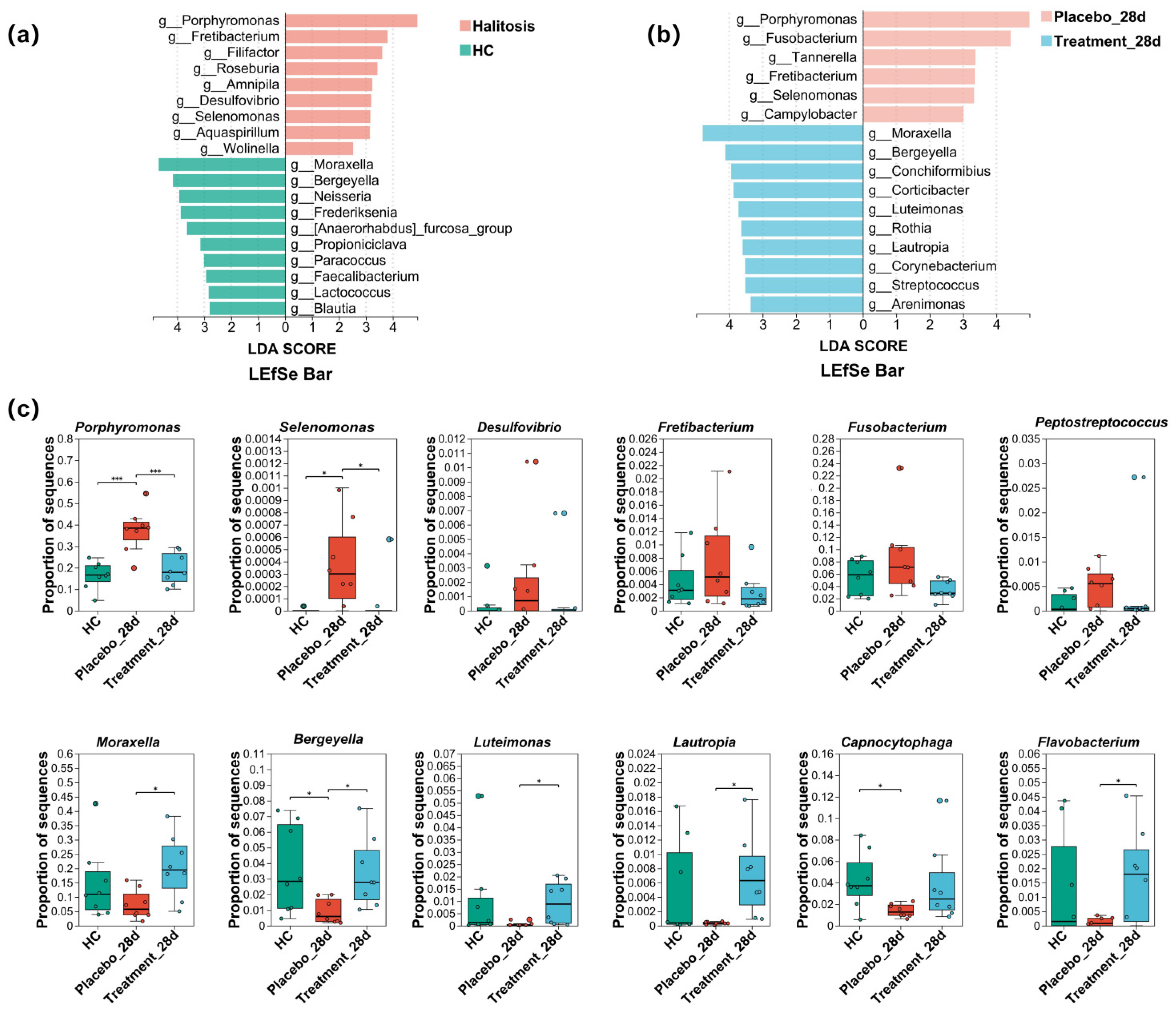
3.6. Changes in the Distribution of Microbiome on the Gingiva of Cats with Halitosis Associated with PL Intervention
3.7. Characteristic Taxa Variation in the Gingival Microbiome in Cats with Halitosis
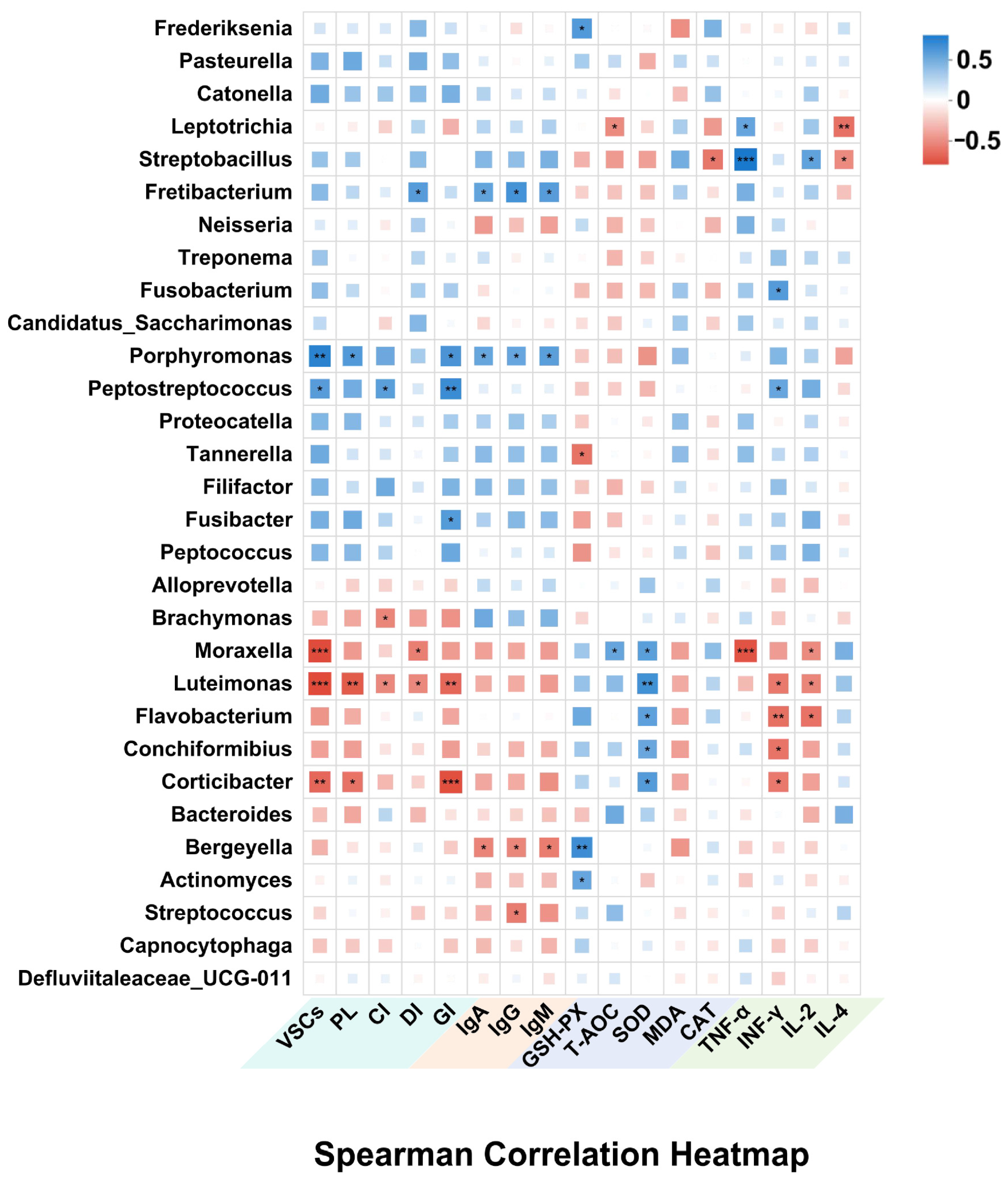
3.8. Correlation Between Oral Microbiota Composition and Systemic Physiological Indicators
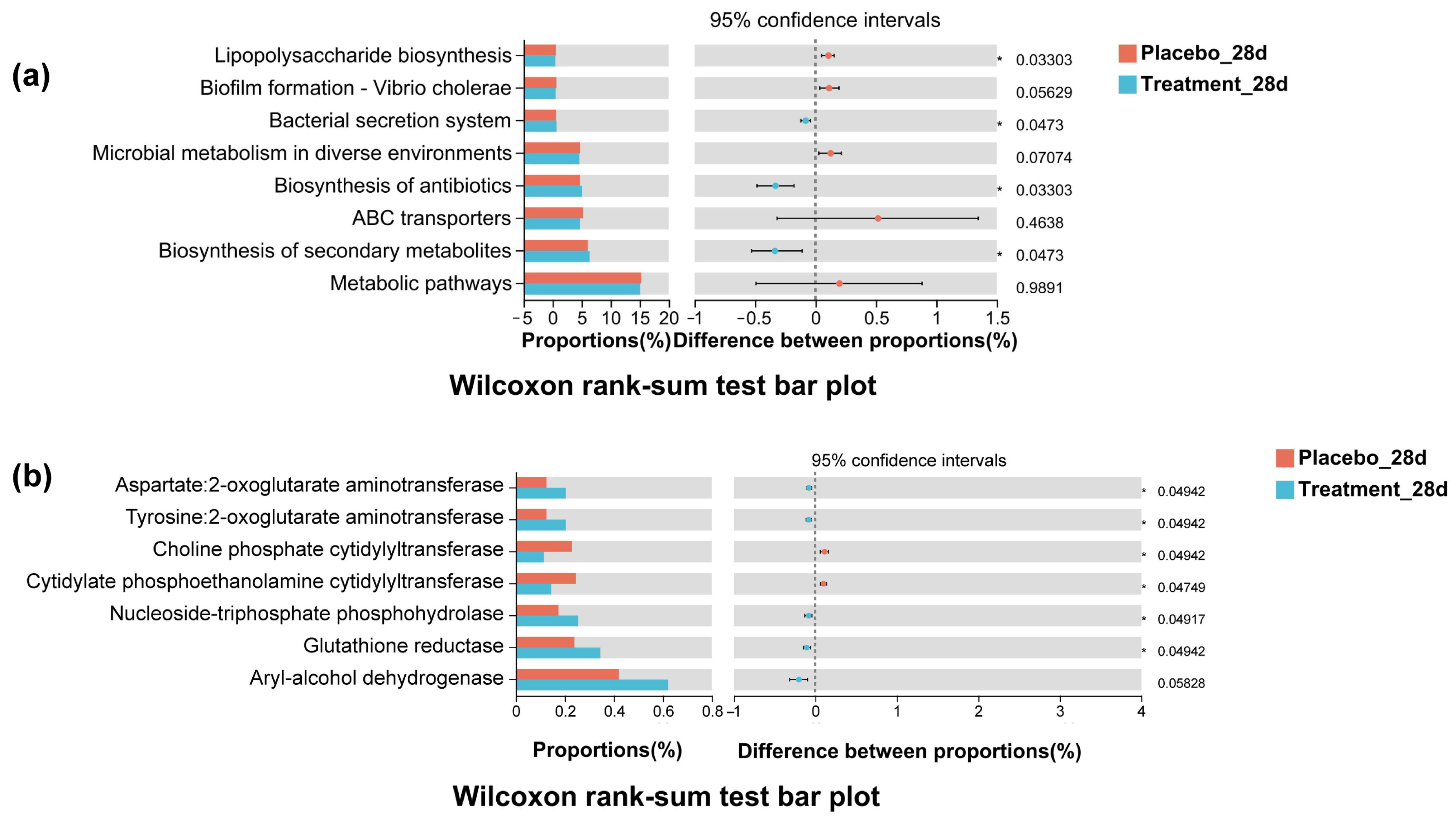
3.9. Functional Prediction of Oral Microbiota via Tax4Fun
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
Abbreviations
| PL | Combining propolis and lysozyme |
| VSCs | Volatile sulfur compounds |
| HC | Health control group |
| PLI | Turesky–Gilmore–Glickman modification of the Quigley–Hein plaque index |
| CI | Ramfjord calculus index |
| GI | Modified Loe and Silness gingival index |
| DI | Greene–Vermillion debris index |
| IgA | Immunoglobulin A |
| IgG | Immunoglobulin G |
| IgM | Immunoglobulin M |
| MDA | Malondialdehyde |
| SOD | Superoxide dismutase |
| T-AOC | Total antioxidant capacity |
| CAT | Catalase |
| GSH-PX | Glutathione peroxidase |
| TNF-α | Tumor necrosis factor-α |
| IL-2 | Interleukin-2 |
| IL-4 | Interleukin-4 |
| IFN-γ | Interferon-γ |
| PCoA | Principal coordinate analysis |
| LEfSe | Linear discriminant analysis effect size |
| LDA | Linear discriminant analysis |
References
- Buckley, C.; Colyer, A.; Skrzywanek, M.; Jodkowska, K.; Kurski, G.; Gawor, J.; Ceregrzyn, M. The impact of home-prepared diets and home oral hygiene on oral health in cats and dogs. Br. J. Nutr. 2011, 106, S124–S127. [Google Scholar] [CrossRef]
- Zhang, M.; Cui, Y.; Mei, X.; Li, L.; Wang, H.; Li, Y.; Wu, Y. Effect of Dietary Composite Probiotic Supplementation on the Microbiota of Different Oral Sites in Cats. Vet. Sci. 2024, 11, 351. [Google Scholar] [CrossRef]
- Rajasekaran, J.J.; Krishnamurthy, H.K.; Bosco, J.; Jayaraman, V.; Krishna, K.; Wang, T.H.; Bei, K. Oral Microbiome: A Review of Its Impact on Oral and Systemic Health. Microorganisms 2024, 12, 1797. [Google Scholar] [CrossRef] [PubMed]
- Giordano-Kelhoffer, B.; Lorca, C.; Llanes, J.M.; Rábano, A.; del Ser, T.; Serra, A.; Gallart-Palau, X. Oral Microbiota, Its Equilibrium and Implications in the Pathophysiology of Human Diseases: A Systematic Review. Biomedicines 2022, 10, 1803. [Google Scholar] [CrossRef]
- Bel’skaya, L.V.; Sarf, E.A.; Shalygin, S.P.; Postnova, T.V.; Kosenok, V.K. Identification of salivary volatile organic compounds as potential markers of stomach and colorectal cancer: A pilot study. J. Oral Biosci. 2020, 62, 212–221. [Google Scholar] [CrossRef]
- Takeshita, T.; Suzuki, N.; Nakano, Y.; Yasui, M.; Yoneda, M.; Shimazaki, Y.; Hirofuji, T.; Yamashita, Y. Discrimination of the oral microbiota associated with high hydrogen sulfide and methyl mercaptan production. Sci. Rep. 2012, 2, 215. [Google Scholar] [CrossRef]
- Xin, X.; Junzhi, H.; Xuedong, Z. Oral microbiota: A promising predictor of human oral and systemic diseases. West China J. Stomatol. 2015, 33, 555–560. [Google Scholar] [CrossRef]
- Xiao, E.; Mattos, M.; Vieira, G.H.A.; Chen, S.S.; Correa, J.D.; Wu, Y.Y.; Albiero, M.L.; Bittinger, K.; Graves, D.T. Diabetes Enhances IL-17 Expression and Alters the Oral Microbiome to Increase Its Pathogenicity. Cell Host Microbe 2017, 22, 120–128. [Google Scholar] [CrossRef]
- James, P.; Worthington, H.V.; Parnell, C.; Harding, M.; Lamont, T.; Cheung, A.; Whelton, H.; Riley, P. Chlorhexidine mouthrinse as an adjunctive treatment for gingival health. Cochrane Database Syst. Rev. 2017, 197, CD008676. [Google Scholar] [CrossRef]
- Ohira, C.; Kaneki, M.; Shirao, D.; Kurauchi, N.; Fukuyama, T. Oral treatment with catechin isolated from Japanese green tea significantly inhibits the growth of periodontal pathogen Porphyromonas gulae and ameliorates the gingivitis and halitosis caused by periodontal disease in cats and dogs. Int. Immunopharmacol. 2025, 146, 113805. [Google Scholar] [CrossRef] [PubMed]
- El-Newary, S.A.; Abd Elkarim, A.S.; Abdelwahed, N.A.M.; Omer, E.A.; Elgamal, A.M.; Elsayed, W.M. Chenopodium murale Juice Shows Anti-Fungal Efficacy in Experimental Oral Candidiasis in Immunosuppressed Rats in Relation to Its Chemical Profile. Molecules 2023, 28, 4304. [Google Scholar] [CrossRef]
- Min, Y.; Xiang, Y.; Hu, Z.; Mao, M.; Xia, L. Effect of Novel Oral Bacteriostatic Agents Based on Antimicrobial Peptides on Oral Bacterial Biofilms. J. Oral Sci. Res. 2023, 39, 1050–1055. [Google Scholar] [CrossRef]
- Ferraboschi, P.; Ciceri, S.; Grisenti, P. Applications of Lysozyme, an Innate Immune Defense Factor, as an Alternative Antibiotic. Antibiotics 2021, 10, 1534. [Google Scholar] [CrossRef]
- Islam, M.S.; Aryasomayajula, A.; Selvaganapathy, P.R. A Review on Macroscale and Microscale Cell Lysis Methods. Micromachines 2017, 8, 83. [Google Scholar] [CrossRef]
- Jiang, S.; Qin, Y.; Yang, J.; Li, M.; Xiong, L.; Sun, Q. Enhanced antibacterial activity of lysozyme immobilized on chitin nanowhiskers. Food Chem. 2017, 221, 1507–1513. [Google Scholar] [CrossRef]
- Pant, K.; Sharma, A.; Chopra, H.K.; Nanda, V. Impact of biodiversification on propolis composition, functionality, and application in foods as natural preservative: A review. Food Control 2024, 155, 110097. [Google Scholar] [CrossRef]
- Przybylek, I.; Karpinski, T.M. Antibacterial Properties of Propolis. Molecules 2019, 24, 2047. [Google Scholar] [CrossRef]
- Zulhendri, F.; Chandrasekaran, K.; Kowacz, M.; Ravalia, M.; Kripal, K.; Fearnley, J.; Perera, C.O. Antiviral, Antibacterial, Antifungal, and Antiparasitic Properties of Propolis: A Review. Foods 2021, 10, 1360. [Google Scholar] [CrossRef]
- Cui, J.; Duan, X.; Ke, L.; Pan, X.; Liu, J.; Song, X.; Ma, W.; Zhang, W.; Liu, Y.; Fan, Y. Extraction, purification, structural character and biological properties of propolis flavonoids: A review. Fitoterapia 2022, 157, 105106. [Google Scholar] [CrossRef] [PubMed]
- Zhang, W.W.; Margarita, G.E.; Wu, D.; Yuan, W.Q.; Yan, S.; Qi, S.Z.; Xue, X.F.; Wang, K.; Wu, L.M. Antibacterial Activity of Chinese Red Propolis against Staphylococcus aureus and MRSA. Molecules 2022, 27, 1693. [Google Scholar] [CrossRef]
- Seemann, R.; Filippi, A.; Michaelis, S.; Lauterbach, S.; John, H.-D.; Huismann, J. Duration of effect of the mouthwash CB12 for the treatment of intra-oral halitosis: A double-blind, randomised, controlled trial. J. Breath Res. 2016, 10, 036002. [Google Scholar] [CrossRef]
- Addy, M.; Moran, J.M. Evaluation of oral hygiene products: Science is true; don’t be misled by the facts. Periodontology 2000 1997, 15, 40–51. [Google Scholar] [CrossRef]
- Oo, M.M.T.; Oo, P.H.; Saddki, N. Efficacy of 0.05% cetylpyridinium chloride mouthwash as an adjunct to toothbrushing compared with 0.12% chlorhexidine gluconate mouthwash in reducing dental plaque and gingival inflammation: A randomized control trial. Int. J. Dent. Hyg. 2023, 21, 195–202. [Google Scholar] [CrossRef]
- Kashi, M.; Noei, M.; Chegini, Z.; Shariati, A. Natural compounds in the fight against Staphylococcus aureus biofilms: A review of antibiofilm strategies. Front. Pharmacol. 2024, 15, 1491363. [Google Scholar] [CrossRef]
- Olson, K.R.; Gao, Y.; Straub, K.D. Oxidation of Hydrogen Sulfide by Quinones: How Polyphenols Initiate Their Cytoprotective Effects. Int. J. Mol. Sci. 2021, 22, 961. [Google Scholar] [CrossRef] [PubMed]
- Gawor, J.; Jank, M. Ascophyllum nodosum as a nutrient supporting oral health in dogs and cats: A review. Pol. J. Vet. Sci. 2023, 26, 511–520. [Google Scholar] [CrossRef]
- Hajishengallis, G. Periodontitis: From microbial immune subversion to systemic inflammation. Nat. Rev. Immunol. 2015, 15, 30–44. [Google Scholar] [CrossRef] [PubMed]
- Pan, Y.N.; Lv, H.L.; Zhang, F.Y.; Chen, S.X.; Cheng, Y.; Ma, S.C.; Hu, H.; Liu, X.Y.; Cai, X.Y.; Fan, F.Y.; et al. Green tea extracts alleviate acetic acid-induced oral inflammation and reconstruct oral microbial balance in mice. J. Food Sci. 2023, 88, 5291–5308. [Google Scholar] [CrossRef]
- Pärnänen, P.; Räisänen, I.T.; Sorsa, T. Oral Anti-Inflammatory and Symbiotic Effects of Fermented Lingonberry Juice-Potential Benefits in IBD. Nutrients 2024, 16, 2896. [Google Scholar] [CrossRef]
- Abu-Seida, A.M. Potential Benefits of Propolis in Large and Small Animal Practices: A Narrative Review of the Literature. World’s Vet. J. 2023, 13, 441–451. [Google Scholar] [CrossRef]
- Lamont, R.J.; Koo, H.; Hajishengallis, G. The oral microbiota: Dynamic communities and host interactions. Nat. Rev. Microbiol. 2018, 16, 745–759. [Google Scholar] [CrossRef] [PubMed]
- Rodrigues, M.X.; Bicalho, R.C.; Fiani, N.; Lima, S.F.; Peralta, S. The subgingival microbial community of feline periodontitis and gingivostomatitis: Characterization and comparison between diseased and healthy cats. Sci. Rep. 2019, 9, 12340. [Google Scholar] [CrossRef]
- Anderson, J.G.; Rojas, C.A.; Scarsella, E.; Entrolezo, Z.; Jospin, G.; Hoffman, S.L.; Force, J.; Maclellan, R.H.; Peak, M.; Shope, B.H.; et al. The Oral Microbiome across Oral Sites in Cats with Chronic Gingivostomatitis, Periodontal Disease, and Tooth Resorption Compared with Healthy Cats. Animals 2023, 13, 3544. [Google Scholar] [CrossRef]
- Chaiyasut, C.; Sivamaruthi, B.S.; Bharathi, M.; Tansrisook, C.; Peerajan, S.; Chaiyasut, K.; Khongtan, S.; Tanongpitchayes, K.; Thongma, N.; Chawnan, N.; et al. Influence of Gallic Acid-Containing Mouth Spray on Dental Health and Oral Microbiota of Healthy Cats-A Pilot Study. Vet. Sci. 2022, 9, 313. [Google Scholar] [CrossRef] [PubMed]
- de Aguiar, S.C.; Zeoula, L.M.; do Prado, O.P.P.; Arcuri, P.B.; Forano, E. Characterization of rumen bacterial strains isolated from enrichments of rumen content in the presence of propolis. WORLD J. Microbiol. Biotechnol. 2014, 30, 2917–2926. [Google Scholar] [CrossRef]
- Liu, B.H.; Piao, X.H.; Niu, W.; Zhang, Q.Y.; Ma, C.; Wu, T.; Gu, Q.C.; Cui, T.F.; Li, S.D. Kuijieyuan Decoction Improved Intestinal Barrier Injury of Ulcerative Colitis by Affecting TLR4-Dependent PI3K/AKT/NF-κB Oxidative and Inflammatory Signaling and Gut Microbiota. Front. Pharmacol. 2020, 11, 1036. [Google Scholar] [CrossRef]
- Mei, F.; Xie, M.; Huang, X.; Long, Y.; Lu, X.; Wang, X.; Chen, L. Porphyromonas gingivalis and Its Systemic Impact: Current Status. Pathogens 2020, 9, 944. [Google Scholar] [CrossRef]
- Wu, P.; Zhang, J.; Zhao, L. Research progress in mechanism of oxidative stress tolerance in Porphyromonas gingivalis. Chin. J. Microbiol. Immunol. 2020, 40, 961–966. [Google Scholar] [CrossRef]
- Kushkevych, I.; Coufalova, M.; Vitezova, M.; Rittmann, S.K.M.R. Sulfate-Reducing Bacteria of the Oral Cavity and Their Relation with Periodontitis-Recent Advances. J. Clin. Med. 2020, 9, 2347. [Google Scholar] [CrossRef]
- Srila, W.; Sripilai, K.; Binlateh, T.; Thammanichanon, P.; Tiskratok, W.; Noisa, P.; Jitprasertwong, P. Relationship Between the Salivary Microbiome and Oral Malodor Metabolites in Older Thai Individuals with Periodontitis and the Cytotoxic Effects of Malodor Compounds on Human Oral Squamous Carcinoma (HSC-4) Cells. Dent. J. 2025, 13, 36. [Google Scholar] [CrossRef]
- Sturgeon, A.; Pinder, S.L.; Costa, M.C.; Weese, J.S. Characterization of the oral microbiota of healthy cats using next-generation sequencing. Vet. J. 2014, 201, 223–229. [Google Scholar] [CrossRef]
- Nakanishi, H.; Furuya, M.; Soma, T.; Hayashiuchi, Y.; Yoshiuchi, R.; Matsubayashi, M.; Tani, H.; Sasai, K. Prevalence of microorganisms associated with feline gingivostomatitis. J. Feline Med. Surg. 2019, 21, 103–108. [Google Scholar] [CrossRef] [PubMed]
- Norris, J.M.; Love, D.N. Associations amongst three feline Porphyromonas species from the gingival margin of cats during periodontal health and disease. Vet. Microbiol. 1999, 65, 195–207. [Google Scholar] [CrossRef]
- Ye, W.; Wu, J.; Jiang, Q.; Su, Z.; Liao, H.; Liu, Z.; Tao, R.; Yong, X. Antibacterial activity of corydalis saxicola bunting total alkaloids against Porphyromonas gingivalis in vitro. Future Microbiol. 2024, 19, 595–606. [Google Scholar] [CrossRef]
- Pinto, C.F.D.; Pignone, V.N.; Silveira, A.K.; Krebs, G.; Trevizan, L. Short-term changes in the oral microbiota of dogs after chewing different types of autoclaved bones. Vet. Arh. 2021, 91, 277–285. [Google Scholar] [CrossRef]
- Xu, J.Y.; Yu, L.; Ye, S.R.; Ye, Z.T.; Yang, L.Y.; Xu, X.X. Oral microbiota-host interaction: The chief culprit of alveolar bone resorption. Front. Immunol. 2024, 15, 1254516. [Google Scholar] [CrossRef]
- Gualtero, D.F.; Lafaurie, G.I.; Buitrago, D.M.; Castillo, Y.; Vargas-Sanchez, P.K.; Castillo, D.M. Oral microbiome mediated inflammation, a potential inductor of vascular diseases: A comprehensive review. Front. Cardiovasc. Med. 2023, 10, 1250263. [Google Scholar] [CrossRef]
- Klimesova, K.; Zakostelska, Z.J.; Tlaskalova-Hogenova, H. Oral Bacterial and Fungal Microbiome Impacts Colorectal Carcinogenesis. Front. Microbiol. 2018, 9, 774. [Google Scholar] [CrossRef]
- Luong, A.; Tawfik, A.N.; Islamoglu, H.; Gobriel, H.S.; Ali, N.; Ansari, P.; Shah, R.; Hung, T.; Patel, T.; Henson, B.; et al. Periodontitis and diabetes mellitus co-morbidity: A molecular dialogue. J. Oral Biosci. 2021, 63, 360–369. [Google Scholar] [CrossRef]
- Belstrom, D. The salivary microbiota in health and disease. J. Oral Microbiol. 2020, 12, 1723975. [Google Scholar] [CrossRef]
- Yachida, S.; Mizutani, S.; Shiroma, H.; Shiba, S.; Nakajima, T.; Sakamoto, T.; Watanabe, H.; Masuda, K.; Nishimoto, Y.; Kubo, M.; et al. Metagenomic and metabolomic analyses reveal distinct stage-specific phenotypes of the gut microbiota in colorectal cancer. Nat. Med. 2019, 25, 968. [Google Scholar] [CrossRef] [PubMed]
- Donia, M.S.; Fischbach, M.A. Small molecules from the human microbiota. Science 2015, 349, 1254766. [Google Scholar] [CrossRef] [PubMed]
- Forman, H.J.; Zhang, H.Q.; Rinna, A. Glutathione: Overview of its protective roles, measurement, and biosynthesis. Mol. Asp. Med. 2009, 30, 1–12. [Google Scholar] [CrossRef] [PubMed]
- Cardia, G.F.E.; Silva-Comar, F.M.d.S.; Bonetti, C.I.; da Rocha, E.M.T.; Zagoto, M.; do Amaral, V.; Bracht, L.; Silva-Filho, S.E.; Bersani-Amado, C.A.; Cuman, R.K.N. Hepatoprotective effect of beta-myrcene pretreatment against acetaminophen-induced liver injury. Avicenna J. Phytomed. 2022, 12, 388–400. [Google Scholar] [CrossRef]


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Health Control Group | Placebo Group | Treatment Group | p-Value | |
|---|---|---|---|---|
| Number | 8 | 8 | 8 | |
| Gender (Male/Female) | 4/4 | 4/4 | 4/4 | |
| Age (month; mean ± s.d.) | 12.65 ± 3.02 | 13.25 ± 1.91 | 13.38 ± 2.5 | 0.8174 |
| Weight (kg; mean ± s.d.) | 3.00 ± 0.78 | 3.08 ± 0.83 | 2.84 ± 0.9 | 0.8481 |
| VSC concentration (ppm; mean ± s.d.) | 332.38 ± 56.22 | 861.67 ± 14.97 | 858.33 ± 165.23 | 0.9669 (P vs. T) |
| 0.0000 (HC vs. P and T) |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2025 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Wang, L.; Chen, Q.; Wang, W.; Dong, H.; Chang, X.; Chen, L.; Wang, R.; Chen, Y.; Wang, P.; Chen, S.; et al. Oral Administration of Propolis and Lysozyme Combination Improves Feline Oral Health and Modulates Systemic Inflammatory and Oxidative Responses. Antioxidants 2025, 14, 639. https://doi.org/10.3390/antiox14060639
Wang L, Chen Q, Wang W, Dong H, Chang X, Chen L, Wang R, Chen Y, Wang P, Chen S, et al. Oral Administration of Propolis and Lysozyme Combination Improves Feline Oral Health and Modulates Systemic Inflammatory and Oxidative Responses. Antioxidants. 2025; 14(6):639. https://doi.org/10.3390/antiox14060639
Chicago/Turabian StyleWang, Longjiao, Qianqian Chen, Weiwei Wang, Hao Dong, Xiaohan Chang, Lishui Chen, Ran Wang, Yaoxing Chen, Pengjie Wang, Shuxing Chen, and et al. 2025. "Oral Administration of Propolis and Lysozyme Combination Improves Feline Oral Health and Modulates Systemic Inflammatory and Oxidative Responses" Antioxidants 14, no. 6: 639. https://doi.org/10.3390/antiox14060639
APA StyleWang, L., Chen, Q., Wang, W., Dong, H., Chang, X., Chen, L., Wang, R., Chen, Y., Wang, P., Chen, S., Xiong, W., & Li, Y. (2025). Oral Administration of Propolis and Lysozyme Combination Improves Feline Oral Health and Modulates Systemic Inflammatory and Oxidative Responses. Antioxidants, 14(6), 639. https://doi.org/10.3390/antiox14060639