
Success and Controversy of Natural Products as Therapeutic Modulators of Wnt Signaling and Its Interplay with Oxidative Stress: Comprehensive Review Across Compound Classes and Experimental Systems

 , , ,
, , , 
Abstract
1. Introduction
- Almost all major cancer types, such as esophageal, breast, lung (predominantly non-small-cell lung carcinoma, NSCLC), liver, colorectal, ovarian, prostate cancers, as well as osteosarcoma, leukemia, melanoma, and glioma [1];
- Involved in cardiac maintenance and recovery, particularly cardiac hypertrophy [21];
- Influences key cell types in the hair follicle to promote hair growth [22];
- Activation of the cascade impairs re-epithelialization by keratinocytes, impeding wound healing [25];
- Pathway inhibition is useful to protect against the consequences of retinal ischemia [26].
- Effects on DNA and protein integrity: RO/NS are capable of directly damaging DNA bases, causing strand breaks and adducts, which, in turn, may cause mutations that lead to genomic instability and may accelerate the carcinogenesis process [40]. Depending on the exact levels and nature, there could also be quantitative effects on protein integrity and cross-linking. Obviously, this effect is mostly directly counteracted by the scavenging ability of antioxidant compounds [40,41].
- Impact on ferroptosis: This type of cell death is characterized by iron-dependent lipid peroxidation. The formation of lipid hydroperoxides initiates the process, which ultimately leads to cell membrane damage and cell death in the presence of iron [42,43]. Fenton-like reactions are then activated to produce lipid radicals [42]. Natural antioxidants can affect ferroptosis in a number of ways, most notably by directly scavenging lipid peroxyl radicals, acting as chain-breaking antioxidants [42], or by altering intracellular GSH levels [43].
2. Review
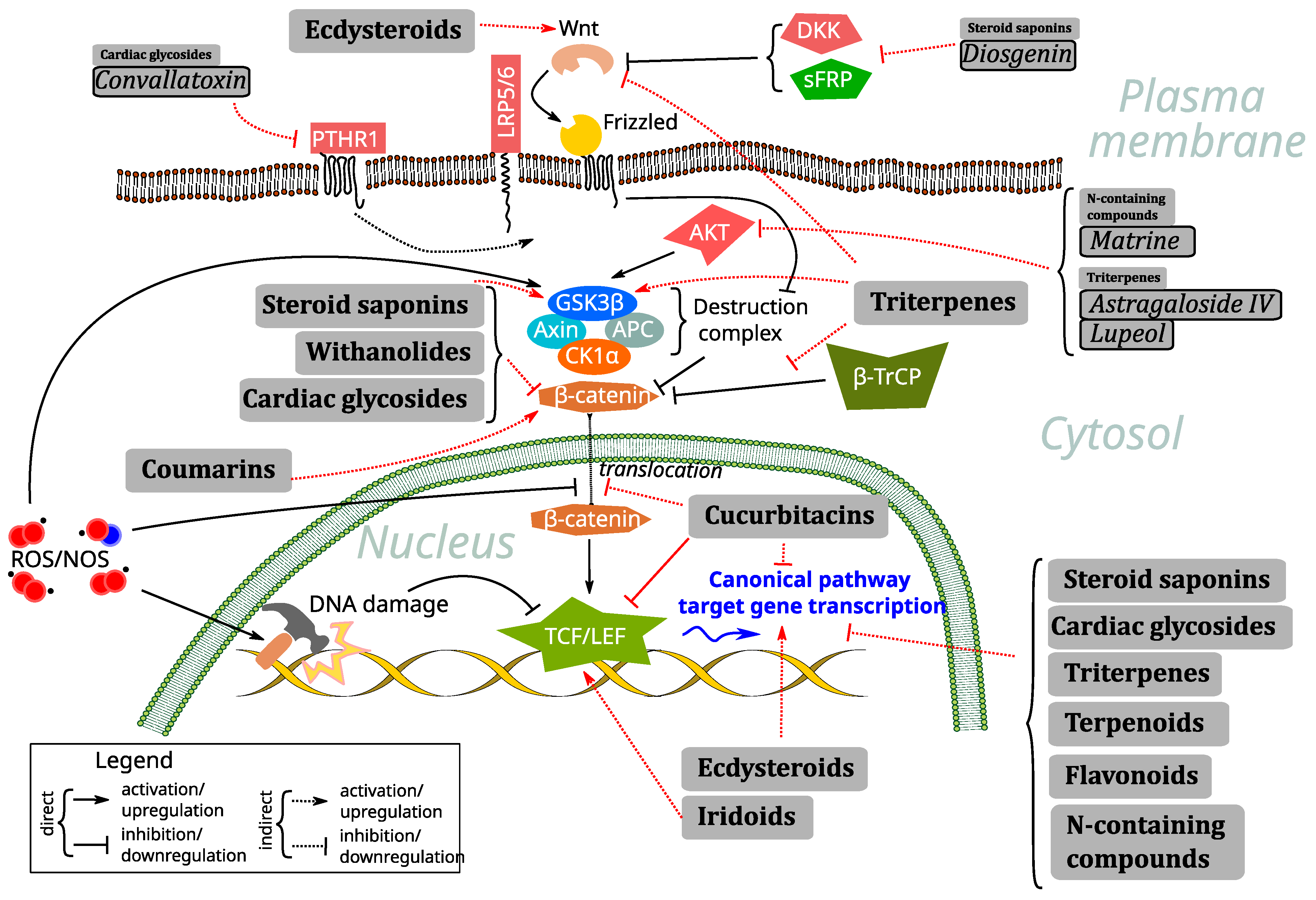
2.1. Natural Compounds as Mediators and Regulators of Wnt Signaling via Oxidative Stress and ROS Regulation
2.2. Cardiac Glycosides
2.3. Steroid Saponins
2.4. Ecdysteroids
2.5. Withanolides
2.6. Cucurbitacins
2.7. Triterpenes
2.8. Iridoids
2.9. Terpenoids
2.10. Coumarins
2.11. Flavonoids and Phenolics
2.12. N-Containing Compounds
3. Conclusions and Perspectives
Author Contributions
Funding
Conflicts of Interest
Abbreviations
| NSCLC | Non-small-cell lung carcinoma |
| FZD | Frizzled |
| DMEM | Dulbecco’s modified eagle medium |
| TNBC | Triple-negative breast cancer |
| CCRK | Cell-cycle-related kinase |
| EMT | Epithelial–mesenchymal transition |
| MBP | Myelin-binding protein |
| ERα | Estrogen receptor α |
| HCC | Hepatocellular carcinoma |
References
- Jung, Y.-S.; Park, J.-I. Wnt Signaling in Cancer: Therapeutic Targeting of Wnt Signaling beyond β-Catenin and the Destruction Complex. Exp. Mol. Med. 2020, 52, 183–191. [Google Scholar] [CrossRef] [PubMed]
- Amaral, L.S.; Martins Ferreira, J.; Predes, D.; Abreu, J.G.; Noël, F.; Quintas, L.E.M. Telocinobufagin and Marinobufagin Produce Different Effects in LLC-PK1 Cells: A Case of Functional Selectivity of Bufadienolides. Int. J. Mol. Sci. 2018, 19, 2769. [Google Scholar] [CrossRef]
- Zhou, P.; Zhang, X.; Guo, M.; Guo, R.; Wang, L.; Zhang, Z.; Lin, Z.; Dong, M.; Dai, H.; Ji, X.; et al. Ginsenoside Rb1 Ameliorates CKD-Associated Vascular Calcification by Inhibiting the Wnt/β-Catenin Pathway. J. Cell. Mol. Med. 2019, 23, 7088–7098. [Google Scholar] [CrossRef]
- Wang, M.; Chen, D.-Q.; Chen, L.; Liu, D.; Zhao, H.; Zhang, Z.-H.; Vaziri, N.D.; Guo, Y.; Zhao, Y.-Y.; Cao, G. Novel RAS Inhibitors Poricoic Acid ZG and Poricoic Acid ZH Attenuate Renal Fibrosis via a Wnt/β-Catenin Pathway and Targeted Phosphorylation of Smad3 Signaling. J. Agric. Food Chem. 2018, 66, 1828–1842. [Google Scholar] [CrossRef]
- Gao, H.; Ruan, T.; Xing, M.; Chen, Y.; Bai, S.; Liu, J.; Yu, X.; Feng, J.; Xu, X.; Wang, Q. Catalpol Attenuates EMT by Inhibiting Wnt/β-Catenin and TGF-β/Smads Signaling to Alleviate Kidney Fibrosis. Trop. J. Pharm. Res. 2024, 23, 57–65. [Google Scholar] [CrossRef]
- Zhang, C.; Peng, J.; Wu, S.; Jin, Y.; Xia, F.; Wang, C.; Liu, K.; Sun, H.; Liu, M. Dioscin Promotes Osteoblastic Proliferation and Differentiation via Lrp5 and ER Pathway in Mouse and Human Osteoblast-like Cell Lines. J. Biomed. Sci. 2014, 21, 30. [Google Scholar] [CrossRef]
- Li, M.; Zhang, N.-D.; Wang, Y.; Han, T.; Jiang, Y.-P.; Rahman, K.; Qin, L.-P.; Xin, H.-L.; Zhang, Q.-Y. Coordinate Regulatory Osteogenesis Effects of Icariin, Timosaponin B II and Ferulic Acid from Traditional Chinese Medicine Formulas on UMR-106 Osteoblastic Cells and Osteoblasts in Neonatal Rat Calvaria Cultures. J. Ethnopharmacol. 2016, 185, 120–131. [Google Scholar] [CrossRef]
- Yan, C.-P.; Wang, X.-K.; Jiang, K.; Yin, C.; Xiang, C.; Wang, Y.; Pu, C.; Chen, L.; Li, Y.-L. β-Ecdysterone Enhanced Bone Regeneration Through the BMP-2/SMAD/RUNX2/Osterix Signaling Pathway. Front. Cell Dev. Biol. 2022, 10, 883228. [Google Scholar] [CrossRef]
- Wang, Y.-Q.; Wang, N.-X.; Luo, Y.; Yu, C.-Y.; Xiao, J.-H. Ganoderal A Effectively Induces Osteogenic Differentiation of Human Amniotic Mesenchymal Stem Cells via Cross-Talk between Wnt/β-Catenin and BMP/SMAD Signaling Pathways. Biomed. Pharmacother. 2020, 123, 109807. [Google Scholar] [CrossRef]
- Ma, T.; Ruan, H.; Lv, L.; Wei, C.; Yu, Y.; Jia, L.; Song, X.; Zhang, J.; Li, Y. Oleanolic Acid, a Small-Molecule Natural Product, Inhibits ECM Degeneration in Osteoarthritis by Regulating the Hippo/YAP and Wnt/β-Catenin Pathways. Food Funct. 2023, 14, 9999–10013. [Google Scholar] [CrossRef]
- Lu, J.; Zhang, T.; Sun, H.; Wang, S.; Liu, M. Protective Effects of Dioscin against Cartilage Destruction in a Monosodium Iodoacetate (MIA)-Indcued Osteoarthritis Rat Model. Biomed. Pharmacother. 2018, 108, 1029–1038. [Google Scholar] [CrossRef]
- Cai, L.; Mu, Y.-R.; Liu, M.-M.; Zhou, M.-Y.; Meng, B.; Liu, F.-Y.; Li, R. Penta-Acetyl Geniposide Suppresses Migration, Invasion, and Inflammation of TNF-α-Stimulated Rheumatoid Arthritis Fibroblast-Like Synoviocytes Involving Wnt/β-Catenin Signaling Pathway. Inflammation 2021, 44, 2232–2245. [Google Scholar] [CrossRef]
- Gautam, J.; Khedgikar, V.; Kushwaha, P.; Choudhary, D.; Nagar, G.K.; Dev, K.; Dixit, P.; Singh, D.; Maurya, R.; Trivedi, R. Formononetin, an Isoflavone, Activates AMP-Activated Protein Kinase/β-Catenin Signalling to Inhibit Adipogenesis and Rescues C57BL/6 Mice from High-Fat Diet-Induced Obesity and Bone Loss. Br. J. Nutr. 2017, 117, 645–661. [Google Scholar] [CrossRef]
- Xu, Y.; Zhang, G.; Kang, Z.; Xu, Y.; Jiang, W.; Zhang, S. Cornin Increases Angiogenesis and Improves Functional Recovery after Stroke via the Ang1/Tie2 Axis and the Wnt/β-Catenin Pathway. Arch. Pharm. Res. 2016, 39, 133–142. [Google Scholar] [CrossRef]
- Sun, F.-L.; Wang, W.; Zuo, W.; Xue, J.-L.; Xu, J.; Ai, H.-X.; Zhang, L.; Wang, X.-M.; Ji, X.-M. Promoting Neurogenesis via Wnt/β-Catenin Signaling Pathway Accounts for the Neurorestorative Effects of Morroniside against Cerebral Ischemia Injury. Eur. J. Pharmacol. 2014, 738, 214–221. [Google Scholar] [CrossRef]
- Li, S. The Wnt/β-Catenin Coupled with HIF-1α/VEGF Signal Pathway Involved Neurovascular Unit Protection by Galangin against Permanent Focal Cerebral Ischemia in Rats (LB628). FASEB J. 2014, 28, LB628. [Google Scholar] [CrossRef]
- Rong, Y.; Liu, W.; Zhou, Z.; Gong, F.; Bai, J.; Fan, J.; Li, L.; Luo, Y.; Zhou, Z.; Cai, W. Harpagide Inhibits Neuronal Apoptosis and Promotes Axonal Regeneration after Spinal Cord Injury in Rats by Activating the Wnt/β-Catenin Signaling Pathway. Brain Res. Bull. 2019, 148, 91–99. [Google Scholar] [CrossRef]
- Yuan, J.; Xu, N.; Tao, Y.; Han, X.; Yang, L.; Liang, J.; Jin, H.; Zhang, X.; Wu, H.; Shi, H.; et al. Total Astragalosides Promote Oligodendrocyte Precursor Cell Differentiation and Enhance Remyelination in Cuprizone-Induced Mice through Suppression of Wnt/β-Catenin Signaling Pathway. J. Ethnopharmacol. 2022, 298, 115622. [Google Scholar] [CrossRef]
- Zu, G.; Guo, J.; Che, N.; Zhou, T.; Zhang, X. Protective Effects of Ginsenoside Rg1 on Intestinal Ischemia/Reperfusion Injury-Induced Oxidative Stress and Apoptosis via Activation of the Wnt/β-Catenin Pathway. Sci. Rep. 2016, 6, 38480. [Google Scholar] [CrossRef]
- He, J.; Wang, M.; Yang, L.; Xin, H.; Bian, F.; Jiang, G.; Zhang, X. Astragaloside IV Alleviates Intestinal Barrier Dysfunction via the AKT-GSK3β-β-Catenin Pathway in Peritoneal Dialysis. Front. Pharmacol. 2022, 13, 873150. [Google Scholar] [CrossRef]
- Li, Q.; Jiang, W.; Wan, Z.; Ni, Y.; Lei, L.; Wei, J. Polyphyllin I Attenuates Pressure Over-Load Induced Cardiac Hypertrophy via Inhibition of Wnt/β-Catenin Signaling Pathway. Life Sci. 2020, 252, 117624. [Google Scholar] [CrossRef]
- Shin, H.-S.; Park, S.-Y.; Hwang, E.-S.; Lee, D.-G.; Song, H.-G.; Mavlonov, G.T.; Yi, T.-H. The Inductive Effect of Ginsenoside F2 on Hair Growth by Altering the WNT Signal Pathway in Telogen Mouse Skin. Eur. J. Pharmacol. 2014, 730, 82–89. [Google Scholar] [CrossRef]
- Li, X.; Liu, X.; Deng, R.; Gao, S.; Jiang, Q.; Liu, R.; Li, H.; Miao, Y.; Zhai, Y.; Zhang, S.; et al. Betulinic Acid Attenuated Bleomycin-Induced Pulmonary Fibrosis by Effectively Intervening Wnt/β-Catenin Signaling. Phytomedicine 2021, 81, 153428. [Google Scholar] [CrossRef]
- Wang, L.; Chi, Y.-F.; Yuan, Z.-T.; Zhou, W.-C.; Yin, P.-H.; Zhang, X.-M.; Peng, W. Astragaloside IV Inhibits the Up-Regulation of Wnt/β-Catenin Signaling in Rats with Unilateral Ureteral Obstruction. Cell. Physiol. Biochem. 2014, 33, 1316–1328. [Google Scholar] [CrossRef]
- Li, F.-L.; Li, X.; Wang, Y.-F.; Xiao, X.-L.; Xu, R.; Chen, J.; Fan, B.; Xu, W.-B.; Geng, L.; Li, B. Astragaloside IV Downregulates β-Catenin in Rat Keratinocytes to Counter LiCl-Induced Inhibition of Proliferation and Migration. Evid.-Based Complement. Altern. Med. ECAM 2012, 2012, 956107. [Google Scholar] [CrossRef]
- Chao, H.W.-H.; Chen, Y.-K.; Liu, J.-H.; Pan, H.-T.; Lin, H.-M.; Chao, H.-M. Emodin Protected against Retinal Ischemia Insulted Neurons through the Downregulation of Protein Overexpression of β-Catenin and Vascular Endothelium Factor. BMC Complement. Med. Ther. 2020, 20, 338. [Google Scholar] [CrossRef]
- Yu, F.; Yu, C.; Li, F.; Zuo, Y.; Wang, Y.; Yao, L.; Wu, C.; Wang, C.; Ye, L. Wnt/β-Catenin Signaling in Cancers and Targeted Therapies. Signal Transduct. Target. Ther. 2021, 6, 307. [Google Scholar] [CrossRef]
- Kim, J.-H.; Kim, Y.H.; Song, G.-Y.; Kim, D.-E.; Jeong, Y.-J.; Liu, K.-H.; Chung, Y.-H.; Oh, S. Ursolic Acid and Its Natural Derivative Corosolic Acid Suppress the Proliferation of APC-Mutated Colon Cancer Cells through Promotion of β-Catenin Degradation. Food Chem. Toxicol. 2014, 67, 87–95. [Google Scholar] [CrossRef]
- Seth, C.; Mas, C.; Conod, A.; Mueller, J.; Siems, K.; Kuciak, M.; Borges, I.; Ruiz i Altaba, A. Long-Lasting WNT-TCF Response Blocking and Epigenetic Modifying Activities of Withanolide F in Human Cancer Cells. PLoS ONE 2016, 11, e0168170. [Google Scholar] [CrossRef]
- Tao, X.; Yin, L.; Xu, L.; Peng, J. Dioscin: A Diverse Acting Natural Compound with Therapeutic Potential in Metabolic Diseases, Cancer, Inflammation and Infections. Pharmacol. Res. 2018, 137, 259–269. [Google Scholar] [CrossRef]
- Koval, A.; Pieme, C.A.; Queiroz, E.F.; Ragusa, S.; Ahmed, K.; Blagodatski, A.; Wolfender, J.-L.; Petrova, T.V.; Kat-anaev, V.L. Tannins from Syzygium Guineense Suppress Wnt Signaling and Proliferation of Wnt-Dependent Tumors through a Direct Effect on Secreted Wnts. Cancer Lett. 2018, 435, 110–120. [Google Scholar] [CrossRef]
- Liu, D.; Chen, L.; Zhao, H.; Vaziri, N.D.; Ma, S.-C.; Zhao, Y.-Y. Small Molecules from Natural Products Targeting the Wnt/β-Catenin Pathway as a Therapeutic Strategy. Biomed. Pharmacother. 2019, 117, 108990. [Google Scholar] [CrossRef]
- Devappa, R.K.; Makkar, H.P.S.; Becker, K. Nutritional, Biochemical, and Pharmaceutical Potential of Proteins and Peptides from Jatropha: Review. J. Agric. Food Chem. 2010, 58, 6543–6555. [Google Scholar] [CrossRef]
- Fatima, I.; El-Ayachi, I.; Taotao, L.; Lillo, M.A.; Krutilina, R.I.; Seagroves, T.N.; Radaszkiewicz, T.W.; Hutnan, M.; Bryja, V.; Krum, S.A.; et al. The Natural Compound Jatrophone Interferes with Wnt/β-Catenin Signaling and Inhibits Proliferation and EMT in Human Triple-Negative Breast Cancer. PLoS ONE 2017, 12, e0189864. [Google Scholar] [CrossRef]
- Wang, L.; Yang, J.; Kong, L.-M.; Deng, J.; Xiong, Z.; Huang, J.; Luo, J.; Yan, Y.; Hu, Y.; Li, X.-N.; et al. Natural and Semisynthetic Tigliane Diterpenoids with New Carbon Skeletons from Euphorbia Dracunculoides as a Wnt Signaling Pathway Inhibitor. Org. Lett. 2017, 19, 3911–3914. [Google Scholar] [CrossRef]
- Ohishi, K.; Toume, K.; Arai, M.A.; Sadhu, S.K.; Ahmed, F.; Mizoguchi, T.; Itoh, M.; Ishibashi, M. Ricinine: A Pyridone Alkaloid from Ricinus Communis That Activates the Wnt Signaling Pathway through Casein Kinase 1α. Bioorg. Med. Chem. 2014, 22, 4597–4601. [Google Scholar] [CrossRef]
- Xiao, X.; Ao, M.; Xu, F.; Li, X.; Hu, J.; Wang, Y.; Li, D.; Zhu, X.; Xin, C.; Shi, W. Effect of Matrine against Breast Cancer by Downregulating the Vascular Endothelial Growth Factor via the Wnt/β-Catenin Pathway. Oncol. Lett. 2018, 15, 1691–1697. [Google Scholar] [CrossRef]
- Zhang, R.; Yang, J.; Wu, J.; Xiao, L.; Miao, L.; Qi, X.; Li, Y.; Sun, W. Berberine Promotes Osteogenic Differentiation of Mesenchymal Stem Cells with Therapeutic Potential in Periodontal Regeneration. Eur. J. Pharmacol. 2019, 851, 144–150. [Google Scholar] [CrossRef]
- Yu, W.-K.; Xu, Z.-Y.; Yuan, L.; Mo, S.; Xu, B.; Cheng, X.-D.; Qin, J.-J. Targeting β-Catenin Signaling by Natural Products for Cancer Prevention and Therapy. Front. Pharmacol. 2020, 11, 984. [Google Scholar] [CrossRef]
- Reddy, V.P. Oxidative Stress in Health and Disease. Biomedicines 2023, 11, 2925. [Google Scholar] [CrossRef]
- Juan, C.A.; Pérez de la Lastra, J.M.; Plou, F.J.; Pérez-Lebeña, E. The Chemistry of Reactive Oxygen Species (ROS) Revisited: Outlining Their Role in Biological Macromolecules (DNA, Lipids and Proteins) and Induced Pathologies. Int. J. Mol. Sci. 2021, 22, 4642. [Google Scholar] [CrossRef] [PubMed]
- Kajarabille, N.; Latunde-Dada, G.O. Programmed Cell-Death by Ferroptosis: Antioxidants as Mitigators. Int. J. Mol. Sci. 2019, 20, 4968. [Google Scholar] [CrossRef]
- Kuang, F.; Liu, J.; Tang, D.; Kang, R. Oxidative Damage and Antioxidant Defense in Ferroptosis. Front. Cell Dev. Biol. 2020, 8, 586578. [Google Scholar] [CrossRef]
- Bartosz, G. Reactive Oxygen Species: Destroyers or Messengers? Biochem. Pharmacol. 2009, 77, 1303–1315. [Google Scholar] [CrossRef] [PubMed]
- Schieber, M.; Chandel, N.S. ROS Function in Redox Signaling and Oxidative Stress. Curr. Biol. 2014, 24, R453–R462. [Google Scholar] [CrossRef]
- Zuo, J.; Zhang, Z.; Luo, M.; Zhou, L.; Nice, E.C.; Zhang, W.; Wang, C.; Huang, C. Redox Signaling at the Crossroads of Human Health and Disease. MedComm 2022, 3, e127. [Google Scholar] [CrossRef]
- Banuls, C.; Rocha, M.; Rovira-Llopis, S.; Falcon, R.; Castello, R.; Herance, J.R.; Polo, M.; Blas-Garcia, A.; Hernandez-Mijares, A.; Victor, V.M. The Pivotal Role of Nitric Oxide: Effects on the Nervous and Immune Systems. Curr. Pharm. Des. 2014, 20, 4679–4689. [Google Scholar] [CrossRef]
- Radak, Z.; Zhao, Z.; Koltai, E.; Ohno, H.; Atalay, M. Oxygen Consumption and Usage during Physical Exercise: The Balance between Oxidative Stress and ROS-Dependent Adaptive Signaling. Antioxid. Redox Signal. 2013, 18, 1208–1246. [Google Scholar] [CrossRef]
- Gostner, J.M.; Becker, K.; Ueberall, F.; Fuchs, D. The Good and Bad of Antioxidant Foods: An Immunological Perspective. Food Chem. Toxicol. Int. J. Publ. Br. Ind. Biol. Res. Assoc. 2015, 80, 72–79. [Google Scholar] [CrossRef]
- Karimaian, A.; Majidinia, M.; Bannazadeh Baghi, H.; Yousefi, B. The Crosstalk between Wnt/β-Catenin Signaling Pathway with DNA Damage Response and Oxidative Stress: Implications in Cancer Therapy. DNA Repair 2017, 51, 14–19. [Google Scholar] [CrossRef]
- Caliceti, C.; Nigro, P.; Rizzo, P.; Ferrari, R. ROS, Notch, and Wnt Signaling Pathways: Crosstalk between Three Major Regulators of Cardiovascular Biology. BioMed Res. Int. 2014, 2014, 318714. [Google Scholar] [CrossRef]
- Chatterjee, S.; Sil, P.C. ROS-Influenced Regulatory Cross-Talk With Wnt Signaling Pathway During Perinatal Development. Front. Mol. Biosci. 2022, 9, 889719. [Google Scholar] [CrossRef]
- Shen, Y.; Wang, Q.; Tian, Y. Reversal Effect of Ouabain on Multidrug Resistance in Esophageal Carcinoma EC109/CDDP Cells by Inhibiting the Translocation of Wnt/β-Catenin into the Nucleus. Tumour Biol. J. Int. Soc. Oncodevelopmental Biol. Med. 2016, 37, 15937–15947. [Google Scholar] [CrossRef]
- Contreras, R.G.; Flores-Maldonado, C.; Lázaro, A.; Shoshani, L.; Flores-Benitez, D.; Larré, I.; Cereijido, M. Ouabain Binding to Na+,K+-ATPase Relaxes Cell Attachment and Sends a Specific Signal (NACos) to the Nucleus. J. Membr. Biol. 2004, 198, 147–158. [Google Scholar] [CrossRef]
- Orellana, A.M.; Leite, J.A.; Kinoshita, P.F.; Vasconcelos, A.R.; Andreotti, D.Z.; de Sá Lima, L.; Xavier, G.F.; Kawamoto, E.M.; Scavone, C. Ouabain Increases Neuronal Branching in Hippocampus and Improves Spatial Memory. Neuropharmacology 2018, 140, 260–274. [Google Scholar] [CrossRef]
- Ma, Y.; Zhu, B.; Liu, X.; Yu, H.; Yong, L.; Liu, X.; Shao, J.; Liu, Z. Inhibition of Oleandrin on the Proliferation Show and Invasion of Osteosarcoma Cells in Vitro by Suppressing Wnt/β-Catenin Signaling Pathway. J. Exp. Clin. Cancer Res. 2015, 34, 115. [Google Scholar] [CrossRef]
- Ko, Y.S.; Rugira, T.; Jin, H.; Park, S.W.; Kim, H.J. Oleandrin and Its Derivative Odoroside A, Both Cardiac Glycosides, Exhibit Anticancer Effects by Inhibiting Invasion via Suppressing the STAT-3 Signaling Pathway. Int. J. Mol. Sci. 2018, 19, 3350. [Google Scholar] [CrossRef]
- Reddy, D.; Kumavath, R.; Tan, T.Z.; Ampasala, D.R.; Kumar, A.P. Peruvoside Targets Apoptosis and Autophagy through MAPK Wnt/β-Catenin and PI3K/AKT/mTOR Signaling Pathways in Human Cancers. Life Sci. 2020, 241, 117147. [Google Scholar] [CrossRef]
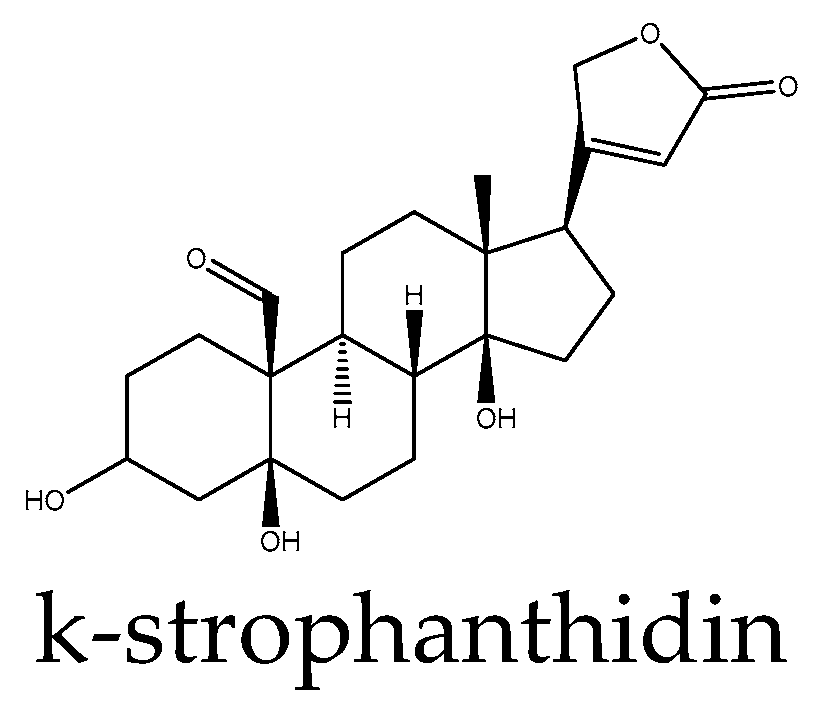
- Reddy, D.; Ghosh, P.; Kumavath, R. Strophanthidin Attenuates MAPK, PI3K/AKT/mTOR, and Wnt/β-Catenin Signaling Pathways in Human Cancers. Front. Oncol. 2019, 9, 1469. [Google Scholar] [CrossRef]
- Wen, S.-Y.; Chen, Y.-Y.; Deng, C.-M.; Zhang, C.-Q.; Jiang, M.-M. Nerigoside Suppresses Colorectal Cancer Cell Growth and Metastatic Potential through Inhibition of ERK/GSK3β/β-Catenin Signaling Pathway. Phytomedicine 2019, 57, 352–363. [Google Scholar] [CrossRef]
- Sun, Z.; Cao, X.; Jiang, M.-M.; Qiu, Y.; Zhou, H.; Chen, L.; Qin, B.; Wu, H.; Jiang, F.; Chen, J.; et al. Inhibition of β-Catenin Signaling by Nongenomic Action of Orphan Nuclear Receptor Nur77. Oncogene 2012, 31, 2653–2667. [Google Scholar] [CrossRef] [PubMed]
- Liu, X.; Geng, Z.; Ding, X.; Lou, Y.; Zhang, X. Convallatoxin Suppresses Osteosarcoma Cell Proliferation, Migration, Invasion, and Enhances Osteogenic Differentiation by Downregulating Parathyroid Hormone Receptor 1 (PTHR1) Expression and Inactivating Wnt/β-Catenin Pathway. Bioengineered 2022, 13, 13280–13292. [Google Scholar] [CrossRef] [PubMed]
- Park, H.Y.; Toume, K.; Arai, M.A.; Sadhu, S.K.; Ahmed, F.; Ishibashi, M. Calotropin: A Cardenolide from Calotropis Gigantea That Inhibits Wnt Signaling by Increasing Casein Kinase 1α in Colon Cancer Cells. Chembiochem 2014, 15, 872–878. [Google Scholar] [CrossRef] [PubMed]
- Qiu, D.-Z.; Zhang, Z.-J.; Wu, W.-Z.; Yang, Y.-K. Bufalin, a Component in Chansu, Inhibits Proliferation and Invasion of Hepatocellular Carcinoma Cells. BMC Complement. Altern. Med. 2013, 13, 185. [Google Scholar] [CrossRef]
- Yu, Z.; Feng, H.; Sun, X.; Zhuo, Y.; Li, M.; Zhou, Z.; Huang, L.; Jiang, Y.; Zhu, X.; Zhang, X.; et al. Bufalin Suppresses Hepatocarcinogenesis by Targeting β-Catenin/TCF Signaling via Cell Cycle-Related Kinase. Sci. Rep. 2018, 8, 3891. [Google Scholar] [CrossRef]
- Zhang, S.; Li, H.; Li, L.; Gao, Q.; Gu, L.; Hu, C.; Chen, M.; Zhang, X. Ophiopogonin B Inhibits Migration and Invasion in Non-Small Cell Lung Cancer Cells through Enhancing the Interaction between Axin and β-Catenin. J. Cancer 2021, 12, 6274–6284. [Google Scholar] [CrossRef]
- Sadava, D.; Kane, S.E. The Effect of Brassinolide, a Plant Steroid Hormone, on Drug Resistant Small-Cell Lung Carcinoma Cells. Biochem. Biophys. Res. Commun. 2017, 493, 783–787. [Google Scholar] [CrossRef]
- Ji, X.; Yang, C.; Xie, J.; Yin, X.; Hu, Q. Effect of Saponin from Tupistra Chinensis Baker on Proliferation and Apoptosis of Ovarian Cancer Cells by Wnt/β-Catenin Pathway. IUBMB Life 2020, 72, 1780–1786. [Google Scholar] [CrossRef]
- Zeng, J.; Liu, X.; Li, X.; Zheng, Y.; Liu, B.; Xiao, Y. Daucosterol Inhibits the Proliferation, Migration, and Invasion of Hepatocellular Carcinoma Cells via Wnt/β-Catenin Signaling. Molecules 2017, 22, 862. [Google Scholar] [CrossRef]
- Chen, T.; Lin, J.; Tang, D.; Zhang, M.; Wen, F.; Xue, D.; Zhang, H. Paris Saponin H Suppresses Human Hepatocellular Carcinoma (HCC) by Inactivation of Wnt/β-Catenin Pathway in Vitro and in Vivo. Int. J. Clin. Exp. Pathol. 2019, 12, 2875–2886. [Google Scholar]
- Ni, B.; Wang, W.; Liu, M.; Xu, Y.; Zhao, J. Paris Saponin Ⅰ Induce Toxicity in Zebrafish by Up-Regulation of P53 Pathway and down-Regulation of Wnt Pathway. Toxicon 2023, 228, 107094. [Google Scholar] [CrossRef] [PubMed]
- Chang, J.; Li, Y.; Wang, X.; Hu, S.; Wang, H.; Shi, Q.; Wang, Y.; Yang, Y. Polyphyllin I Suppresses Human Osteosarcoma Growth by Inactivation of Wnt/β-Catenin Pathway in Vitro and in Vivo. Sci. Rep. 2017, 7, 7605. [Google Scholar] [CrossRef] [PubMed]
- Liao, M.; Du, H.; Wang, B.; Huang, J.; Huang, D.; Tong, G. Anticancer Effect of Polyphyllin I in Suppressing Stem Cell-Like Properties of Hepatocellular Carcinoma via the AKT/GSK-3β/β-Catenin Signaling Pathway. Oxid. Med. Cell. Longev. 2022, 2022, 4031008. [Google Scholar] [CrossRef] [PubMed]
- Ge, Y.; Ding, S.; Feng, J.; Du, J.; Gu, Z. Diosgenin Inhibits Wnt/β-Catenin Pathway to Regulate the Proliferation and Differentiation of MG-63 Cells. Cytotechnology 2021, 73, 169–178. [Google Scholar] [CrossRef]
- Raju, J.; Bird, R.P. Diosgenin, a Naturally Occurring Furostanol Saponin Suppresses 3-Hydroxy-3-Methylglutaryl CoA Reductase Expression and Induces Apoptosis in HCT-116 Human Colon Carcinoma Cells. Cancer Lett. 2007, 255, 194–204. [Google Scholar] [CrossRef]
- Bhuvanalakshmi, G.; Basappa; Rangappa, K.S.; Dharmarajan, A.; Sethi, G.; Kumar, A.P.; Warrier, S. Breast Cancer Stem-Like Cells Are Inhibited by Diosgenin, a Steroidal Saponin, by the Attenuation of the Wnt β-Catenin Signaling via the Wnt Antagonist Secreted Frizzled Related Protein-4. Front. Pharmacol. 2017, 8, 124. [Google Scholar] [CrossRef]
- Zhang, S.; Pang, H.; Sun, M.; Li, H. Timosaponin AIII Inhibits the Growth of Human Leukaemia Cells HL-60 by down-Regulation of PI3K/AKT and Wnt/β-Catenin Pathways. Biotechnol. Biotechnol. Equip. 2018, 32, 150–155. [Google Scholar] [CrossRef]
- Zhang, X.; Han, X.; Yin, L.; Xu, L.; Qi, Y.; Xu, Y.; Sun, H.; Lin, Y.; Liu, K.; Peng, J. Potent Effects of Dioscin against Liver Fibrosis. Sci. Rep. 2015, 5, 9713. [Google Scholar] [CrossRef]
- Wang, X.D.; Su, G.Y.; Zhao, C.; Qu, F.Z.; Wang, P.; Zhao, Y.Q. Anticancer Activity and Potential Mechanisms of 1C, a Ginseng Saponin Derivative, on Prostate Cancer Cells. J. Ginseng Res. 2018, 42, 133–143. [Google Scholar] [CrossRef]
- He, B.-C.; Gao, J.-L.; Luo, X.; Luo, J.; Shen, J.; Wang, L.; Zhou, Q.; Wang, Y.-T.; Luu, H.H.; Haydon, R.C.; et al. Ginsenoside Rg3 Inhibits Colorectal Tumor Growth through the Down-Regulation of Wnt/ß-Catenin Signaling. Int. J. Oncol. 2011, 38, 437–445. [Google Scholar] [CrossRef]
- Liu, J.J.; Zhang, H.Y.; Chen, X.; Zhang, G.B.; Lin, J.K.; Feng, H.; Chu, W.H. 20-Hydroxyecdysone Improves Neuronal Differentiation of Adult Hippocampal Neural Stem Cells in High Power Microwave Radiation-Exposed Rats. Biomed. Environ. Sci. BES 2022, 35, 504–517. [Google Scholar] [CrossRef] [PubMed]
- Ye, Z.-N.; Yuan, F.; Liu, J.-Q.; Peng, X.-R.; An, T.; Li, X.; Kong, L.-M.; Qiu, M.-H.; Li, Y. Physalis Peruviana-Derived 4β-Hydroxywithanolide E, a Novel Antagonist of Wnt Signaling, Inhibits Colorectal Cancer In Vitro and In Vivo. Molecules 2019, 24, 1146. [Google Scholar] [CrossRef] [PubMed]
- Kuang, Z.; Bai, J.; Ni, L.; Hang, K.; Xu, J.; Ying, L.; Xue, D.; Pan, Z. Withanolide B Promotes Osteogenic Differentiation of Human Bone Marrow Mesenchymal Stem Cells via ERK1/2 and Wnt/β-Catenin Signaling Pathways. Int. Immunopharmacol. 2020, 88, 106960. [Google Scholar] [CrossRef]
- Chen, C.; Zhu, D.; Zhang, H.; Han, C.; Xue, G.; Zhu, T.; Luo, J.; Kong, L. YAP-Dependent Ubiquitination and Degradation of β-Catenin Mediates Inhibition of Wnt Signalling Induced by Physalin F in Colorectal Cancer. Cell Death Dis. 2018, 9, 591. [Google Scholar] [CrossRef]
- Shukla, S.; Sinha, S.; Khan, S.; Kumar, S.; Singh, K.; Mitra, K.; Maurya, R.; Meeran, S.M. Cucurbitacin B Inhibits the Stemness and Metastatic Abilities of NSCLC via Downregulation of Canonical Wnt/β-Catenin Signaling Axis. Sci. Rep. 2016, 6, 21860. [Google Scholar] [CrossRef]
- Dakeng, S.; Duangmano, S.; Jiratchariyakul, W.; U-Pratya, Y.; Bögler, O.; Patmasiriwat, P. Inhibition of Wnt Signaling by Cucurbitacin B in Breast Cancer Cells: Reduction of Wnt-Associated Proteins and Reduced Translocation of Galectin-3-Mediated β-Catenin to the Nucleus. J. Cell. Biochem. 2012, 113, 49–60. [Google Scholar] [CrossRef]
- Feng, H.; Zang, L.; Zhao, Z.-X.; Kan, Q.-C. Cucurbitacin-E Inhibits Multiple Cancer Cells Proliferation through Attenuation of Wnt/β-Catenin Signaling. Cancer Biother. Radiopharm. 2014, 29, 210–214. [Google Scholar] [CrossRef]
- Yang, P.; Liu, W.; Fu, R.; Ding, G.-B.; Amin, S.; Li, Z. Cucurbitacin E Chemosensitizes Colorectal Cancer Cells via Mitigating TFAP4/Wnt/β-Catenin Signaling. J. Agric. Food Chem. 2020, 68, 14148–14160. [Google Scholar] [CrossRef]
- Zhao, H.; Tang, S.; Tao, Q.; Ming, T.; Lei, J.; Liang, Y.; Peng, Y.; Wang, M.; Liu, M.; Yang, H.; et al. Ursolic Acid Suppresses Colorectal Cancer by Down-Regulation of Wnt/β-Catenin Signaling Pathway Activity. J. Agric. Food Chem. 2023, 71, 3981–3993. [Google Scholar] [CrossRef]
- Park, J.-H.; Kwon, H.-Y.; Sohn, E.J.; Kim, K.A.; Kim, B.; Jeong, S.-J.; Song, J.H.; Koo, J.S.; Kim, S.-H. Inhibition of Wnt/β-Catenin Signaling Mediates Ursolic Acid-Induced Apoptosis in PC-3 Prostate Cancer Cells. Pharmacol. Rep. 2013, 65, 1366–1374. [Google Scholar] [CrossRef]
- Fan, X.; Wang, P.; Sun, Y.; Jiang, J.; Du, H.; Wang, Z.; Duan, Z.; Lei, H.; Li, H. Oleanolic Acid Derivatives Inhibit the Wnt/β-Catenin Signaling Pathway by Promoting the Phosphorylation of β-Catenin in Human SMMC-7721 Cells. Die Pharm. 2016, 71, 398–401. [Google Scholar] [CrossRef]
- Zhang, L.; Tu, Y.; He, W.; Peng, Y.; Qiu, Z. A Novel Mechanism of Hepatocellular Carcinoma Cell Apoptosis Induced by Lupeol via Brain-Derived Neurotrophic Factor Inhibition and Glycogen Synthase Kinase 3 Beta Reactivation. Eur. J. Pharmacol. 2015, 762, 55–62. [Google Scholar] [CrossRef]
- Saleem, M.; Murtaza, I.; Tarapore, R.S.; Suh, Y.; Adhami, V.M.; Johnson, J.J.; Siddiqui, I.A.; Khan, N.; Asim, M.; Hafeez, B.B.; et al. Lupeol Inhibits Proliferation of Human Prostate Cancer Cells by Targeting Beta-Catenin Signaling. Carcinogenesis 2009, 30, 808–817. [Google Scholar] [CrossRef] [PubMed]
- Tarapore, R.S.; Siddiqui, I.A.; Adhami, V.M.; Spiegelman, V.S.; Mukhtar, H. The Dietary Terpene Lupeol Targets Colorectal Cancer Cells with Constitutively Active Wnt/β-Catenin Signaling. Mol. Nutr. Food Res. 2013, 57, 1950–1958. [Google Scholar] [CrossRef]
- Tarapore, R.S.; Siddiqui, I.A.; Saleem, M.; Adhami, V.M.; Spiegelman, V.S.; Mukhtar, H. Specific Targeting of Wnt/β-Catenin Signaling in Human Melanoma Cells by a Dietary Triterpene Lupeol. Carcinogenesis 2010, 31, 1844–1853. [Google Scholar] [CrossRef]
- Cheng, X.; Wei, B.; Sun, L.; Hu, X.; Liang, J.; Chen, Y. Astragaloside I Stimulates Osteoblast Differentiation Through the Wnt/β-Catenin Signaling Pathway. Phytother. Res. 2016, 30, 1680–1688. [Google Scholar] [CrossRef]
- Fan, J.; Jia, F.; Liu, Y.; Zhou, X. Astragalus Polysaccharides and Astragaloside IV Alleviate Inflammation in Bovine Mammary Epithelial Cells by Regulating Wnt/β-Catenin Signaling Pathway. PLoS ONE 2022, 17, e0271598. [Google Scholar] [CrossRef]
- Sun, N.-Y.; Liu, X.-L.; Gao, J.; Wu, X.-H.; Dou, B. Astragaloside-IV Modulates NGF-induced Osteoblast Differentiation via the GSK3β/β-catenin Signalling Pathway. Mol. Med. Rep. 2021, 23, 19. [Google Scholar] [CrossRef]
- Cao, Y.; Lv, Q.; Huang, Z.; Li, Y. Astragaloside-IV Induces the Differentiation of Bone Marrow Mesenchymal Stem Cells into Osteoblasts through NMUR2-Mediated Wnt/β-Catenin Pathway. Regen. Med. 2023, 18, 471–485. [Google Scholar] [CrossRef]
- Qin, C.-D.; Ma, D.-N.; Ren, Z.-G.; Zhu, X.-D.; Wang, C.-H.; Wang, Y.-C.; Ye, B.-G.; Cao, M.-Q.; Gao, D.-M.; Tang, Z.-Y. Astragaloside IV Inhibits Metastasis in Hepatoma Cells through the Suppression of Epithelial-Mesenchymal Transition via the Akt/GSK-3β/β-Catenin Pathway. Oncol. Rep. 2017, 37, 1725–1735. [Google Scholar] [CrossRef]
- Cui, X.; Jiang, X.; Wei, C.; Xing, Y.; Tong, G. Astragaloside IV Suppresses Development of Hepatocellular Carcinoma by Regulating miR-150-5p/β-Catenin Axis. Environ. Toxicol. Pharmacol. 2020, 78, 103397. [Google Scholar] [CrossRef] [PubMed]
- Han, J.; Shen, X.; Zhang, Y.; Wang, S.; Zhou, L. Astragaloside IV Suppresses Transforming Growth Factor-Β1-Induced Epithelial-Mesenchymal Transition through Inhibition of Wnt/β-Catenin Pathway in Glioma U251 Cells. Biosci. Biotechnol. Biochem. 2020, 84, 1345–1352. [Google Scholar] [CrossRef] [PubMed]
- An, W.F.; Germain, A.R.; Bishop, J.A.; Nag, P.P.; Metkar, S.; Ketterman, J.; Walk, M.; Weiwer, M.; Liu, X.; Patnaik, D.; et al. Discovery of Potent and Highly Selective Inhibitors of GSK3b. In Probe Reports from the NIH Molecular Libraries Program; National Center for Biotechnology Information (US): Bethesda, MD, USA, 2010. [Google Scholar]
- Zhu, Y.; Wang, Y.; Jia, Y.; Xu, J.; Chai, Y. Catalpol Promotes the Osteogenic Differentiation of Bone Marrow Mesenchymal Stem Cells via the Wnt/β-Catenin Pathway. Stem Cell Res. Ther. 2019, 10, 37. [Google Scholar] [CrossRef] [PubMed]
- Chung, H.-J.; Kim, W.K.; Oh, J.; Kim, M.-R.; Shin, J.-S.; Lee, J.; Ha, I.-H.; Lee, S.K. Anti-Osteoporotic Activity of Harpagoside by Upregulation of the BMP2 and Wnt Signaling Pathways in Osteoblasts and Suppression of Differentiation in Osteoclasts. J. Nat. Prod. 2017, 80, 434–442. [Google Scholar] [CrossRef] [PubMed]
- Zhou, L.; Wang, H.; Jing, J.; Yu, L.; Wu, X.; Lu, Z. Morroniside Regulates Hair Growth and Cycle Transition via Activation of the Wnt/β-Catenin Signaling Pathway. Sci. Rep. 2018, 8, 13785. [Google Scholar] [CrossRef]
- Takayama, H.; Jia, Z.-J.; Kremer, L.; Bauer, J.O.; Strohmann, C.; Ziegler, S.; Antonchick, A.P.; Waldmann, H. Discovery of Inhibitors of the Wnt and Hedgehog Signaling Pathways through the Catalytic Enantioselective Synthesis of an Iridoid-Inspired Compound Collection. Angew. Chem. Int. Ed. 2013, 52, 12404–12408. [Google Scholar] [CrossRef]
- Tapia-Rojas, C.; Schüller, A.; Lindsay, C.B.; Ureta, R.C.; Mejías-Reyes, C.; Hancke, J.; Melo, F.; Inestrosa, N.C. Andrographolide Activates the Canonical Wnt Signalling Pathway by a Mechanism That Implicates the Non-ATP Competitive Inhibition of GSK-3β: Autoregulation of GSK-3β in Vivo. Biochem. J. 2015, 466, 415–430. [Google Scholar] [CrossRef]
- Tang, L.; Zhu, H.; Yang, X.; Xie, F.; Peng, J.; Jiang, D.; Xie, J.; Qi, M.; Yu, L. Shizukaol D, a Dimeric Sesquiterpene Isolated from Chloranthus Serratus, Represses the Growth of Human Liver Cancer Cells by Modulating Wnt Signalling Pathway. PLoS ONE 2016, 11, e0152012. [Google Scholar] [CrossRef]
- Pelusi, L.; Mandatori, D.; Di Pietrantonio, N.; Del Pizzo, F.; Di Tomo, P.; Di Pietro, N.; Buda, R.; Genovese, S.; Epifano, F.; Pandolfi, A.; et al. Estrogen Receptor 1 (ESR1) and the Wnt/β-Catenin Pathway Mediate the Effect of the Coumarin Derivative Umbelliferon on Bone Mineralization. Nutrients 2022, 14, 3209. [Google Scholar] [CrossRef] [PubMed]
- Ge, L.; Cui, Y.; Liu, B.; Yin, X.; Pang, J.; Han, J. ERα and Wnt/Β-catenin Signaling Pathways Are Involved in Angelicin-dependent Promotion of Osteogenesis. Mol. Med. Rep. 2019, 19, 3469–3476. [Google Scholar] [CrossRef]
- Hong, W.; Zhang, W. Hesperidin Promotes Differentiation of Alveolar Osteoblasts via Wnt/β-Catenin Signaling Pathway. J. Recept. Signal Transduct. 2020, 40, 442–448. [Google Scholar] [CrossRef] [PubMed]
- Lan, X.; Gui, Z.; Chen, T.; Tang, M.; Wang, H. Genistin Represses the Proliferation and Angiogenesis While Accelerating the Apoptosis of Glioma Cells by Modulating the FOXC1-Mediated Wnt Signaling Pathway. Discov. Med. 2024, 36, 332–342. [Google Scholar] [CrossRef] [PubMed]
- Dong, Z.; Shi, Y.; Zhao, H.; Li, N.; Ye, L.; Zhang, S.; Zhu, H. Sulphonated Formononetin Induces Angiogenesis through Vascular Endothelial Growth Factor/cAMP Response Element-Binding Protein/Early Growth Response 3/Vascular Cell Adhesion Molecule 1 and Wnt/β-Catenin Signaling Pathway. Pharmacology 2017, 101, 76–85. [Google Scholar] [CrossRef]
- Fu, D.-J.; Zhang, L.; Song, J.; Mao, R.-W.; Zhao, R.-H.; Liu, Y.-C.; Hou, Y.-H.; Li, J.-H.; Yang, J.-J.; Jin, C.-Y.; et al. Design and Synthesis of Formononetin-Dithiocarbamate Hybrids That Inhibit Growth and Migration of PC-3 Cells via MAPK/Wnt Signaling Pathways. Eur. J. Med. Chem. 2017, 127, 87–99. [Google Scholar] [CrossRef]
- Hu, C.; Dong, T.; Li, R.; Lu, J.; Wei, X.; Liu, P. Emodin Inhibits Epithelial to Mesenchymal Transition in Epithelial Ovarian Cancer Cells by Regulation of GSK-3β/β-Catenin/ZEB1 Signaling Pathway. Oncol. Rep. 2016, 35, 2027–2034. [Google Scholar] [CrossRef]
- Du, M.; Shen, P.; Tan, R.; Wu, D.; Tu, S. Aloe-Emodin Inhibits the Proliferation, Migration, and Invasion of Melanoma Cells via Inactivation of the Wnt/Beta-Catenin Signaling Pathway. Ann. Transl. Med. 2021, 9, 1722. [Google Scholar] [CrossRef]
- Hussain, T.; Alafnan, A.; Almazni, I.A.; Helmi, N.; Moin, A.; Baeissa, H.M.; Awadelkareem, A.M.; Elkhalifa, A.O.; Bakhsh, T.; Alzahrani, A.; et al. Aloe-Emodin Exhibits Growth-Suppressive Effects on Androgen-Independent Human Prostate Cancer DU145 Cells via Inhibiting the Wnt/β-Catenin Signaling Pathway: An in Vitro and in Silico Study. Front. Pharmacol. 2024, 14, 1325184. [Google Scholar] [CrossRef]
- Gebhardt, R.; Lerche, K.S.; Götschel, F.; Günther, R.; Kolander, J.; Teich, L.; Zellmer, S.; Hofmann, H.-J.; Eger, K.; Hecht, A.; et al. 4-Aminoethylamino-Emodin—A Novel Potent Inhibitor of GSK-3β– Acts as an Insulin-Sensitizer Avoiding Downstream Effects of Activated β-Catenin. J. Cell. Mol. Med. 2010, 14, 1276–1293. [Google Scholar] [CrossRef]
- Albring, K.F.; Weidemüller, J.; Mittag, S.; Weiske, J.; Friedrich, K.; Geroni, M.C.; Lombardi, P.; Huber, O. Berberine Acts as a Natural Inhibitor of Wnt/β-Catenin Signaling--Identification of More Active 13-Arylalkyl Derivatives. BioFactors Oxf. Engl. 2013, 39, 652–662. [Google Scholar] [CrossRef]
- Liu, Y.; Qi, Y.; Bai, Z.-H.; Ni, C.-X.; Ren, Q.-H.; Xu, W.-H.; Xu, J.; Hu, H.-G.; Qiu, L.; Li, J.-Z.; et al. A Novel Matrine Derivate Inhibits Differentiated Human Hepatoma Cells and Hepatic Cancer Stem-like Cells by Suppressing PI3K/AKT Signaling Pathways. Acta Pharmacol. Sin. 2017, 38, 120–132. [Google Scholar] [CrossRef]
- Liu, W.; Zhao, Z.; Wang, Y.; Li, W.; Su, Q.; Jia, Q.; Zhang, J.; Zhang, X.; Shen, J.; Yin, J. Dioscin Inhibits Stem-Cell-like Properties and Tumor Growth of Osteosarcoma through Akt/GSK3/β-Catenin Signaling Pathway. Cell Death Dis. 2018, 9, 343. [Google Scholar] [CrossRef] [PubMed]
- Amado, N.G.; Fonseca, B.F.; Cerqueira, D.M.; Neto, V.M.; Abreu, J.G. Flavonoids: Potential Wnt/Beta-Catenin Signaling Modulators in Cancer. Life Sci. 2011, 89, 545–554. [Google Scholar] [CrossRef] [PubMed]
- Qin, B.; Zeng, Z.; Xu, J.; Shangwen, J.; Ye, Z.J.; Wang, S.; Wu, Y.; Peng, G.; Wang, Q.; Gu, W.; et al. Emodin Inhibits Invasion and Migration of Hepatocellular Carcinoma Cells via Regulating Autophagy-Mediated Degradation of Snail and β-Catenin. BMC Cancer 2022, 22, 671. [Google Scholar] [CrossRef] [PubMed]
- Kim, S.; Lim, S.-W.; Choi, J. Drug Discovery Inspired by Bioactive Small Molecules from Nature. Anim. Cells Syst. 2022, 26, 254–265. [Google Scholar] [CrossRef]
- Katanaev, V.L.; Blagodatski, A.; Xu, J.; Khotimchenko, Y.; Koval, A. Mining Natural Compounds to Target WNT Signaling: Land and Sea Tales. Handb. Exp. Pharmacol. 2021, 269, 215–248. [Google Scholar] [CrossRef]
- Quiros-Guerrero, L.-M.; Marcourt, L.; Chaiwangrach, N.; Koval, A.; Ferreira Queiroz, E.; David, B.; Grondin, A.; Katanaev, V.L.; Wolfender, J.-L. Integration of Wnt-Inhibitory Activity and Structural Novelty Scoring Results to Uncover Novel Bioactive Natural Products: New Bicyclo[3.3.1]Non-3-Ene-2,9-Diones from the Leaves of Hymenocardia Punctata. Front. Chem. 2024, 12, 1371982. [Google Scholar] [CrossRef]
- Olivon, F.; Allard, P.-M.; Koval, A.; Righi, D.; Genta-Jouve, G.; Neyts, J.; Apel, C.; Pannecouque, C.; Nothias, L.-F.; Cachet, X.; et al. Bioactive Natural Products Prioritization Using Massive Multi-Informational Molecular Networks. ACS Chem. Biol. 2017, 12, 2644–2651. [Google Scholar] [CrossRef]
- Mamadalieva, N.Z.; Koval, A.; Dusmuratov, M.M.; Hussain, H.; Katanaev, V.L. Chemical and Biological Characterization of Metabolites from Silene Viridiflora Using Mass Spectrometric and Cell-Based Assays. Biomolecules 2024, 14, 1285. [Google Scholar] [CrossRef]
- Eshboev, F.; Mamadalieva, N.; Nazarov, P.A.; Hussain, H.; Katanaev, V.; Egamberdieva, D.; Azimova, S. Antimicrobial Action Mechanisms of Natural Compounds Isolated from Endophytic Microorganisms. Antibiot. Basel Switz. 2024, 13, 271. [Google Scholar] [CrossRef]
- Ma, Y.; Zhao, Y.; Luo, M.; Jiang, Q.; Liu, S.; Jia, Q.; Bai, Z.; Wu, F.; Xie, J. Advancements and Challenges in Pharmacokinetic and Pharmacodynamic Research on the Traditional Chinese Medicine Saponins: A Comprehensive Review. Front. Pharmacol. 2024, 15, 1393409. [Google Scholar] [CrossRef]
- Khotimchenko, Y.S.; Silachev, D.N.; Katanaev, V.L. Marine Natural Products from the Russian Pacific as Sources of Drugs for Neurodegenerative Diseases. Mar. Drugs 2022, 20, 708. [Google Scholar] [CrossRef] [PubMed]
- Klimenko, A.; Huber, R.; Marcourt, L.; Tabakaev, D.; Koval, A.; Dautov, S.S.; Dautova, T.N.; Wolfender, J.-L.; Thew, R.; Khotimchenko, Y.; et al. Shallow- and Deep-Water Ophiura Species Produce a Panel of Chlorin Compounds with Potent Photodynamic Anticancer Activities. Antioxid. Basel Switz. 2023, 12, 386. [Google Scholar] [CrossRef] [PubMed]
- Stan, D.; Enciu, A.-M.; Mateescu, A.L.; Ion, A.C.; Brezeanu, A.C.; Stan, D.; Tanase, C. Natural Compounds With Antimicrobial and Antiviral Effect and Nanocarriers Used for Their Transportation. Front. Pharmacol. 2021, 12, 723233. [Google Scholar] [CrossRef] [PubMed]
- Huber, R.; Marcourt, L.; Koval, A.; Schnee, S.; Righi, D.; Michellod, E.; Katanaev, V.L.; Wolfender, J.-L.; Gindro, K.; Queiroz, E.F. Chemoenzymatic Synthesis of Complex Phenylpropanoid Derivatives by the Botrytis Cinerea Secretome and Evaluation of Their Wnt Inhibition Activity. Front. Plant Sci. 2021, 12, 805610. [Google Scholar] [CrossRef]




{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Compound | Wnt Signaling Influence | Experimental System | Ref. | |
|---|---|---|---|---|
| 2.2. Cardiac glycosides | Ouabain | Inhibition (reduces β-catenin nuclear translocation) | Esophageal cancer cell line EC109 | [53] |
| Activation (promotes β-catenin nuclear shuttle via tyrosine phosphorylation) | MDCK cells | [54] | ||
| Activation (inactivates GSK3β, leading to β-catenin stabilization and nuclear translocation) | Hippocampus of adult rats | [55] | ||
| Oleandrin | Inhibition (suppresses TopFlash reporter and target genes, inhibits β-catenin nuclear translocation) | Osteosarcoma cell lines U2OS and SaOS-2 | [56] | |
| Inhibition (suppresses TopFlash reporter and target genes, inhibits β-catenin nuclear translocation) | Breast cancer cell line MDA-MB-231 and its radioresistant subclone | [57] | ||
| Odoroside A | Inhibition (contributes to reduced invasiveness and radioresistance, partly through decreased β-catenin levels) | Breast cancer cell line MDA-MB-231 and its radioresistant subclone | [57] | |
| Peruvoside | Inhibition (decreases β-catenin levels in nucleus and cytoplasm, reduces expression of Wnt target genes) | MCF-7 breast cancer, A549 lung cancer, and HepG2 liver cancer cell lines | [58] | |
| Strophant-hidine | Inhibition (decreases β-catenin levels in nucleus and cytoplasm, reduces expression of Wnt target genes) | MCF-7 breast cancer, A549 lung cancer, and HepG2 liver cancer cell lines | [59] | |
| Nerigoside | Inhibition (suppresses β-catenin degradation via ERK crosstalk, promoting GSK3β degradation) | Colorectal cancer cell lines HT29 and SW620 | [60] | |
| H-9 | Inhibition (downregulates Wnt signaling through direct interaction and induction of β-catenin degradation) | Colorectal cancer cells | [61] | |
| ATE-i2-b4 | [61] | |||
| Convalla-toxin | Inhibition (through crosstalk with parathyroid hormone receptor 1 (PTHR1)) | Osteosarcoma cells | [62] | |
| Calotropin | Inhibition (increases CK1a levels leading to increased β-catenin degradation) | Colon cancer cells | [63] | |
| Bufalin | Activation (inhibits AKT phosphorylation, potentially leading to decreased GSK3β activity and increased β-catenin stability) | Hepatoma cell lines | [64] | |
| Inhibition (suppresses CCRK-driven transcription, leading to loss of active β-catenin) | Liver cancer | [65] | ||
| Telocino- bufagin | Inhibition (inhibits Wnt/β-catenin signaling via modulation of GSK3β activity and by negatively affecting β-catenin stability, with lower potency for marinobufagin) | LLC-PK1 porcine kidney epithelial cells | [2] | |
| Marino-bufagin | ||||
| 2.3. Steroid saponins | Ophiopo-gonin B | Inhibition (reduces β-catenin, cyclin D1, c-Myc, strengthens Axin/β-catenin interaction) | Non-small-cell lung carcinoma cell lines | [66] |
| Epi-brassinolide | Inhibition (reduces β-catenin, c-Myc, CCND1, Sox9, c-Jun, survivin, MMP1, MMP7, mPar) | NCI-H69 and VPA17 small-cell lung carcinoma line | [67] | |
| Saponin (Tupistra) | Inhibition (reduces β-catenin and c-Myc) | Ovarian cancer cells SKOV3 | [68] | |
| Daucosterol | Inhibition (reduces β-catenin and Wnt5a, increases GSK3β) | Hepatocellular carcinoma lines HepG2 and SMMC-7721 | [69] | |
| Paris saponin H | Inhibition (downregulates β-catenin and p-GSK3β) | Hepatocellular carcinoma PLC/PRF/5 and Huh7 cells, in vivo xenograft | [70] | |
| Paris saponin I | Inhibition (decreases β-catenin) | Zebrafish | [71] | |
| Polyphyllin I (Chong Lou) | Inhibition (suppresses p-GSK3β, decreases β-catenin) | Osteosarcoma cell lines (in vitro), xenograft orthotopic mouse model | [72] | |
| Inhibition (suppresses p-GSK3β, decreases β-catenin, requires AKT crosstalk) | Liver cancer stem cells | [73] | ||
| Inhibition (counteracts aberrant Wnt activation by AngII, decreases β-catenin, p-GSK3β, c-Myc, c-Jun, c-Fos, cyclin D1) | Mouse model of pressure overload-induced cardiac hypertrophy, cultured neonatal rat ventricular myocytes | [21] | ||
| Diosgenin | Inhibition (decreases β-catenin) | MG-63 osteoblast-like cells | [74] | |
| Inhibition (decreases β-catenin) | HCT-116 human colon carcinoma cells | [75] | ||
| Inhibition (upregulates sFRP4 and DKK1) | Breast cancer cell lines (mammosphere-enriched) | [76] | ||
| Timosapo-nin AIII | Inhibition (decreases β-catenin, cyclin D1, c-Myc, crosstalk with AKT) | Human leukemia HL-60 cells | [77] | |
| Timosaponin B II | Activation (increases Dishevelled, increases β-catenin) | UMR-106 osteoblastic cells and primary rat osteoblasts | [7] | |
| Dioscin | Inhibition (increases GSK3β activity by decreased phosphorylation, downregulates nuclear β-catenin, blocks AKT) | Osteosarcoma model, xenograft tumor | [69] | |
| Inhibition (increases GSK3β activity by decreased phosphorylation, downregulates nuclear β-catenin) | Rat model of CCl4-induced liver fibrosis, primary rat hepatic stellate cells (in vitro and in vivo) | [78] | ||
| Inhibition (counteracts increased Wnt3a and β-catenin) | Osteoarthritis mouse model | [11] | ||
| Activation (increases LRP5 mRNA, β-catenin mRNA and protein) | Mouse osteoblast-like MC3T3-E1 and MG-63 cells | [6] | ||
| Ginsenoside 1C | Inhibition (decreases β-catenin, TCF4, CCND1, c-Myc) | LNCaP prostate cancer cell line | [79] | |
| Inhibition (blocks transcriptional activity, decreases nuclear β-catenin, c-Myc) | Colorectal cancer cells SW480 and HCT116 cell lines, xenograft model | [80] | ||
| Inhibition (inhibits p-β-catenin Ser675 and nuclear translocation) | Adenine-induced chronic kidney disease in rats, rat vascular smooth muscle cells | [3] | ||
| Activation (increases Wnt1 and β-catenin, reduces GSK3β and p-GSK3β) | Mouse model of intestinal ischemia–reperfusion injury (in vivo), DKK1 blocking experiments | [19] | ||
| Activation (increases β-catenin and LEF1, decreases DKK1) | Human hair dermal papilla cell line, C57BL/6 mice | [22] | ||
| 2.4. Ecdysteroids | β-Ecdysterone (20-hydroxyecdysone) | Activation (enriched gene signature of Wnt/β-catenin activation, synergy with BMP-2) | MC3T3-E1 preosteoblasts | [8] |
| Activation (upregulates Wnt3a followed by increasing β-catenin) | Adult rat hippocampal neural stem cells (in vitro), rat model of microwave-radiation-induced learning and memory deficits (in vivo) | [81] | ||
| 2.5. Withanolides | 4β-hydroxy-withanolide E | Inhibition (increases phosphorylated β-catenin, decreases active and total β-catenin) | Colon cancer HCT116 cells, HCT116 xenografts (in vivo) | [82] |
| Withanolide B | Potentiation/activation (enhances osteogenic differentiation through Wnt/β-catenin pathway) | Human bone marrow mesenchymal stem cells (in vitro), rat tibial osteotomy model (in vivo) | [83] | |
| Physalin F | Inhibition (antagonist of Wnt/β-catenin signaling, accelerates β-catenin ubiquitination and degradation in a YAP-dependent manner) | Colorectal cancer cells (in vitro and in vivo) | [84] | |
| 2.6. Cucurbitacins | Cucurbitacin B | Inhibition (directly affects β-catenin and TCF1 interaction) | Lung cancer H1299 and A549 cell lines, in vivo mouse model (NNK-induced) | [85] |
| Inhibition (prevents β-catenin nuclear translocation, loss of p-GSK3β, downregulates cyclin D1 and c-Myc) | Breast cancer T47D and SKBR-3 cell lines | [86] | ||
| Cucurbitacin E | Inhibition (prevents β-catenin nuclear accumulation by inducing menin, downregulates cyclin D1) | Lung (A549), hepatocellular (Hep3B), and colon (SW480) cancer cell lines | [87] | |
| Inhibition (downregulates ABCC1, MDR1, PKM2, cyclin D1, Axin2, c-Myc, and β-catenin combined with 5-FU) | Colorectal cancer cell lines DLD1 and HCT-116 | [88] | ||
| 2.7. Triterpenes | Betulinic acid | Inhibition (reduces Wnt3a-/LiCl-induced transcription, suppresses β-catenin nuclear accumulation, promotes β-catenin phosphorylation and degradation, loss of p-LRP6) | NIH-3T3 fibroblasts, primary and immortalized lung fibroblasts (Mlg line), bleomycin-induced mouse model of pulmonary fibrosis (in vivo) | [23] |
| Poricoic acid | Inhibition (suppresses Wnt1, β-catenin, active β-catenin, Snail1, Twist, MMP-7, PAI-1, Fsp-1) | Proximal kidney epithelial cells HK-2 and podocytes MPC5 (stimulated with TGF-β1 and Ang II) | [4] | |
| Ganoderal A | Activation (upregulates β-catenin, Wnt3, FZD receptors, osteogenic-specific proteins and genes) | Human amniotic mesenchymal stem cells | [9] | |
| Corosolic acid | Inhibition (antagonist of Wnt/β-catenin pathway) | Colon cancer cells | [28] | |
| Ursolic acid | Inhibition (reduces Wnt4, TCF4, LEF1, β-catenin, inhibits β-catenin nuclear translocation, increases GSK3β) | CRC cell lines | [89] | |
| Inhibition (suppresses Wnt5a/b and β-catenin, increases p-GSK3β Ser9) | PC-3 prostate cancer cells | [90] | ||
| Inhibition (promotes Wnt3a-induced β-catenin degradation via b-TrCP) | HEK293-FL reporter cells, HCT-15 colon cancer cells | [28] | ||
| Oleanolic acid | Inhibition (decreases β-catenin, promotes p-GSK3β, decreases Wnt3a, WISP1) | Primary rat chondrocytes (induced by IL-1β) | [10] | |
| Inhibition (low β-catenin in cytoplasm and nucleus by promoting phosphorylation, decreases c-Myc and Cyclin D1) | Human hepatocellular carcinoma SMMC-7721 cells | [91] | ||
| Lupeol | Inhibition (decreases p-GSK3β via PI3K/AKT, suppresses LiCl-induced activity) | Human hepatocellular carcinoma cell line Huh-7 | [92] | |
| Inhibition (decreases p-GSK3β, increases Axin, reduces cyclin D1 and MMP-2) | Prostate cancer LNCaP and DU145 cells | [93] | ||
| Inhibition (prevents β-catenin translocation by affecting Ser552 and Ser675 phosphorylation, reduces Axin2 and TCF1 expression) | Colorectal cancer cells HCT 116 and DLD 1 | [94] | ||
| Inhibition (decreases proliferation and viability, reduces MITF, CRD-BP, cyclin D1, loss of β-catenin translocation) | Melanoma cells with APC or β-catenin mutations, in vivo xenografts, HEK293T reporter cells | [95] | ||
| Astra-galoside I | Activation (upregulates β-catenin and Runx2, increases OPG/RANKL ratio) | MC3T3-E1 cells | [96] | |
| Total astragaloside fraction | Inhibition (increases p-GSK3β Tyr216, decreases p-GSK3β Ser9, decreases TCF4 and Id2) | MO3.13 oligodendroglial cell line, CPZ-induced mouse model of demyelination (corpus callosum) | [18] | |
| Astragaloside IV | Activation (increases β-catenin stability, antagonizes LPS-induced inhibition) | Bovine mammary epithelial cells | [97] | |
| Activation (promotes osteogenesis, reduces p-GSK3β Ser9, crosstalk with NGF) | Bone-marrow-derived mesenchymal stem cells | [98] | ||
| Activation (enhances osteogenic differentiation, NMUR2 crosstalk, reversed by DKK1) | Rat tibia defect models and rat bone marrow mesenchymal stem cells | [99] | ||
| Activation (increases p-GSK3β levels) | T84 cells (in vitro), peritoneal dialysis mouse models (in vivo) | [20] | ||
| Inhibition (suppresses EMT, controls AKT phosphorylation via p-GSK3β Ser9) | Hepatocellular carcinoma Huh7 and MHCC97-H cells | [100] | ||
| Inhibition (via miR-150-5p action, β-catenin overexpression counteracts) | Hepatocellular carcinoma SMMC7721 and Huh7 lines, in vivo assays | [101] | ||
| Inhibition (prevents TGF-β1-induced Wnt activation, reverses N-cadherin, vimentin, cyclin-D1, β-catenin overexpression counteracts) | U251 glioma cells | [102] | ||
| Inhibition (antagonizes fibrosis, decreases β-catenin, Wnt3, Wnt4, FZD4, p-LRPs, LEF1, TCF1, Snail, Jagged, E-cadherin, Twist, MMP2, MMP7) | Rat model of unilateral ureteral obstruction | [24] | ||
| Inhibition (suppresses β-catenin expression) | Mouse keratinocytes (LiCl-induced impaired re-epithelialization model) | [25] | ||
| 2.8. Iridoids | Iridoid-inspired | Inhibition (20 derivatives) | HEK293 reporter cell lines | [103] |
| Penta-acetyl geniposide | Inhibition (reduces Wnt1, p-GSK3β, β-catenin, inhibits β-catenin nuclear translocation) | TNFalpha-stimulated MH7A cell line (in vitro), rat model of collagen-induced arthritis (in vivo) | [12] | |
| Cornin | Activation (enhances Wnt5a, β-catenin, cyclin D1) | Arterial smooth muscle cell line (in vitro), rat model of stroke (in vivo) | [14] | |
| Catalpol | Inhibition (reduces Wnt3a, p-GSK3β, β-catenin, Snail1) | Rat model of renal fibrosis, proximal tubular line HK-2 | [5] | |
| Catalpol | Activation (enhances β-catenin, LEF1, TCF1/7, increases p-GSK3β, nuclear β-catenin) | Bone marrow mesenchymal stem cells | [104] | |
| Harpa-goside | Activation (increases DKK1, cyclin D1, c-Myc, β-catenin, suppresses p-β-catenin) | Osteoblast line MC3T3-E1 | [105] | |
| Harpagide | Activation (increases β-catenin) | Rat model of spinal cord injury (in vivo) | [17] | |
| Morroniside | Activation (upregulates Wnt10b, β-catenin, LEF1) | Outer root sheath cells (in vitro) | [106] | |
| Activation (increases Wnt3a, β-catenin, TCF4, Ngn2, Pax6) | Rat model of middle cerebral artery occlusion (in vivo) | [15] | ||
| Harpa-goside | Activation (upregulates β-catenin, cyclin D1, c-Myc, downregulates Dkk1) | Osteoblast line MC3T3-E1 | [105] | |
| Iridoid-inspired | Inhibition (potency related to C-1 substituents) | HEK293 reporter cell line stimulated by Wnt3a | [107] | |
| 2.9. Terpenoids | Jatrophone | Inhibition (interferes with Wnt/β-catenin signaling) | Human triple-negative breast cancer cells | [34] |
| Andrographolide | Activation (inhibits GSK3β) | Primary hippocampal neuron cultures, in vivo in rats | [108] | |
| Tigliane diterpenoids | Inhibition (reduces Axin2, c-myc, cyclin D, promotes β-catenin phosphorylation and degradation) | HEK293 and HEK293W cells (luciferase assay) | [35,109] | |
| Shizukaol D | Inhibition (downregulates β-catenin, LRP, Dvl2, Axin2) | Liver cancer cells | [109] | |
| 2.10. Coumarins | Umbelli-feron | Activation (increases β-catenin, ESR1/β-catenin pathway) | Cultured hOBs | [110] |
| Angelicin | Activation (promotes β-catenin and runt-related transcription factor 2, increases ERα) | Human osteoblast culture | [111] | |
| 2.11. Flavonoids and phenolics | Galangin | Inhibition (decreases β-catenin, LRP6, FZD1, reduces p-GSK3β, increases p-β-catenin) | Rat model of middle cerebral artery occlusion (stroke model) | [16] |
| Hesperidin | Activation (increases RUNX2, cyclin D1, β-catenin) | Human alveolar osteoblasts | [112] | |
| Genistin | Inhibition (decreases Wnt1 and Wnt3a, increases p-GSK3β/GSK3β ratio) | Glioma U-87 cells | [113] | |
| Formo-nonetin | Activation (increases Wnt10b and LRP5, promotes nuclear β-catenin, p-GSK3β Ser9, decreases DKK2 via AMPK) | 3T3-L1 pre-adipocytes | [13] | |
| Sulfonated formononetin | Activation (increases Wnt5a, β-catenin, cyclin D1) | Human umbilical vein endothelial cells | [114] | |
| Formononetin + dithiocarbamate | Inhibition (increases Axin, decreases β-catenin and TCF4) | PC-3 cells | [115] | |
| Emodin | Inhibition (reduces β-catenin) | Rat retinal ischemia model | [26] | |
| Inhibition (reduces p-GSK3β Ser9, β-catenin, EMT-related Wnt target genes) | HepG2 hepatocellular carcinoma | [69] | ||
| Inhibition (reduces p-GSK3β Ser9, β-catenin, EMT-related Wnt target genes) | A2780 and SK-OV-3 epithelial ovarian cancer lines | [116] | ||
| Aloe-emodin | Inhibition (reduces Wnt3a, increases p-β-catenin/β-catenin ratio, reduces p-GSK3β) | Melanoma cell lines A375 and SK-MEL-28, in vivo model | [117] | |
| Inhibition (decreases β-catenin, cyclin D1, c-Myc, pro-oxidant effects) | Androgen-independent DU145 prostate cancer cells | [118] | ||
| ethylamino-emodin | Inhibition (direct inhibitor of GSK3β, complex effects on β-catenin localization) | HepG2, HEK293, and primary hepatocytes | [119] | |
| 2.12. N-containing compounds | Ricinine | Activation (stimulates Wnt cascade, target is CK1) | HEK293 TopFlash reporter cells | [36] |
| Berberine | Activation (promotes total and nuclear β-catenin) | Bone mesenchymal stem cells | [38] | |
| Inhibition (reduces cytoplasmic β-catenin) | HCT116 human colon carcinoma cells | [120] | ||
| Matrine | Inhibition (inhibits VEGF, regulates Wnt/β-catenin signaling) | Breast cancer cells | [37] | |
| Inhibition (via PI3K/AKT/mTOR and AKT/GSK3/β-catenin pathways) | Hepatocellular carcinoma (HCC) cells | [121] | ||
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2025 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Koval, A.; Mamadalieva, N.Z.; Mamadalieva, R.; Jalilov, F.; Katanaev, V.L. Success and Controversy of Natural Products as Therapeutic Modulators of Wnt Signaling and Its Interplay with Oxidative Stress: Comprehensive Review Across Compound Classes and Experimental Systems. Antioxidants 2025, 14, 591. https://doi.org/10.3390/antiox14050591
Koval A, Mamadalieva NZ, Mamadalieva R, Jalilov F, Katanaev VL. Success and Controversy of Natural Products as Therapeutic Modulators of Wnt Signaling and Its Interplay with Oxidative Stress: Comprehensive Review Across Compound Classes and Experimental Systems. Antioxidants. 2025; 14(5):591. https://doi.org/10.3390/antiox14050591
Chicago/Turabian StyleKoval, Alexey, Nilufar Z. Mamadalieva, Rano Mamadalieva, Fazliddin Jalilov, and Vladimir L. Katanaev. 2025. "Success and Controversy of Natural Products as Therapeutic Modulators of Wnt Signaling and Its Interplay with Oxidative Stress: Comprehensive Review Across Compound Classes and Experimental Systems" Antioxidants 14, no. 5: 591. https://doi.org/10.3390/antiox14050591
APA StyleKoval, A., Mamadalieva, N. Z., Mamadalieva, R., Jalilov, F., & Katanaev, V. L. (2025). Success and Controversy of Natural Products as Therapeutic Modulators of Wnt Signaling and Its Interplay with Oxidative Stress: Comprehensive Review Across Compound Classes and Experimental Systems. Antioxidants, 14(5), 591. https://doi.org/10.3390/antiox14050591