
A Novel Hybrid Peptide VLP-Aβ Exhibits Antioxidant Activity In Vitro and In Vivo via KEAP1-NRF2-ARE Signaling Pathway

 , ,
, ,
Abstract
1. Introduction
2. Materials and Methods
2.1. Chemicals and Reagents
2.2. Cell Culture
2.3. H2O2-Induced HepG2 Cell Damage Model
2.4. Protective Effect of the Hybrid Peptide, VA, on H2O2-Induced Cell Damage
2.5. Detection of Cellular ROS Levels
2.6. Detection of Cellular Mitochondrial Damage
2.7. Measurement of the Cellular Antioxidant Enzyme Activity
2.8. Small Interfering RNA (siRNA) Transfection
2.9. qRT-PCR
2.10. Western Blot
2.11. Animal Models
2.11.1. Serum Biochemical Analysis
2.11.2. Measurement of Antioxidant Enzyme Activity in the Serum and Liver
2.11.3. Liver Tissue Histological Analysis
2.11.4. Liver Tissue qRT-PCR Analysis
2.12. Statistical Analysis
3. Results
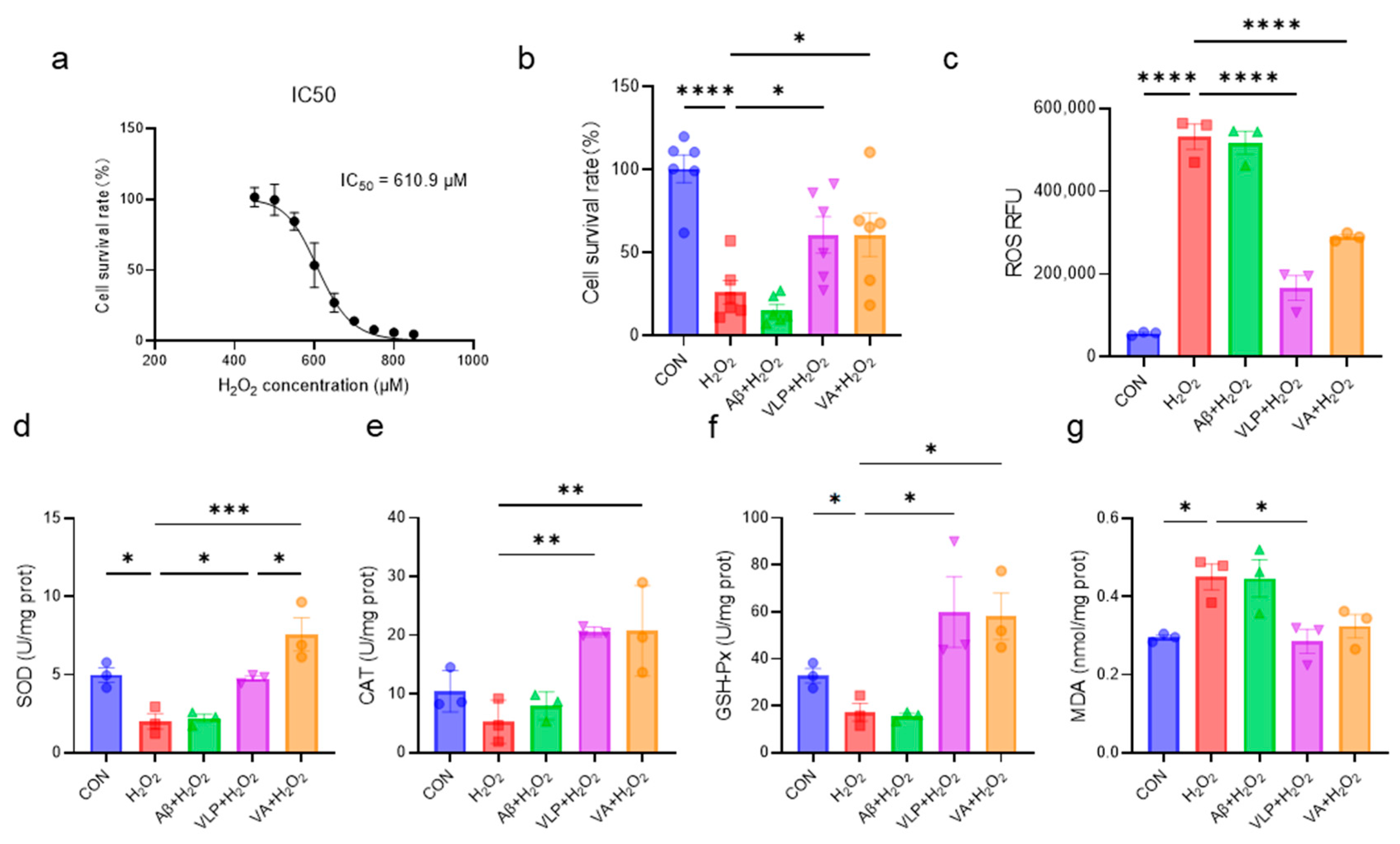
3.1. The Novel Hybrid Peptide, VA, Alleviates H2O2-Induced Cell Damage More Effectively than Its Parental Peptides
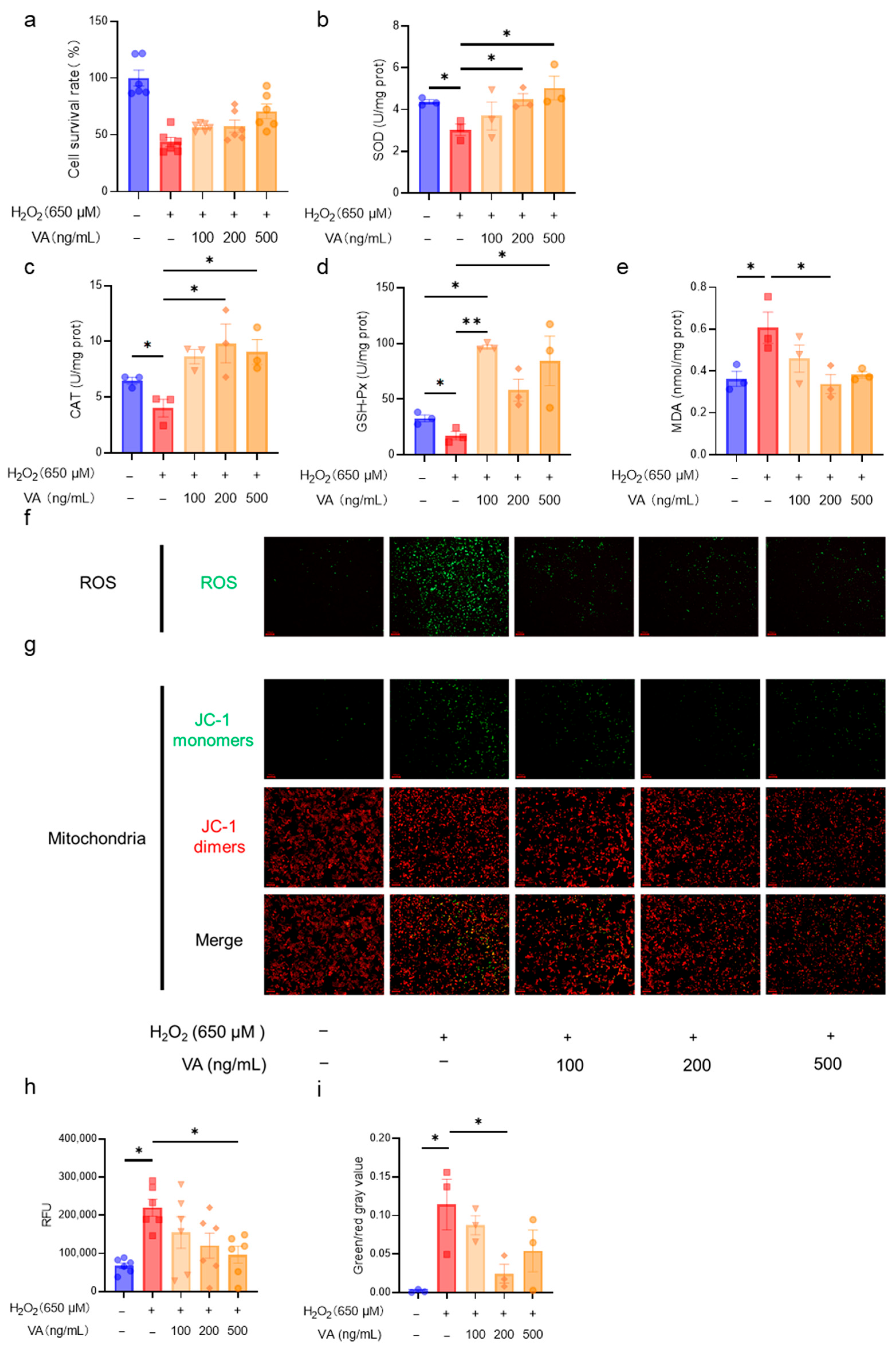
3.2. The Novel Hybrid Peptide, VA, Alleviates H2O2-Induced Cell Damage in a Dose-Dependent Manner
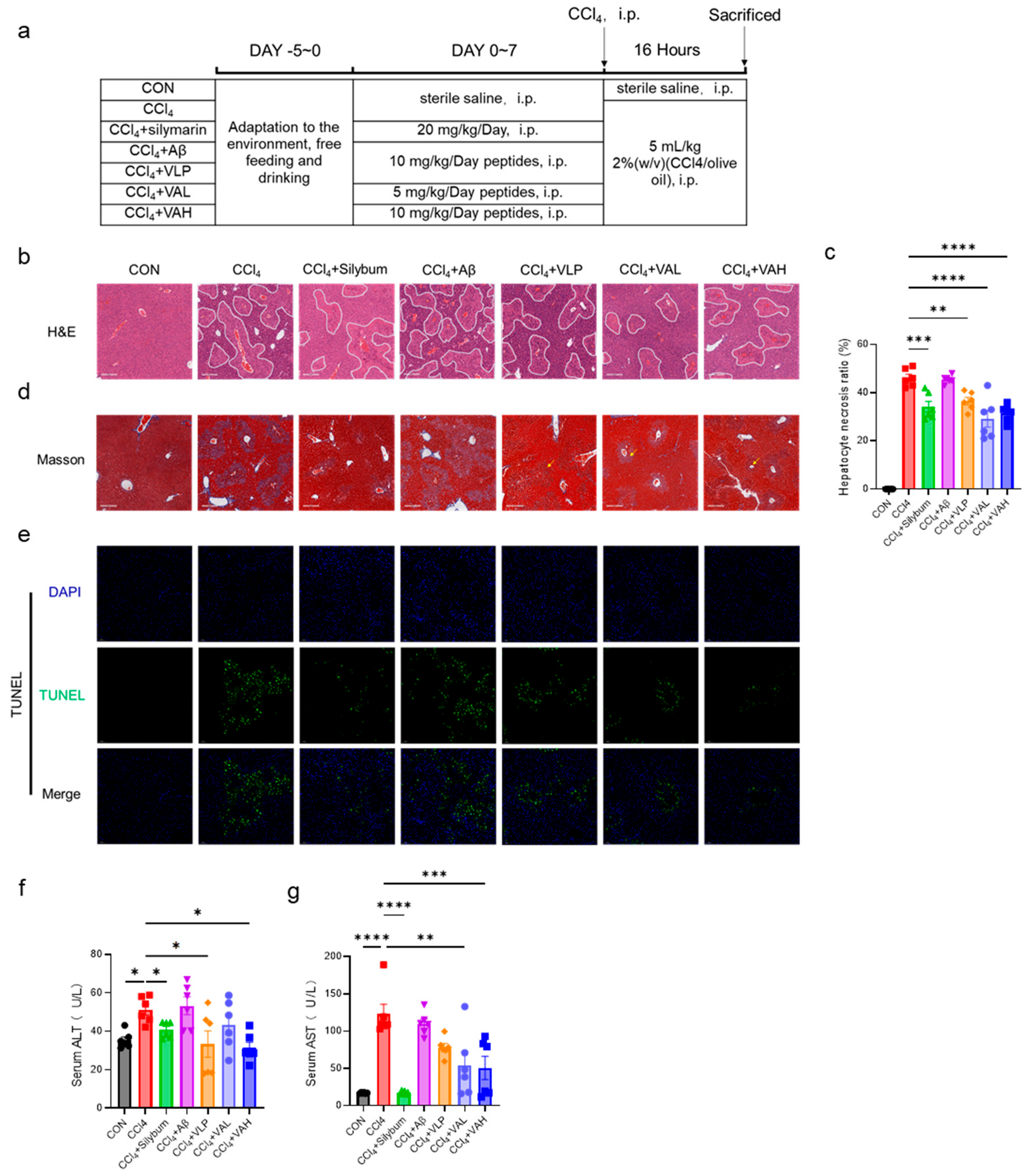
3.3. The Novel Hybrid Peptide, VA, Alleviates CCl4-Induced Liver Damage in Mice by Enhancing the Antioxidant Enzyme Activity
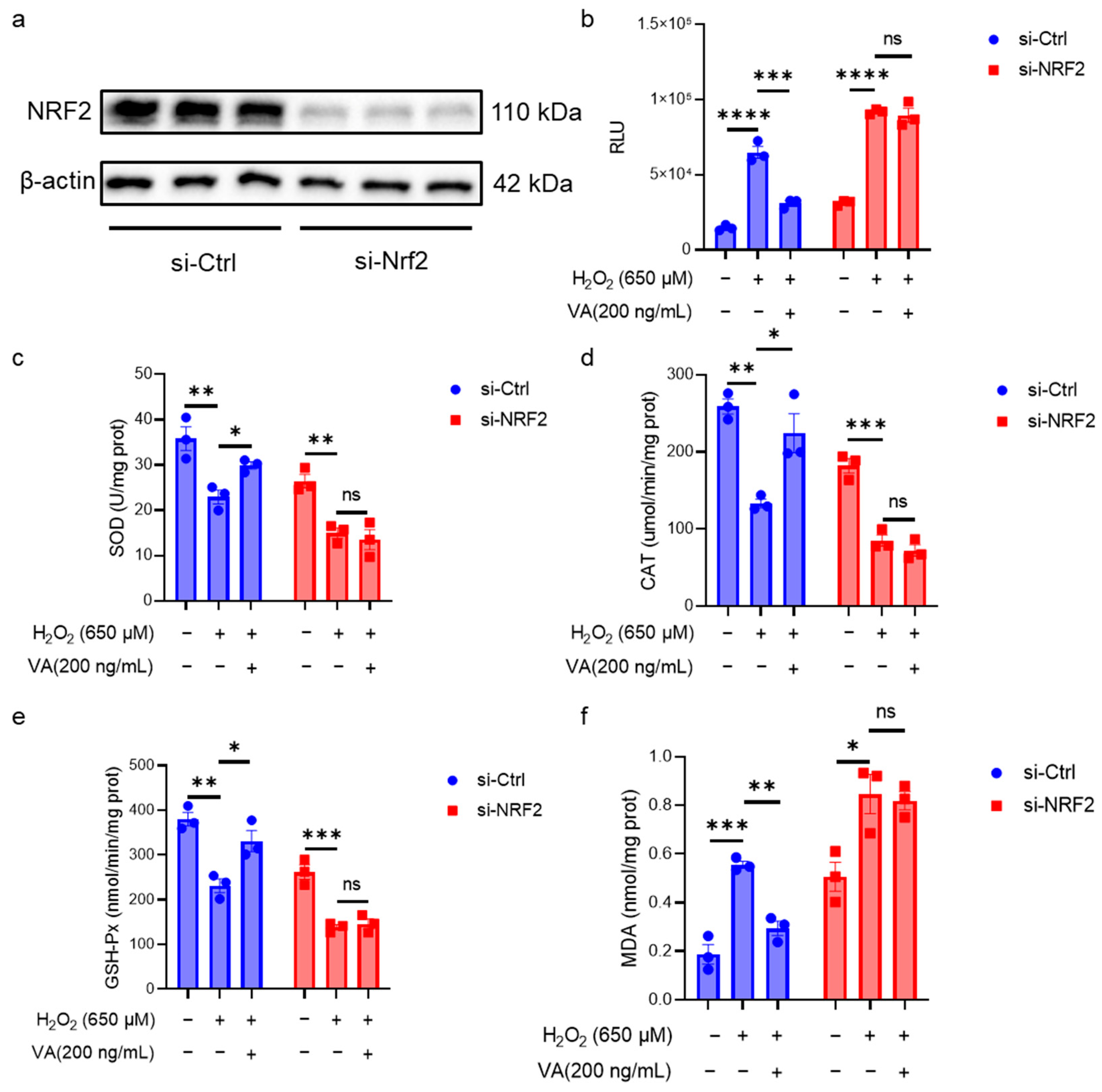
3.4. The Antioxidant Activity of the Hybrid Peptide, VA, Is Dependent on NRF2
3.5. The Novel Hybrid Peptide, VA, Alleviates Oxidative Stress by Modulating the KEAP1-NRF2-ARE Signaling Pathway
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Halliwell, B. Understanding mechanisms of antioxidant action in health and disease. Nat. Rev. Mol. Cell Biol. 2023, 25, 13–33. [Google Scholar] [CrossRef]
- Kishida, K.T.; Hoeffer, C.A.; Hu, D.; Pao, M.; Holland, S.M.; Klann, E. Synaptic plasticity deficits and mild memory impairments in mouse models of chronic granulomatous disease. Mol. Cell. Biol. 2006, 26, 5908–5920. [Google Scholar] [CrossRef]
- Lv, C.; Huang, Y.; Wang, Q.; Wang, C.; Hu, H.; Zhang, H.; Lu, D.; Jiang, H.; Shen, R.; Zhang, W.; et al. Ainsliadimer A induces ROS-mediated apoptosis in colorectal cancer cells via directly targeting peroxiredoxin 1 and 2. Cell Chem. Biol. 2023, 30, 295–307.e295. [Google Scholar] [CrossRef]
- Milton, V.J.; Sweeney, S.T. Oxidative stress in synapse development and function. Dev. Neurobiol. 2012, 72, 100–110. [Google Scholar] [CrossRef] [PubMed]
- Halliwell, B.; Adhikary, A.; Dingfelder, M.; Dizdaroglu, M. Hydroxyl radical is a significant player in oxidative DNA damage in vivo. Chem. Soc. Rev. 2021, 50, 8355–8360. [Google Scholar] [CrossRef] [PubMed]
- Davis, S.J.; Zhao, Y.; Yu, T.C.; Maytin, E.V.; Anand, S.; Hasan, T.; Pogue, B.W. Singlet Molecular Oxygen: From COIL Lasers to Photodynamic Cancer Therapy. J. Phys. Chem. B 2023, 127, 2289–2301. [Google Scholar] [CrossRef] [PubMed]
- Sies, H.; Belousov, V.V.; Chandel, N.S.; Davies, M.J.; Jones, D.P.; Mann, G.E.; Murphy, M.P.; Yamamoto, M.; Winterbourn, C. Defining roles of specific reactive oxygen species (ROS) in cell biology and physiology. Nat. Rev. Mol. Cell Biol. 2022, 23, 499–515. [Google Scholar] [CrossRef]
- Dizdaroglu, M.; Coskun, E.; Jaruga, P. Measurement of oxidatively induced DNA damage and its repair, by mass spectrometric techniques. Free Radic. Res. 2015, 49, 525–548. [Google Scholar] [CrossRef]
- Li, C.; Xue, Y.; Ba, X.; Wang, R. The Role of 8-oxoG Repair Systems in Tumorigenesis and Cancer Therapy. Cells 2022, 11, 3798. [Google Scholar] [CrossRef]
- Miao, X.; Liu, X.; Chen, H.; Wang, C.; Diao, J. Restoration and preservation effects of mung bean antioxidant peptides on H2O2-induced WRL-68 cells via Keap1-Nrf2 pathway. Food Sci. Nutr. 2023, 11, 7130–7144. [Google Scholar] [CrossRef]
- Pei, X.-D.; He, Y.-N.; Wu, Q.-L.; Zhang, Y.-M.; Li, F.; Jiao, D.-Q.; Liu, X.-L.; Wang, C.-H. Novel Antioxidant Peptides Derived from Feather Keratin Alleviate H2O2-Induced Oxidative Damage in HepG2 Cells via Keap1/Nrf2 Pathway. J. Agric. Food Chem. 2023, 71, 20062–20072. [Google Scholar] [CrossRef] [PubMed]
- Xiao, Y.; Yang, J.; Deng, Y.; Zhang, L.; Xu, Q.; Li, H. Tartary buckwheat protein-derived peptide AFYRW alleviates H2O2-induced vascular injury via the PI3K/AKT/NF-κB pathway. Prostaglandins Other Lipid Mediat. 2023, 169, 106768. [Google Scholar] [CrossRef]
- Wang, J.; Zhang, R.; Zhao, X.; Zhang, J.; Tong, Y.; Abbas, Z.; Li, Z.; Zhang, H.; Si, D.; Wei, X. Hybridization Design and High-Throughput Screening of Peptides with Immunomodulatory and Antioxidant Activities. Int. J. Mol. Sci. 2025, 26, 505. [Google Scholar] [CrossRef]
- Shanmugam, V.P.; Kapila, S.; Sonfack, T.K.; Kapila, R. Antioxidative peptide derived from enzymatic digestion of buffalo casein. Int. Dairy J. 2015, 42, 1–5. [Google Scholar] [CrossRef]
- Devi, S.; Kumar, N.; Kapila, S.; Mada, S.B.; Reddi, S.; Vij, R.; Kapila, R. Buffalo casein derived peptide can alleviates H(2)O(2) induced cellular damage and necrosis in fibroblast cells. Exp. Toxicol. Pathol. 2017, 69, 485–495. [Google Scholar] [CrossRef]
- Li, D.; Cao, J.; Zhang, J.; Mu, T.; Wang, R.; Li, H.; Tang, H.; Chen, L.; Lin, X.; Peng, X.; et al. The Effects and Regulatory Mechanism of Casein-Derived Peptide VLPVPQK in Alleviating Insulin Resistance of HepG2 Cells. Foods 2023, 12, 2627. [Google Scholar] [CrossRef]
- Mada, S.B.; Reddi, S.; Kumar, N.; Kumar, R.; Kapila, S.; Kapila, R.; Trivedi, R.; Karvande, A.; Ahmad, N. Antioxidative peptide from milk exhibits antiosteopenic effects through inhibition of oxidative damage and bone-resorbing cytokines in ovariectomized rats. Nutrition 2017, 43–44, 21–31. [Google Scholar] [CrossRef] [PubMed]
- Kumar, N.; Reddi, S.; Devi, S.; Mada, S.B.; Kapila, R.; Kapila, S. Nrf2 dependent antiaging effect of milk-derived bioactive peptide in old fibroblasts. J. Cell. Biochem. 2019, 120, 9677–9691. [Google Scholar] [CrossRef]
- Liao, W.; Liu, S.; Liu, X.; Duan, S.; Xiao, S.; Yang, Z.; Cao, Y.; Miao, J. The purification, identification and bioactivity study of a novel calcium-binding peptide from casein hydrolysate. Food Funct. 2019, 10, 7724–7732. [Google Scholar] [CrossRef]
- He, Z.; Li, X.; Wang, Z.; Cao, Y.; Han, S.; Li, N.; Cai, J.; Cheng, S.; Liu, Q. Protective effects of luteolin against amyloid beta-induced oxidative stress and mitochondrial impairments through peroxisome proliferator-activated receptor γ-dependent mechanism in Alzheimer’s disease. Redox Biol. 2023, 66, 102848. [Google Scholar] [CrossRef]
- Li, R.; Zhang, P.; Li, C.; Yang, W.; Yin, Y.; Tao, K. Tert-butylhydroquinone mitigates Carbon Tetrachloride induced Hepatic Injury in mice. Int. J. Med. Sci. 2020, 17, 2095–2103. [Google Scholar] [CrossRef] [PubMed]
- Down, W.H.; Chasseaud, L.F.; Grundy, R.K. Effect of silybin on the hepatic microsomal drug-metabolising enzyme system in the rat. Arzneimittelforschung 1974, 24, 1986–1988. [Google Scholar]
- Mostafa-Hedeab, G.; Ewaiss Hassan, M.; Halawa, T.F.; Ahmed Wani, F. Epigallocatechin gallate ameliorates tetrahydrochloride-induced liver toxicity in rats via inhibition of TGFβ/p-ERK/p-Smad1/2 signaling, antioxidant, anti-inflammatory activity. Saudi Pharm. J. SPJ Off. Publ. Saudi Pharm. Soc. 2022, 30, 1293–1300. [Google Scholar] [CrossRef]
- Wang, B.; Cui, S.; Mao, B.; Zhang, Q.; Tian, F.; Zhao, J.; Tang, X.; Chen, W. Cyanidin Alleviated CCl(4)-Induced Acute Liver Injury by Regulating the Nrf2 and NF-κB Signaling Pathways. Antioxidants 2022, 11, 2383. [Google Scholar] [CrossRef] [PubMed]
- Lu, X.; Gu, X.; Shi, Y. A review on lignin antioxidants: Their sources, isolations, antioxidant activities and various applications. Int. J. Biol. Macromol. 2022, 210, 716–741. [Google Scholar] [CrossRef]
- Chaudhary, P.; Janmeda, P.; Docea, A.O.; Yeskaliyeva, B.; Abdull Razis, A.F.; Modu, B.; Calina, D.; Sharifi-Rad, J. Oxidative stress, free radicals and antioxidants: Potential crosstalk in the pathophysiology of human diseases. Front. Chem. 2023, 11, 1158198. [Google Scholar] [CrossRef]
- Abed, D.A.; Goldstein, M.; Albanyan, H.; Jin, H.; Hu, L. Discovery of direct inhibitors of Keap1–Nrf2 protein–protein interaction as potential therapeutic and preventive agents. Acta Pharm. Sin. B 2015, 5, 285–299. [Google Scholar] [CrossRef] [PubMed]
- Chen, B.; Miao, J.; Ye, H.; Xia, Z.; Huang, W.; Guo, J.; Liang, X.; Yin, Y.; Zheng, Y.; Cao, Y. Purification, Identification, and Mechanistic Investigation of Novel Selenium-Enriched Antioxidant Peptides from Moringa oleifera Seeds. J. Agric. Food Chem. 2023, 71, 4625–4637. [Google Scholar] [CrossRef]
- Sun, C.; Tang, X.; Ren, Y.; Wang, E.; Shi, L.; Wu, X.; Wu, H. Novel Antioxidant Peptides Purified from Mulberry (Morus atropurpurea Roxb.) Leaf Protein Hydrolysates with Hemolysis Inhibition Ability and Cellular Antioxidant Activity. J. Agric. Food Chem. 2019, 67, 7650–7659. [Google Scholar] [CrossRef]
- Wei, X.; Zhang, L.; Yang, Y.; Hou, Y.; Xu, Y.; Wang, Z.; Su, H.; Han, F.; Han, J.; Liu, P.; et al. LL-37 transports immunoreactive cGAMP to activate STING signaling and enhance interferon-mediated host antiviral immunity. Cell Rep. 2022, 39, 110880. [Google Scholar] [CrossRef]
- Mailloux, R.J. An update on methods and approaches for interrogating mitochondrial reactive oxygen species production. Redox Biol. 2021, 45, 102044. [Google Scholar] [CrossRef]
- Arzumanian, V.A.; Kiseleva, O.I.; Poverennaya, E.V. The Curious Case of the HepG2 Cell Line: 40 Years of Expertise. Int. J. Mol. Sci. 2021, 22, 3135. [Google Scholar] [CrossRef] [PubMed]
- Long, X.; Wang, P.; Zhou, Y.; Wang, Q.; Ren, L.; Li, Q.; Zhao, X. Preventive effect of Lactobacillus plantarum HFY15 on carbon tetrachloride (CCl(4) )-induced acute liver injury in mice. J. Food Sci. 2022, 87, 2626–2639. [Google Scholar] [CrossRef]
- Hilpert, K.; Gani, J.; Rumancev, C.; Simpson, N.; Lopez-Perez, P.M.; Garamus, V.M.; von Gundlach, A.R.; Markov, P.; Scocchi, M.; Mikut, R.; et al. Rational Designed Hybrid Peptides Show up to a 6-Fold Increase in Antimicrobial Activity and Demonstrate Different Ultrastructural Changes as the Parental Peptides Measured by BioSAXS. Front. Pharmacol. 2021, 12, 769739. [Google Scholar] [CrossRef]
- Zhang, Y.; Zhong, Y.; Connor, A.L.; Miller, D.P.; Cao, R.; Shen, J.; Song, B.; Baker, E.S.; Tang, Q.; Pulavarti, S.; et al. Folding and Assembly of Short α, β, γ-Hybrid Peptides: Minor Variations in Sequence and Drastic Differences in Higher-Level Structures. J. Am. Chem. Soc. 2019, 141, 14239–14248. [Google Scholar] [CrossRef]
- Liu, S.; Pi, J.; Zhang, Q. Signal amplification in the KEAP1-NRF2-ARE antioxidant response pathway. Redox Biol. 2022, 54, 102389. [Google Scholar] [CrossRef] [PubMed]
- Arroyave-Ospina, J.C.; Wu, Z.; Geng, Y.; Moshage, H. Role of Oxidative Stress in the Pathogenesis of Non-Alcoholic Fatty Liver Disease: Implications for Prevention and Therapy. Antioxidants 2021, 10, 174. [Google Scholar] [CrossRef] [PubMed]
- Guo, Y.; Wu, Q.; Wu, Y.; Gao, L.; Yang, J. Effect of Lycium barbarum polysaccharide and aerobic exercise on rats with non-alcoholic fatty liver disease and its mechanism. J. Shanghai Jiaotong Univ. Med. Sci. 2020, 40, 30–36. [Google Scholar]
- Li, H.; Isaac, N.; He, S.; Ou, X.; Xiang, J.; Tian, X.; Zhu, H.; Liu, L.; Wen, L.; Chu, W. Dietary supplementation with protein hydrolysates from the shell of red swamp crayfish (Procambarus clarkii) affects growth, muscle antioxidant capacity and circadian clock genes expression of zebrafish (Danio rerio). Aquac. Rep. 2022, 27, 101390. [Google Scholar] [CrossRef]
- Barteková, M.; Adameová, A.; Görbe, A.; Ferenczyová, K.; Pecháňová, O.; Lazou, A.; Dhalla, N.S.; Ferdinandy, P.; Giricz, Z. Natural and synthetic antioxidants targeting cardiac oxidative stress and redox signaling in cardiometabolic diseases. Free Radic. Biol. Med. 2021, 169, 446–477. [Google Scholar] [CrossRef]
- Reis, R.; Charehsaz, M.; Sipahi, H.; Ekici, A.I.; Macit, Ç.; Akkaya, H.; Aydın, A. Energy Drink Induced Lipid Peroxidation and Oxidative Damage in Rat Liver and Brain When Used Alone or Combined with Alcohol. J. Food Sci. 2017, 82, 1037–1043. [Google Scholar] [CrossRef] [PubMed]
- Li, J.; Li, Y.; Lin, S.; Zhao, W.; Chen, Y.; Jin, H. Collagen peptides from Acaudina molpadioides prevent CCl(4)—induced liver injury via Keap1/Nrf2-ARE, PI3K/AKT, and MAPKs pathways. J. Food Sci. 2022, 87, 2185–2196. [Google Scholar] [CrossRef] [PubMed]
- Zhou, Y.; Zhang, R.; Wang, J.; Tong, Y.; Zhang, J.; Li, Z.; Zhang, H.; Abbas, Z.; Si, D.; Wei, X. Isolation, Characterization, and Functional Properties of Antioxidant Peptides from Mulberry Leaf Enzymatic Hydrolysates. Antioxidants 2024, 13, 854. [Google Scholar] [CrossRef]
- Raghunath, A.; Sundarraj, K.; Nagarajan, R.; Arfuso, F.; Bian, J.; Kumar, A.P.; Sethi, G.; Perumal, E. Antioxidant response elements: Discovery, classes, regulation and potential applications. Redox Biol. 2018, 17, 297–314. [Google Scholar] [CrossRef] [PubMed]
- Zhu, L.; He, S.; Huang, L.; Ren, D.; Nie, T.; Tao, K.; Xia, L.; Lu, F.; Mao, Z.; Yang, Q. Chaperone-mediated autophagy degrades Keap1 and promotes Nrf2-mediated antioxidative response. Aging Cell 2022, 21, e13616. [Google Scholar] [CrossRef]
- Li, L.; Liu, J.; Nie, S.; Ding, L.; Wang, L.; Liu, J.; Liu, W.; Zhang, T. Direct inhibition of Keap1–Nrf2 interaction by egg-derived peptides DKK and DDW revealed by molecular docking and fluorescence polarization. RSC Adv. 2017, 7, 34963–34971. [Google Scholar] [CrossRef]



{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Peptide | Sequence |
|---|---|
| Aβ | DAEFRHDSGYEVHHQKLVFFAEDVGSNKGAIIGLMVG |
| VLP | VLPVPQK |
| VLP-Aβ | VLPVPQKDAEFRHDSGYEVHHQKLVFFAEDVGSNKGAIIGLMVG |
| Gene Name | Gene Accession Numbers | Sequence (5′–3′) | Length | |
|---|---|---|---|---|
| mNrf2 | NM_010902.5 | F | GCTGGCTGATACTACCGCTGTTC | 23 |
| R | AGTGGAGAGGATGCTGCTGAAAGA | 24 | ||
| mHmox1 | NM_010442.2 | F | AGACCGCCTTCCTGCTCAACA | 21 |
| R | CTCTGACGAAGTGACGCCATCTG | 23 | ||
| mNqo1 | NM_008706.5 | F | GCGAGAAGAGCCCTGATTGTACTG | 24 |
| R | GCCTCTACAGCAGCCTCCTTCA | 22 | ||
| mSod2 | NM_013671.3 | F | TCCCAGACCTGCCTTACGACTATG | 24 |
| R | TTGATAGCCTCCAGCAACTCTCCTT | 25 | ||
| mβ-actin | NM_007393.5 | F | TCACTATTGGCAACGAGCGGTTC | 23 |
| R | CAGCACTGTGTTGGCATAGAGGTC | 24 | ||
| hHmox1 | NM_002133.3 | F | GCCAGCAACAAAGTGCAAGA | 20 |
| R | TAAGGACCCATCGGAGAAGC | 20 | ||
| hNqo1 | NM_000903.3 | F | TGGTGGAGTCGGACCTCTATG | 21 |
| R | CATGGCAGCGTAAGTGTAAGC | 21 | ||
| hβ-actin | NM_001101.5 | F | ATCGTCCACCGCAAATGCTTCT | 22 |
| R | TGCTGTCACCTTCACCGTTCCA | 22 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2025 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Wang, J.; Zhang, W.; Zhang, R.; Zhao, X.; Zhang, J.; Zhou, Y.; Tong, Y.; Abbas, Z.; Li, Z.; Zhang, H.; et al. A Novel Hybrid Peptide VLP-Aβ Exhibits Antioxidant Activity In Vitro and In Vivo via KEAP1-NRF2-ARE Signaling Pathway. Antioxidants 2025, 14, 583. https://doi.org/10.3390/antiox14050583
Wang J, Zhang W, Zhang R, Zhao X, Zhang J, Zhou Y, Tong Y, Abbas Z, Li Z, Zhang H, et al. A Novel Hybrid Peptide VLP-Aβ Exhibits Antioxidant Activity In Vitro and In Vivo via KEAP1-NRF2-ARE Signaling Pathway. Antioxidants. 2025; 14(5):583. https://doi.org/10.3390/antiox14050583
Chicago/Turabian StyleWang, Junyong, Wenxiu Zhang, Rijun Zhang, Xuelian Zhao, Jing Zhang, Yichen Zhou, Yucui Tong, Zaheer Abbas, Zhenzhen Li, Haosen Zhang, and et al. 2025. "A Novel Hybrid Peptide VLP-Aβ Exhibits Antioxidant Activity In Vitro and In Vivo via KEAP1-NRF2-ARE Signaling Pathway" Antioxidants 14, no. 5: 583. https://doi.org/10.3390/antiox14050583
APA StyleWang, J., Zhang, W., Zhang, R., Zhao, X., Zhang, J., Zhou, Y., Tong, Y., Abbas, Z., Li, Z., Zhang, H., Yang, D., Chen, S., Hu, C., Si, D., & Wei, X. (2025). A Novel Hybrid Peptide VLP-Aβ Exhibits Antioxidant Activity In Vitro and In Vivo via KEAP1-NRF2-ARE Signaling Pathway. Antioxidants, 14(5), 583. https://doi.org/10.3390/antiox14050583