
Anti-Melanogenic Activity of Ethanolic Extract from Garcinia atroviridis Fruits Using In Vitro Experiments, Network Pharmacology, Molecular Docking, and Molecular Dynamics Simulation


 , ,
, ,  ,
,
Abstract
1. Introduction
2. Materials and Methods
2.1. Materials
2.2. Plant Materials
2.3. Plant Preparation
2.4. Determination of Total Phenolic Content (TPC) and Total Flavonoid Content (TFC)
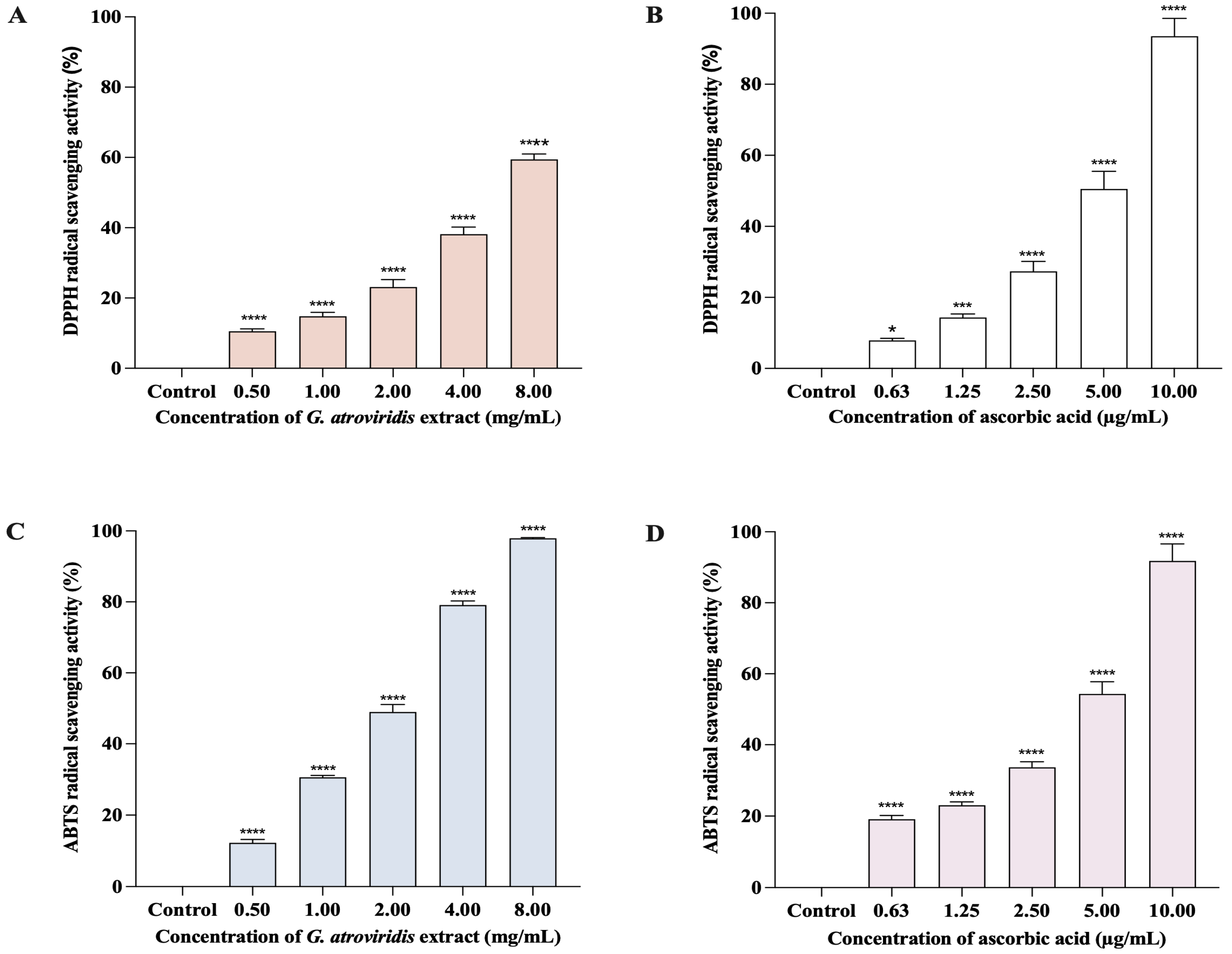
2.5. DPPH Radical Scavenging Activity
2.6. ABTS Radical Scavenging Activity
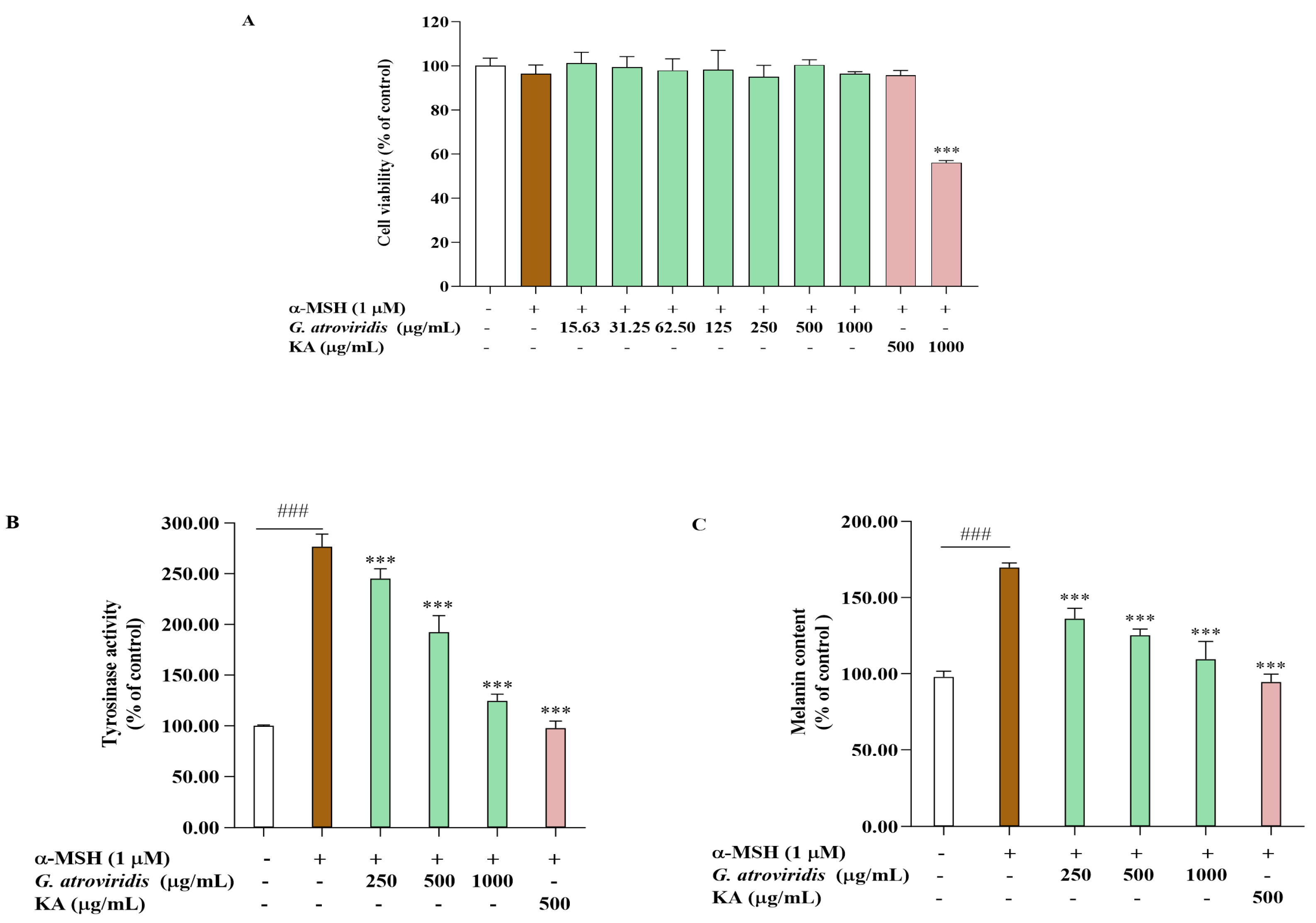
2.7. Cell Culture and Cell Viability Assay
2.8. Measurement of Intracellular Tyrosinase Activity and Melanin Content
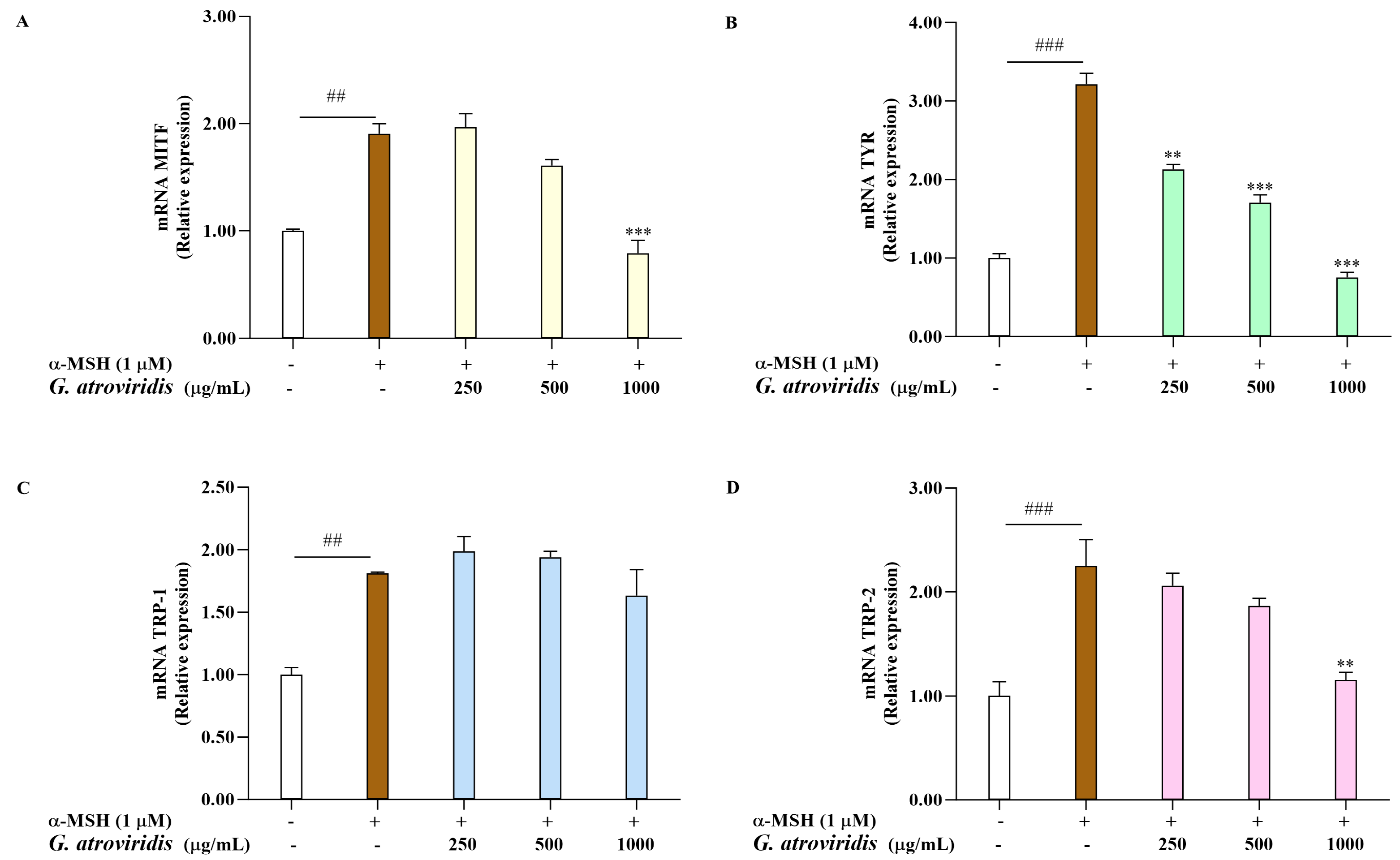
2.9. Quantitative Real-Time PCR (qRT-PCR)
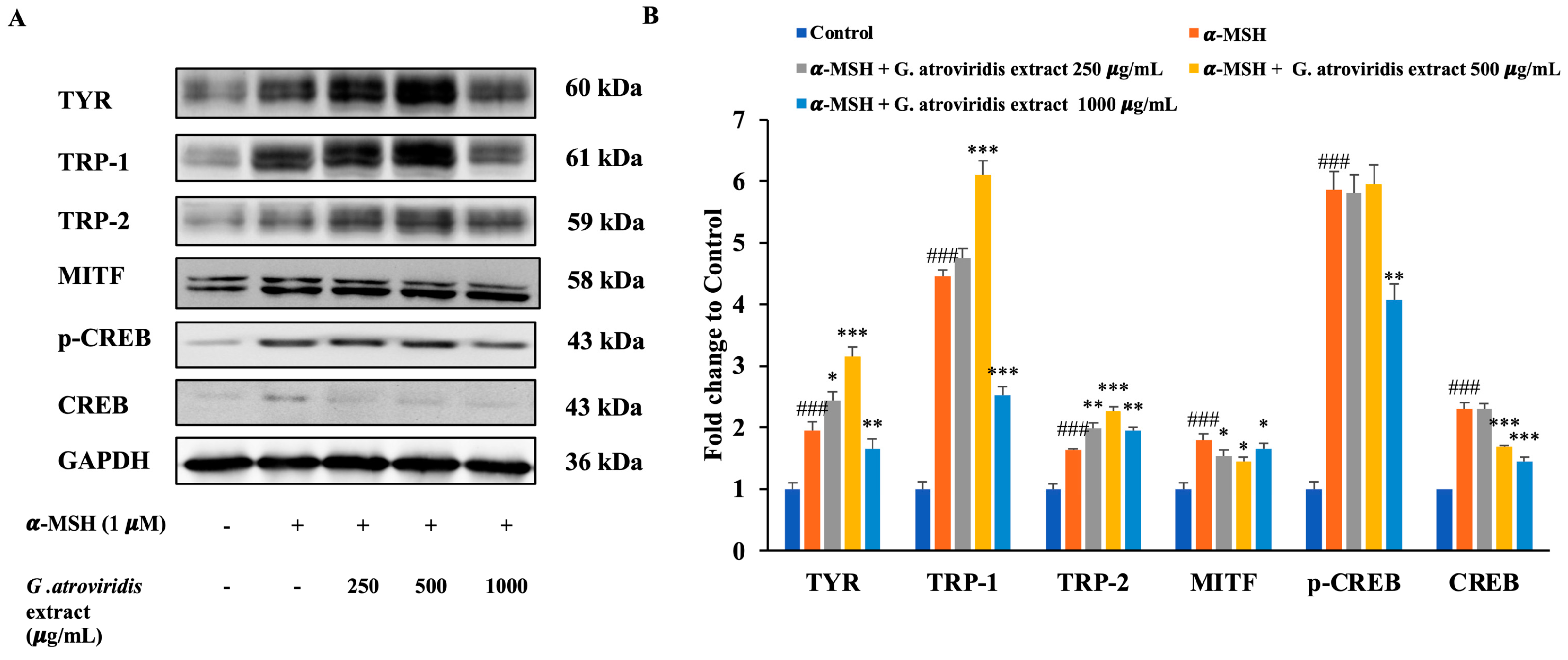
2.10. Western Blot Analysis
2.11. Copper-Chelating Assay (CCA)
2.12. Liquid Chromatography–Mass Spectrometry (LC-MS) Analysis
2.13. Chemoinformatics, Drug Likeness and Pharmacokinetic Prediction
2.14. Network Pharmacology Analysis
2.14.1. Target Protein Prediction
2.14.2. Disease Potential Targets
2.14.3. GO and KEGG Pathway Enrichment Analysis
2.14.4. Construction of Protein–Protein Interaction Network
2.15. Molecular Docking Verification
2.16. Molecular Dynamics Simulation
2.17. Statistical Analysis
3. Results and Discussion
3.1. Extraction Yield, Total Phenolic and Total Flavonoid Contents of Ethanolic Extract of G. atroviridis Fruits
3.2. Free Radical Scavenging Activities of Ethanolic Extract of G. atroviridis Fruits
3.3. Effects of Ethanolic Extract of G. atroviridis Fruits on Cell Viability, Intracellular Tyrosinase Activity, and Melanin Content
3.4. Effects of Ethanolic Extract of G. atroviridis Fruits on Melanogenesis-Related Gene Expression
3.5. Effects of Ethanolic Extract of G. atroviridis Fruits on Protein Expression of Melanogenesis Related Proteins
3.6. Effects of Ethanolic Extract of G. atroviridis Fruits on Copper-Chelating Activity
3.7. Liquid Chromatography–Mass Spectrometry (LC-MS) Analysis
3.8. Chemoinformatics, Drug Likeness, and Pharmacokinetic Properties of Compounds from Ethanolic Extract of G. atroviridis Fruits
3.9. Network Pharmacology Analysis
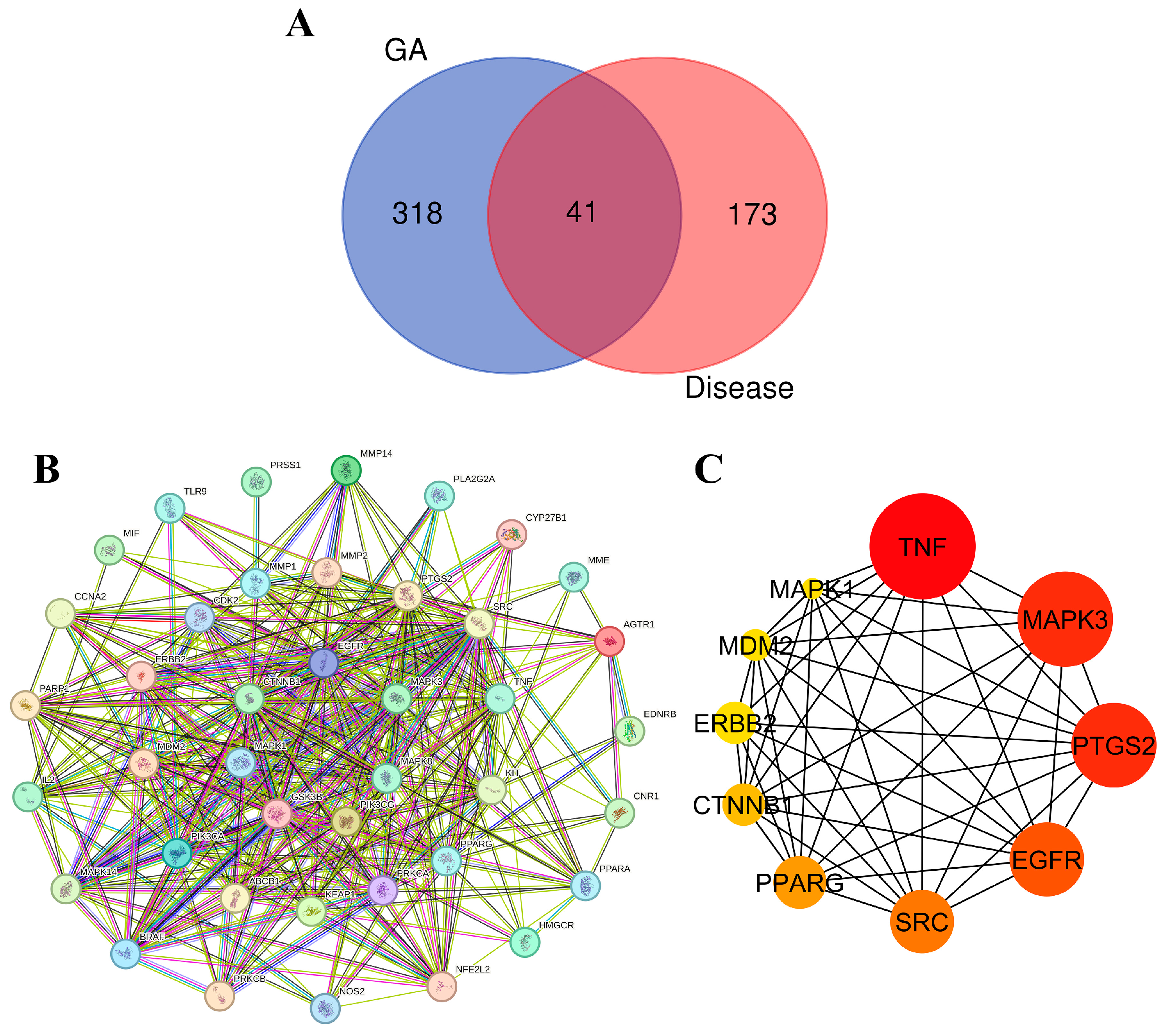
3.9.1. Target Identification and Analysis
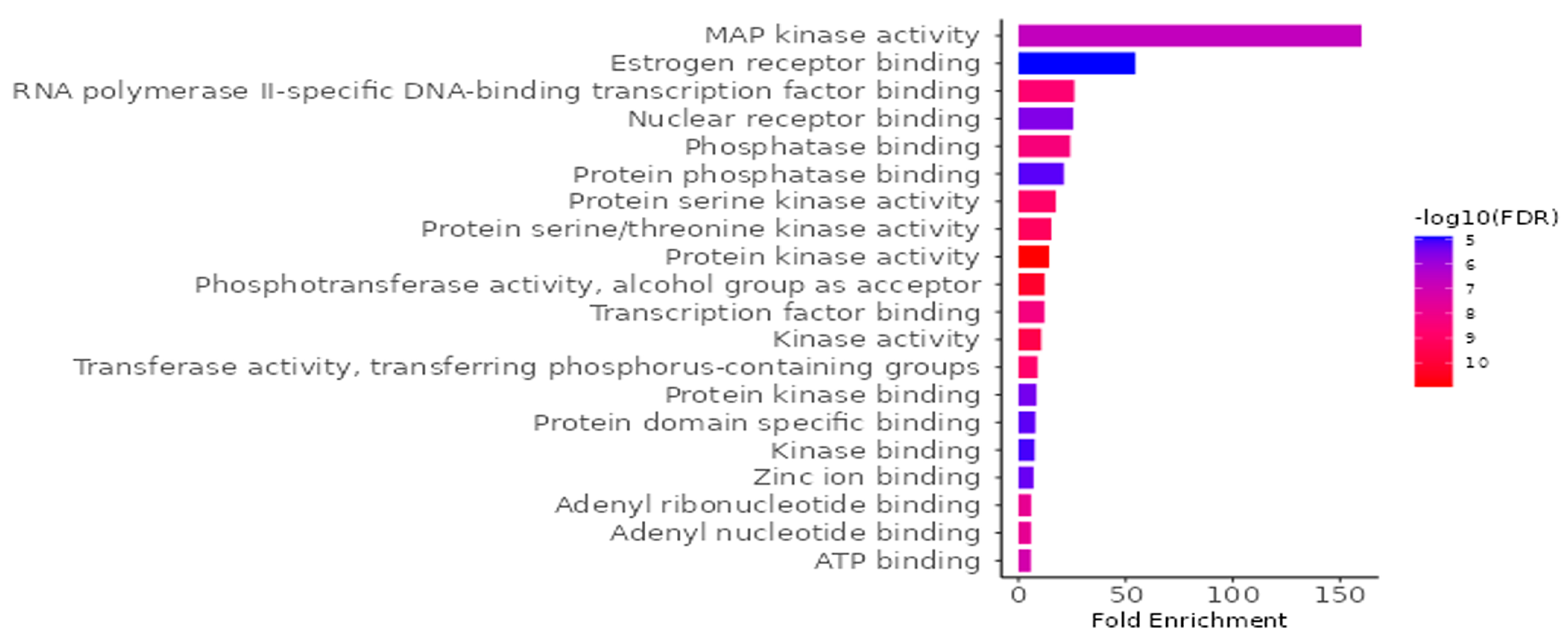
3.9.2. GO and KEGG Enrichment Analysis
3.9.3. Network Analysis of Protein–Protein Interactions (PPI) and Identification of Key Targets
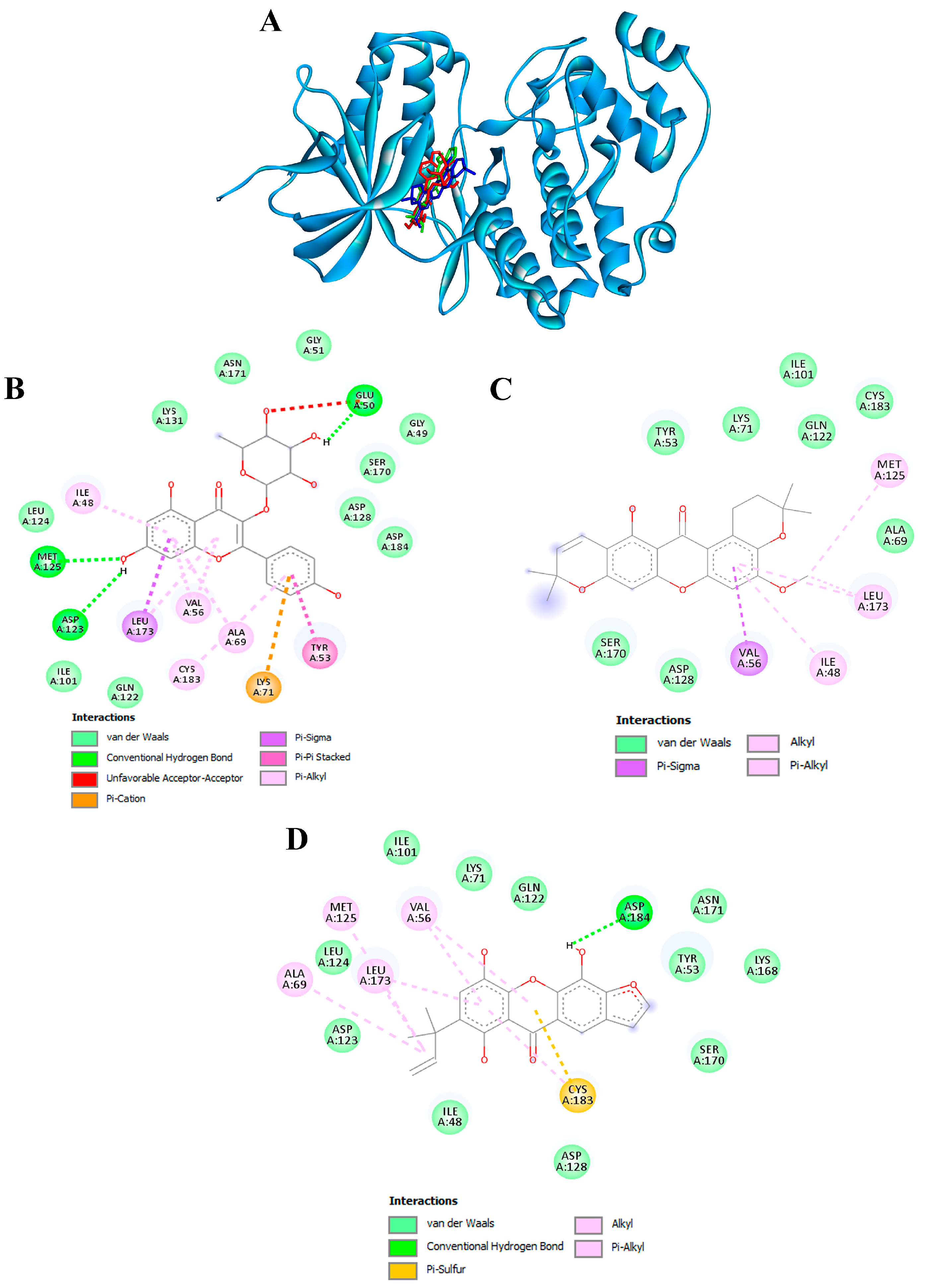
3.10. Confirmation of Hub Targets through Molecular Docking
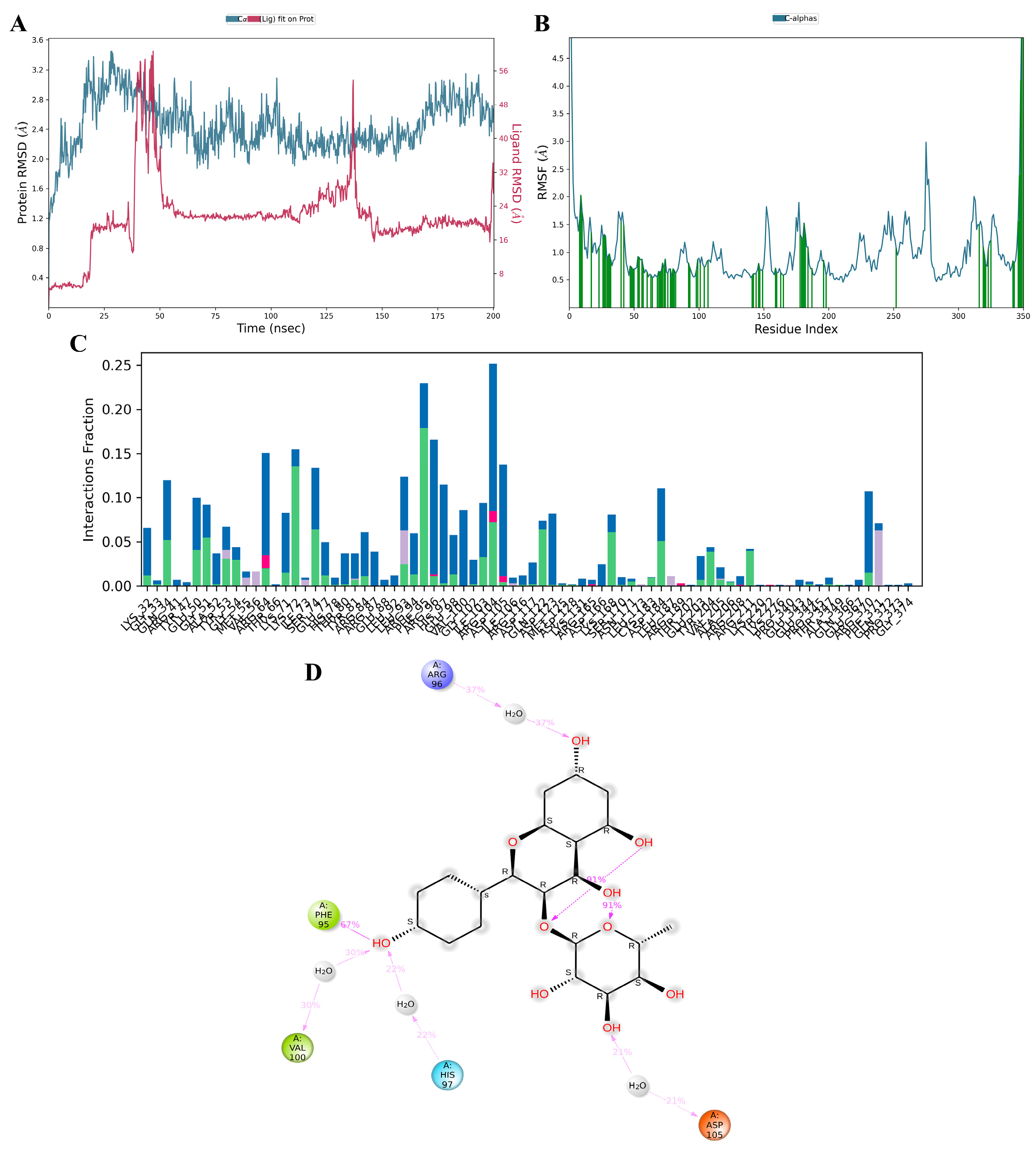
3.11. MD Simulation
4. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Brenner, M.; Hearing, V.J. The protective role of melanin against UV damage in human skin. Photochem. Photobiol. 2008, 84, 539–549. [Google Scholar] [CrossRef]
- Goelzer Neto, C.F.; do Nascimento, P.; da Silveira, V.C.; de Mattos, A.B.N.; Bertol, C.D. Natural sources of melanogenic inhibitors: A systematic review. Int. J. Cosmet. Sci. 2022, 44, 143–153. [Google Scholar] [CrossRef]
- Maddaleno, A.S.; Camargo, J.; Mitjans, M.; Vinardell, M.P. Melanogenesis and melasma treatment. Cosmetics 2021, 8, 82. [Google Scholar] [CrossRef]
- Gillbro, J.M.; Olsson, M.J. The melanogenesis and mechanisms of skin-lightening agents—Existing and new approaches. Int. J. Cosmet. Sci. 2011, 33, 210–221. [Google Scholar] [CrossRef]
- Shahid, M.; Law, D.; Azfaralariff, A.; Mackeen, M.M.; Chong, T.F.; Fazry, S. Phytochemicals and biological activities of Garcinia atroviridis: A critical review. Toxics 2022, 10, 656. [Google Scholar] [CrossRef]
- Tomar, M.; Rao, R.P.; Dorairaj, P.; Koshta, A.; Suresh, S.; Rafiq, M.; Kumawat, R.; Paramesh, R.; V, B.U.; Venkatesh, K.V. A clinical and computational study on anti-obesity effects of hydroxycitric acid. RSC Adv. 2019, 9, 18578–18588. [Google Scholar] [CrossRef]
- Mackeen, M.M.; Ali, A.M.; Lajis, N.H.; Kawazu, K.; Kikuzaki, H.; Nakatani, N. Antifungal garcinia acid esters from the fruits of Garcinia atroviridis. Z. Naturforsch. C. J. Biosci. 2002, 57, 291–295. [Google Scholar] [CrossRef]
- Liu, C.; Ho, P.C.; Wong, F.C.; Sethi, G.; Wang, L.Z.; Goh, B.C. Garcinol: Current status of its anti-oxidative, anti-inflammatory and anti-cancer effects. Cancer Lett. 2015, 362, 8–14. [Google Scholar] [CrossRef]
- Schobert, R.; Biersack, B. Chemical and biological aspects of garcinol and isogarcinol: Recent developments. Chem. Biodivers. 2019, 16, e1900366. [Google Scholar] [CrossRef] [PubMed]
- Tan, W.-N.; Wong, K.-C.; Khairuddean, M.; Eldeen, I.M.; Asmawi, M.Z.; Sulaiman, B. Volatile constituents of the fruit of Garcinia atroviridis and their antibacterial and anti-inflammatory activities. Flavour Fragr. J. 2013, 28, 2–9. [Google Scholar] [CrossRef]
- Shahid, M.; Fazry, S.; Azfaralariff, A.; Najm, A.A.K.; Law, D.; Mackeen, M.M. Bioactive compound identification and In Vitro evaluation of antidiabetic and cytotoxic potential of Garcinia atroviridis fruit extract. Food Biosci. 2023, 51, 102285. [Google Scholar] [CrossRef]
- Kongchian, A.; Keawboonlert, N.; Boonrak, T.; Lookyee, S.; Buasri, K.; Surongkul, N.; Tangpong, J. Anti-hyperlipidemia and anti-obesity properties of Garcinia atroviridis and Camellia sinensis extracts in high-fat diet mice. Walailak J. Sci. Technol. 2020, 17, 1126–1138. [Google Scholar] [CrossRef]
- Mackeen, M.M.; Ali, A.M.; Lajis, N.H.; Kawazu, K.; Hassan, Z.; Amran, M.; Habsah, M.; Mooi, L.Y.; Mohamed, S.M. Antimicrobial, antioxidant, antitumour-promoting and cytotoxic activities of different plant part extracts of Garcinia atroviridis Griff. ex T. Anders. J. Ethnopharmacol. 2000, 72, 395–402. [Google Scholar] [CrossRef] [PubMed]
- Muchtaridi, M.; Nuwarda, R.F.; Ikram, E.H.K.; Abdul Rahim, A.S.; Gazzali, A.M.; Wahab, H.A. Neuraminidase inhibitor of Garcinia atroviridis L. fruits and leaves using partial purification and molecular characterization. Molecules 2022, 27, 949. [Google Scholar] [CrossRef] [PubMed]
- Chatatikun, M.; Supjaroen, P.; Promlat, P.; Chantarangkul, P.; Waranuntakul, S.; Nawarat, J.; Tangpong, J.; Chiabchalard, A. Antioxidant and tyrosinase inhibitory properties of an aqueous extract of Garcinia atroviridis Griff. ex. T. anderson fruit pericarps. Pharmacogn. J. 2020, 12, 71–78. [Google Scholar] [CrossRef]
- Yang, M.; Chen, J.-L.; Xu, L.-W.; Ji, G. Navigating traditional chinese medicine network pharmacology and computational tools. Evid.-Based Complement. Altern. Med. 2013, 2013, 731969. [Google Scholar] [CrossRef] [PubMed]
- Zhou, Z.; Chen, B.; Chen, S.; Lin, M.; Chen, Y.; Jin, S.; Chen, W.; Zhang, Y. Applications of network pharmacology in traditional Chinese medicine research. Evid.-Based Complement. Altern. Med. 2020, 2020, 1646905. [Google Scholar] [CrossRef] [PubMed]
- Hopkins, A.L. Network pharmacology. Nat. Biotechnol. 2007, 25, 1110–1111. [Google Scholar] [CrossRef] [PubMed]
- Chakraborty, C.; Doss, C.G.; Chen, L.; Zhu, H. Evaluating protein-protein interaction (PPI) networks for diseases pathway, target discovery, and drug-design using ‘In Silico pharmacology’. Curr. Protein Pept Sci. 2014, 15, 561–571. [Google Scholar] [CrossRef]
- Zheng, S.; Xue, T.; Wang, B.; Guo, H.; Liu, Q. Application of network pharmacology in the study of mechanism of Chinese medicine in the treatment of ulcerative colitis: A review. Front. Bioinform. 2022, 2, 928116. [Google Scholar] [CrossRef]
- Agu, P.C.; Afiukwa, C.A.; Orji, O.U.; Ezeh, E.M.; Ofoke, I.H.; Ogbu, C.O.; Ugwuja, E.I.; Aja, P.M. Molecular docking as a tool for the discovery of molecular targets of nutraceuticals in diseases management. Sci. Rep. 2023, 13, 13398. [Google Scholar] [CrossRef] [PubMed]
- Macalino, S.J.Y.; Gosu, V.; Hong, S.; Choi, S. Role of computer-aided drug design in modern drug discovery. Arch. Pharm. Res. 2015, 38, 1686–1701. [Google Scholar] [CrossRef] [PubMed]
- Pinzi, L.; Rastelli, G. Molecular docking: Shifting paradigms in drug discovery. Int. J. Mol. Sci. 2019, 20, 4331. [Google Scholar] [CrossRef] [PubMed]
- Eggel, M.; Würbel, H. Internal consistency and compatibility of the 3Rs and 3Vs principles for project evaluation of animal research. Lab. Anim. 2020, 55, 233–243. [Google Scholar] [CrossRef] [PubMed]
- Tekleyes, B.; Huluka, S.A.; Wondu, K.; Wondmkun, Y.T. Wound healing activity of 80% methanol leaf extract of Zehneria scabra (L.f) sond (Cucurbitaceae) in mice. J. Exp. Pharmacol. 2021, 13, 537–544. [Google Scholar] [CrossRef] [PubMed]
- Chatatikun, M.; Chiabchalard, A. Thai plants with high antioxidant levels, free radical scavenging activity, anti-tyrosinase and anti-collagenase activity. BMC Complement. Altern. Med. 2017, 17, 487. [Google Scholar] [CrossRef] [PubMed]
- Lefahal, M.; Zaabat, N.; Ayad, R.; Makhloufi, E.H.; Djarri, L.; Benahmed, M.; Laouer, H.; Nieto, G.; Akkal, S. In Vitro assessment of total phenolic and flavonoid contents, antioxidant and photoprotective activities of crude methanolic extract of aerial parts of Capnophyllum peregrinum (L.) Lange (Apiaceae) growing in Algeria. Medicines 2018, 5, 26. [Google Scholar] [CrossRef] [PubMed]
- Chatatikun, M.; Tedasen, A.; Pattaranggoon, N.C.; Palachum, W.; Chuaijit, S.; Mudpan, A.; Pruksaphanrat, S.; Sohbenalee, S.; Yamasaki, K.; Klangbud, W.K. Antioxidant activity, anti-tyrosinase activity, molecular docking studies, and molecular dynamic simulation of active compounds found in nipa palm vinegar. PeerJ 2023, 11, e16494. [Google Scholar] [CrossRef] [PubMed]
- Wairata, J.; Fadlan, A.; Setyo Purnomo, A.; Taher, M.; Ersam, T. Total phenolic and flavonoid contents, antioxidant, antidiabetic and antiplasmodial activities of Garcinia forbesii King: A correlation study. Arab. J. Chem. 2022, 15, 103541. [Google Scholar] [CrossRef]
- Choi, M.R.; Lee, H.; Kim, H.K.; Han, J.; Seol, J.E.; Vasileva, E.A.; Mishchenko, N.P.; Fedoreyev, S.A.; Stonik, V.A.; Ju, W.S.; et al. Echinochrome A inhibits melanogenesis in B16F10 cells by downregulating CREB signaling. Mar. Drugs 2022, 20, 555. [Google Scholar] [CrossRef]
- Yu, C.-L.; Wu, H.; Chen, Y.-P.; Chen, F.; Wang, G.-H. Orcinol inhibits melanogenesis in B16F10 cells via the upregulation of the MAPK/ERK signaling pathway. Nat. Prod. Commun. 2023, 18, 1934578X231156704. [Google Scholar] [CrossRef]
- Kooltheat, N.; Tedasen, A.; Yamasaki, K.; Chatatikun, M. Melanogenesis inhibitory activity, chemical components and molecular docking studies of Prunus cerasoides Buch.-Ham. D. Don. Flowers. J. Evid.-Based Integr. Med. 2023, 28, 2515690x231152928. [Google Scholar] [CrossRef] [PubMed]
- Chatatikun, M.; Yamauchi, T.; Yamasaki, K.; Aiba, S.; Chiabchalard, A. Anti melanogenic effect of Croton roxburghii and Croton sublyratus leaves in α-MSH stimulated B16F10 cells. J. Tradit. Complement. Med. 2019, 9, 66–72. [Google Scholar] [CrossRef] [PubMed]
- Su, J.; Sun, Y.-Q.; Huo, F.-J.; Yang, Y.-T.; Yin, C.-X. Naked-eye determination of oxalate anion in aqueous solution with copper ion and pyrocatechol violet. Analyst 2010, 135, 2918–2923. [Google Scholar] [CrossRef]
- Zubair, M.S.; Maulana, S.; Widodo, A.; Pitopang, R.; Arba, M.; Hariono, M. GC-MS, LC-MS/MS, docking and molecular dynamics approaches to identify potential SARS-CoV-2 3-chymotrypsin-like protease inhibitors from Zingiber officinale Roscoe. Molecules 2021, 26, 5230. [Google Scholar] [CrossRef] [PubMed]
- Daina, A.; Michielin, O.; Zoete, V. SwissADME: A free web tool to evaluate pharmacokinetics, drug-likeness and medicinal chemistry friendliness of small molecules. Sci. Rep. 2017, 7, 42717. [Google Scholar] [CrossRef]
- Stelzer, G.; Rosen, N.; Plaschkes, I.; Zimmerman, S.; Twik, M.; Fishilevich, S.; Stein, T.I.; Nudel, R.; Lieder, I.; Mazor, Y.; et al. The GeneCards suite: From gene data mining to disease genome sequence analyses. Curr. Protoc. Bioinform. 2016, 54, 1.30.31–31.30.33. [Google Scholar] [CrossRef]
- Ge, S.X.; Jung, D.; Yao, R. ShinyGO: A graphical gene-set enrichment tool for animals and plants. Bioinformatics 2019, 36, 2628–2629. [Google Scholar] [CrossRef]
- Szklarczyk, D.; Kirsch, R.; Koutrouli, M.; Nastou, K.; Mehryary, F.; Hachilif, R.; Gable, A.L.; Fang, T.; Doncheva, N.T.; Pyysalo, S.; et al. The STRING database in 2023: Protein–protein association networks and functional enrichment analyses for any sequenced genome of interest. Nucleic Acids Res. 2022, 51, D638–D646. [Google Scholar] [CrossRef]
- Eberhardt, J.; Santos-Martins, D.; Tillack, A.F.; Forli, S. AutoDock Vina 1.2.0: New docking methods, expanded force field, and python bindings. J. Chem. Inf. Model. 2021, 61, 3891–3898. [Google Scholar] [CrossRef]
- Sama-ae, I.; Pattaranggoon, N.C.; Tedasen, A. In Silico prediction of antifungal compounds from natural sources towards lanosterol 14-alpha demethylase (CYP51) using molecular docking and molecular dynamic simulation. J. Mol. Graph. Model. 2023, 121, 108435. [Google Scholar] [CrossRef] [PubMed]
- Durrant, J.D.; McCammon, J.A. Molecular dynamics simulations and drug discovery. BMC Biol. 2011, 9, 71. [Google Scholar] [CrossRef] [PubMed]
- Childers, M.C.; Daggett, V. Validating molecular dynamics simulations against experimental observables in light of underlying conformational ensembles. J. Phys. Chem. B 2018, 122, 6673–6689. [Google Scholar] [CrossRef] [PubMed]
- Essmann, U.; Perera, L.; Berkowitz, M.L.; Darden, T.; Lee, H.; Pedersen, L.G. A smooth particle mesh Ewald method. J. Chem. Phys. 1995, 103, 8577–8593. [Google Scholar] [CrossRef]
- Palaiogiannis, D.; Chatzimitakos, T.; Athanasiadis, V.; Bozinou, E.; Makris, D.P.; Lalas, S.I. Successive solvent extraction of polyphenols and flavonoids from Cistus creticus L. leaves. Oxygen 2023, 3, 274–286. [Google Scholar] [CrossRef]
- Khan, S.; Jaafar, A.; Rukayadi, Y. Antibacterial activity of asam gelugur (Garcinia atroviridis Griff) fruit extract against foodborne pathogens and its effect on microbial population in raw chicken. Food Res. 2022, 6, 223–234. [Google Scholar] [CrossRef] [PubMed]
- Chailap, B.; Nuanyai, T. Effect of ethanol polarity and temperature on antioxidant activity, tyrosinase inhibition, and total phenol content of Garcinia atroviridis (Asam gelugor) fruits extract. Huachiew Chalermprakiet Sci. Technol. J. 2022, 8, 70–80. [Google Scholar]
- Jantan, I.; Jumuddin, F.A.; Saputri, F.; Rahman, K. Inhibitory effects of the extracts of Garcinia species on human low-density lipoprotein peroxidation and platelet aggregation in relation to their total phenolic contents. J. Med. Plants Res. 2011, 5, 2699–2709. [Google Scholar]
- Al-Mansoub, M.A.; Asmawi, M.Z.; Murugaiyah, V. Effect of extraction solvents and plant parts used on the antihyperlipidemic and antioxidant effects of Garcinia atroviridis: A comparative study. J. Sci. Food Agric. 2014, 94, 1552–1558. [Google Scholar] [CrossRef]
- Čižmárová, B.; Hubková, B.; Tomečková, V.; Birková, A. Flavonoids as promising natural compounds in the prevention and treatment of selected skin diseases. Int. J. Mol. Sci. 2023, 24, 6324. [Google Scholar] [CrossRef]
- Miean, K.H.; Mohamed, S. Flavonoid (myricetin, quercetin, kaempferol, luteolin, and apigenin) content of edible tropical plants. J. Agric. Food Chem. 2001, 49, 3106–3112. [Google Scholar] [CrossRef]
- Phuyal, N.; Jha, P.K.; Raturi, P.P.; Rajbhandary, S. Total Phenolic, flavonoid contents, and antioxidant activities of fruit, seed, and bark extracts of Zanthoxylum armatum DC. Sci. World J. 2020, 2020, 8780704. [Google Scholar] [CrossRef]
- Martínez, S.; Fuentes, C.; Carballo, J. Antioxidant activity, total phenolic content and total flavonoid content in sweet chestnut (Castanea sativa Mill.) cultivars grown in Northwest Spain under different environmental conditions. Foods 2022, 11, 3519. [Google Scholar] [CrossRef] [PubMed]
- Nantitanon, W.; Yotsawimonwat, S.; Okonogi, S. Factors influencing antioxidant activities and total phenolic content of guava leaf extract. LWT Food Sci. Technol. 2010, 43, 1095–1103. [Google Scholar] [CrossRef]
- Gulcin, İ.; Alwasel, S.H. DPPH radical scavenging assay. Processes 2023, 11, 2248. [Google Scholar] [CrossRef]
- Nahhas, A.F.; Abdel-Malek, Z.A.; Kohli, I.; Braunberger, T.L.; Lim, H.W.; Hamzavi, I.H. The potential role of antioxidants in mitigating skin hyperpigmentation resulting from ultraviolet and visible light-induced oxidative stress. Photodermatol. Photoimmunol. Photomed. 2019, 35, 420–428. [Google Scholar] [CrossRef] [PubMed]
- Ilyasov, I.R.; Beloborodov, V.L.; Selivanova, I.A.; Terekhov, R.P. ABTS/PP decolorization assay of antioxidant capacity reaction pathways. Int. J. Mol. Sci. 2020, 21, 1131. [Google Scholar] [CrossRef]
- El-Nashar, H.A.S.; El-Din, M.I.G.; Hritcu, L.; Eldahshan, O.A. Insights on the inhibitory power of flavonoids on tyrosinase activity: A Survey from 2016 to 2021. Molecules 2021, 26, 7546. [Google Scholar] [CrossRef]
- Działo, M.; Mierziak, J.; Korzun, U.; Preisner, M.; Szopa, J.; Kulma, A. The potential of plant phenolics in prevention and therapy of skin disorders. Int. J. Mol. Sci. 2016, 17, 160. [Google Scholar] [CrossRef]
- Guo, L.; Li, W.; Gu, Z.; Wang, L.; Guo, L.; Ma, S.; Li, C.; Sun, J.; Han, B.; Chang, J. Recent advances and progress on melanin: From source to application. Int. J. Mol. Sci. 2023, 24, 4360. [Google Scholar] [CrossRef]
- Kobayashi, T.; Urabe, K.; Winder, A.; Jiménez-Cervantes, C.; Imokawa, G.; Brewington, T.; Solano, F.; García-Borrón, J.C.; Hearing, V.J. Tyrosinase related protein 1 (TRP1) functions as a DHICA oxidase in melanin biosynthesis. EMBO J. 1994, 13, 5818–5825. [Google Scholar] [CrossRef] [PubMed]
- Dall’Olmo, L.; Papa, N.; Surdo, N.C.; Marigo, I.; Mocellin, S. Alpha-melanocyte stimulating hormone (α-MSH): Biology, clinical relevance and implication in melanoma. J. Transl. Med. 2023, 21, 562. [Google Scholar] [CrossRef] [PubMed]
- Nguyen, N.T.; Fisher, D.E. MITF and UV responses in skin: From pigmentation to addiction. Pigment. Cell Melanoma Res. 2019, 32, 224–236. [Google Scholar] [CrossRef] [PubMed]
- Seo, G.-Y.; Ha, Y.; Park, A.-H.; Kwon, O.W.; Kim, Y.-J. Leathesia difformis extract inhibits α-MSH-induced melanogenesis in B16F10 cells via down-regulation of CREB signaling pathway. Int. J. Mol. Sci. 2019, 20, 536. [Google Scholar] [CrossRef] [PubMed]
- Zhou, X.; Oh, J.H.; Karadeniz, F.; Yang, J.; Lee, H.; Seo, Y.; Kong, C.-S. Anti-melanogenesis effect of Rosa rugosa on α-MSH-induced B16F10 cells via PKA/CREB pathway activation. Appl. Sci. 2023, 13, 184. [Google Scholar] [CrossRef]
- Matoba, Y.; Bando, N.; Oda, K.; Noda, M.; Higashikawa, F.; Kumagai, T.; Sugiyama, M. A molecular mechanism for copper transportation to tyrosinase that is assisted by a metallochaperone, caddie protein. J. Biol. Chem. 2011, 286, 30219–30231. [Google Scholar] [CrossRef] [PubMed]
- Hamann, J.N.; Tuczek, F. New catalytic model systems of tyrosinase: Fine tuning of the reactivity with pyrazole-based N-donor ligands. Chem. Commun. 2014, 50, 2298–2300. [Google Scholar] [CrossRef] [PubMed]
- Stylianou, M.; Keramidas, A.D.; Drouza, C. pH-potentiometric investigation towards chelating tendencies of p-hydroquinone and phenol iminodiacetate copper(II) complexes. Bioinorg. Chem. Appl 2010, 2010, 125717. [Google Scholar] [CrossRef] [PubMed]
- Zolghadri, S.; Bahrami, A.; Hassan Khan, M.T.; Munoz-Munoz, J.; Garcia-Molina, F.; Garcia-Canovas, F.; Saboury, A.A. A comprehensive review on tyrosinase inhibitors. J. Enzyme Inhib. Med. Chem. 2019, 34, 279–309. [Google Scholar] [CrossRef]
- Noh, H.; Lee, S.J.; Jo, H.J.; Choi, H.W.; Hong, S.; Kong, K.H. Histidine residues at the copper-binding site in human tyrosinase are essential for its catalytic activities. J. Enzyme Inhib. Med. Chem. 2020, 35, 726–732. [Google Scholar] [CrossRef]
- Waterman, P.G.; Hussain, R.A. Systematic significance of xanthones, benzophenones and biflavonoids in Garcinia. Biochem. Syst. Ecol 1983, 11, 21–28. [Google Scholar] [CrossRef]
- Ali, S.; Goundar, R.; Sotheeswaran, S.; Beaulieu, C.; Spino, C. Benzophenones of Garcinia pseudoguttifera (Clusiaceae). Phytochem. 2000, 53, 281–284. [Google Scholar] [CrossRef]
- Baharuddin, F.F.; Mad Nasir, N.; Tejo, B.A.; Koh, S.P.; Ramakrishnan, S.; Nordin, N.; Adzahar, A.N.; Devakrishnan, P.; Mohd Razib, S. Potent halogenated xanthone derivatives: Synthesis, molecular docking and study on antityrosinase activity. J. Asian Nat. Prod. Res. 2023, 26, 575–582. [Google Scholar] [CrossRef]
- Huang, Y.L.; Chen, C.C.; Chen, Y.J.; Huang, R.L.; Shieh, B.J. Three xanthones and a benzophenone from Garcinia mangostana. J. Nat. Prod. 2001, 64, 903–906. [Google Scholar] [CrossRef]
- Khamthong, N.; Hutadilok-Towatana, N. Phytoconstituents and biological activities of Garcinia dulcis (Clusiaceae): A review. Nat. Prod. Commun. 2017, 12, 453–460. [Google Scholar] [CrossRef] [PubMed]
- Fuller, R.W.; Westergaard, C.K.; Collins, J.W.; Cardellina, J.H.; Boyd, M.R. Vismiaphenones D−G, new prenylated benzophenones from Vismia cayennensis. J. Nat. Prod. 1999, 62, 67–69. [Google Scholar] [CrossRef]
- Yun, Y.; Shioura, M.; Hitotsuyanagi, Y.; Yotsumoto, S.; Takahashi, Y.; Aoyagi, Y.; Kinoshita, T.; Takeya, K.; Inoue, H. Garcinielliptone G from Garcinia subelliptica induces apoptosis in acute leukemia cells. Molecules 2021, 26, 2422. [Google Scholar] [CrossRef] [PubMed]
- Hay, A.E.; Merza, J.; Landreau, A.; Litaudon, M.; Pagniez, F.; Le Pape, P.; Richomme, P. Antileishmanial polyphenols from Garcinia vieillardii. Fitoterapia 2008, 79, 42–46. [Google Scholar] [CrossRef]
- Liu, K.; Abdullah, A.A.; Huang, M.; Nishioka, T.; Altaf-Ul-Amin, M.; Kanaya, S. Novel approach to classify plants based on metabolite-content similarity. Biomed. Res. Int. 2017, 2017, 5296729. [Google Scholar] [CrossRef]
- Alam, M.B.; Ahmed, A.; Motin, M.A.; Kim, S.; Lee, S.-H. Attenuation of melanogenesis by Nymphaea nouchali (Burm. f) flower extract through the regulation of cAMP/CREB/MAPKs/MITF and proteasomal degradation of tyrosinase. Sci. Rep. 2018, 8, 13928. [Google Scholar] [CrossRef]
- Fu, C.; Chen, J.; Lu, J.; Yi, L.; Tong, X.; Kang, L.; Pei, S.; Ouyang, Y.; Jiang, L.; Ding, Y.; et al. Roles of inflammation factors in melanogenesis (Review). Mol. Med. Rep. 2020, 21, 1421–1430. [Google Scholar] [CrossRef] [PubMed]
- Hemesath, T.J.; Price, E.R.; Takemoto, C.; Badalian, T.; Fisher, D.E. MAP kinase links the transcription factor Microphthalmia to c-Kit signalling in melanocytes. Nature 1998, 391, 298–301. [Google Scholar] [CrossRef]
- Imokawa, G.; Yada, Y.; Kimura, M. Signalling mechanisms of endothelin-induced mitogenesis and melanogenesis in human melanocytes. Biochem. J. 1996, 314 Pt 1, e305–e312. [Google Scholar] [CrossRef] [PubMed]
- Imran, M.; Salehi, B.; Sharifi-Rad, J.; Aslam Gondal, T.; Saeed, F.; Imran, A.; Shahbaz, M.; Tsouh Fokou, P.V.; Umair Arshad, M.; Khan, H.; et al. Kaempferol: A key emphasis to its anticancer potential. Molecules 2019, 24, 2277. [Google Scholar] [CrossRef] [PubMed]
- Akter, M.; Parvin, M.S.; Hasan, M.M.; Rahman, M.A.A.; Islam, M.E. Anti-tumor and antioxidant activity of kaempferol-3-O-alpha-L-rhamnoside (Afzelin) isolated from Pithecellobium dulce leaves. BMC Complement. Med. Ther. 2022, 22, 169. [Google Scholar] [CrossRef] [PubMed]
- Buscà, R.; Ballotti, R. Cyclic AMP a key messenger in the regulation of skin pigmentation. Pigment Cell Res. 2000, 13, 60–69. [Google Scholar] [CrossRef] [PubMed]
- Lee, H.J.; Lee, W.J.; Chang, S.E.; Lee, G.-Y. Hesperidin, a popular antioxidant inhibits melanogenesis via Erk1/2 mediated MITF degradation. Int. J. Mol. Sci. 2015, 16, 18384–18395. [Google Scholar] [CrossRef]
- Yoon, Y.; Bae, S.; Kim, T.J.; An, S.; Lee, J.H. Nodakenin inhibits melanogenesis via the ERK/MSK1 signaling pathway. Pharmazie 2023, 78, 6–12. [Google Scholar]




{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| No. | RT (min) | Mass | Chemical Formula | Error (ppm) | Identification |
|---|---|---|---|---|---|
| Positive Mode of Analysis | |||||
| 1 | 1.707 | 598.3291 | C38H46O6 | −0.6 | Gambogin |
| 2 | 4.552 | 168.0422 | C8H8O4 | −0.49 | 2,6-Dimethoxy-p-benzoquinone |
| 3 | 7.745 | 484.3538 | C31H48O4 | −2.99 | Garcihombronane J |
| 4 | 14.752 | 196.0524 | C13H8O2 | −0.19 | Xanthone |
| 5 | 16.833 | 372.2166 | C19H32O7 | 4.86 | Blumenol-C-O-β-D-glucoside |
| 6 | 19.329 | 408.1577 | C24H24O6 | 0.98 | Garcimangosone B |
| 7 | 19.818 | 442.1618 | C24H26O6 | −2.21 | Mangostenone C |
| 8 | 22.106 | 300.1365 | C18H20O4 | 1.10 | (S)-3-hydroxygarcibenzopyran |
| Negative Mode of Analysis | |||||
| 1 | 2.319 | 192.0267 | C6H8O7 | −1.32 | Citric acid |
| 2 | 5.051 | 190.0109 | C6H6O7 | −2.18 | Garcinia lactone |
| 3 | 5.733 | 352.0951 | C20H16O6 | 1.06 | Subelliptenone C |
| 4 | 12.335 | 132.0422 | C5H8O4 | −0.71 | 2-Acetoxypropanoic acid |
| 5 | 15.068 | 164.0472 | C9H8O3 | −0.96 | p-Coumaric acid |
| 6 | 15.524 | 432.1071 | C21H20O10 | 3.43 | Kaempferol-3-O-a-L-rhamnoside |
| 7 | 17.401 | 182.0575 | C9H10O4 | −1.99 | Methyl orsellinate |
| 8 | 19.109 | 398.1745 | C23H26O6 | 3.95 | Nigrolineaxanthone L |
| 9 | 19.166 | 420.2287 | C27H32O4 | −3.29 | Clusiachromene D |
| 10 | 19.848 | 362.1717 | C20H26O6 | −3.29 | Parvifoliol B |
| 11 | 20.133 | 384.1209 | C21H20O7 | 0.12 | Dulxanthone F |
| 12 | 20.759 | 250.1941 | C16H26O2 | 3.32 | Scortechterpene B |
| 13 | 21.329 | 636.3679 | C38H52O8 | 2.65 | Garcimultiflorone F |
| 14 | 21.955 | 576.4400 | C35H60O6 | 1.69 | β-Sitosterol-3-O-β-D-glucoside |
| 15 | 22.012 | 380.1634 | C23H24O5 | 2.78 | Vismiaphenone E |
| 16 | 22.012 | 426.1691 | C24H26O7 | 2.90 | Parvixanthones H |
| 17 | 22.126 | 428.1845 | C24H28O7 | 2.22 | Garcinone D |
| 18 | 22.581 | 502.3085 | C33H42O4 | 0.33 | Kolanone |
| 19 | 22.582 | 400.3348 | C27H44O2 | 1.63 | Garcinielliptone |
| 20 | 23.150 | 410.1747 | C24H26O6 | 4.40 | Mangostin |
| 21 | 23.150 | 364.1685 | C23H24O4 | 2.92 | Clusiachromene C |
| 22 | 24.857 | 586.3672 | C38H50O5 | 2.28 | Guttiferone J |
| 23 | 25.429 | 394.1797 | C24H26O5 | 4.31 | Cudraxanthone G |
| 24 | 26.453 | 574.4245 | C35H58O6 | 1.97 | Stigmasta-5,22-dien-3-O-β-glucopyranoside |
| 25 | 26.909 | 570.3724 | C38H50O4 | 2.55 | 13,14-Didehydroxyisogarcinol |
| Identification | Pubchem CID | Physicochemical Properties | Drug-Likeness | ||||
|---|---|---|---|---|---|---|---|
| MW (g/mol) | HBA | HBD | TPSA (Å2) | logP | |||
| Gambogin | 15298998 | 598.77 | 6 | 1 | 82.06 | 7.04 | No |
| 2,6-dimethoxy-p-benzoquinone | 68262 | 168.15 | 4 | 0 | 52.6 | 0.22 | Yes |
| Garcihombronane J | 11670274 | 484.71 | 4 | 1 | 63.6 | 5.69 | Yes |
| Xanthone | 7020 | 196.2 | 2 | 0 | 30.21 | 2.84 | Yes |
| Blumenol-C-O-β-D-glucoside | 14135394 | 372.45 | 7 | 4 | 116.45 | 0.91 | Yes |
| Garcimangosone B | 11143989 | 408.44 | 6 | 1 | 78.13 | 4.33 | Yes |
| Mangostenone C | 11546716 | 442.46 | 8 | 4 | 129.59 | 3.13 | Yes |
| (S)-3-hydroxygarcibenzopyran | - | 300.35 | 4 | 2 | 58.92 | 2.94 | Yes |
| Citric acid | 311 | 192.12 | 7 | 4 | 132.13 | −1.51 | Yes |
| Garcinia lactone | 9991606 | 190.11 | 7 | 3 | 121.13 | −1.34 | Yes |
| Subelliptenone C | 101681084 | 352.34 | 6 | 3 | 104.04 | 3.51 | Yes |
| 2-acetoxypropanoic acid | 79041 | 132.11 | 4 | 1 | 63.6 | 0.06 | Yes |
| p-Coumaric acid | 637542 | 164.16 | 3 | 2 | 57.53 | 1.26 | Yes |
| Kaempferol-3-O-a-L-rhamnoside | 5835713 | 432.38 | 10 | 6 | 170.05 | 0.6 | Yes |
| Methyl orsellinate | 76658 | 182.17 | 4 | 2 | 66.76 | 1.4 | Yes |
| Nigrolineaxanthone L | 11749894 | 398.45 | 6 | 3 | 100.13 | 3.8 | Yes |
| Clusiachromene D | 21602027 | 420.54 | 4 | 3 | 69.92 | 5.73 | Yes |
| Parvifoliol B | 163184032 | 306.35 | 5 | 4 | 97.99 | 3.39 | Yes |
| Dulxanthone F | 10500218 | 384.38 | 7 | 1 | 87.36 | 3.34 | Yes |
| Scortechterpene B | 101746891 | 236.35 | 2 | 0 | 26.3 | 2.96 | Yes |
| Garcimultiflorone F | 46919309 | 636.81 | 8 | 5 | 152.36 | 5.88 | No |
| β-sitosterol-3-O-β-D-glucoside | 12309057 | 576.85 | 6 | 4 | 99.38 | 5.51 | No |
| Vismiaphenone E | 474315 | 380.43 | 5 | 3 | 86.99 | 4.02 | Yes |
| Parvixanthones H | 11080433 | 426.46 | 7 | 4 | 120.36 | 3.86 | Yes |
| Garcinone D | 5495926 | 428.47 | 7 | 4 | 120.36 | 3.91 | Yes |
| Kolanone | 6439598 | 502.68 | 4 | 2 | 74.6 | 7.01 | No |
| Garcinielliptone | - | 400.64 | 2 | 1 | 37.3 | 5.7 | Yes |
| Mangostin | 5281650 | 410.46 | 6 | 3 | 100.13 | 4.64 | Yes |
| Clusiachromene C | 16070714 | 364.43 | 4 | 2 | 66.76 | 4.43 | Yes |
| Guttiferone J | 102031302 | 586.8 | 5 | 2 | 91.67 | 7.8 | No |
| Cudraxanthone G | 42645953 | 394.46 | 5 | 2 | 79.9 | 5.02 | Yes |
| Stigmasta-5,22-dien-3-O-β-glucopyranoside | 6602508 | 574.83 | 6 | 4 | 99.38 | 5.1 | No |
| 13,14-didehydroxyisogarcinol | 25243255 | 570.8 | 4 | 0 | 60.44 | 7.74 | No |
| Identification | Pharmacokinetic Properties | |||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| GI Absorption | BBB Permeant | P-gp Substrate | CYP1A2 Inhibitor | CYP2C19 Inhibitor | CYP2C9 Inhibitor | CYP2D6 Inhibitor | CYP3A4 Inhibitor | Log Kp (cm/s) | AMES Toxicity | hERG I Inhibitor | hERG II Inhibitor | Hepato-Toxicity | Skin Sensitisation | |
| Gambogin | Low | No | Yes | No | No | No | No | Yes | −3.82 | No | No | No | No | No |
| 2,6-Dimethoxy-p-benzoquinone | High | No | No | No | No | No | No | No | −7.37 | No | No | No | No | Yes |
| Garcihombronane J | High | No | No | No | No | No | No | Yes | −5.17 | No | No | No | No | No |
| Xanthone | High | Yes | No | Yes | No | No | No | No | −5.09 | Yes | No | No | No | Yes |
| Blumenol-C-O-β-D-glucoside | High | No | Yes | No | No | No | No | No | −8.36 | No | No | No | No | No |
| Garcimangosone B | High | No | Yes | No | Yes | Yes | No | No | −5.22 | No | No | Yes | No | No |
| Mangostenone C | High | No | No | No | No | Yes | No | No | −6.05 | Yes | No | Yes | No | No |
| (S)-3-hydroxygarcibenzopyran | High | Yes | Yes | Yes | No | No | Yes | Yes | −5.88 | Yes | No | Yes | No | No |
| Citric acid | Low | No | No | No | No | No | No | No | −8.69 | No | No | No | No | No |
| Garcinia lactone | Low | No | No | No | No | No | No | No | −8.33 | No | No | No | No | No |
| Subelliptenone C | High | No | No | Yes | No | Yes | No | No | −5.04 | No | No | Yes | No | No |
| 2-Acetoxypropanoic acid | High | No | No | No | No | No | No | No | −7.03 | No | No | No | No | No |
| p-Coumaric acid | High | Yes | No | No | No | No | No | No | −6.26 | No | No | No | No | No |
| Kaempferol-3-O-a-L-rhamnoside | Low | No | No | No | No | No | No | No | −8.07 | No | No | Yes | No | No |
| Methyl orsellinate | High | Yes | No | No | No | No | No | No | −6.01 | Yes | No | No | No | No |
| Nigrolineaxanthone L | High | No | No | No | No | Yes | No | No | −5.66 | Yes | No | Yes | Yes | No |
| Clusiachromene D | High | No | No | No | Yes | No | No | No | −3.71 | No | No | Yes | No | No |
| Parvifoliol B | High | No | No | No | No | Yes | No | No | −4.43 | No | No | No | No | No |
| Dulxanthone F | High | No | No | No | Yes | Yes | No | Yes | −5.8 | Yes | No | Yes | No | No |
| Scortechterpene B | High | Yes | No | No | No | No | No | No | −5.64 | No | No | No | No | Yes |
| Garcimultiflorone F | Low | No | Yes | No | No | No | No | Yes | −4.5 | No | No | No | Yes | No |
| β-Sitosterol-3-O-β-D-glucoside | Low | No | No | No | No | No | No | No | −4.32 | No | No | No | No | No |
| Vismiaphenone E | High | No | No | Yes | No | Yes | No | Yes | −4.64 | No | No | Yes | No | No |
| Parvixanthones H | High | No | No | No | No | Yes | No | Yes | −5.24 | Yes | No | Yes | No | No |
| Garcinone D | High | No | No | No | No | No | No | No | −5.43 | No | No | Yes | No | No |
| Kolanone | Low | No | Yes | No | Yes | No | No | Yes | −2.81 | No | No | No | No | No |
| Garcinielliptone | High | No | No | No | No | No | No | No | −3.72 | No | No | No | No | No |
| Mangostin | High | No | No | No | No | Yes | No | No | −4.35 | Yes | No | Yes | No | No |
| Clusiachromene C | High | No | No | Yes | Yes | Yes | No | Yes | −4.28 | No | No | Yes | No | No |
| Guttiferone J | Low | No | Yes | No | No | No | No | Yes | −2.26 | No | No | No | No | No |
| Cudraxanthone G | High | No | No | No | Yes | Yes | No | No | −4 | Yes | No | Yes | No | No |
| Stigmasta-5,22-dien-3-O-β-glucopyranoside | High | No | Yes | No | No | No | No | No | −4.86 | No | No | No | No | No |
| 13,14-Didehydroxyisogarcinol | Low | No | Yes | No | No | No | No | No | −2.96 | No | No | No | No | No |
| Ligands | Docking Scores (kcal/mol) | |||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| TNF | MAPK3 | PTGS2 | EGFR | SRC | PPARG | CTNNB1 | ERBB2 | MDM2 | MAPK1 | MIFT | TYR | TYRP1 | MC1R | |
| 2,6-Dimethoxy-p-benzoquinone | −5.0 | −5.5 | −5.4 | −5.4 | −4.8 | −5.1 | −4.8 | −5.2 | −3.8 | −5.0 | −4.5 | −5.0 | −3.8 | −4.4 |
| Garcihombronane J | −7.8 | −7.1 | −7.1 | −7.6 | −7.0 | −6.6 | −5.8 | −7.3 | −7.3 | −5.7 | −6.1 | −7.3 | −4.6 | −5.6 |
| Blumenol-C-O-β-D-glucoside | −7.6 | −8.4 | −7.1 | −7.4 | −7.7 | −6.6 | −6.7 | −6.5 | −7.0 | −7.3 | −6.5 | −7.1 | −4.9 | −6.1 |
| Garcimangosone B | −8.5 | −9.8 | −9.3 | −10.3 | −9.8 | −8.6 | −7.8 | −7.7 | −7.9 | −8.4 | −7.8 | −7.6 | −6.5 | −7.7 |
| Citric acid | −4.7 | −4.8 | −5.0 | −5.7 | −5.0 | −5.1 | −5.1 | −5.5 | −3.8 | −4.7 | −4.5 | −5.6 | −4.4 | −4.5 |
| Garcinia lactone | −5.4 | −5.7 | −5.8 | −5.7 | −5.8 | −4.9 | −6.0 | −5.8 | −4.5 | −5.2 | −5.1 | −5.7 | −5.8 | −4.5 |
| Subelliptenone C | −8.6 | −9.5 | −10.9 | −9.1 | −8.9 | −9.1 | −7.5 | −8.7 | −7.2 | −8.6 | −7.4 | −7.4 | −7.0 | −7.4 |
| 2-Acetoxypropanoic acid | −4.2 | −4.7 | −4.8 | −5.1 | −4.6 | −4.5 | −4.4 | −5.3 | −3.0 | −4.4 | −4.0 | −5.0 | −4.2 | −4.0 |
| p-Coumaric acid | −5.7 | −6.6 | −5.2 | −5.9 | −5.7 | −5.5 | −5.0 | −5.8 | −4.9 | −5.5 | −5.1 | −5.3 | −6.6 | −5.0 |
| Kaempferol-3-O-a-L-rhamnoside | −7.5 | −10.4 | −7.7 | −8.3 | −7.8 | −6.4 | −6.7 | −7.5 | −6.4 | −9.0 | −7.4 | −6.5 | −5.6 | −7.2 |
| Clusiachromene D | −8.6 | −7.5 | −7.6 | −8.0 | −6.8 | −6.4 | −7.2 | −7.6 | −7.9 | −5.9 | −6.6 | −6.5 | −5.8 | −8.1 |
| Parvifoliol B | −6.6 | −8.2 | −7.3 | −7.0 | −6.7 | −6.1 | −5.5 | −8.5 | −6.6 | −7.2 | −4.7 | −7.0 | −5.4 | −5.5 |
| Scortechterpene B | −6.7 | −6.9 | −5.6 | −7.4 | −7.1 | −5.6 | −5.4 | −6.5 | −6.4 | −6.2 | −4.8 | −5.8 | −4.5 | −5.5 |
| Vismiaphenone E | −8.1 | −8.7 | −7.1 | −9.3 | −7.9 | −7.5 | −7.1 | −8.0 | −7.9 | −7.3 | −7.2 | −8.2 | −6.6 | −7.6 |
| Garcinone D | −7.4 | −8.0 | −8.5 | −8.2 | −7.6 | −5.9 | −6.0 | −6.6 | −6.5 | −7.4 | −6.1 | −6.2 | −5.6 | −5.7 |
| Garcinielliptone | −8.8 | −8.4 | −9.2 | −9.3 | −8.5 | −7.6 | −6.9 | −8.8 | −9.3 | −7.1 | −8.1 | −7.7 | −6.3 | −8.4 |
| Clusiachromene C | −8.7 | −9.1 | −8.3 | −9.7 | −8.0 | −8.2 | −6.1 | −7.5 | −7.5 | −8.3 | −5.9 | −6.5 | −5.8 | −8.5 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Tedasen, A.; Chiabchalard, A.; Tencomnao, T.; Yamasaki, K.; Majima, H.J.; Phongphithakchai, A.; Chatatikun, M. Anti-Melanogenic Activity of Ethanolic Extract from Garcinia atroviridis Fruits Using In Vitro Experiments, Network Pharmacology, Molecular Docking, and Molecular Dynamics Simulation. Antioxidants 2024, 13, 713. https://doi.org/10.3390/antiox13060713
Tedasen A, Chiabchalard A, Tencomnao T, Yamasaki K, Majima HJ, Phongphithakchai A, Chatatikun M. Anti-Melanogenic Activity of Ethanolic Extract from Garcinia atroviridis Fruits Using In Vitro Experiments, Network Pharmacology, Molecular Docking, and Molecular Dynamics Simulation. Antioxidants. 2024; 13(6):713. https://doi.org/10.3390/antiox13060713
Chicago/Turabian StyleTedasen, Aman, Anchalee Chiabchalard, Tewin Tencomnao, Kenshi Yamasaki, Hideyuki J. Majima, Atthaphong Phongphithakchai, and Moragot Chatatikun. 2024. "Anti-Melanogenic Activity of Ethanolic Extract from Garcinia atroviridis Fruits Using In Vitro Experiments, Network Pharmacology, Molecular Docking, and Molecular Dynamics Simulation" Antioxidants 13, no. 6: 713. https://doi.org/10.3390/antiox13060713
APA StyleTedasen, A., Chiabchalard, A., Tencomnao, T., Yamasaki, K., Majima, H. J., Phongphithakchai, A., & Chatatikun, M. (2024). Anti-Melanogenic Activity of Ethanolic Extract from Garcinia atroviridis Fruits Using In Vitro Experiments, Network Pharmacology, Molecular Docking, and Molecular Dynamics Simulation. Antioxidants, 13(6), 713. https://doi.org/10.3390/antiox13060713