


Telomerase Inhibition in the Treatment of Leukemia: A Comprehensive Review

Abstract
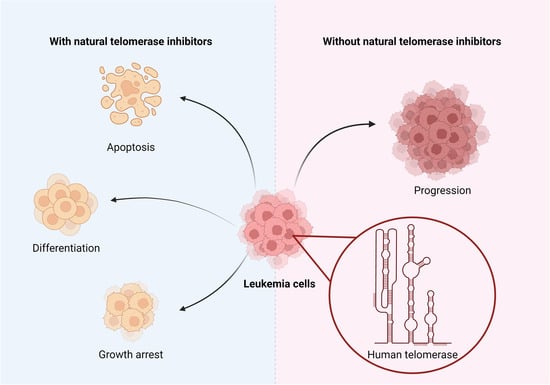
1. Introduction
2. Materials and Methods
3. Natural Substances Inhibiting Telomerase
3.1. Polyphenols and Flavones
3.1.1. Polyphenols
3.1.2. Flavones and Flavonolignans
3.2. Vitamins
3.2.1. Vitamin A
3.2.2. Vitamin D
3.2.3. Vitamin E
3.2.4. Vitamin C
3.3. Fatty Acids and Their Esters
3.4. Substances Used in Traditional Chinese Medicine
3.5. Other Substances
3.5.1. Alkaloids
3.5.2. Lactones
3.5.3. Quinones
3.5.4. Sapogenins
3.5.5. Saponins
3.5.6. Substances Containing Sulfur
3.5.7. Xanthenes
3.6. Substances Produced by Animals
3.6.1. Substances Produced by Organisms from Marine Ecosystems
3.6.2. Substances Produced by Bacteria
3.7. Substances Produced by Fungi

| Substance | Natural Occurrence | Cell Line | Dose Applied | Durance | IC50 or ED50 | |
|---|---|---|---|---|---|---|
| Alkaloids | ||||||
| Camptothecin | Camptotheca acuminata | HL-60 | 1 mg/L | 2, 4, 6 h | not specified | [113] |
| Homoharringtonine | Chinese evergreen Cephalotaxus harringtonia | HL-60 | 5, 10, 50, 100, 500 μg/L | 3, 6, 12, 24, 48 h | not specified | [114] |
| Bacteria | ||||||
| Telomestatin | Streptomyces anulatus | U937 | 1, 2, 5, 10 μM | 48 h, 0–60 days | not specified | [153] |
| Fatty acids and their esters | ||||||
| 23-Hydroxybetulinic acid | Pulsatilla chinensis | HL-60 | 1–1000 μM | 3, 6, 12, 24 h | not specified | [79] |
| HL-60, HL-60A | 0.5, 1, 2, 4, 8 mM | 24, 48, 72 h | 2.83, 3.90 mM | [77] | ||
| Valproic acid | Valeriana officinalis | 18 + 114 patients of AML | 400 mg/day + from day 3, 60–150 mg/L | 2 × 21 days, 3 × 21 days | not specified | [86] |
| 31 cases of elderly AML/RAEB | 5 mg/kg daily | 2–23 months | not specified | [87] | ||
| Ω-3 fatty acids (DHA, EPA) | Seafood, especially fatty fish, nuts, seeds, plant oils | 16 cases of Rai Stage 0–1 CLL | 1. month: 3 × 1250 mg/day 2. month: 6 × 1250 mg/day 3.–12. month: 9 × 1250 mg/day | 12 months | not specified | [82] |
| 14 cases of AML | 100 mL/day | 2 years | not specified | [84] | ||
| 60 cases of cALL (30 study/30 control) | 1000 mg/day | 6 months | not specified | [83] | ||
| Caffeic acid undecyl ester | Daphne oleoides | NALM-6 | 0.1, 0.3, 0.6, 1, 3 μM | 4, 18 h | not specified | [91] |
| NALM-6 | 0.1, 0.3, 0.6, 1 μM | 6, 12, 24, 72 h | 0.33, 0.16 μM | [90] | ||
| Trichostatin A | Culture broth of Streptomyces platensis | U-937 | 15, 30, 45, 60, 75 nM | 48 h | not specified | [88] |
| Fungi | ||||||
| Cordycepin | Cordyceps militaris | THP-1, H937 | 10, 20, 30 μM | 24 h | not specified | [155] |
| FA-2-b-ß fraction (an RNA-protein complex) | Agaricus blazei Murill | HL-60 | 5, 10, 20, 40, 80 μg/mL | 24, 48, 72, 96 h | 211, 187.35, 89.61, 42.72 µg/mL | [159] |
| Lactones | ||||||
| Constunolide | Stem bark of Magnolia sieboldii | NALM-6 | 2, 6, 8, 10 μM | 1, 2, 4, 6 h | 3.31 ± 0.32 mM | [115] |
| Withaferin-A | Withania somnifera | U937 | 0.25, 0.5, 1, 2 μM | 12, 24 h | not specified | [116] |
| Polyphenols | ||||||
| Curcumin | Curcuma longa | HL-60 | 1, 10, 50 μM | 24 h | 9.8 μM | [23] |
| HL-60 | 10, 15, 20, 40 μM | 24 and 48 h | not specified | [24] | ||
| K-562 | 1, 10, 50 μM | 6, 16, 24, 48 h | 14 μM | [25] | ||
| Epigallocatechin-3-gallate (EGCG) | Camellia sinensis | HL-60 | 50 μM | 3, 6, 9, 12 days | not specified | [26] |
| Jurkat | 30, 50, 70, 85, 100 μM | 24, 48, 72 h | not specified | [27] | ||
| Indole-3-carbinol | Brassica oleracea | NALM-6 | 20, 30, 40, 50, 60 μM | 24, 48 h | not specified | [29] |
| K-562 | 100, 200, 400 μM | 24, 48 h | not specified | [30] | ||
| Butein | Stem bark of Anacardium occidentale, the heartwood of Dalbergia odonifea, Rhus verniciflua, and Caragena jubata | U937, THP-1, HL60, K562 | 5, 10, 15, 20, 30 μM | 24 h | not specified | [31] |
| Genistein | Fabaceae | HL-60 | 50 μM | 24, 48, 72 h | 50 μM | [17] |
| Apigenin | Lamiaceae, Allium cepa, Petroselinum crispum, wine, tea, beer | HL-60, U937, THP-1 | 25, 50, 75, 100 μM | 24 h | not specified | [33] |
| Silymarin | Silybum marianum | K562 | 10, 25, 50, 75, 100 μg/mL | 24, 48, 72 h | not specified | [35] |
| Baicalin | Scutellaria baicalensis | HL-60 | HL-60 5, 10, 20, 40, 80 μg/mL | 12, 24, 48 h | 21.8 μg/mL | [36] |
| Quinones | ||||||
| β-Lapachone | Handroanthus impetiginosus | U937, K562, HL60, THP-1 | 1, 2, 3, 4 μM | 24 h | not specified | [119] |
| Salvicine | Salvia prionitis (modified) | HL-60 | 2.5, 5, 10, 20 μM | 2, 4, 6 h | not specified | [120] |
| Sapogenin | ||||||
| Diosgenin | Trigonella foenum-graecum | K562 | 5, 10, 15, 20, 30 μM | 48 h | not specified | [123] |
| Saponins | ||||||
| Ginsenoside Rg1 | Panax ginseng | CD34+CD38-LSCs | 20, 40, 80 μM | 24, 48, 72 h | not specified | [124] |
| U937 | 0.2, 0.4, 0.6, 0.8, 1, 1.2, 1.4, 1.6, 0.8, 2 mg/mL | 24, 48, 72 h | not specified | [125] | ||
| Patensin | Pulsatilla patens var. multifida | HL-60 | 0.001, 0.01, 0.1, 1, 10 mmol/L | 3, 6, 12, 24 h | not specified | [130] |
| Platycodin D | Platycodon grandiflorus | U937, THP-1, K562 | 5, 10, 15, 20 μM | 48 h | not specified | [132] |
| Substances produced by animals | ||||||
| D-galactan sulfate | Gymnodinium sp. A3 | K562 | 0.003, 0.01, 0.03, 0.1, 0.3, 1 μg/mL | 15, 45 min | not specified | [150] |
| Dideoxypetrosynol A | Marine sponge Petrosia sp. | U-937 | 0.2, 0.4, 0.6, 0.8, 1 μg/mL | 48 h | not specified | [148] |
| Interferon alpha | Synthesized in the human organism by plasmacytoid dendritic cells | 42 study/42 control of AML | 3 × week 3 mln IU | 12–18 months | not specified | [144] |
| 36 | −1. before HCT, 14., 28., 42. (+/−7) days 45, 90, 180 μg | 24 months | not specified | [145] | ||
| Manisa propolis | Substance collected by honeybees | 4 childhood leukemia cases (3 ALL, 1 CML) | 15, 30, 60 ng/mL | 24, 48, 72 h | not specified | [146] |
| CCFR-CEM | 0.03 μg/mL | 24, 48, 72, 96 h | 0.03 μg/mL | [147] | ||
| Melatonin | Synthesized in a human pineal gland, eggs, fish, nuts | RS4-11 (MLL-AF4+ B-ALL), MOLM-13 (MLL-AF9+ AML), Nalm-6 (non MLL-r B-ALL) | 1, 2, 3 mM | 24, 48, 72 h | 1.523, 0.957, 1.869, 0.728, 0.748, 1.877 mM | [139] |
| Pectenotoxin-2 | Dinophysis species | U937, THP-1, HL-60 | 2, 4, 6, 8, 10 ng/mL | 72 h | not specified | [141] |
| (Z)-stellettic acid C | Stelletta sp. sponges | U-937 | 5, 10, 20, 30, 40 μg/mL | 48 h | not specified | [149] |
| Substances containing sulfur | ||||||
| SQDG | Azadirachta indica | MOLT-4 | 5, 10, 15, 20, 25 μM | 2, 4, 8, 12, 16, 20, 24, 48, 72 h | 15.32 ± 0.58, 22.52 ± 0.64, 19.63 ± 0.23, 75.67 ± 6.4 μM | [134] |
| Sulforaphane | Cruciferous vegetables | HL-60 | 6, 7, 8, 9, 10 μM | 24, 48 h | not specified | [136] |
| Traditional Chinese medicine | ||||||
| Angelica sinensis polysaccharide | Angelica sinensis | K562, CD34+CD38− | 20, 40, 80 μg/mL | 48 h | not specified | [101] |
| Tanshinone I | Salvia miltiorrhiza | U937, THP-1, SHI 1 | 10, 20, 30, 40, 50 μM | 24, 48, 72 h | not specified | [97] |
| Tanshinone IIA | Salvia miltiorrhiza | HL-60, K562 | 0.5 μg/mL | 5, 6 days | not specified | [95] |
| Tianshengyuan-1 | A mixture of various Chinese herbs | HL-60, PBMCs, CD34+ HSCs | 31.2, 62.5 μg/mL | 24 h | not specified | [99] |
| Triptonide | Tripterygium wilfordii Hook F | U937, HL-60 | 0–10, 15, 20 nM 2, 4 mg/kg | 72 h, 3, 6, 9, 14, 21 days | 7.5, 12 nM | [107] |
| Wogonin | Scutellaria baicalensis | HL-60 | 0.5, 1, 2, 3 mg/mL 10, 25, 50, 75, 100 μM | 24 h | not specified | [110] |
| Vitamins | ||||||
| Ascorbic acid | Citrus fruits, green and leafy vegetables | CD34+, HL-60, U937 | 8, 20 mM | 24 h | not specified | [73] |
| Retinoic acid | Yellow, green, red, and leafy vegetables, yellow fruits | HL-60 | 1 μM | 12 days | not specified | [40] |
| 242 cases of APL | 60 mg/kg/day | 2 years | not specified | [160] | ||
| 124 cases of cAPL | 25 mg/m2/day | 5 years | not specified | [161] | ||
| Eμ-TCL1 mice | 1 μM | 24, 48 h | not specified | [47] | ||
| 9cUAB30 | Novel retinoid | HL-60d | 5 μM | 12 days | not specified | [162] |
| Growth factor binding protein 7 | Human insulin recombinant | HL60, KG1a, THP1, HEK293T | 100 μg/mL, 300 μg/mL, 10 mg/kg, 12 mg/kg | 48, 72, 120 h, 7, 14 days, 16 weeks | not specified | [42] |
| Vitamin D | Produced in the human body form 7-dehydrocholesterol; fish oil, egg yolk | HL-60 | 10−8 M | 3 days | not specified | [163] |
| U937 | 10−9–10−7 M | 48, 72 h | not specified | [164] | ||
| 26 cases of AML | 1 μg/day | 2 × 5 weeks | not specified | [165] | ||
| 17 cases of elderly AML | 100,000 IU/week | 6 months | not specified | [166] | ||
| EB 1089 | Vitamin D3 analog | HL-60, U937 | 5 × 10−10 M | 96 h | not specified | [54] |
| 1,25(OH)2-16-ene-5,6-trans-D3 | Vitamin D3 analog | HL-60 | 10−7 M | 4 days | 1.9 × 10−12 M | [55] |
| Vitamin E | Vegetable oil, seeds, nuts, grains | U937, KG-1 | 10–50 μM | 24 h | 29.43, 25.23 µM | [64] |
| K562 | 100 μM | 48 h | not specified | [167] | ||
| 25 cases of AML | 400 IU/day | 30 days | not specified | [66] | ||
| 2396 study/2235 control | 600 IU/day | 10 years | not specified | [68] | ||
| Xanthene | ||||||
| Crocin | Crocus sativus | Jurkat | 0.625, 1.25, 2.5, 5, 10 mg/mL | 12, 24, 26, 48 h | not specified | [138] |
| Group | Substance | Structure | Properties |
|---|---|---|---|
| Alkaloids | Camptothecin |  |
|
| Homoharringtonine |  |
| |
| Bacteria | Telomestatin |  |
|
| Fatty acids and their esters | 23-Hydroxybetulinic acid |  |
|
| Caffeic acid undecyl ester |  |
| |
| Docosahexaenoic acid |  |
| |
| Eicosapentaenoic acid |  | ||
| Oleic acid |  |
| |
| Sodium butyrate |  |
| |
| Trichostatin A |  |
| |
| Valproic acid |  |
| |
| Flavones | Apigenin |  |
|
| Baicalin |  |
| |
| Genistein |  |
| |
| Flavonolignans | Silymarin |  |
|
| Fungi | Cordycepin |  |
|
| FA-2-b-ß fraction (an RNA-protein complex) |
| ||
| Lactones | Constunolide |  |
|
| Withaferin-A |  |
| |
| Polyphenols | Butein |  |
|
| Curcumin |  |
| |
| Epigallocatechin-3-gallate |  |
| |
| Indole-3-carbinol |  |
| |
| Quinones | β-Lapachone |  |
|
| Salvicine |  |
| |
| Sapogenins | Diosgenin |  |
|
| Saponins | Ginsenoside Rg1 |  |
|
| Patensin |
| ||
| Platycodin D |  |
| |
| Substances produced by animals | D-galactan sulfate |
| |
| Dideoxypetrosynol A |  |
| |
| Interferon alpha |  |
| |
| Manisa propolis |
| ||
| Melatonin |  |
| |
| Pectenotoxin-2 |  |
| |
| (Z)-stellettic acid C |  |
| |
| Substances containing sulfur | SQDG |  |
|
| Sulforaphane |  |
| |
| Substances used in traditional Chinese medicine | Angelica sinensis polysaccharide |
| |
| Tanshinone I |  |
| |
| Tanshinone IIA |  |
| |
| Tianshengyuan-1 |
| ||
| Triptonide |  |
| |
| Wogonin |  |
| |
| Vitamins | Ascorbic acid |  |
|
| Calcitriol |  |
| |
| Retinoic acid |  |
| |
| Tocotrienol |  |
| |
| Xanthenes | Crocin |  |
|
4. Conclusions
- Their ambiguous effect on chemoprevention;
- The lack of data indicating the optimal and toxic doses;
- The lack of data regarding their potential side effects;
- The lack of data evaluating their pharmacodynamic properties.
Author Contributions
Funding
Conflicts of Interest
References
- Pandit, S.; Wasekar, N.; Badarkhe, G.; Yasam, V.R.; Nagarkar, R. Acute Lymphoblastic Leukemia to Acute Myeloid Leukemia: An Unusual Case Report of Lineage Switching. Hematol. Transfus. Cell Ther. 2022, 44, 112. [Google Scholar] [CrossRef] [PubMed]
- Da Mota, T.H.A.; Camargo, R.; Biojone, E.R.; Guimarães, A.F.R.; Pittella-Silva, F.; de Oliveira, D.M. The Relevance of Telomerase and Telomere-Associated Proteins in B-Acute Lymphoblastic Leukemia. Genes 2023, 14, 691. [Google Scholar] [CrossRef] [PubMed]
- Li, X.; Yao, Y.; Wu, F.; Song, Y. A Proteolysis-Targeting Chimera Molecule Selectively Degrades ENL and Inhibits Malignant Gene Expression and Tumor Growth. J. Hematol. Oncol. 2022, 15, 41. [Google Scholar] [CrossRef] [PubMed]
- Li, X.; Song, Y. Structure, Function and Inhibition of Critical Protein-Protein Interactions Involving Mixed Lineage Leukemia 1 and Its Fusion Oncoproteins. J. Hematol. Oncol. 2021, 14, 56. [Google Scholar] [CrossRef]
- Wu, F.; Nie, S.; Yao, Y.; Huo, T.; Li, X.; Wu, X.; Zhao, J.; Lin, Y.L.; Zhang, Y.; Mo, Q.; et al. Small-Molecule Inhibitor of AF9/ENL-DOT1L/AF4/AFF4 Interactions Suppresses Malignant Gene Expression and Tumor Growth. Theranostics 2021, 11, 8172–8184. [Google Scholar] [CrossRef] [PubMed]
- Shen, Z.; Wang, Y.; Wang, G.; Gu, W.; Zhao, S.; Hu, X.; Liu, W.; Cai, Y.; Ma, Z.; Gautam, R.K.; et al. Research Progress of Small-Molecule Drugs in Targeting Telomerase in Human Cancer and Aging. Chem. Biol. Interact. 2023, 382, 110631. [Google Scholar] [CrossRef] [PubMed]
- Zvereva, M.I.; Shcherbakova, D.M.; Dontsova, O.A. Telomerase: Structure, Functions, and Activity Regulation. Biochemistry 2010, 75, 1563–1583. [Google Scholar] [CrossRef] [PubMed]
- Wang, Y.; Sušac, L.; Feigon, J. Structural Biology of Telomerase. Cold Spring Harb. Perspect. Biol. 2019, 11, a032383. [Google Scholar] [CrossRef]
- Dratwa, M.; Wysoczańska, B.; Łacina, P.; Kubik, T.; Bogunia-Kubik, K. TERT—Regulation and Roles in Cancer Formation. Front. Immunol. 2020, 11, 589929. [Google Scholar] [CrossRef]
- Sandin, S.; Rhodes, D. Telomerase Structure. Curr. Opin. Struct. Biol. 2014, 25, 104–110. [Google Scholar] [CrossRef]
- Eitsuka, T.; Nakagawa, K.; Kato, S.; Ito, J.; Otoki, Y.; Takasu, S.; Shimizu, N.; Takahashi, T.; Miyazawa, T. Modulation of Telomerase Activity in Cancer Cells by Dietary Compounds: A Review. Int. J. Mol. Sci. 2018, 19, 478. [Google Scholar] [CrossRef] [PubMed]
- Kim, N.W.; Piatyszek, M.A.; Prowse, K.R.; Harley, C.B.; West, M.D.; Ho, P.L.C.; Coviello, G.M.; Wright, W.E.; Weinrich, S.L.; Shay, J.W. Specific Association of Human Telomerase Activity with Immortal Cells and Cancers. Science 1994, 266, 2011–2015. [Google Scholar] [CrossRef] [PubMed]
- Trybek, T.; Kowalik, A.; Góźdź, S.; Kowalska, A. Telomeres and Telomerase in Oncogenesis (Review). Oncol. Lett. 2020, 20, 1015–1027. [Google Scholar] [CrossRef] [PubMed]
- Cesare, A.J.; Reddel, R.R. Alternative Lengthening of Telomeres: Models, Mechanisms and Implications. Nat. Rev. Genet. 2010, 11, 319–330. [Google Scholar] [CrossRef] [PubMed]
- Ganesan, K.; Xu, B. Telomerase Inhibitors from Natural Products and Their Anticancer Potential. Int. J. Mol. Sci. 2018, 19, 13. [Google Scholar] [CrossRef] [PubMed]
- Wei, Z.-Z.; Qin, Q.-P.; Chen, J.-N.; Chen, Z.-F. Oxoisoaporphine as Potent Telomerase Inhibitor. Molecules 2016, 21, 1534. [Google Scholar] [CrossRef] [PubMed]
- Balci Okcanoglu, T.; Avci, C.B.; Yılmaz Süslüer, S.; Kayabasi, C.; Saydam, G.; Gunduz, C. Genistein-Induced Apoptosis Affects Human Telomerase Reverse Transcriptase Activity in Acute Promyelocytic Leukemia. Cyprus J. Med. Sci. 2020, 5, 153–156. [Google Scholar] [CrossRef]
- Zhou, Y.; Zheng, J.; Li, Y.; Xu, D.-P.; Li, S.; Chen, Y.-M.; Li, H.-B. Natural Polyphenols for Prevention and Treatment of Cancer. Nutrients 2016, 8, 515. [Google Scholar] [CrossRef] [PubMed]
- Liu, C.; Dong, S.; Wang, X.; Xu, H.; Yang, X.; Wu, S.; Jiang, X.; Kan, M.; Xu, C. Research Progress of Polyphenols in Nanoformulations for Antibacterial Application. Mater. Today Bio 2023, 21, 100729. [Google Scholar] [CrossRef]
- Grabska-Kobyłecka, I.; Szpakowski, P.; Król, A.; Książek-Winiarek, D.; Kobyłecki, A.; Głąbiński, A.; Nowak, D. Polyphenols and Their Impact on the Prevention of Neurodegenerative Diseases and Development. Nutrients 2023, 15, 3454. [Google Scholar] [CrossRef]
- Pi, X.; Sun, Y.; Cheng, J.; Fu, G.; Guo, M. A Review on Polyphenols and Their Potential Application to Reduce Food Allergenicity. Crit. Rev. Food Sci. Nutr. 2023, 63, 10014–10031. [Google Scholar] [CrossRef] [PubMed]
- Li, L.; Chong, L.; Huang, T.; Ma, Y.; Li, Y.; Ding, H. Natural Products and Extracts from Plants as Natural UV Filters for Sunscreens: A Review. Anim. Model. Exp. Med. 2023, 6, 183–195. [Google Scholar] [CrossRef] [PubMed]
- Mukherjee (nee Chakraborty), S.; Ghosh, U.; Bhattacharyya, N.P.; Bhattacharya, R.K.; Dey, S.; Roy, M. Curcumin-Induced Apoptosis in Human Leukemia Cell HL-60 Is Associated with Inhibition of Telomerase Activity. Mol. Cell. Biochem. 2007, 297, 31–39. [Google Scholar] [CrossRef] [PubMed]
- Dikmen, M.; Canturk, Z.; Ozturk, Y.; Tunali, Y. Investigation of the Apoptotic Effect of Curcumin in Human Leukemia HL-60 Cells by Using Flow Cytometry. Cancer Biother. Radiopharm. 2010, 25, 749–755. [Google Scholar] [CrossRef] [PubMed]
- Chakraborty, S.; Ghosh, U.; Bhattacharyya, N.P.; Bhattacharya, R.K.; Roy, M. Inhibition of Telomerase Activity and Induction of Apoptosis by Curcumin in K-562 Cells. Mutat. Res./Fundam. Mol. Mech. Mutagen. 2006, 596, 81–90. [Google Scholar] [CrossRef] [PubMed]
- Berletch, J.B.; Liu, C.; Love, W.K.; Andrews, L.G.; Katiyar, S.K.; Tollefsbol, T.O. Epigenetic and Genetic Mechanisms Contribute to Telomerase Inhibition by EGCG. J. Cell. Biochem. 2008, 103, 509–519. [Google Scholar] [CrossRef] [PubMed]
- Ghasemi-Pirbaluti, M.; Pourgheysari, B.; Shirzad, H.; Motaghi, E.; Askarian Dehkordi, N.; Surani, Z.; Shirzad, M.; Beshkar, P.; Pirayesh, A. Effect of Epigallocatechin-3-Gallate (EGCG) on Cell Proliferation Inhibition and Apoptosis Induction in Lymphoblastic Leukemia Cell Line. J. HerbMed Pharmacol. 2015, 4, 65–68. [Google Scholar]
- Ghasemimehr, N.; Farsinejad, A.; Mirzaee Khalilabadi, R.; Yazdani, Z.; Fatemi, A. The Telomerase Inhibitor MST-312 Synergistically Enhances the Apoptotic Effect of Doxorubicin in Pre-B Acute Lymphoblastic Leukemia Cells. Biomed. Pharmacother. 2018, 106, 1742–1750. [Google Scholar] [CrossRef] [PubMed]
- Safa, M.; Tavasoli, B.; Manafi, R.; Kiani, F.; Kashiri, M.; Ebrahimi, S.; Kazemi, A. Indole-3-Carbinol Suppresses NF-ΚB Activity and Stimulates the P53 Pathway in Pre-B Acute Lymphoblastic Leukemia Cells. Tumor Biol. 2015, 36, 3919–3930. [Google Scholar] [CrossRef]
- Safa, M.; Jafari, L.; Alikarami, F.; Manafi Shabestari, R.; Kazemi, A. Indole-3-Carbinol Induces Apoptosis of Chronic Myelogenous Leukemia Cells through Suppression of STAT5 and Akt Signaling Pathways. Tumor Biol. 2017, 39, 101042831770576. [Google Scholar] [CrossRef]
- Moon, D.-O.; Kim, M.-O.; Lee, J.-D.; Choi, Y.H.; Kim, G.-Y. Butein Suppresses C-Myc-Dependent Transcription and Akt-Dependent Phosphorylation of HTERT in Human Leukemia Cells. Cancer Lett. 2009, 286, 172–179. [Google Scholar] [CrossRef] [PubMed]
- Salehi, B.; Venditti, A.; Sharifi-Rad, M.; Kręgiel, D.; Sharifi-Rad, J.; Durazzo, A.; Lucarini, M.; Santini, A.; Souto, E.; Novellino, E.; et al. The Therapeutic Potential of Apigenin. Int. J. Mol. Sci. 2019, 20, 1305. [Google Scholar] [CrossRef] [PubMed]
- Jayasooriya, R.G.P.T.; Kang, S.H.; Kang, C.H.; Choi, Y.H.; Moon, D.O.; Hyun, J.W.; Chang, W.Y.; Kim, G.Y. Apigenin Decreases Cell Viability and Telomerase Activity in Human Leukemia Cell Lines. Food Chem. Toxicol. 2012, 50, 2605–2611. [Google Scholar] [CrossRef] [PubMed]
- Chambers, C.S.; Holečková, V.; Petrásková, L.; Biedermann, D.; Valentová, K.; Buchta, M.; Křen, V. The Silymarin Composition… and Why Does It Matter??? Food Res. Int. 2017, 100, 339–353. [Google Scholar] [CrossRef] [PubMed]
- Faezizadeh, Z.; Ali, S.; Mesbah-Namin, R.; Allameh, A. The Effect of Silymarin on Telomerase Activity in the Human Leukemia Cell Line K562. Planta Med. 2012, 78, 899–902. [Google Scholar] [CrossRef] [PubMed]
- Ren, X.; Zhang, Z.; Tian, J.; Wang, H.; Song, G.; Guo, Q.; Tian, J.; Han, Y.; Liao, Q.; Liu, G.; et al. The Downregulation of C-Myc and Its Target Gene HTERT Is Associated with the Antiproliferative Effects of Baicalin on HL-60 Cells. Oncol. Lett. 2017, 14, 6833–6840. [Google Scholar] [CrossRef] [PubMed][Green Version]
- Blaner, W.S. Present Knowledge in Nutrition, 11th ed.; Elsevier: Amsterdam, The Netherlands, 2020; Volume 5, ISBN 9780323661621. [Google Scholar]
- Wolf, G. The Discovery of the Visual Function of Vitamin A. J. Nutr. 2001, 131, 1647–1650. [Google Scholar] [CrossRef]
- Chang, Q.; Chen, Z.; You, J.; McNutt, M.A.; Zhang, T.; Han, Z.; Zhang, X.; Gong, E.; Gu, J. All-Trans-Retinoic Acid Induces Cell Growth Arrest in a Human Medulloblastoma Cell Line. J. Neurooncol. 2007, 84, 263–267. [Google Scholar] [CrossRef] [PubMed]
- Liu, L.; Berletch, J.B.; Green, J.G.; Pate, M.S.; Andrews, L.G.; Tollefsbol, T.O. Telomerase Inhibition by Retinoids Precedes Cytodifferentiation of Leukemia Cells and May Contribute to Terminal Differentiation. Mol. Cancer Ther. 2004, 3, 1003–1009. [Google Scholar] [CrossRef]
- Zhang, J.-W.; Gu, J.; Wang, Z.-Y.; Chen, S.-J.; Chen, Z. Mechanisms Ofall-Trans Retinoic Acid-Induced Differentiation of Acute Promyelocytic Leukemia Cells. J. Biosci. 2000, 25, 275–284. [Google Scholar] [CrossRef]
- Van Gils, N.; Verhagen, H.J.M.P.; Rutten, A.; Menezes, R.X.; Tsui, M.L.; Vermue, E.; Dekens, E.; Brocco, F.; Denkers, F.; Kessler, F.L.; et al. IGFBP7 Activates Retinoid Acid-Induced Responses in Acute Myeloid Leukemia Stem and Progenitor Cells. Blood Adv. 2020, 4, 6368–6383. [Google Scholar] [CrossRef] [PubMed]
- De Thé, H. Differentiation Therapy Revisited. Nat. Rev. Cancer 2018, 18, 117–127. [Google Scholar] [CrossRef] [PubMed]
- Yilmaz, M.; Kantarjian, H.; Ravandi, F. Acute Promyelocytic Leukemia Current Treatment Algorithms. Blood Cancer J. 2021, 11, 123. [Google Scholar] [CrossRef] [PubMed]
- Masetti, R.; Vendemini, F.; Zama, D.; Biagi, C.; Gasperini, P.; Pession, A. All-Trans Retinoic Acid in the Treatment of Pediatric Acute Promyelocytic Leukemia. Expert Rev. Anticancer Ther. 2012, 12, 1191–1204. [Google Scholar] [CrossRef] [PubMed]
- Kutny, M.A.; Alonzo, T.A.; Abla, O.; Rajpurkar, M.; Gerbing, R.B.; Wang, Y.C.; Hirsch, B.A.; Raimondi, S.; Kahwash, S.; Hardy, K.K.; et al. Assessment of Arsenic Trioxide and All-Trans Retinoic Acid for the Treatment of Pediatric Acute Promyelocytic Leukemia A Report from the Children’s Oncology Group AAML1331 Trial. JAMA Oncol. 2021, 8, 79–81. [Google Scholar] [CrossRef] [PubMed]
- Farinello, D.; Wozińska, M.; Lenti, E.; Genovese, L.; Bianchessi, S.; Migliori, E.; Sacchetti, N.; di Lillo, A.; Bertilaccio, M.T.S.; de Lalla, C.; et al. A Retinoic Acid-Dependent Stroma-Leukemia Crosstalk Promotes Chronic Lymphocytic Leukemia Progression. Nat. Commun. 2018, 9, 1787. [Google Scholar] [CrossRef]
- Luong, Q.T.; Koeffler, H.P. Vitamin D Compounds in Leukemia. J. Steroid Biochem. Mol. Biol. 2005, 97, 195–202. [Google Scholar] [CrossRef] [PubMed]
- Kulling, P.M.; Olson, K.C.; Olson, T.L.; Feith, D.J.; Loughran, T.P. Vitamin D in Hematological Disorders and Malignancies. Eur. J. Haematol. 2017, 98, 187–197. [Google Scholar] [CrossRef] [PubMed]
- Holick, M.F. Vitamin D: Importance in the Prevention of Cancers, Type 1 Diabetes, Heart Disease, and Osteoporosis. Am. J. Clin. Nutr. 2004, 79, 362–371. [Google Scholar] [CrossRef]
- Christakos, S.; Dhawan, P.; Verstuyf, A.; Verlinden, L.; Carmeliet, G. Vitamin D: Metabolism, Molecular Mechanism of Action, and Pleiotropic Effects. Physiol. Rev. 2016, 96, 365–408. [Google Scholar] [CrossRef]
- Trump, D.L.; Deeb, K.K.; Johnson, C.S. Vitamin D: Considerations in the Continued Development as an Agent for Cancer Prevention and Therapy. Cancer J. 2010, 16, 1–9. [Google Scholar] [CrossRef] [PubMed]
- Kasiappan, R.; Shen, Z.; Tse, A.K.-W.; Jinwal, U.; Tang, J.; Lungchukiet, P.; Sun, Y.; Kruk, P.; Nicosia, S.V.; Zhang, X.; et al. 1,25-Dihydroxyvitamin D3 Suppresses Telomerase Expression and Human Cancer Growth through MicroRNA-498. J. Biol. Chem. 2012, 287, 41297–41309. [Google Scholar] [CrossRef] [PubMed]
- James, S.; Williams, M.; Newland, A.; Colston, K. Leukemia Cell Differentiation: Cellular and Molecular Interactions of Retinoids and Vitamin D. Gen. Pharmacol. Vasc. Syst. 1999, 32, 143–154. [Google Scholar] [CrossRef] [PubMed]
- Hisatake, J.; Kubota, T.; Hisatake, Y.; Uskokovic, M.; Tomoyasu, S.; Koeffler, H.P. 5,6-Trans-16-Ene-Vitamin D3: A New Class of Potent Inhibitors of Proliferation of Prostate, Breast, and Myeloid Leukemic Cells. Cancer Res. 1999, 59, 4023–4029. [Google Scholar] [PubMed]
- Delvin, E.; Alos, N.; Rauch, F.; Marcil, V.; Morel, S.; Boisvert, M.; Lecours, M.-A.; Laverdière, C.; Sinnett, D.; Krajinovic, M.; et al. Vitamin D Nutritional Status and Bone Turnover Markers in Childhood Acute Lymphoblastic Leukemia Survivors: A PETALE Study. Clin. Nutr. 2019, 38, 912–919. [Google Scholar] [CrossRef] [PubMed]
- Cao, H.; Xu, Y.; de Necochea-Campion, R.; Baylink, D.J.; Payne, K.J.; Tang, X.; Ratanatharathorn, C.; Ji, Y.; Mirshahidi, S.; Chen, C.-S. Application of Vitamin D and Vitamin D Analogs in Acute Myelogenous Leukemia. Exp. Hematol. 2017, 50, 1–12. [Google Scholar] [CrossRef] [PubMed]
- Lee, H.J.; Muindi, J.R.; Tan, W.; Hu, Q.; Wang, D.; Liu, S.; Wilding, G.E.; Ford, L.A.; Sait, S.N.J.; Block, A.W.; et al. Low 25(OH) Vitamin D3 Levels Are Associated with Adverse Outcome in Newly Diagnosed, Intensively Treated Adult Acute Myeloid Leukemia. Cancer 2014, 120, 521–529. [Google Scholar] [CrossRef] [PubMed]
- Thomas, X.; Chelghoum, Y.; Fanari, N.; Cannas, G. Serum 25-Hydroxyvitamin D Levels Are Associated with Prognosis in Hematological Malignancies. Hematology 2011, 16, 278–283. [Google Scholar] [CrossRef] [PubMed]
- Shanafelt, T.D.; Drake, M.T.; Maurer, M.J.; Allmer, C.; Rabe, K.G.; Slager, S.L.; Weiner, G.J.; Call, T.G.; Link, B.K.; Zent, C.S.; et al. Vitamin D Insufficiency and Prognosis in Chronic Lymphocytic Leukemia. Blood 2011, 117, 1492–1498. [Google Scholar] [CrossRef]
- Tan, B.; Watson, R.R.; Preedy, V.R. (Eds.) Tocotrienols: Vitamin E Beyond Tocopherols, 2nd ed.; CRC Press: Boca Raton, FL, USA, 2012; ISBN 9781439884430. [Google Scholar]
- Aggarwal, B.B.; Sundaram, C.; Prasad, S.; Kannappan, R. Tocotrienols, the Vitamin E of the 21st Century: Its Potential against Cancer and Other Chronic Diseases. Biochem. Pharmacol. 2010, 80, 1613–1631. [Google Scholar] [CrossRef]
- Traber, M.G. Vitamin E, Nuclear Receptors and Xenobiotic Metabolism. Arch. Biochem. Biophys. 2004, 423, 6–11. [Google Scholar] [CrossRef] [PubMed]
- Ghanem, P.; Zouein, A.; Mohamad, M.; Hodroj, M.H.; Haykal, T.; Abou Najem, S.; Naim, H.Y.; Rizk, S. The Vitamin E Derivative Gamma Tocotrienol Promotes Anti-Tumor Effects in Acute Myeloid Leukemia Cell Lines. Nutrients 2019, 11, 2808. [Google Scholar] [CrossRef] [PubMed]
- Shvachko, L.P.; Zavelevich, M.P.; Gluzman, D.F.; Kravchuk, I.V.; Telegeev, G.D. Vitamin E; Activates Expression of C/EBP Alpha Transcription Factor and G-CSF Receptor in Leukemic K562 Cells. Exp. Oncol. 2018, 40, 328–331. [Google Scholar] [CrossRef]
- Mustafa, K.N.; Sheikha, A.; Thanoon, I.A.-J.; Hussein, F.N. Effect of Vitamin E on Chemotherapy Induced Oxidative Stress and Immunoglobulin Levels in Patients with Acute Myeloid Leukemia. Tikrit Med. J. 2008, 14, 146–151. [Google Scholar]
- Telegeev, G.D.; Gluzman, D.F.; Zavelevich, M.P.; Shvachko, L.P. Aberrant Expression of Placental-like Alkaline Phosphatase in Chronic Myeloid Leukemia Cells in Vitro and Its Modulation by Vitamin E. Exp. Oncol. 2020, 42, 31–34. [Google Scholar] [CrossRef]
- Hall, K.T.; Buring, J.E.; Mukamal, K.J.; Vinayaga Moorthy, M.; Wayne, P.M.; Kaptchuk, T.J.; Battinelli, E.M.; Ridker, P.M.; Sesso, H.D.; Weinstein, S.J.; et al. COMT and Alpha-Tocopherol Effects in Cancer Prevention: Gene-Supplement Interactions in Two Randomized Clinical Trials. JNCI J. Natl. Cancer Inst. 2019, 111, 684–694. [Google Scholar] [CrossRef]
- Institute of Medicine, Dietary Reference Intakes for Vitamin C, Vitamin E, Selenium, and Carotenoids; National Academies Press: Washington, DC, USA, 2000; Volume 3, ISBN 978-0-309-06935-9.
- Travica, N.; Ried, K.; Sali, A.; Scholey, A.; Hudson, I.; Pipingas, A. Vitamin C Status and Cognitive Function: A Systematic Review. Nutrients 2017, 9, 960. [Google Scholar] [CrossRef] [PubMed]
- Carr, A.; Maggini, S. Vitamin C and Immune Function. Nutrients 2017, 9, 1211. [Google Scholar] [CrossRef] [PubMed]
- Li, R.; Guo, C.; Li, Y.; Liang, X.; Su, M. Functional Benefit and Molecular Mechanism of Vitamin C against Perfluorooctanesulfonate-Associated Leukemia. Chemosphere 2021, 263, 128242. [Google Scholar] [CrossRef]
- Lin, X.-J.; Zou, X.-L.; Zhao, Z.-Y.; Wang, J.; Yang, Z.; Ni, X.; Wei, J. Effect of High Dose Vitamin C on Proliferation and Apoptosis of Acute Myeloid Leukemia Cells. Zhongguo Shi Yan Xue Ye Xue Za Zhi 2020, 28, 833–841. [Google Scholar] [CrossRef]
- Lunn, J.; Theobald, H.E. The Health Effects of Dietary Unsaturated Fatty Acids. Nutr. Bull. 2006, 31, 178–224. [Google Scholar] [CrossRef]
- Eitsuka, T.; Nakagawa, K.; Suzuki, T.; Miyazawa, T. Polyunsaturated Fatty Acids Inhibit Telomerase Activity in DLD-1 Human Colorectal Adenocarcinoma Cells: A Dual Mechanism Approach. Biochim. Biophys. Acta (BBA)-Mol. Cell Biol. Lipids 2005, 1737, 1–10. [Google Scholar] [CrossRef] [PubMed]
- Liu, W.H.; Hsu, C.Y.; Yung, B.Y.M. Nucleophosmin/B23 Regulates the Susceptibility of Human Leukemia HL-60 Cells to Sodium Butyrate-Induced Apoptosis and Inhibition of Telomerase Activity. Int. J. Cancer 1999, 83, 765–771. [Google Scholar] [CrossRef]
- Nie, D.; Huang, K.; Yin, S.; Li, Y.; Xie, S.; Ma, L.; Wang, X.; Wu, Y.; Xiao, J. Synergistic/Additive Interaction of Valproic Acid with Bortezomib on Proliferation and Apoptosis of Acute Myeloid Leukemia Cells. Leuk. Lymphoma 2012, 53, 2487–2495. [Google Scholar] [CrossRef]
- Li, Y.-Q.; Yin, S.-M.; Feng, S.-Q.; Nie, D.-N.; Xie, S.-F.; Ma, L.-P.; Wang, X.-J.; Wu, Y.-D. Effect of Valproic Acid on Apoptosis of Leukemia HL-60 Cells and Expression of h-Tert Gene. Zhongguo Shi Yan Xue Ye Xue Za Zhi 2010, 18, 1445–1450. [Google Scholar] [PubMed]
- Ji, Z.-N.; Ye, W.-C.; Liu, G.-G.; Hsiao, W.L.W. 23-Hydroxybetulinic Acid-Mediated Apoptosis Is Accompanied by Decreases in Bcl-2 Expression and Telomerase Activity in HL-60 Cells. Life Sci. 2002, 72, 1–9. [Google Scholar] [CrossRef]
- Oda, M.; Ueno, T.; Kasai, N.; Takahashi, H.; Yoshida, H.; Sugawara, F.; Sakaguchi, K.; Hayashi, H.; Mizushina, Y. Inhibition of Telomerase by Linear-Chain Fatty Acids: A Structural Analysis. Biochem. J. 2002, 367, 329–334. [Google Scholar] [CrossRef]
- Choi, Y.H. Apoptosis of U937 Human Leukemic Cells by Sodium Butyrate Is Associated with Inhibition of Telomerase Activity. Int. J. Oncol. 2006, 29, 1207–1213. [Google Scholar] [CrossRef][Green Version]
- Fahrmann, J.F.; Ballester, O.F.; Ballester, G.; Witte, T.R.; Salazar, A.J.; Kordusky, B.; Cowen, K.G.; Ion, G.; Primerano, D.A.; Boskovic, G.; et al. Inhibition of Nuclear Factor Kappa B Activation in Early-Stage Chronic Lymphocytic Leukemia by Omega-3 Fatty Acids. Cancer Investig. 2013, 31, 24–38. [Google Scholar] [CrossRef]
- El Amrousy, D.; El-Afify, D.; Khedr, R.; Ibrahim, A.M. Omega 3 Fatty Acids Can Reduce Early Doxorubicin-induced Cardiotoxicity in Children with Acute Lymphoblastic Leukemia. Pediatr. Blood Cancer 2022, 69, e29496. [Google Scholar] [CrossRef]
- Bükki, J.; Stanga, Z.; Tellez, F.B.; Duclos, K.; Kolev, M.; Krähenmann, P.; Pabst, T.; Iff, S.; Jüni, P. Omega-3 Poly-Unsaturated Fatty Acids for the Prevention of Severe Neutropenic Enterocolitis in Patients with Acute Myeloid Leukemia. Nutr. Cancer 2013, 65, 834–842. [Google Scholar] [CrossRef] [PubMed]
- Chagas, T.R.; Borges, D.S.; de Oliveira, P.F.; Mocellin, M.C.; Barbosa, A.M.; Camargo, C.Q.; del Moral, J.Â.G.; Poli, A.; Calder, P.C.; Trindade, E.B.S.M.; et al. Oral Fish Oil Positively Influences Nutritional-Inflammatory Risk in Patients with Haematological Malignancies during Chemotherapy with an Impact on Long-Term Survival: A Randomised Clinical Trial. J. Hum. Nutr. Diet. 2017, 30, 681–692. [Google Scholar] [CrossRef]
- Rücker, F.G.; Lang, K.M.; Fütterer, M.; Komarica, V.; Schmid, M.; Döhner, H.; Schlenk, R.F.; Döhner, K.; Knudsen, S.; Bullinger, L. Molecular Dissection of Valproic Acid Effects in Acute Myeloid Leukemia Identifies Predictive Networks. Epigenetics 2016, 11, 517–525. [Google Scholar] [CrossRef] [PubMed]
- Corsetti, M.T.; Salvi, F.; Perticone, S.; Baraldi, A.; de Paoli, L.; Gatto, S.; Pietrasanta, D.; Pini, M.; Primon, V.; Zallio, F.; et al. Hematologic Improvement and Response in Elderly AML/RAEB Patients Treated with Valproic Acid and Low-Dose Ara-C. Leuk. Res. 2011, 35, 991–997. [Google Scholar] [CrossRef] [PubMed]
- Woo, H.J.; Lee, S.J.; Choi, B.T.; Park, Y.-M.; Choi, Y.H. Induction of Apoptosis and Inhibition of Telomerase Activity by Trichostatin A, a Histone Deacetylase Inhibitor, in Human Leukemic U937 Cells. Exp. Mol. Pathol. 2007, 82, 77–84. [Google Scholar] [CrossRef] [PubMed]
- Caffeic Acid Undecyl Ester. Available online: https://pubchem.ncbi.nlm.nih.gov/compound/25022448 (accessed on 28 July 2023).
- Tomizawa, A.; Kanno, S.I.; Osanai, Y.; Goto, A.; Sato, C.; Yomogida, S.; Ishikawa, M. Induction of Apoptosis by a Potent Caffeic Acid Derivative, Caffeic Acid Undecyl Ester, Is Mediated by Mitochondrial Damage in NALM-6 Human B Cell Leukemia Cells. Oncol. Rep. 2013, 29, 425–429. [Google Scholar] [CrossRef]
- Tomizawa, A.; Kanno, S.I.; Osanai, Y.; Yomogida, S.; Ishikawa, M. Cytotoxic Effects of Caffeic Acid Undecyl Ester Are Involved in the Inhibition of Telomerase Activity in NALM-6 Human B-Cell Leukemia Cells. Oncol. Lett. 2013, 6, 875–877. [Google Scholar] [CrossRef] [PubMed][Green Version]
- Maiti, A.; Cortes, J.; Ferrajoli, A.; Estrov, Z.; Borthakur, G.; Garcia-Manero, G.; Jabbour, E.; Ravandi, F.; O’Brien, S.; Kantarjian, H. Phase II Trial of Homoharringtonine with Imatinib in Chronic, Accelerated, and Blast Phase Chronic Myeloid Leukemia. Leuk. Lymphoma 2017, 58, 2240–2242. [Google Scholar] [CrossRef] [PubMed]
- Innocenti, F.; Sibley, A.B.; Patil, S.A.; Etheridge, A.S.; Jiang, C.; Ou, F.S.; Howell, S.D.; Plummer, S.J.; Casey, G.; Bertagnolli, M.M.; et al. Genomic Analysis of Germline Variation Associated with Survival of Patients with Colorectal Cancer Treated with Chemotherapy Plus Biologics in CALGB/SWOG 80405 (Alliance). Clin. Cancer Res. 2021, 27, 267–276. [Google Scholar] [CrossRef]
- Ansari, M.A.; Khan, F.B.; Safdari, H.A.; Almatroudi, A.; Alzohairy, M.A.; Safdari, M.; Amirizadeh, M.; Rehman, S.; Equbal, M.J.; Hoque, M. Prospective Therapeutic Potential of Tanshinone IIA: An Updated Overview. Pharmacol. Res. 2021, 164, 105364. [Google Scholar] [CrossRef]
- Song, Y.; Yuan, S.-L.; Yang, Y.-M.; Wang, X.-J.; Huang, G.-Q.; Expand, A. Alteration of Activities of Telomerase in Tanshinone IIA Inducing Apoptosis of the Leukemia Cells. Zhongguo Zhong Yao Za Zhi = Zhongguo Zhongyao Zazhi = China J. Chin. Mater. Medica 2005, 30, 207–211. [Google Scholar]
- Tanshinone, I. Available online: https://pubchem.ncbi.nlm.nih.gov/compound/114917 (accessed on 27 August 2023).
- Liu, X.D.; Fan, R.F.; Zhang, Y.; Yang, H.Z.; Fang, Z.G.; Guan, W.B.; Lin, D.J.; Xiao, R.Z.; Huang, R.W.; Huang, H.Q.; et al. Down-Regulation of Telomerase Activity and Activation of Caspase-3 Are Responsible for Tanshinone I-Induced Apoptosis in Monocyte Leukemia Cells in Vitro. Int. J. Mol. Sci. 2010, 11, 2267–2280. [Google Scholar] [CrossRef]
- Lu, S.; Qin, X.; Yuan, S.; Li, Y.; Wang, L.; Jin, Y.; Zeng, G.; Yen, L.; Hu, J.; Dang, T.; et al. Effect of Tianshengyuan-1 (TSY-1) on Telomerase Activity and Hematopoietic Recovery—In Vitro, Ex Vivo, and in Vivo Studies. Int. J. Clin. Exp. Med. 2014, 7, 597–606. [Google Scholar] [PubMed]
- Yu, W.; Qin, X.; Jin, Y.; Li, Y.; Santiskulvong, C.; Vu, V.; Zeng, G.; Zhang, Z.; Chow, M.; Rao, J. Tianshengyuan-1 (TSY-1) Regulates Cellular Telomerase Activity by Methylation of TERT Promoter. Oncotarget 2017, 8, 7977–7988. [Google Scholar] [CrossRef]
- Nai, J.; Zhang, C.; Shao, H.; Li, B.; Li, H.; Gao, L.; Dai, M.; Zhu, L.; Sheng, H. Extraction, Structure, Pharmacological Activities and Drug Carrier Applications of Angelica Sinensis Polysaccharide. Int. J. Biol. Macromol. 2021, 183, 2337–2353. [Google Scholar] [CrossRef] [PubMed]
- Liu, J.; Xu, C.Y.; Cai, S.Z.; Zhou, Y.; Li, J.; Jiang, R.; Wang, Y.P. Senescence Effects of Angelica Sinensis Polysaccharides on Human Acute Myelogenous Leukemia Stem and Progenitor Cells. Asian Pac. J. Cancer Prev. 2013, 14, 6549–6556. [Google Scholar] [CrossRef] [PubMed][Green Version]
- Bao, J.; Dai, S.-M. A Chinese Herb Tripterygium Wilfordii Hook F in the Treatment of Rheumatoid Arthritis: Mechanism, Efficacy, and Safety. Rheumatol. Int. 2011, 31, 1123–1129. [Google Scholar] [CrossRef] [PubMed]
- Chang, Z.; Qin, W.; Zheng, H.; Schegg, K.; Han, L.; Liu, X.; Wang, Y.; Wang, Z.; McSwiggin, H.; Peng, H.; et al. Triptonide Is a Reversible Non-Hormonal Male Contraceptive Agent in Mice and Non-Human Primates. Nat. Commun. 2021, 12, 1253. [Google Scholar] [CrossRef]
- Zhang, M.; Tan, S.; Yu, D.; Zhao, Z.; Zhang, B.; Zhang, P.; Lv, C.; Zhou, Q.; Cao, Z. Triptonide Inhibits Lung Cancer Cell Tumorigenicity by Selectively Attenuating the Shh-Gli1 Signaling Pathway. Toxicol. Appl. Pharmacol. 2019, 365, 1–8. [Google Scholar] [CrossRef]
- Zhang, M.; Meng, M.; Liu, Y.; Qi, J.; Zhao, Z.; Qiao, Y.; Hu, Y.; Lu, W.; Zhou, Z.; Xu, P.; et al. Triptonide Effectively Inhibits Triple-Negative Breast Cancer Metastasis through Concurrent Degradation of Twist1 and Notch1 Oncoproteins. Breast Cancer Res. 2021, 23, 116. [Google Scholar] [CrossRef]
- Fu, L.; Niu, X.; Jin, R.; Xu, F.; Ding, J.; Zhang, L.; Huang, Z. Triptonide Inhibits Metastasis Potential of Thyroid Cancer Cells via Astrocyte Elevated Gene-1. Transl. Cancer Res. 2020, 9, 1195–1204. [Google Scholar] [CrossRef]
- Pan, Y.; Meng, M.; Zheng, N.; Cao, Z.; Yang, P.; Xi, X.; Zhou, Q. Targeting of Multiple Senescence-Promoting Genes and Signaling Pathways by Triptonide Induces Complete Senescence of Acute Myeloid Leukemia Cells. Biochem. Pharmacol. 2017, 126, 34–50. [Google Scholar] [CrossRef]
- Huynh, D.L.; Ngau, T.H.; Nguyen, N.H.; Tran, G.-B.; Nguyen, C.T. Potential Therapeutic and Pharmacological Effects of Wogonin: An Updated Review. Mol. Biol. Rep. 2020, 47, 9779–9789. [Google Scholar] [CrossRef]
- Khan, N.M.; Haseeb, A.; Ansari, M.Y.; Devarapalli, P.; Haynie, S.; Haqqi, T.M. Wogonin, a Plant Derived Small Molecule, Exerts Potent Anti-Inflammatory and Chondroprotective Effects through the Activation of ROS/ERK/Nrf2 Signaling Pathways in Human Osteoarthritis Chondrocytes. Free Radic. Biol. Med. 2017, 106, 288–301. [Google Scholar] [CrossRef]
- Huang, S.T.; Wang, C.Y.; Yang, R.C.; Chu, C.J.; Wu, H.T.; Pang, J.H.S. Wogonin, an Active Compound in Scutellaria baicalensis, Induces Apoptosis and Reduces Telomerase Activity in the HL-60 Leukemia Cells. Phytomedicine 2010, 17, 47–54. [Google Scholar] [CrossRef] [PubMed]
- Khaiwa, N.; Maarouf, N.R.; Darwish, M.H.; Alhamad, D.W.M.; Sebastian, A.; Hamad, M.; Omar, H.A.; Orive, G.; Al-Tel, T.H. Camptothecin’s Journey from Discovery to WHO Essential Medicine: Fifty Years of Promise. Eur. J. Med. Chem. 2021, 223, 113639. [Google Scholar] [CrossRef] [PubMed]
- Lazareva, N.F.; Baryshok, V.P.; Lazarev, I.M. Silicon-Containing Analogs of Camptothecin as Anticancer Agents. Arch. Pharm. 2018, 351, 1700297. [Google Scholar] [CrossRef] [PubMed]
- Jiang, J.F.; Liu, W.J.; Ding, J. Regulation of Telomerase Activity in Camptothecin-Induced Apoptosis of Human Leukemia HL-60 Cells. Acta Pharmacol. Sin. 2000, 21, 759–764. [Google Scholar]
- Xie, W.-Z.; Lin, M.-F.; Huang, H.; Cai, Z. Homoharringtonine-Induced Apoptosis of Human Leukemia HL-60 Cells Is Associated with Down-Regulation of Telomerase. Am. J. Chin. Med. 2006, 34, 233–244. [Google Scholar] [CrossRef]
- Kanno, S.; Kitajima, Y.; Kakuta, M.; Osanai, Y.; Kurauchi, K.; Ujibe, M.; Ishikawa, M. Costunolide-Induced Apoptosis Is Caused by Receptor-Mediated Pathway and Inhibition of Telomerase Activity in NALM-6 Cells. Biol. Pharm. Bull. 2008, 31, 1024–1028. [Google Scholar] [CrossRef]
- Oh, J.H.; Lee, T.-J.; Kim, S.H.; Choi, Y.H.; Lee, S.H.; Lee, J.M.; Kim, Y.-H.; Park, J.-W.; Kwon, T.K. Induction of Apoptosis by Withaferin A in Human Leukemia U937 Cells through Down-Regulation of Akt Phosphorylation. Apoptosis 2008, 13, 1494–1504. [Google Scholar] [CrossRef] [PubMed]
- Yu, Y.; Katiyar, S.P.; Sundar, D.; Kaul, Z.; Miyako, E.; Zhang, Z.; Kaul, S.C.; Reddel, R.R.; Wadhwa, R. Withaferin-A Kills Cancer Cells with and without Telomerase: Chemical, Computational and Experimental Evidences. Cell Death Dis. 2017, 8, e2755. [Google Scholar] [CrossRef] [PubMed]
- Gomes, C.L.; de Albuquerque Wanderley Sales, V.; Gomes de Melo, C.; Ferreira da Silva, R.M.; Vicente Nishimura, R.H.; Rolim, L.A.; Rolim Neto, P.J. Beta-Lapachone: Natural Occurrence, Physicochemical Properties, Biological Activities, Toxicity and Synthesis. Phytochemistry 2021, 186, 112713. [Google Scholar] [CrossRef] [PubMed]
- Moon, D.-O.; Kang, C.-H.; Kim, M.-O.; Jeon, Y.-J.; Lee, J.-D.; Choi, Y.H.; Kim, G.-Y. β-Lapachone (LAPA) Decreases Cell Viability and Telomerase Activity in Leukemia Cells: Suppression of Telomerase Activity by LAPA. J. Med. Food 2010, 13, 481–488. [Google Scholar] [CrossRef] [PubMed]
- Liu, W.-J.; Jiang, J.-F.; Xiao, D.; Ding, J. Down-Regulation of Telomerase Activity via Protein Phosphatase 2A Activation in Salvicine-Induced Human Leukemia HL-60 Cell Apoptosis. Biochem. Pharmacol. 2002, 64, 1677–1687. [Google Scholar] [CrossRef] [PubMed]
- Khathayer, F.; Ray, S.K. Diosgenin as a Novel Alternative Therapy for Inhibition of Growth, Invasion, and Angiogenesis Abilities of Different Glioblastoma Cell Lines. Neurochem. Res. 2020, 45, 2336–2351. [Google Scholar] [CrossRef] [PubMed]
- Semwal, P.; Painuli, S.; Abu-Izneid, T.; Rauf, A.; Sharma, A.; Daştan, S.D.; Kumar, M.; Alshehri, M.M.; Taheri, Y.; Das, R.; et al. Diosgenin: An Updated Pharmacological Review and Therapeutic Perspectives. Oxid. Med. Cell. Longev. 2022, 2022, 1035441. [Google Scholar] [CrossRef] [PubMed]
- Jiang, S.; Fan, J.; Wang, Q.; Ju, D.; Feng, M.; Li, J.; Guan, Z.; An, D.; Wang, X.; Ye, L. Diosgenin Induces ROS-Dependent Autophagy and Cytotoxicity via MTOR Signaling Pathway in Chronic Myeloid Leukemia Cells. Phytomedicine 2016, 23, 243–252. [Google Scholar] [CrossRef] [PubMed]
- Tang, Y.; Wang, X.; Zhou, Y.; Wang, Y.; Ding, J. Ginsenoside Rg1 Induces Senescence of Leukemic Stem Cells by Upregulating P16INK4a and Downregulating HTERT Expression. Adv. Clin. Exp. Med. 2021, 30, 599–605. [Google Scholar] [CrossRef]
- Park, S.E.; Park, C.; Kim, S.H.; Hossain, M.A.; Kim, M.Y.; Chung, H.Y.; Son, W.S.; Kim, G.-Y.; Choi, Y.H.; Kim, N.D. Korean Red Ginseng Extract Induces Apoptosis and Decreases Telomerase Activity in Human Leukemia Cells. J. Ethnopharmacol. 2009, 121, 304–312. [Google Scholar] [CrossRef]
- Gao, Y.; Chu, S.; Zhang, Z.; Chen, N. Hepataprotective Effects of Ginsenoside Rg1—A Review. J. Ethnopharmacol. 2017, 206, 178–183. [Google Scholar] [CrossRef] [PubMed]
- Chen, J.; Zhang, X.; Liu, X.; Zhang, C.; Shang, W.; Xue, J.; Chen, R.; Xing, Y.; Song, D.; Xu, R. Ginsenoside Rg1 Promotes Cerebral Angiogenesis via the PI3K/Akt/MTOR Signaling Pathway in Ischemic Mice. Eur. J. Pharmacol. 2019, 856, 172418. [Google Scholar] [CrossRef]
- Hou, T.; Liu, Y.; Wang, X.; Jiao, D.; Xu, H.; Shi, Q.; Wang, Y.; Li, W.; Wu, T.; Liang, Q. Ginsenoside Rg1 Promotes Lymphatic Drainage and Improves Chronic Inflammatory Arthritis. J. Musculoskelet. Neuronal Interact. 2020, 20, 526–534. [Google Scholar] [PubMed]
- Zhang, Z.; Song, Z.; Shen, F.; Xie, P.; Wang, J.; Zhu, A.; Zhu, G. Ginsenoside Rg1 Prevents PTSD-Like Behaviors in Mice through Promoting Synaptic Proteins, Reducing Kir4.1 and TNF-α in the Hippocampus. Mol. Neurobiol. 2021, 58, 1550–1563. [Google Scholar] [CrossRef] [PubMed]
- Ji, Z.N.; Ye, W.C.; Rui, J.; Wang, L.; Liu, G.Q. Patensin-Induced Apoptosis Is Accompanied by Decreased Bcl-2 Expression and Telomerase Activity in HL-60 Cells. J. Asian Nat. Prod. Res. 2004, 6, 107–114. [Google Scholar] [CrossRef] [PubMed]
- Platycodin, D. Available online: https://pubchem.ncbi.nlm.nih.gov/compound/Platycodin-D#section=Associated-Disorders-and-Diseases (accessed on 28 August 2023).
- Kim, M.O.; Moon, D.O.; Choi, Y.H.; Shin, D.Y.; Kang, H.S.; Choi, B.T.; Lee, J.D.; Li, W.; Kim, G.Y. Platycodin D Induces Apoptosis and Decreases Telomerase Activity in Human Leukemia Cells. Cancer Lett. 2008, 261, 98–107. [Google Scholar] [CrossRef]
- Eitsuka, T.; Nakagawa, K.; Igarashi, M.; Miyazawa, T. Telomerase Inhibition by Sulfoquinovosyldiacylglycerol from Edible Purple Laver (Porphyra yezoensis). Cancer Lett. 2004, 212, 15–20. [Google Scholar] [CrossRef] [PubMed]
- Jain, C.K.; Pradhan, B.S.; Banerjee, S.; Mondal, N.B.; Majumder, S.S.; Bhattacharyya, M.; Chakrabarti, S.; Roychoudhury, S.; Majumder, H.K. Sulfonoquinovosyl Diacylglyceride Selectively Targets Acute Lymphoblastic Leukemia Cells and Exerts Potent Anti-Leukemic Effects in Vivo. Sci. Rep. 2015, 5, 12082. [Google Scholar] [CrossRef] [PubMed]
- Dart, D.A.; Adams, K.E.; Akerman, I.; Lakin, N.D. Recruitment of the Cell Cycle Checkpoint Kinase ATR to Chromatin during S-Phase. J. Biol. Chem. 2004, 279, 16433–16440. [Google Scholar] [CrossRef]
- Shang, H.-S.; Shih, Y.-L.; Lee, C.-H.; Hsueh, S.-C.; Liu, J.-Y.; Liao, N.-C.; Chen, Y.-L.; Huang, Y.-P.; Lu, H.-F.; Chung, J.-G. Sulforaphane-Induced Apoptosis in Human Leukemia HL-60 Cells through Extrinsic and Intrinsic Signal Pathways and Altering Associated Genes Expression Assayed by CDNA Microarray. Environ. Toxicol. 2017, 32, 311–328. [Google Scholar] [CrossRef]
- Veisi, A.; Akbari, G.; Mard, S.A.; Badfar, G.; Zarezade, V.; Mirshekar, M.A. Role of Crocin in Several Cancer Cell Lines: An Updated Review. Iran. J. Basic. Med. Sci. 2020, 23, 3–12. [Google Scholar] [CrossRef] [PubMed]
- Sun, Y.; Wang, Z.; Wang, L.; Wang, L.-Z.; Zang, C.; Sun, L.-R. The Effect and Mechanisms of Proliferative Inhibition of Crocin on Human Leukaemia Jurkat Cells. West Indian Med. J. 2016, 64, 473. [Google Scholar] [CrossRef][Green Version]
- Tang, Y.-L.; Sun, X.; Huang, L.-B.; Liu, X.-J.; Qin, G.; Wang, L.-N.; Zhang, X.-L.; Ke, Z.-Y.; Luo, J.-S.; Liang, C.; et al. Melatonin Inhibits MLL-Rearranged Leukemia via RBFOX3/HTERT and NF-ΚB/COX-2 Signaling Pathways. Cancer Lett. 2019, 443, 167–178. [Google Scholar] [CrossRef] [PubMed]
- Kiladjian, J.-J.; Giraudier, S.; Cassinat, B. Interferon-Alpha for the Therapy of Myeloproliferative Neoplasms: Targeting the Malignant Clone. Leukemia 2016, 30, 776–781. [Google Scholar] [CrossRef] [PubMed]
- Kim, M.O.; Moon, D.O.; Kang, S.H.; Heo, M.S.; Choi, Y.H.; Jung, J.H.; Lee, J.D.; Kim, G.Y. Pectenotoxin-2 Represses Telomerase Activity in Human Leukemia Cells through Suppression of HTERT Gene Expression and Akt-Dependent HTERT Phosphorylation. FEBS Lett. 2008, 582, 3263–3269. [Google Scholar] [CrossRef] [PubMed]
- Meng, X.; Li, Y.; Li, S.; Zhou, Y.; Gan, R.-Y.; Xu, D.-P.; Li, H.-B. Dietary Sources and Bioactivities of Melatonin. Nutrients 2017, 9, 367. [Google Scholar] [CrossRef]
- Simonsson, B.; Hjorth-Hansen, H.; Weis Bjerrum, O.; Porkka, K. Interferon Alpha for Treatment of Chronic Myeloid Leukemia. Curr. Drug Targets 2011, 12, 420–428. [Google Scholar] [CrossRef] [PubMed]
- Jiang, H.; Liu, X.-H.; Kong, J.; Wang, J.; Jia, J.-S.; Lu, S.-Y.; Gong, L.-Z.; Zhao, X.-S.; Jiang, Q.; Chang, Y.-J.; et al. Interferon-α as Maintenance Therapy Can Significantly Reduce Relapse in Patients with Favorable-Risk Acute Myeloid Leukemia. Leuk. Lymphoma 2021, 62, 2949–2956. [Google Scholar] [CrossRef]
- Magenau, J.M.; Peltier, D.; Riwes, M.; Pawarode, A.; Parkin, B.; Braun, T.; Anand, S.; Ghosh, M.; Maciejewski, J.; Yanik, G.; et al. Type 1 Interferon to Prevent Leukemia Relapse after Allogeneic Transplantation. Blood Adv. 2021, 5, 5047–5056. [Google Scholar] [CrossRef]
- Cogulu, O.; Biray, C.; Gunduz, C.; Karaca, E.; Aksoylar, S.; Sorkun, K.; Salih, B.; Ozkinay, F. Effects of Manisa Propolis on Telomerase Activity in Leukemia Cells Obtained from the Bone Marrow of Leukemia Patients. Int. J. Food Sci. Nutr. 2009, 60, 601–605. [Google Scholar] [CrossRef]
- Gunduz, C.; Biray, C.; Kosova, B.; Yilmaz, B.; Eroglu, Z.; Şahin, F.; Omay, S.B.; Cogulu, O. Evaluation of Manisa Propolis Effect on Leukemia Cell Line by Telomerase Activity. Leuk. Res. 2005, 29, 1343–1346. [Google Scholar] [CrossRef] [PubMed]
- Park, C.; Jung, J.H.; Kim, N.D.; Choi, Y.H. Inhibition of Cyclooxygenase-2 and Telomerase Activities in Human Leukemia Cells by Dideoxypetrosynol A, a Polyacetylene from the Marine Sponge Petrosia sp. Int. J. Oncol. 2007, 30, 291–298. [Google Scholar] [CrossRef] [PubMed]
- Park, C.; Kim, G.Y.; Kim, W.I.; Hong, S.H.; Park, D.I.; Kim, N.D.; Bae, S.J.; Jung, J.H.; Choi, Y.H. Induction of Apoptosis by (Z)-Stellettic Acid C, an Acetylenic Acid from the Sponge Stelletta Sp., Is Associated with Inhibition of Telomerase Activity in Human Leukemic U937 Cells. Chemotherapy 2007, 53, 160–168. [Google Scholar] [CrossRef] [PubMed]
- Sogawa, K.; Yamada, T.; Sumida, T.; Hamakawa, H.; Kuwabara, H.; Matsuda, M.; Muramatsu, Y.; Kose, H.; Matsumoto, K.; Sasaki, Y.; et al. Induction of Apoptosis and Inhibition of DNA Topoisomerase-I in K-562 Cells by a Marine Microalgal Polysaccharide. Life Sci. 2000, 66, PL227–PL231. [Google Scholar] [CrossRef] [PubMed]
- Sogawa, K.; Sumida, T.; Hamakawa, H.; Yamada, T.; Matsumoto, K.; Matsuda, M.; Oda, H.; Miyake, H.; Tashiro, S.; Okutani, K. Inhibitory Effect of a Marine Microalgal Polysaccharide on the Telomerase Activity in K562 Cells. Res. Commun. Mol. Pathol. Pharmacol. 1998, 99, 259–265. [Google Scholar]
- Shin-ya, K. Telomerase Inhibitor, Telomestatin, a Specific Mechanism to Interact with Telomere Structure. Nihon Rinsho 2004, 62, 1277–1282. [Google Scholar] [PubMed]
- Tauchi, T.; Shin-ya, K.; Sashida, G.; Sumi, M.; Okabe, S.; Ohyashiki, J.H.; Ohyashiki, K. Telomerase Inhibition with a Novel G-Quadruplex-Interactive Agent, Telomestatin: In Vitro and in Vivo Studies in Acute Leukemia. Oncogene 2006, 25, 5719–5725. [Google Scholar] [CrossRef] [PubMed]
- Ashraf, S.A.; Elkhalifa, A.E.O.; Siddiqui, A.J.; Patel, M.; Awadelkareem, A.M.; Snoussi, M.; Ashraf, M.S.; Adnan, M.; Hadi, S. Cordycepin for Health and Wellbeing: A Potent Bioactive Metabolite of an Entomopathogenic Medicinal Fungus Cordyceps with Its Nutraceutical and Therapeutic Potential. Molecules 2020, 25, 2735. [Google Scholar] [CrossRef]
- Jang, K.J.; Kwon, G.S.; Jeong, J.W.; Kim, C.H.; Yoon, H.M.; Kim, G.Y.; Shim, J.H.; Moon, S.K.; Kim, W.J.; Choi, Y.H. Cordyceptin Induces Apoptosis through Repressing HTERT Expression and Inducing Extranuclear Export of HTERT. J. Biosci. Bioeng. 2015, 119, 351–357. [Google Scholar] [CrossRef]
- Hetland, G.; Tangen, J.-M.; Mahmood, F.; Mirlashari, M.R.; Nissen-Meyer, L.S.H.; Nentwich, I.; Therkelsen, S.P.; Tjønnfjord, G.E.; Johnson, E. Antitumor, Anti-Inflammatory and Antiallergic Effects of Agaricus Blazei Mushroom Extract and the Related Medicinal Basidiomycetes Mushrooms, Hericium Erinaceus and Grifola Frondosa: A Review of Preclinical and Clinical Studies. Nutrients 2020, 12, 1339. [Google Scholar] [CrossRef]
- Al-Dbass, A.M.; Al-Daihan, S.K.; Bhat, R.S. Agaricus Blazei Murill as an Efficient Hepatoprotective and Antioxidant Agent against CCl4-Induced Liver Injury in Rats. Saudi J. Biol. Sci. 2012, 19, 303–309. [Google Scholar] [CrossRef] [PubMed]
- Zhao, Y.; Tian, N.; Wang, H.; Yan, H. Chemically Sulfated Polysaccharides from Agaricus Blazei Murill: Synthesis, Characterization and Anti-HIV Activity. Chem. Biodivers. 2021, 18, e2100338. [Google Scholar] [CrossRef] [PubMed]
- Gao, L.; Sun, Y.; Chen, C.; Xi, Y.; Wang, J.; Wang, Z. Primary Mechanism of Apoptosis Induction in a Leukemia Cell Line by Fraction FA-2-b-ß Prepared from the Mushroom Agaricus Blazei Murill. Braz. J. Med. Biol. Res. 2007, 40, 1545–1555. [Google Scholar] [CrossRef] [PubMed]
- Zhu, H.-H.; Wu, D.-P.; Jin, J.; Li, J.-Y.; Ma, J.; Wang, J.-X.; Jiang, H.; Chen, S.-J.; Huang, X.-J. Oral Tetra-Arsenic Tetra-Sulfide Formula Versus Intravenous Arsenic Trioxide As First-Line Treatment of Acute Promyelocytic Leukemia: A Multicenter Randomized Controlled Trial. J. Clin. Oncol. 2013, 31, 4215–4221. [Google Scholar] [CrossRef] [PubMed]
- Testi, A.M. GIMEMA-AIEOPAIDA Protocol for the Treatment of Newly Diagnosed Acute Promyelocytic Leukemia (APL) in Children. Blood 2005, 106, 447–453. [Google Scholar] [CrossRef] [PubMed]
- Love, W.K.; Tyson DeAngelis, J.; Berletch, J.B.; Phipps, S.M.O.; Andrews, L.G.; Brouillette, W.J.; Muccio, D.D.; Tollefsbol, T.O. The Novel Retinoid, 9cUAB30, Inhibits Telomerase and Induces Apoptosis in HL60 Cells. Transl. Oncol. 2008, 1, 148–152. [Google Scholar] [CrossRef] [PubMed]
- Seol, J.G.; Kim, E.S.; Park, W.H.; Jung, C.W.; Kim, B.K.; Lee, Y.Y. Telomerase Activity in Acute Myelogenous Leukaemia: Clinical and Biological Implications. Br. J. Haematol. 1998, 100, 156–165. [Google Scholar] [CrossRef] [PubMed]
- Hewison, M.; Barker, S.; Brennan, A.; Nathan, J.; Katz, D.R.; O’Riordan, J.L. Autocrine Regulation of 1,25-Dihydroxycholecalciferol Metabolism in Myelomonocytic Cells. Immunology 1989, 68, 247–252. [Google Scholar] [PubMed]
- Ferrero, D.; Campa, E.; Dellacasa, C.; Campana, S.; Foli, C.; Boccadoro, M. Differentiating Agents + Low-Dose Chemotherapy in the Management of Old/Poor Prognosis Patients with Acute Myeloid Leukemia or Myelodysplastic Syndrome. Haematologica 2004, 89, 619–620. [Google Scholar]
- Paubelle, E.; Zylbersztejn, F.; Alkhaeir, S.; Suarez, F.; Callens, C.; Dussiot, M.; Isnard, F.; Rubio, M.T.; Damaj, G.; Gorin, N.C.; et al. Deferasirox and Vitamin D Improves Overall Survival in Elderly Patients with Acute Myeloid Leukemia after Demethylating Agents Failure. PLoS ONE 2013, 8, e65998. [Google Scholar] [CrossRef]
- Shvachko, L.; Zavelevich, M.; Gluzman, D.; Telegeev, G. Vitamin E in Chronic Myeloid Leukemia (CML) Prevention. In Vitamin E in Health and Disease-Interactions, Diseases and Health Aspects; IntechOpen: London, UK, 2021. [Google Scholar]




{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}




Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Bartoszewska, E.; Molik, K.; Woźniak, M.; Choromańska, A. Telomerase Inhibition in the Treatment of Leukemia: A Comprehensive Review. Antioxidants 2024, 13, 427. https://doi.org/10.3390/antiox13040427
Bartoszewska E, Molik K, Woźniak M, Choromańska A. Telomerase Inhibition in the Treatment of Leukemia: A Comprehensive Review. Antioxidants. 2024; 13(4):427. https://doi.org/10.3390/antiox13040427
Chicago/Turabian StyleBartoszewska, Elżbieta, Klaudia Molik, Marta Woźniak, and Anna Choromańska. 2024. "Telomerase Inhibition in the Treatment of Leukemia: A Comprehensive Review" Antioxidants 13, no. 4: 427. https://doi.org/10.3390/antiox13040427
APA StyleBartoszewska, E., Molik, K., Woźniak, M., & Choromańska, A. (2024). Telomerase Inhibition in the Treatment of Leukemia: A Comprehensive Review. Antioxidants, 13(4), 427. https://doi.org/10.3390/antiox13040427