


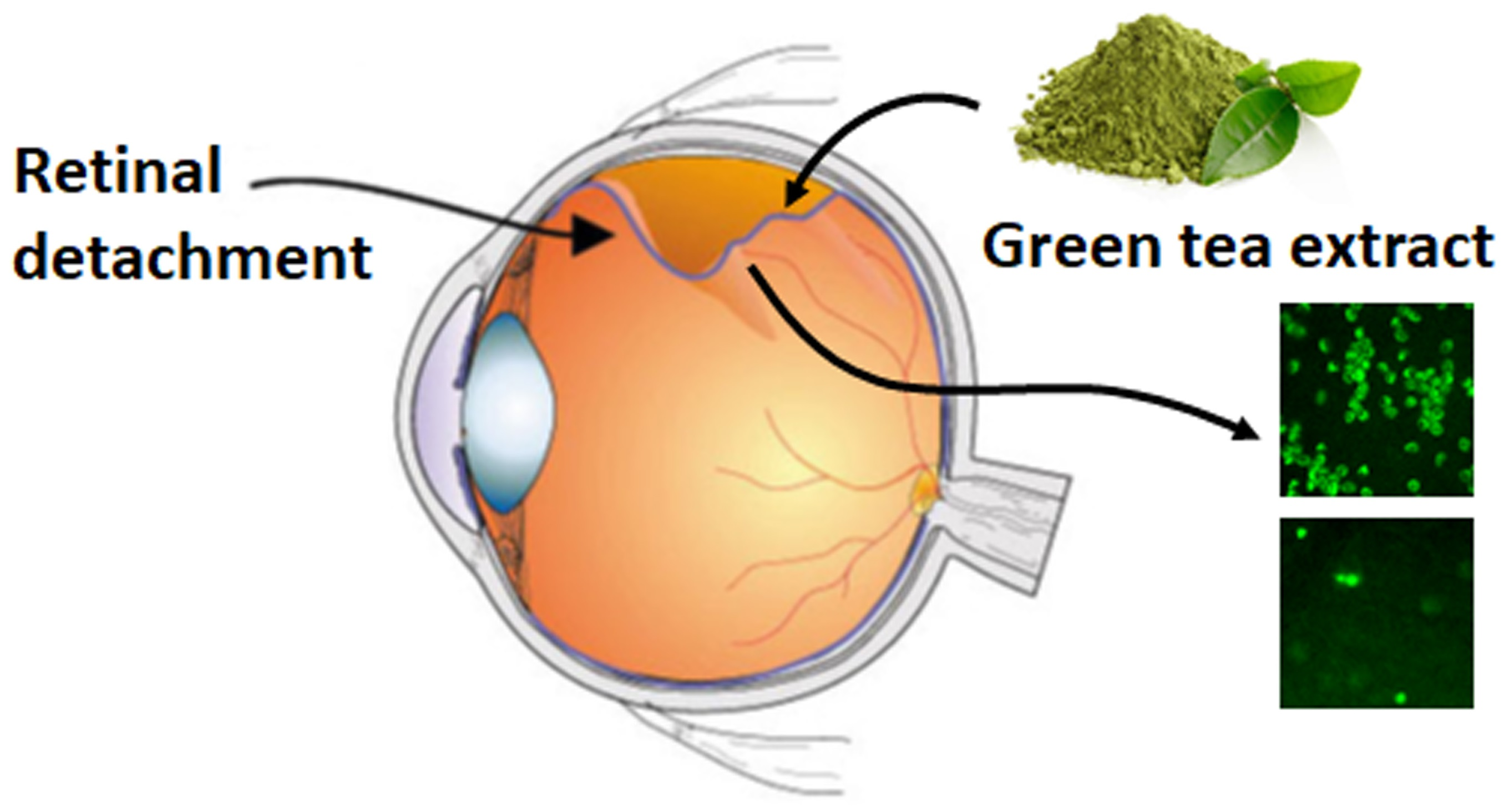
Amelioration of Functional, Metabolic, and Morphological Deterioration in the Retina following Retinal Detachment by Green Tea Extract


 ,
, 
Abstract

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
1. Introduction
2. Materials and Methods
2.1. Materials
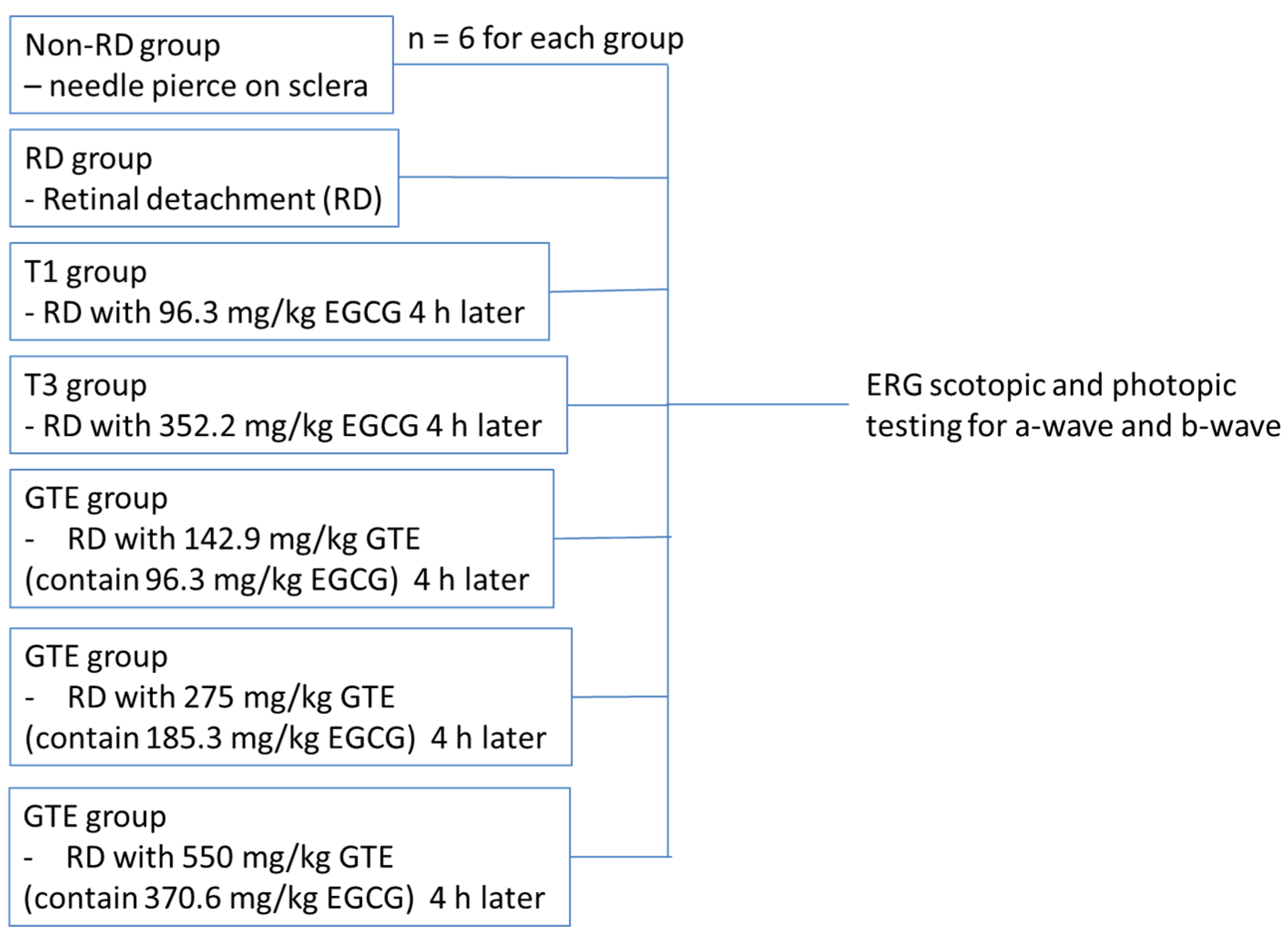
2.2. Animal Experiments
2.3. Retinal Detachment Induction
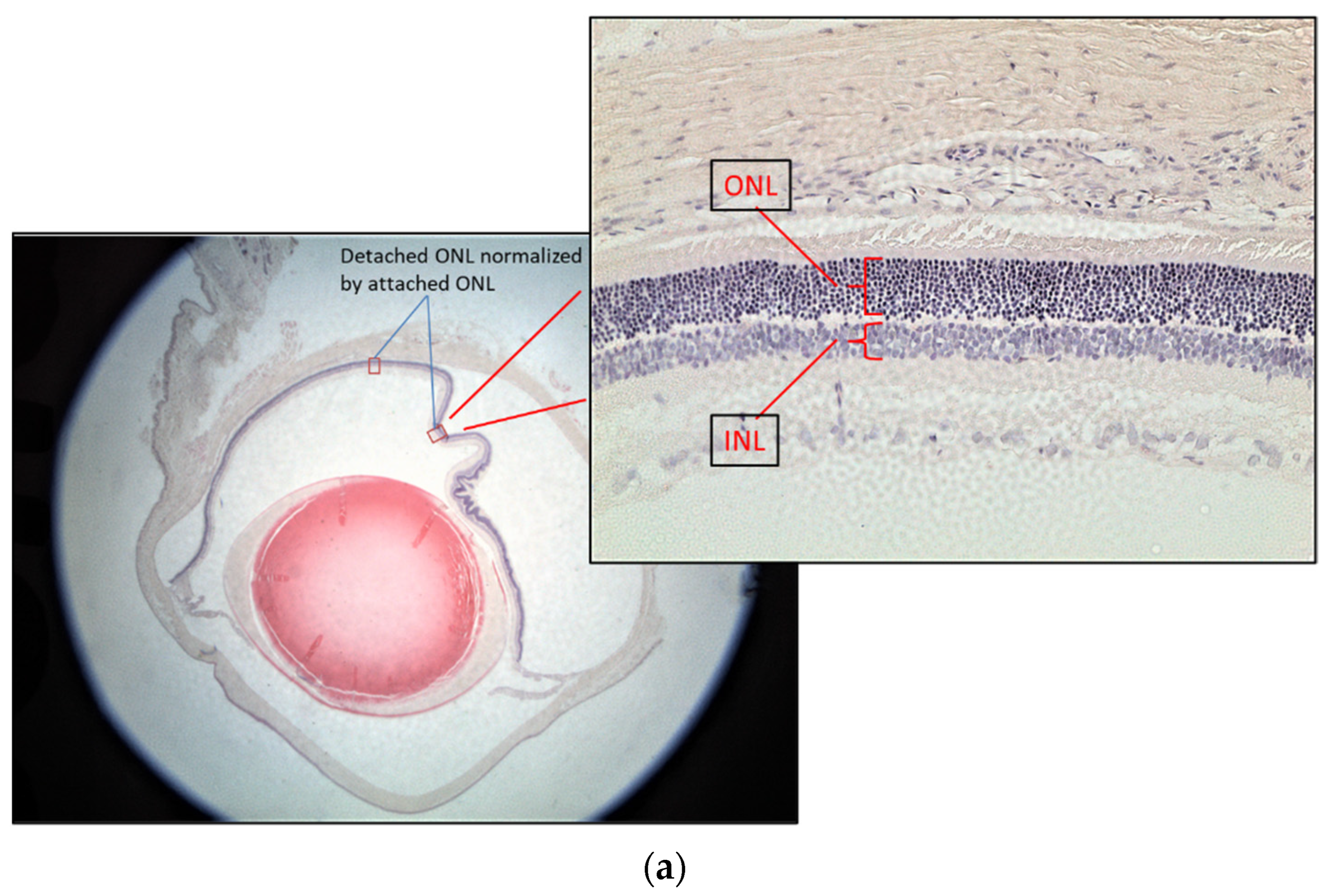
2.4. Histology and Retinal Ganglion Cell Layer Thickness Measurement
2.5. Electroretinography
2.6. TUNEL Staining
2.7. Caspase-3 Assay
2.8. Sphingomyelin Assay
2.9. Protein Carbonyl Content Assay
2.10. Hypoxia Inducible Factor 1 Alpha (HIF-α) Assay
2.11. Cytokine Array Analysis
2.12. Fatty Acids Profiling and Cholesterol Level in Retina
2.13. GC/MS Analysis
2.14. Statistical Analysis
3. Results
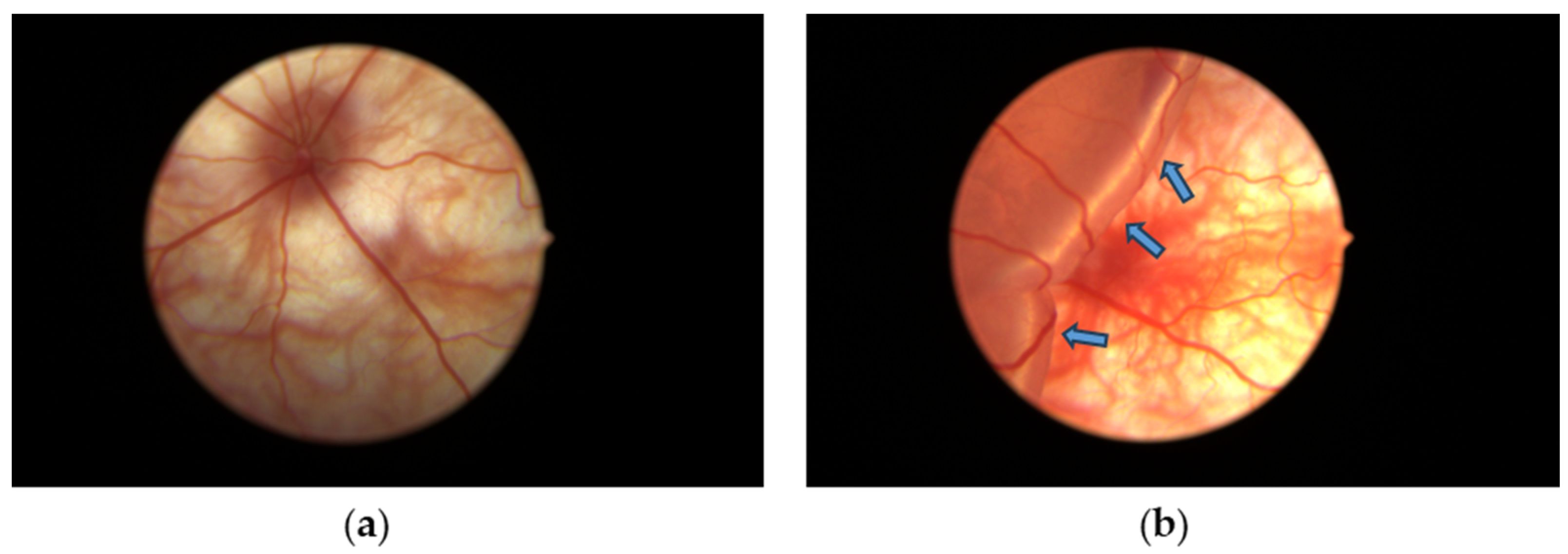
3.1. Retinal Detachment Features
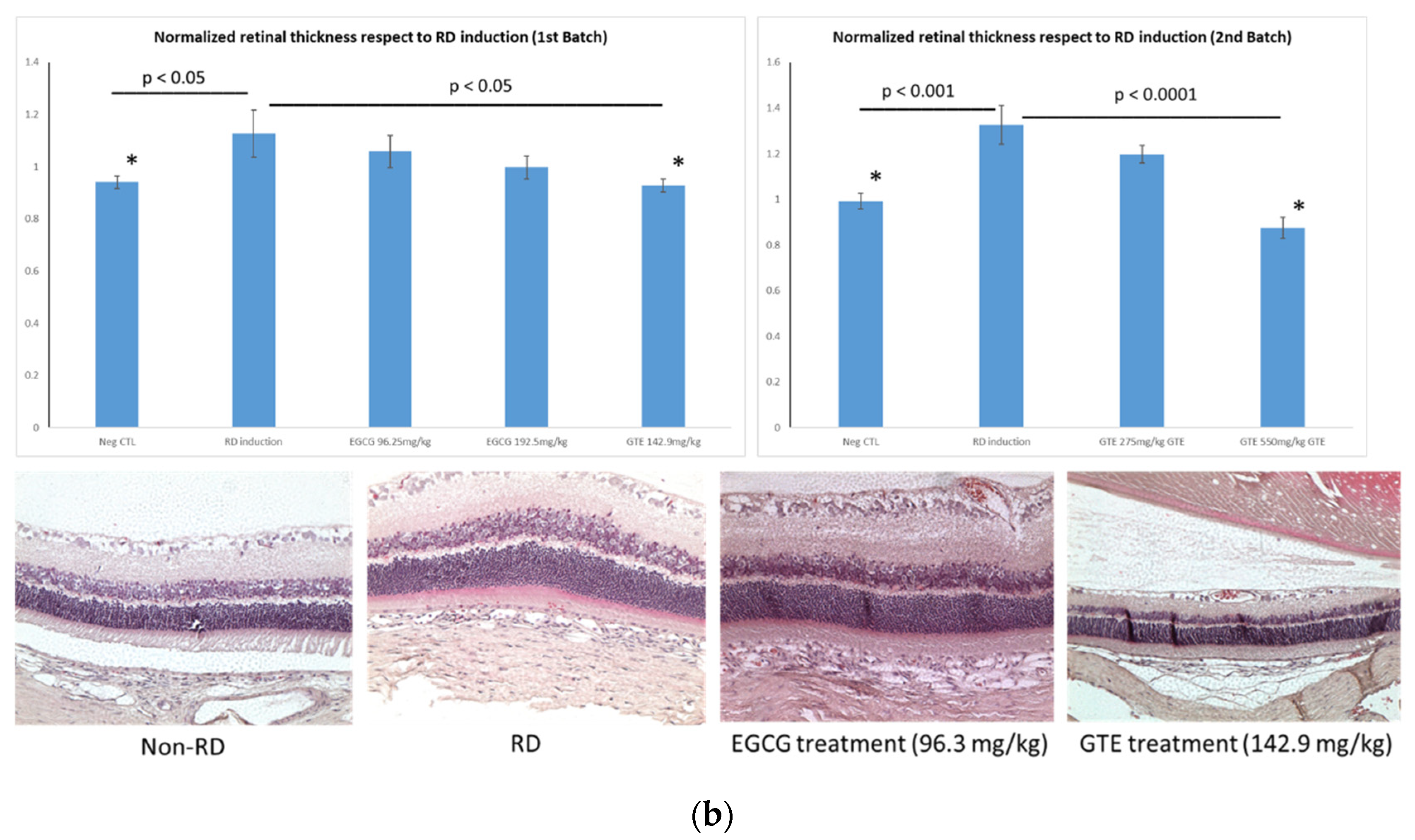
3.2. Comparison of Retina Thickness under Different Treatments
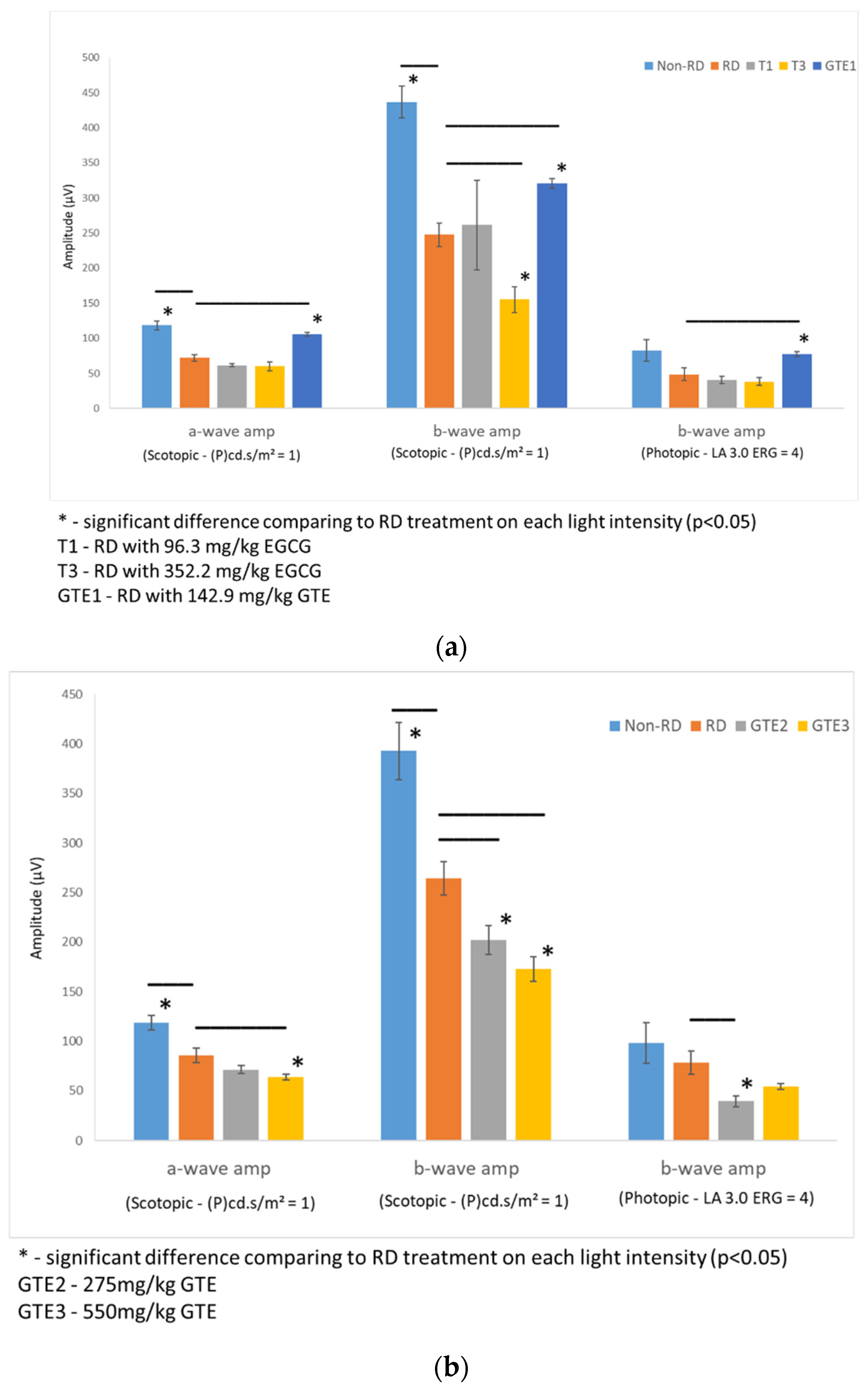
3.3. Comparison of Electroretinography Responses under Different Treatments
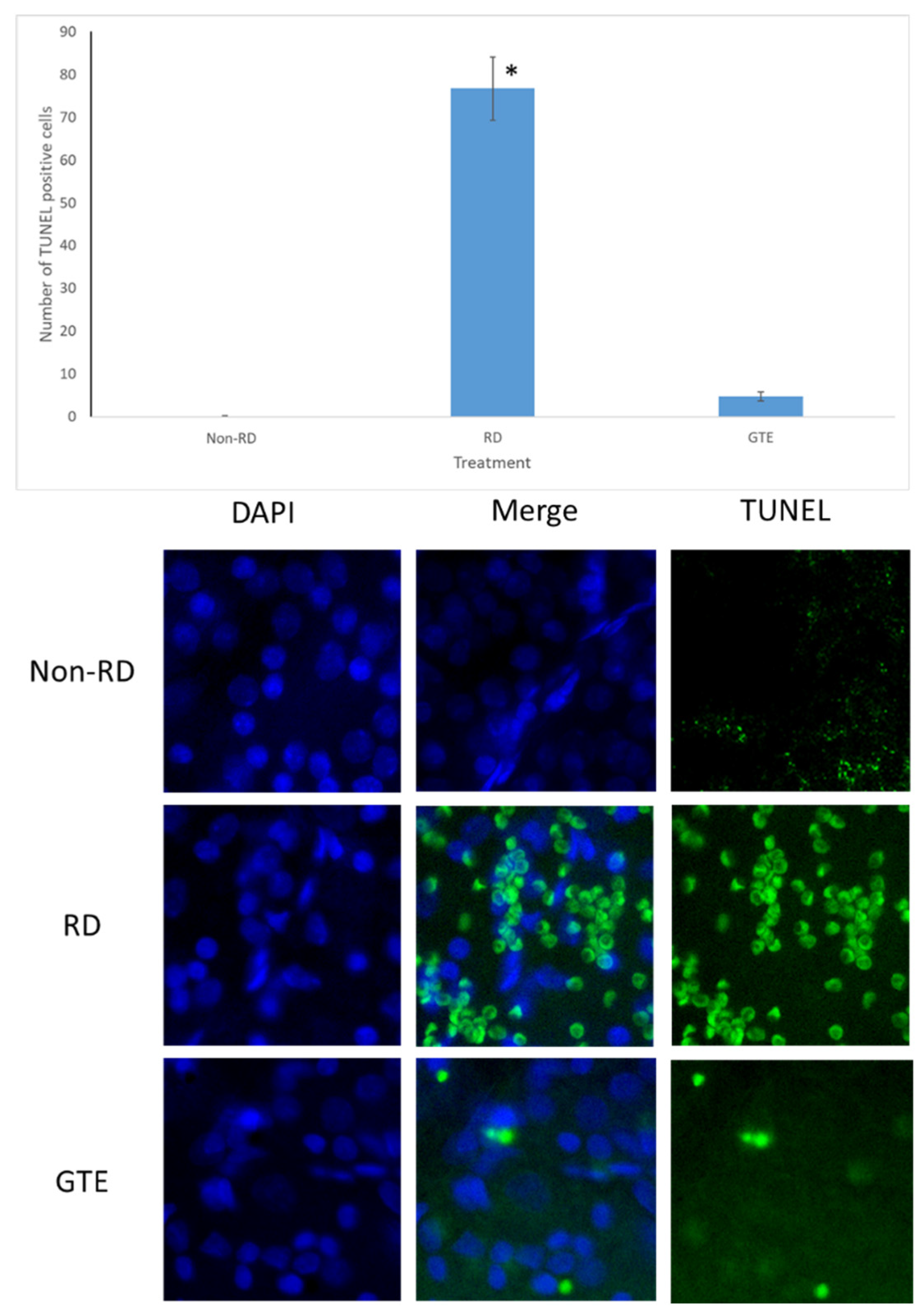
3.4. Comparison of the Number of Apoptosis Cells under Different Treatments by TUNEL Assay
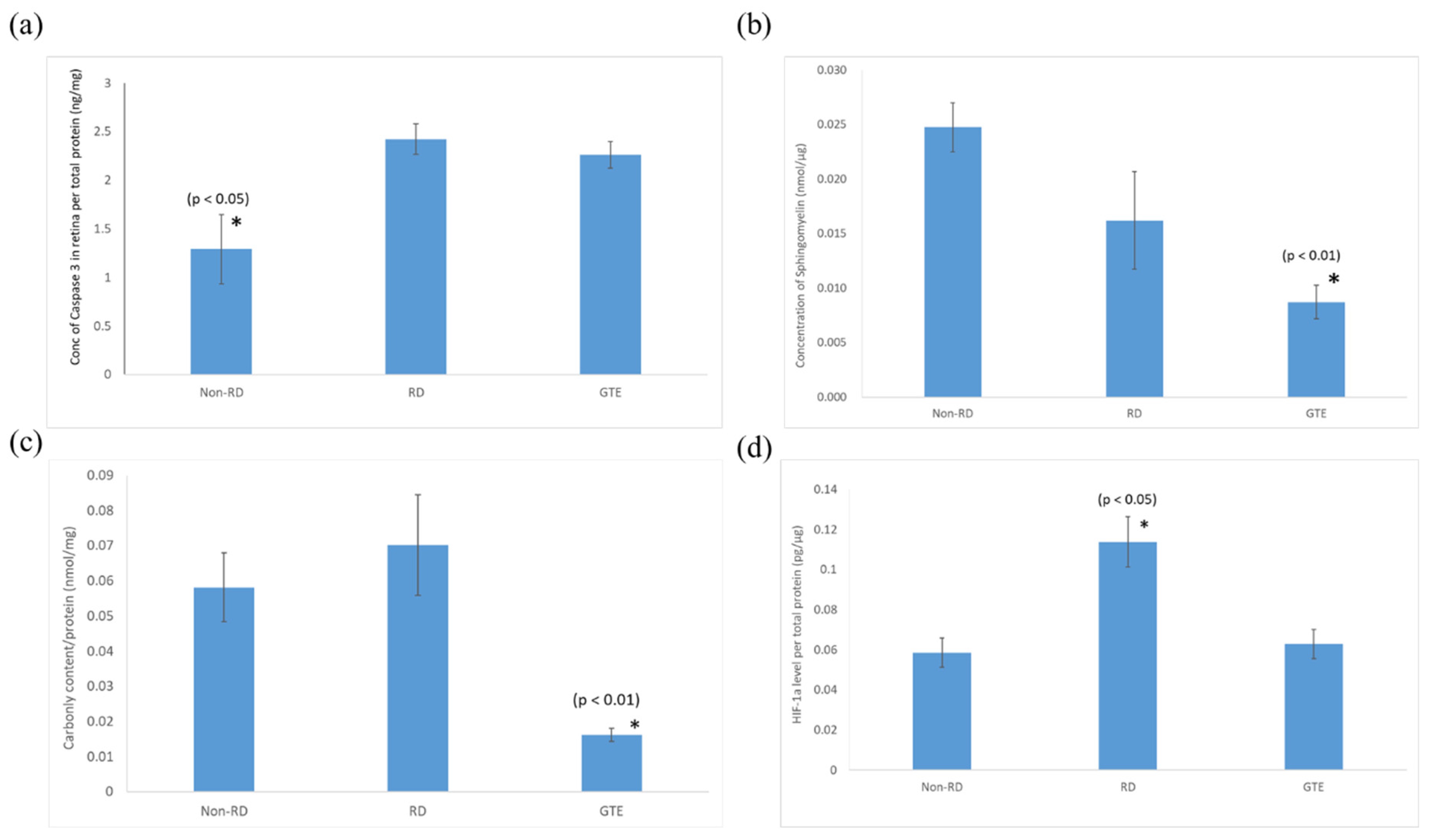
3.5. Comparison of the Expression of Pro-Apoptotic Caspase-3, Sphingomyelin, Protein Carbonyl, and HIF-1α Levels in the Retinae under Different Treatments
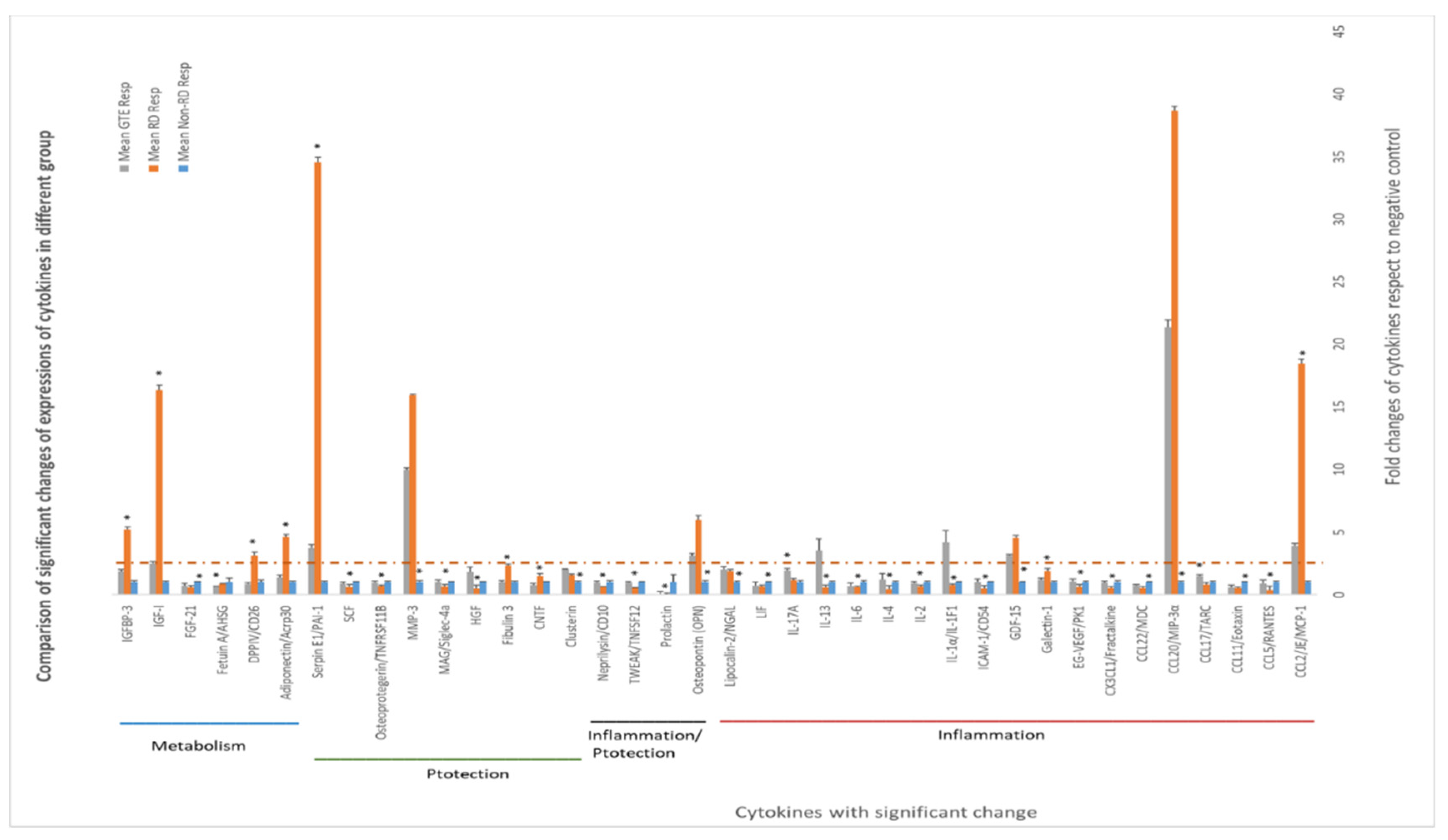
3.6. Comparing the Cytokines Proteome Profile in the Vitreous Humor under Different Treatments by Cytokine Array Analysis
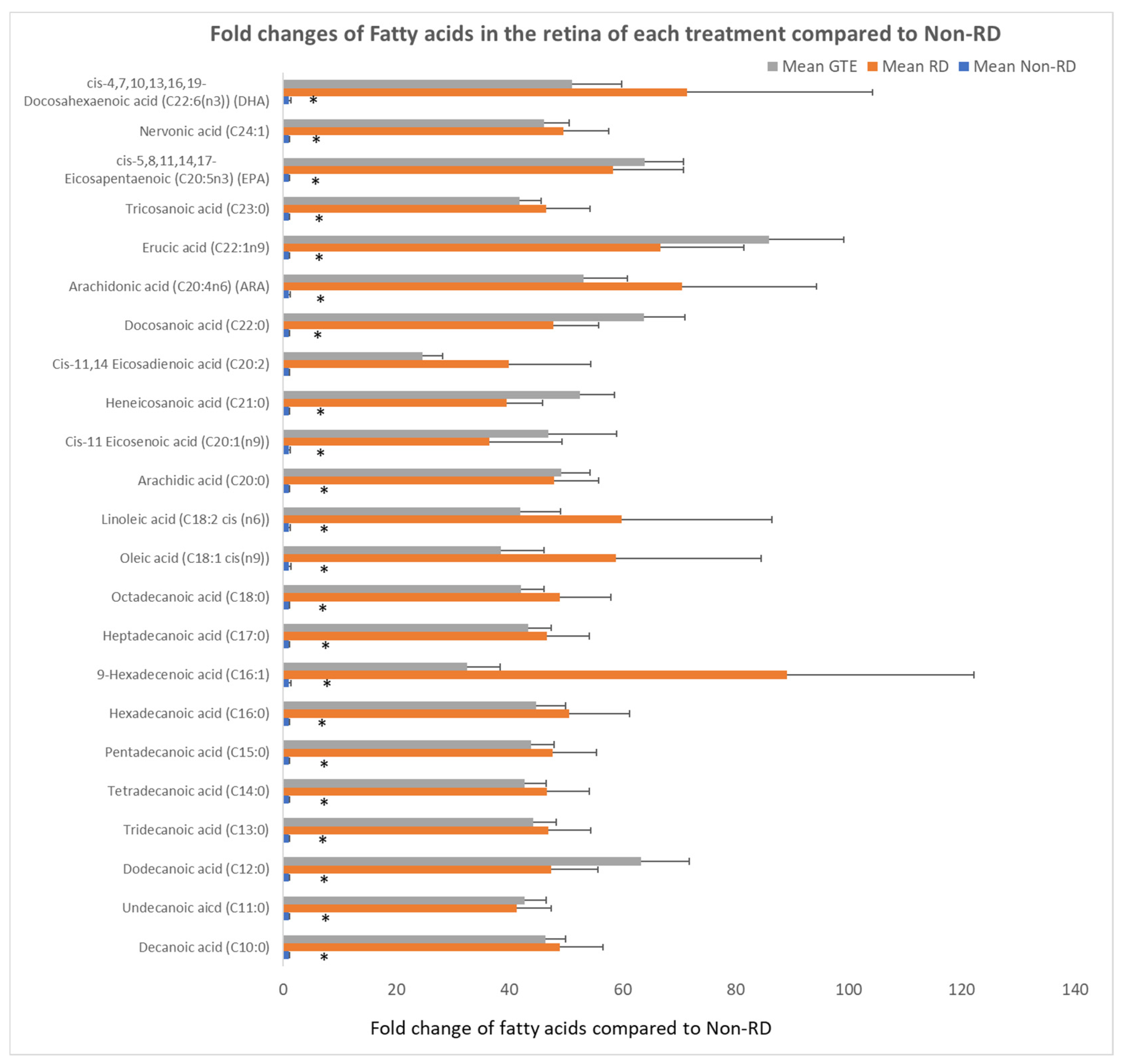
3.7. Comparing the Fatty Acids Profile in the Retinae under Different Treatments by GC/MS Analysis
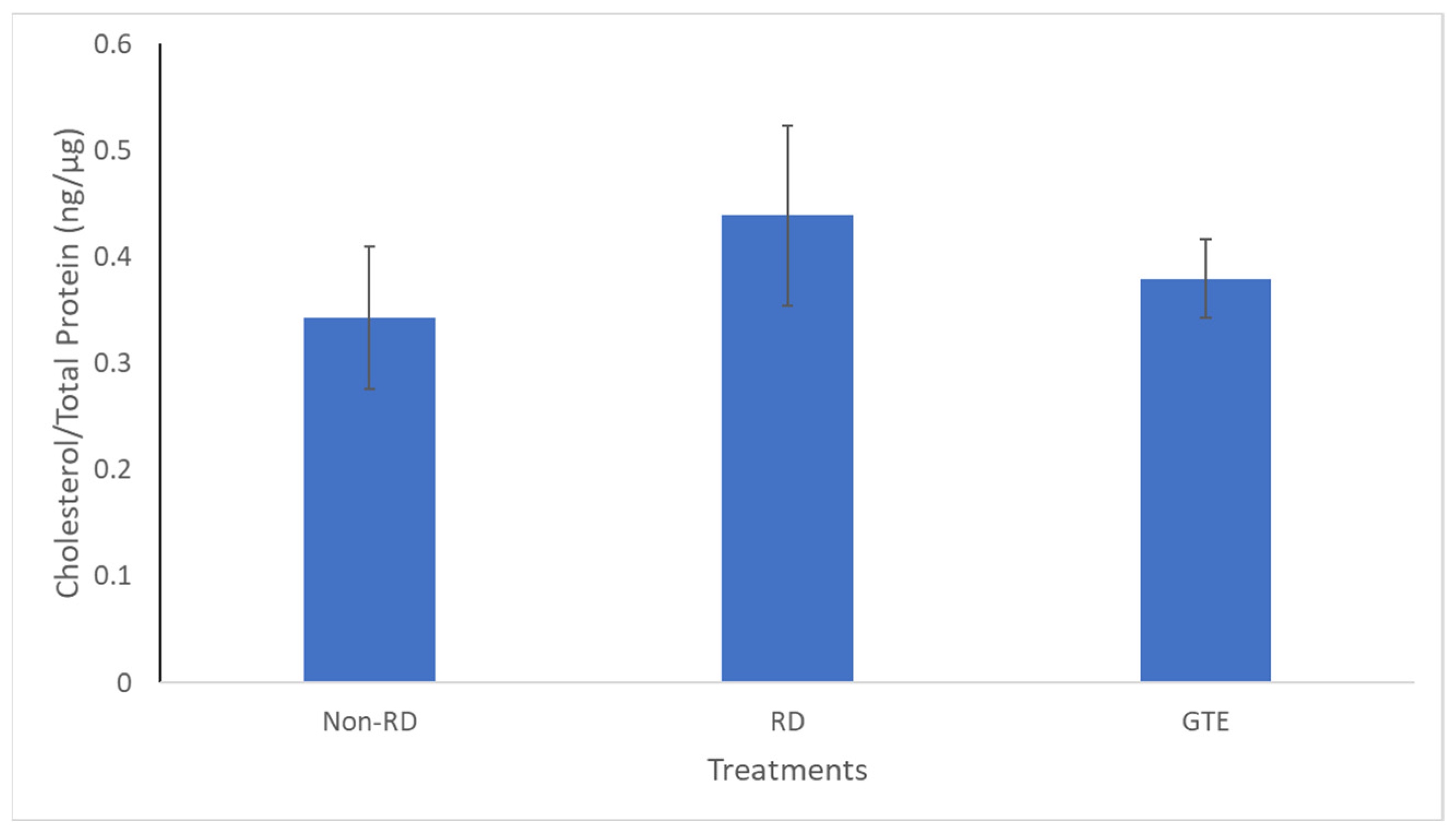
3.8. Comparing the Cholesterol Level in the Retinae under Different Treatments by GC/MS Analysis
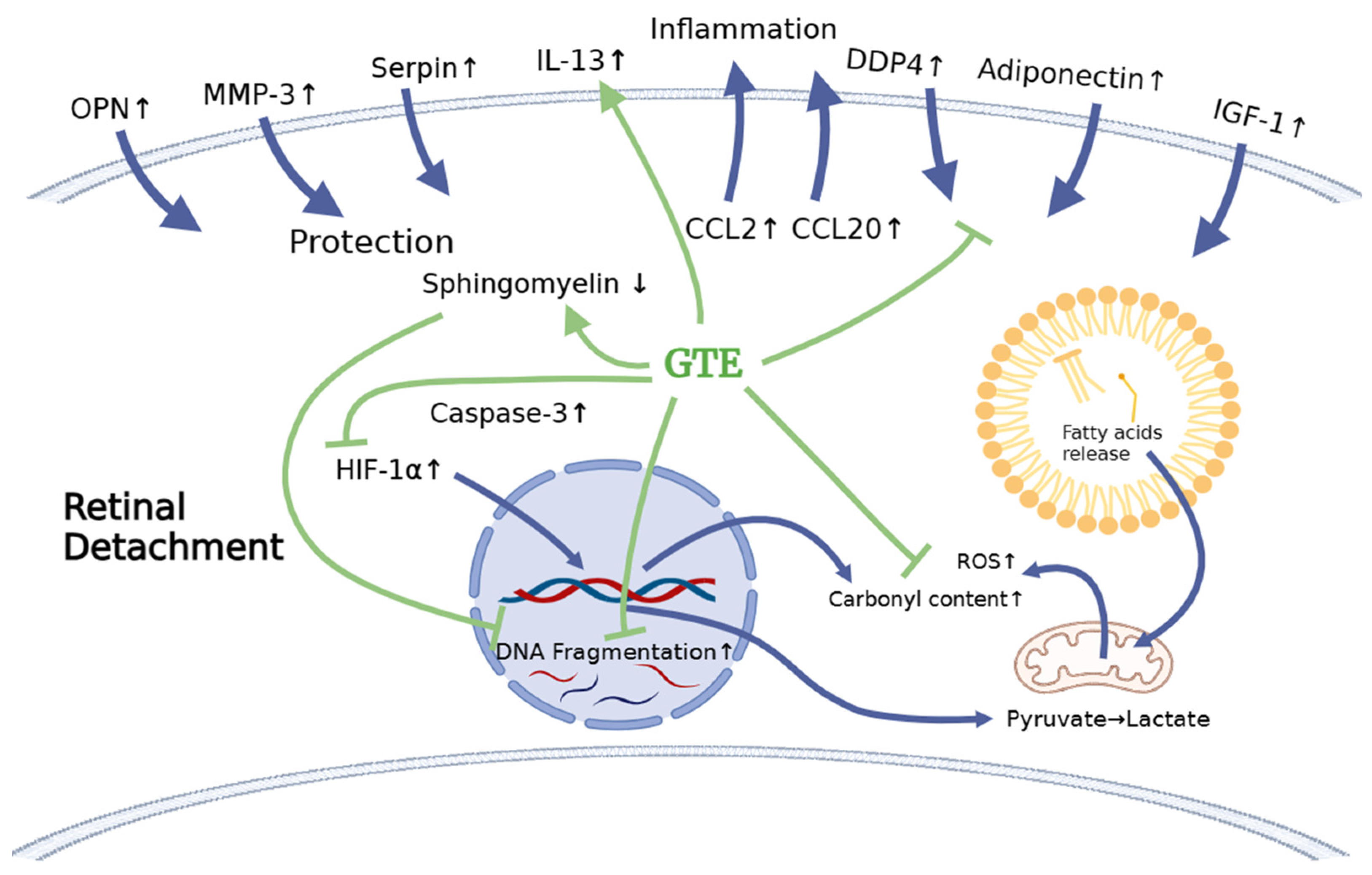
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Eijk, E.S.V.; Busschbach, J.J.V.; Timman, R.; Monteban, H.C.; Vissers, J.M.H.; Meurs, J.C.V. What made you wait so long? Delays in presentation of retinal detachment: Knowledge is related to an attached macula. Acta Ophthalmol. 2016, 94, 434–440. [Google Scholar] [CrossRef] [PubMed]
- Mowatt, L.; Shun-Shin, G.A.; Arora, S.; Price, N. Macula off retinal detachments. How long can they wait before it is too late? Eur. J. Ophthalmol. 2005, 15, 109–117. [Google Scholar] [CrossRef]
- Tang, E.W.H.; Wong, D.H.T.; Chan, Y.Y.Y.; Li, K.K.W. Emergency ophthalmic surgeries during COVID-19—A Hong Kong perspective. Graefes. Arch. Clin. Exp. Ophthalmol. 2020, 258, 2867–2868. [Google Scholar] [CrossRef] [PubMed]
- Grabowska, A.; Neffendorf, J.E.; Yorston, D.; Williamson, T.H. Urgency of retinal detachment repair: Is it time to re-think our priorities? Eye 2021, 35, 1035–1036. [Google Scholar] [CrossRef] [PubMed]
- Chu, K.O.; Chan, K.P.; Wang, C.C.; Chu, C.Y.; Li, W.Y.; Choy, K.W.; Rogers, M.S.; Pang, C.P. Green tea catechins and their oxidative protection in the rat eye. J. Agric. Food Chem. 2010, 58, 1523–1534. [Google Scholar] [CrossRef]
- Li, J.; Yip, Y.W.Y.; Ren, J.; Hui, W.K.; He, J.N.; Yu, Q.X.; Chu, K.O.; Ng, T.K.; Chan, S.O.; Pang, C.P.; et al. Green tea catechins alleviate autoimmune symptoms and visual impairment in a murine model for human chronic intraocular inflammation by inhibiting Th17-associated pro-inflammatory gene expression. Sci. Rep. 2019, 9, 2301. [Google Scholar] [CrossRef]
- Chu, K.O.; Chan, K.P.; Yip, Y.W.Y.; Chu, W.K.; Wang, C.C.; Pang, C.P. Systemic and Ocular Anti-Inflammatory Mechanisms of Green Tea Extract on Endotoxin-Induced Ocular Inflammation. Front. Endocrinol. 2022, 13, 899271. [Google Scholar] [CrossRef]
- Matsumoto, H.; Miller, J.W.; Vavas, D.G. Retinal Detachment Model in Rodents by Subretinal Injection of Sodium Hyaluronate. J. Vis. Exp. 2013, 79, 50660. [Google Scholar]
- Zadro-Lamoureux, L.A.; Zacks, D.N.; Baker, A.N.; Zheng, Q.D.; Hauswirth, W.W.; Tsilfidis, C. Effects on XIAP Retinal Detachment–Induced Photoreceptor Apoptosis. Investig. Ophthalmol. Vis. Sci. 2009, 50, 1448–1453. [Google Scholar] [CrossRef][Green Version]
- Dong, K.; Zhu, H.; Song, Z.; Gong, Y.; Wang, F.; Wang, W.; Zheng, Z.; Yu, Z.; Gu, Q.; Xu, X.; et al. Necrostatin-1 protects photoreceptors from cell death and improves functional outcome after experimental retinal detachment. Am. J. Pathol. 2012, 181, 1634–1641. [Google Scholar] [CrossRef]
- Xie, Z.; Chen, F.; Wu, X.; Zhuang, C.; Zhu, J.; Wang, J.; Ji, H.; Wang, Y.; Hua, X. Safety and efficacy of intravitreal injection of recombinant erythropoietin for protection of photoreceptor cells in a rat model of retinal detachment. Eye 2012, 26, 144–152. [Google Scholar] [CrossRef][Green Version]
- Natalia, M.; Stepnowski, P.; Paszkiewicz, M.; Gołebiowski, M.; Kumirska, J. Trimethylsilyldiazomethane (TMSD) as a new derivatization reagent for trace analysis of selected non-steroidal anti-inflammatory drugs (NSAIDs) by gas chromatography methods. Anal. Bioanal. Chem. 2010, 397, 3029–3034. [Google Scholar]
- Topolewska, A.; Czarnowska, K.; Haliński, Ł.P.; Stepnowski, P. Evaluation of four derivatization methods for the analysis of fatty acids from green leafy vegetables by gas chromatography. J. Chromatogr. B Analyt. Technol. Biomed. Life Sci. 2015, 990, 150–157. [Google Scholar] [CrossRef]
- Saleem, M.A.; Benitez, B.; Yaros, C.; Yamar, G.; Bhattacharya, S.K. Analysis of Cholesterol from the Liver Using Gas Chromatography-Mass Spectrometry. Methods Mol. Biol. 2023, 2571, 143–148. [Google Scholar] [PubMed]
- Baranowska, M.; Suliborska, K.; Chrzanowski, W.; Kusznierewicz, B.; Namieśnik, J.; Bartoszek, A. The relationship between standard reduction potentials of catechins and biological activities involved in redox control. Redox. Biol. 2018, 17, 355–366. [Google Scholar] [CrossRef] [PubMed]
- Byun, E.B.; Kim, W.S.; Sung, N.Y.; Byun, E.H. Epigallocatechin-3-Gallate Regulates Anti-Inflammatory Action Through 67-kDa Laminin Receptor-Mediated Tollip Signaling Induction in Lipopolysaccharide-Stimulated Human Intestinal Epithelial Cells. Cell Physiol. Biochem. 2018, 46, 2072–2081. [Google Scholar] [CrossRef] [PubMed]
- Chu, K.O.; Chan, K.P.; Yang, Y.; Qin, Y.J.; Li, W.Y.; Chan, S.O.; Wang, C.C.; Pang, C.P. Effects of EGCG content in green tea extract on pharmacokinetics, oxidative status and expression of inflammatory and apoptotic genes in the rat ocular tissues. J. Nutr. Biochem. 2015, 26, 1357–1367. [Google Scholar] [CrossRef] [PubMed]
- Hong, J.; Lambert, J.D.; Lee, S.H.; Sinko, P.J.; Yang, C.S. Involvement of multidrug resistance-associated proteins in regulating cellular levels of (-)-epigallocatechin-3-gallate and its methyl metabolites. Biochem. Biophys. Res. Commun. 2003, 310, 222–227. [Google Scholar] [CrossRef]
- Chen, F.; Jiang, L.; Shen, C.; Wan, H.; Xu, L.; Wang, N.; Jonas, J.B. Neuroprotective effect of epigallocatechin-3-gallate against N-methyl-D-aspartate-induced excitotoxicity in the adult rat retina. Acta Ophthalmol. 2012, 90, e609–e615. [Google Scholar] [CrossRef]
- Qin, Y.J.; Chu, K.O.; Yip, Y.W.; Li, W.Y.; Yang, Y.P.; Chan, K.P.; Ren, J.L.; Chan, S.O.; Pang, C.P. Green tea extract treatment alleviates ocular inflammation in a rat model of endotoxin-induced uveitis. PLoS ONE 2014, 9, e103995. [Google Scholar] [CrossRef]
- Yang, Y.; Qin, Y.J.; Yip, Y.W.; Chan, K.P.; Chu, K.O.; Chu, W.K.; Ng, T.K.; Pang, C.P.; Chan, S.O. Green tea catechins are potent anti-oxidants that ameliorate sodium iodate-induced retinal degeneration in rats. Sci. Rep. 2016, 6, 29546. [Google Scholar] [CrossRef] [PubMed]
- Xie, J.; Jiang, L.; Zhang, T.; Jin, Y.; Yang, D.; Chen, F. Neuroprotective effects of Epigallocatechin-3-gallate (EGCG) in optic nerve crush model in rats. Neurosci. Lett. 2010, 479, 26–30. [Google Scholar] [CrossRef] [PubMed]
- Linsenmeier, R.A.; Padnick-Silver, L. Metabolic dependence of photoreceptors on the choroid in the normal and detached retina. Investig. Ophthalmol. Vis. Sci. 2000, 41, 3117–3123. [Google Scholar]
- Bhatt, L.; Groeger, G.; McDermott, K.; Cotter, T.G. Rod and cone photoreceptor cells produce ROS in response to stress in a live retinal explant system. Mol. Vis. 2010, 16, 283–293. [Google Scholar]
- Chu, K.O.; Chan, K.P.; Chan, S.O.; Ng, T.K.; Jhanji, V.; Wang, C.C.; Pang, C.P. Metabolomics of Green-Tea Catechins on Vascular-Endothelial-Growth-Factor-Stimulated Human-Endothelial-Cell Survival. J. Agric Food Chem. 2018, 66, 12866–12875. [Google Scholar] [CrossRef] [PubMed]
- Yang, C.S.; Lambert, J.D.; Ju, J.; Lu, G.; Sang, S. Tea and cancer prevention: Molecular mechanisms and human relevance. Toxicol. Appl. Pharmacol. 2007, 224, 265–273. [Google Scholar] [CrossRef]
- Woo, T.T.Y.; Li, S.Y.; Lai, W.W.K.; Wong, D.; Lo, A.C.Y. Neuroprotective effects of lutein in a rat model of retinal detachment. Graefes Arch. Clin. Exp. Ophthalmol. 2013, 251, 41–51. [Google Scholar] [CrossRef]
- Ding, J.; Yang, N.; Yan, Y.; Wang, Y.; Wang, X.; Lu, L.; Dong, K. Rapamycin Inhibited Photoreceptor Necroptosis and Protected the Retina by Activation of Autophagy in Experimental Retinal Detachment. Curr. Eye Res. 2019, 44, 739–745. [Google Scholar] [CrossRef]
- Khalil, H.; Peltzer, N.; Walicki, J.; Yang, J.Y.; Dubuis, G.; Gardiol, N.; Held, W.; Bigliardi, P.; Marsland, B.; Liaudet, L.; et al. Caspase-3 protects stressed organs against cell death. Mol. Cell. Biol. 2012, 32, 4523–4533. [Google Scholar] [CrossRef]
- Porta, C.C.; Paglino, M.A. Targeting PI3K/Akt/mTOR signaling in cancer. Front. Oncol. 2014, 4, 64. [Google Scholar] [CrossRef]
- Hussain, A.R.; Ahmed, S.O.; Ahmed, M.; Khan, O.S.; Al Abdul Mohsen, S.; Platanias, L.C.; Al-Kuraya, K.S.; Uddin, S. Cross-talk between NFkB and the PI3-kinase/AKT pathway can Be targeted in primary effusion lymphoma (PEL) cell lines for efficient apoptosis. PLoS ONE 2012, 7, e39945. [Google Scholar] [CrossRef]
- Uchiyama, T.M.; Richard, E.; Nilanjana, M.; Das Dipak, K. Role of Akt signaling in mitochondrial survival pathway triggered by hypoxic preconditioning. Circulation 2004, 109, 3042–3049. [Google Scholar] [CrossRef]
- Weidemann, A.; Johnson, R.S. Biology of HIF-1alpha. Cell Death Differ. 2008, 15, 621–627. [Google Scholar] [CrossRef]
- Zhang, Q.; Tang, X.; Lu, Q.; Zhang, Z.; Rao, J.; D Le, A. Green tea extract and (-)-epigallocatechin-3-gallate inhibit hypoxia- and serum-induced HIF-1alpha protein accumulation and VEGF expression in human cervical carcinoma and hepatoma cells. Mol. Cancer Ther. 2006, 5, 1227–1238. [Google Scholar] [CrossRef]
- Taniguchi, M.; Okazaki, T. The role of sphingomyelin and sphingomyelin synthases in cell death, proliferation and migration-from cell and animal models to human disorders. Biochim. Biophys. Acta. 2014, 1841, 692–703. [Google Scholar] [CrossRef]
- van Blitterswijk, W.J.; Klarenbeek, J.B.; van der Luit, A.H.; Alderliesten, M.C.; van Lummel, M.; Verheij, M. Fas/CD95 down-regulation in lymphoma cells through acquired alkyllysophospholipid resistance: Partial role of associated sphingomyelin deficiency. Biochem. J. 2009, 425, 225–234. [Google Scholar] [CrossRef][Green Version]
- Gschwandtner, M.; Derler, R.; Midwood, K.S. More Than Just Attractive: How CCL2 Influences Myeloid Cell Behavior Beyond Chemotaxis. Front. Immunol. 2019, 10, 2759. [Google Scholar] [CrossRef]
- Bellak, J.M.; Hanson, K.; Busse, W.W.; Gern, J.E. MIP-3alpha/CCL20 expression is highly induced in respiratory epithelium and in human subjects infected with rhinoviruses. J. Allergy Clinic. Immunol. 2004, 113, S267. [Google Scholar] [CrossRef]
- Wischhusen, J.; Melero, I.; Fridman, W.H. Growth/Differentiation Factor-15 (GDF-15): From Biomarker to Novel Targetable Immune Checkpoint. Front. Immunol. 2020, 11, 951. [Google Scholar] [CrossRef] [PubMed]
- Da Silva, A.P.; Pollett, A.; Rittling, S.R.; Denhardt, D.T.; Sodek, J.; Zohar, R. Exacerbated tissue destruction in DSS-induced acute colitis of OPN-null mice is associated with downregulation of TNF-alpha expression and non-programmed cell death. J. Cell. Physiol. 2006, 208, 629–639. [Google Scholar] [CrossRef] [PubMed]
- Van Hove, I.; Lemmens, K.; Van de Velde, S.; Verslegers, M.; Moons, L. Matrix metalloproteinase-3 in the central nervous system: A look on the bright side. J. Neurochem. 2012, 123, 203–216. [Google Scholar] [CrossRef]
- Sanrattana, W.; Maas, C.; de Maat, S. SERPINs-From Trap to Treatment. Front. Med. 2019, 6, 25. [Google Scholar] [CrossRef]
- Chandran, M.; Phillips, S.A.; Ciaraldi, T.; Henry, R.R. Adiponectin: More than just another fat cell hormone? Diabetes Care 2003, 26, 2442–2450. [Google Scholar] [CrossRef]
- Deacon, C.F. Physiology and Pharmacology of DPP-4 in Glucose Homeostasis and the Treatment of Type 2 Diabetes. Front. Endocrinol. 2019, 10, 80. [Google Scholar] [CrossRef]
- Clemmons, D.R. Metabolic Actions of IGF-I in Normal Physiology and Diabetes. Endocrinol. Metab. Clin. N. Am. 2012, 41, 425–443. [Google Scholar] [CrossRef]
- Shrivastav, S.V.; Bhardwaj, A.; Pathak, K.A.; Shrivastav, A. Insulin-Like Growth Factor Binding Protein-3 (IGFBP-3): Unraveling the Role in Mediating IGF-Independent Effects Within the Cell. Front. Cell. Dev. Biol. 2020, 8, 286. [Google Scholar] [CrossRef]
- Incardona, J.P.; Eaton, S. Cholesterol in signal transduction. Curr. Opin. Cell Biol. 2000, 12, 193–203. [Google Scholar] [CrossRef] [PubMed]
- Steinhauser, M.L.; Olenchock, B.A.; O'Keefe, J.; Lun, M.; Pierce, K.A.; Lee, H.; Pantano, L.; Klibanski, A.; Shulman, G.I.; Clish, C.B.; et al. The circulating metabolome of human starvation. JCI Insight 2018, 3, e121434. [Google Scholar] [CrossRef] [PubMed]
- Kolosowska, N.; Keuters, M.H.; Wojciechowski, S.; Keksa-Goldsteine, V.; Laine, M.; Malm, T.; Goldsteins, G.; Koistinaho, J.; Dhungana, H. Peripheral Administration of IL-13 Induces Anti-inflammatory Microglial/Macrophage Responses and Provides Neuroprotection in Ischemic Stroke. Neurotherapeutics. 2019, 16, 1304–1319. [Google Scholar] [CrossRef] [PubMed]
- Takahashi, S.; Nakamura, E.; Okabe, S. Effects of cytokines, without and with Helicobacter pylori components, on mucus secretion by cultured gastric epithelial cells. Dig. Dis. Sci. 1998, 43, 2301–2308. [Google Scholar] [CrossRef] [PubMed]
- Al-Sadi, R.; Ye, D.; Dokladny, K.; Ma, T.Y. Mechanism of IL-1beta-induced increase in intestinal epithelial tight junction permeability. J. Immunol. 2008, 180, 5653–5661. [Google Scholar] [CrossRef] [PubMed]
- Al-Sadi, R.; Ye, D.; Said, H.M.; Ma, T.Y. Cellular and molecular mechanism of interleukin-1β modulation of Caco-2 intestinal epithelial tight junction barrier. J. Cell. Mol. Med. 2011, 15, 970–982. [Google Scholar] [CrossRef] [PubMed]
- Mohamed, E.; Udata, C.; Liao, K.; Yin, D. Considerations on the Calculation of the Human Equivalent Dose from Toxicology Studies for Biologic Anticancer Agents. Clin. Pharmacokinet. 2021, 60, 563–567. [Google Scholar]
- Shanafelt, T.D.; Call, T.G.; Zent, C.S.; Leis, J.F.; LaPlant, B.; Bowen, D.A.; Roos, M.; Laumann, K.; Ghosh, A.K.; Lesnick, C.; et al. Phase 2 Trial of Daily, Oral Polyphenon E in Patients with Asymptomatic, Rai Stage 0-II Chronic Lymphocytic Leukemia (CLL). Cancer 2013, 119, 363–370. [Google Scholar] [CrossRef]











Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Chu, K.O.; Yip, Y.W.Y.; Chan, K.P.; Wang, C.C.; Ng, D.S.C.; Pang, C.P. Amelioration of Functional, Metabolic, and Morphological Deterioration in the Retina following Retinal Detachment by Green Tea Extract. Antioxidants 2024, 13, 235. https://doi.org/10.3390/antiox13020235
Chu KO, Yip YWY, Chan KP, Wang CC, Ng DSC, Pang CP. Amelioration of Functional, Metabolic, and Morphological Deterioration in the Retina following Retinal Detachment by Green Tea Extract. Antioxidants. 2024; 13(2):235. https://doi.org/10.3390/antiox13020235
Chicago/Turabian StyleChu, Kai On, Yolanda Wong Ying Yip, Kwok Ping Chan, Chi Chiu Wang, Danny Siu Chun Ng, and Chi Pui Pang. 2024. "Amelioration of Functional, Metabolic, and Morphological Deterioration in the Retina following Retinal Detachment by Green Tea Extract" Antioxidants 13, no. 2: 235. https://doi.org/10.3390/antiox13020235
APA StyleChu, K. O., Yip, Y. W. Y., Chan, K. P., Wang, C. C., Ng, D. S. C., & Pang, C. P. (2024). Amelioration of Functional, Metabolic, and Morphological Deterioration in the Retina following Retinal Detachment by Green Tea Extract. Antioxidants, 13(2), 235. https://doi.org/10.3390/antiox13020235