
Silymarin and Inflammation: Food for Thoughts



Abstract
1. Introduction
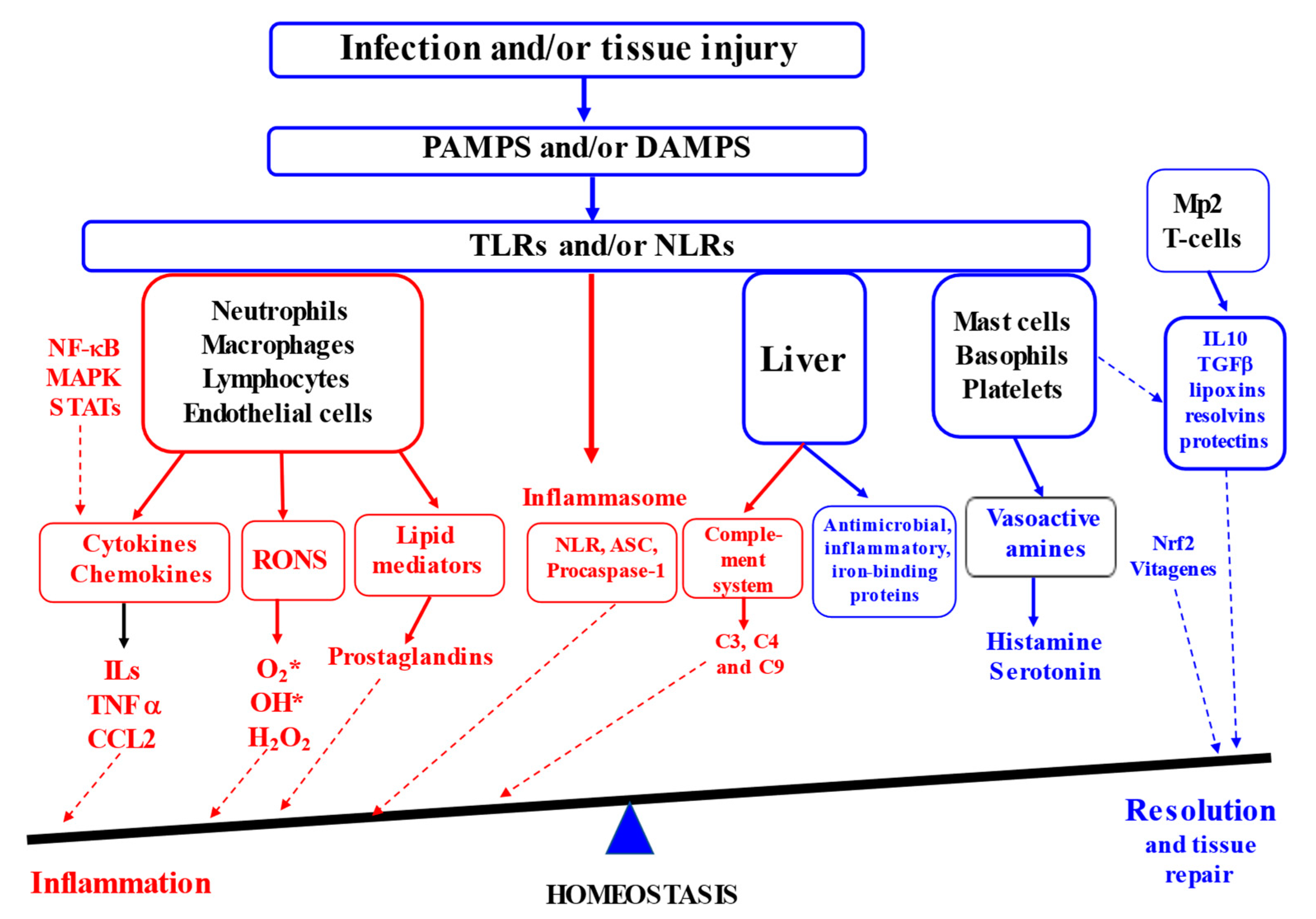
2. Molecular Mechanisms of Inflammation
3. Oxidative Stress and Inflammation
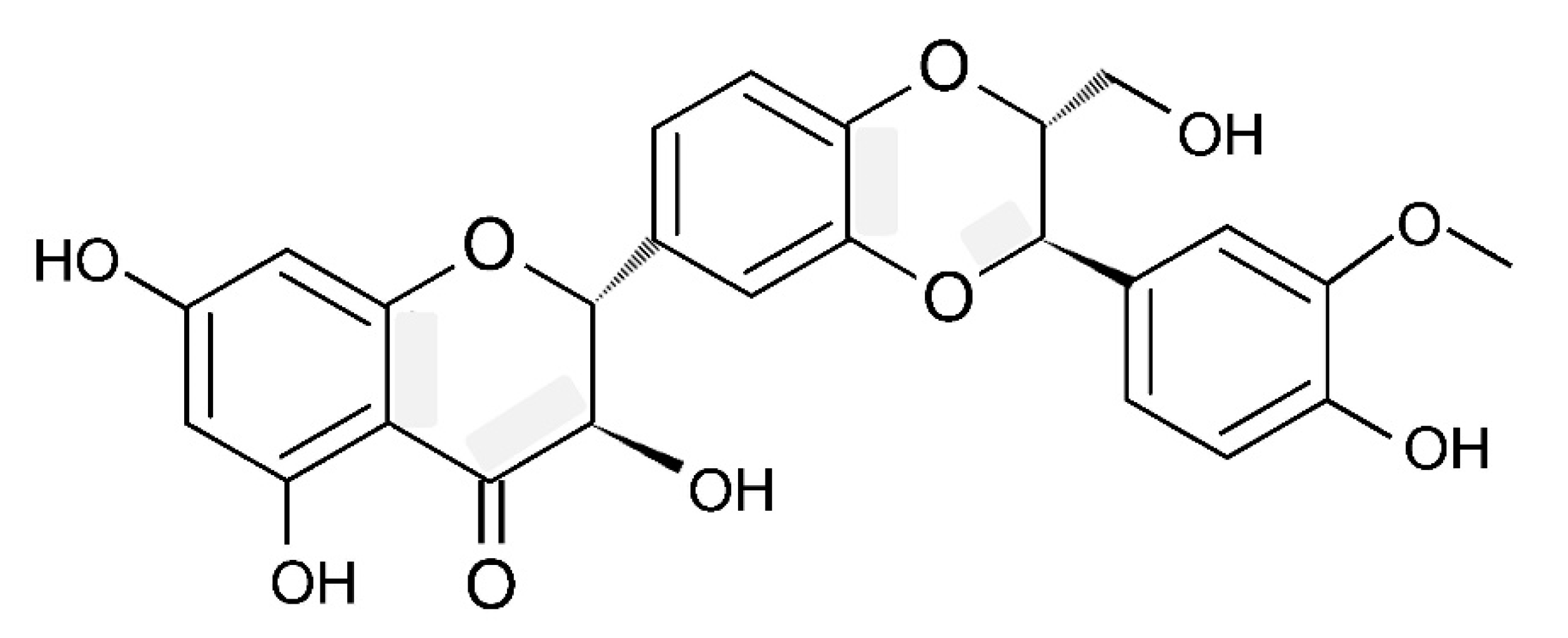
4. Chemical Composition and Structure of Silymarin
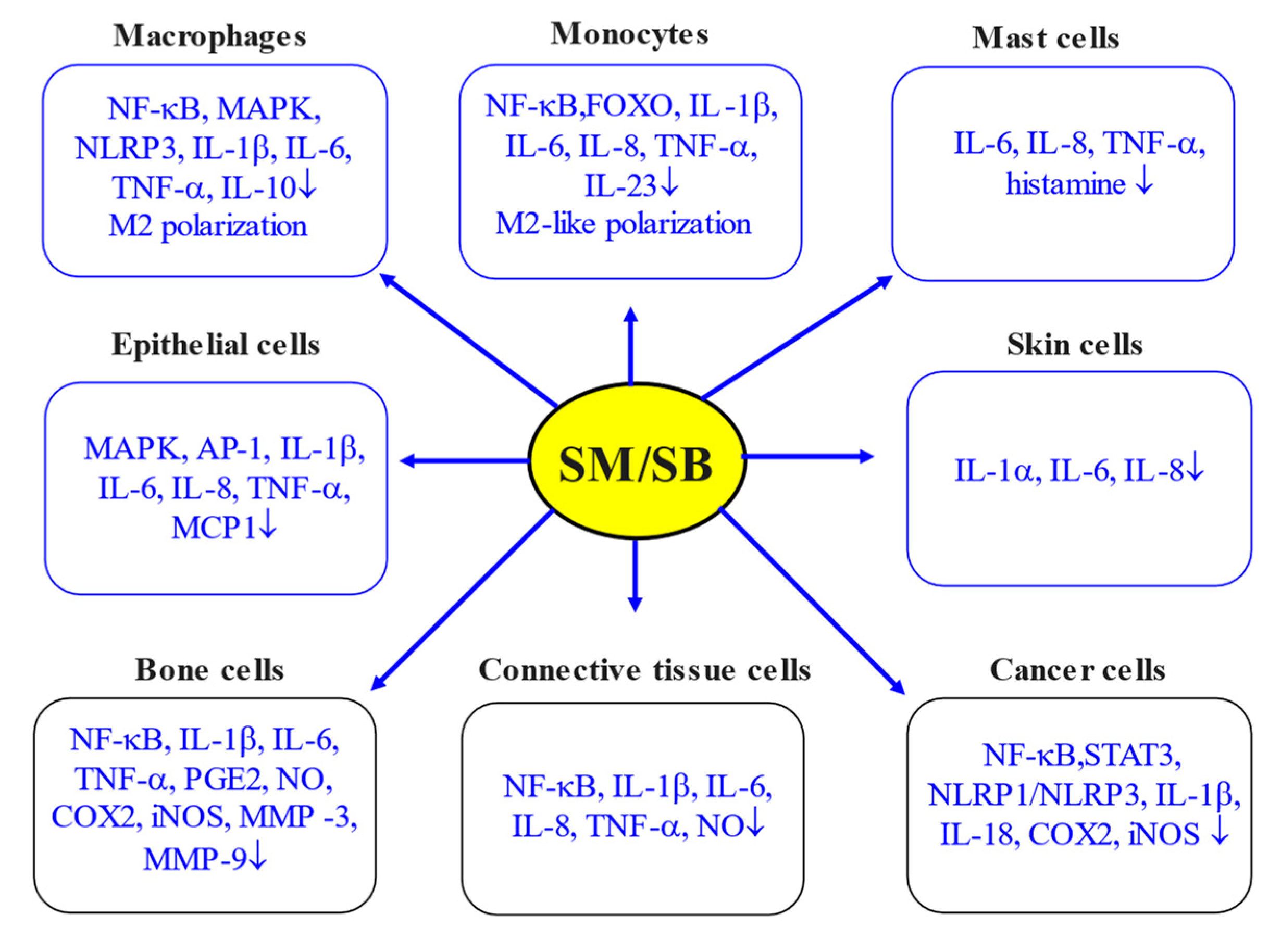
5. Silymarin and Inflammation: In Vitro Studies
5.1. Macrophages
5.2. Mononuclear Cells
5.3. Epithelial Cells
5.4. Skin and Bone Cells
5.5. Connective Tissue Cells
5.6. Cancer Cells
5.7. Other Cell Types
6. Silymarin and Inflammation: In Vivo Studies
6.1. Nonalcoholic Fatty Liver Disease (NAFLD)/Steatohepatitis/Diet-Induced Obesity
6.2. Toxicity Models
6.2.1. CCl4
6.2.2. Other Toxicants
6.3. Ischemia/Reperfusion (I/R) Models
6.4. Ageing and Exercising Models
6.5. Stress-Induced Injuries
6.6. Other In Vivo Model Systems and Disease States
6.7. Wound Healing
7. Silymarin and NF-κB Regulation
7.1. In Vitro Studies
7.2. In Vivo Studies
8. Conclusions
- Oxidative stress response;
- DNA damage response;
- Unfolded protein response;
- Heat shock response;
- Autophagy modulation;
- Formation of inflammasome;
- Non-coding RNA response;
- Inducible networks of signalling pathway formation and epigenetic changes.
Author Contributions
Funding
Conflicts of Interest
References
- Lei, Y.; Wang, K.; Deng, L.; Chen, Y.; Nice, E.C.; Huang, C. Redox regulation of inflammation: Old elements, a new story. Med. Res. Rev. 2015, 35, 306–340. [Google Scholar] [CrossRef] [PubMed]
- Surai, P.F.; Surai, A. Silymarin and inflammation: From understanding molecular mechanisms to practical applications. In Silymarin Puzzle; Surai, P.F., Surai, A., Eds.; Wageningen Academic Publishers: Wageningen, The Netherlands, 2023; pp. 287–317. [Google Scholar]
- Yu, W.; Tu, Y.; Long, Z.; Liu, J.; Kong, D.; Peng, J.; Wu, H.; Zheng, G.; Zhao, J.; Chen, Y.; et al. Reactive Oxygen Species Bridge the Gap between Chronic Inflammation and Tumor Development. Oxid. Med. Cell Longev. 2022, 2022, 2606928. [Google Scholar] [CrossRef] [PubMed]
- Surai, P.F. Vitagenes in Avian Biology and Poultry Health; Wageningen Academic Publishers: Wageningen, The Netherlands, 2020. [Google Scholar]
- Surai, P.F. Silymarin as a natural antioxidant: An overview of the current evidence and perspectives. Antioxidants 2015, 4, 204–247. [Google Scholar] [CrossRef] [PubMed]
- Surai, P.F. Silymarin as a vitagene modulator: Effects on mitochondria integrity in stress conditions. In Molecular Nutrition and Mitochondria; Ostojic, S.M., Ed.; Academic Press: Cambridge, MA, USA; Elsevier: London, UK, 2023; pp. 535–559. [Google Scholar]
- Sharma, S.; Kumar, P.; Ashawat, M.S.; Pandit, V.; Verma, C.S.; Sharma, D.K. Silymarin: A Phytoconstituent with Significant Therapeutic Potential-A Narrative Review. Curr. Drug Ther. 2023, 18, 89–97. [Google Scholar]
- Pérez, S.; Rius-Pérez, S. Macrophage Polarization and Reprogramming in Acute Inflammation: A Redox Perspective. Antioxidants 2022, 11, 1394. [Google Scholar] [CrossRef]
- Chen, L.; Deng, H.; Cui, H.; Fang, J.; Zuo, Z.; Deng, J.; Li, Y.; Wang, X.; Zhao, L. Inflammatory responses and inflammation-associated diseases in organs. Oncotarget 2018, 9, 7204. [Google Scholar] [CrossRef] [PubMed]
- Schmid-Schönbein, G.W. Analysis of inflammation. Annu. Rev. Biomed. Eng. 2006, 8, 93–151. [Google Scholar] [CrossRef]
- Ahmed, A.U. An overview of inflammation: Mechanism and consequences. Front. Biol. 2011, 6, 274–281. [Google Scholar] [CrossRef]
- Liu, D.; Zhong, Z.; Karin, M. NF-κB: A double-edged sword controlling inflammation. Biomedicines 2022, 10, 1250. [Google Scholar] [CrossRef]
- Broom, L.J.; Kogut, M.H. Inflammation: Friend or foe for animal production? Poult. Sci. 2018, 97, 510–514. [Google Scholar] [CrossRef]
- Robinson, M.W.; Harmon, C.; O’Farrelly, C. Liver immunology and its role in inflammation and homeostasis. Cell. Mol. Immunol. 2016, 13, 267–276. [Google Scholar] [CrossRef] [PubMed]
- Jain, P.; Pandey, R.; Shukla, S.S. Inflammation: Natural Resources and Its Applications; Springer: New Dehli, India, 2015. [Google Scholar]
- Fernandes, Q.; Inchakalody, V.P.; Bedhiafi, T.; Mestiri, S.; Taib, N.; Uddin, S.; Merhi, M.; Dermime, S. Chronic inflammation and cancer; the two sides of a coin. Life Sci. 2023, 338, 122390. [Google Scholar] [CrossRef] [PubMed]
- Surai, P.F.; Surai, A. Antioxidant defence systems in health and diseases. In Silymarin Puzzle; Surai, P.F., Surai, A., Eds.; Wageningen Academic Publishers: Wageningen, The Netherlands, 2023; pp. 85–102. [Google Scholar]
- Sies, H.; Jones, D.P. Reactive oxygen species (ROS) as pleiotropic physiological signalling agents. Nat. Rev. Mol. Cell Biol. 2020, 21, 363–383. [Google Scholar] [CrossRef]
- Bhattacharya, S.; Rattan, S.I. Primary stress response pathways for preconditioning and physiological hormesis. In The Science of Hormesis in Health and Longevity; Rattan, S.I.S., Kyriazis, M., Eds.; Academic Press: London, UK, 2019; pp. 35–51. [Google Scholar]
- Surai, P.F.; Kochish, I.I.; Fisinin, V.I.; Kidd, M.T. Antioxidant Defence Systems and Oxidative Stress in Poultry Biology: An Update. Antioxidants 2019, 8, 235. [Google Scholar] [CrossRef] [PubMed]
- Surai, P.F.; Surai, A. Silymarin and Nrf2. In Silymarin Puzzle; Surai, P.F., Surai, A., Eds.; Wageningen Academic Publishers: Wageningen, The Netherlands, 2023; pp. 127–150. [Google Scholar]
- Surai, P.F.; Kochish, I.I.; Kidd, M.T. Redox Homeostasis in Poultry: Regulatory Roles of NF-κB. Antioxidants 2021, 10, 186. [Google Scholar] [CrossRef] [PubMed]
- Surai, P.F.; Surai, A. Silymarin and NF-κB. In Silymarin Puzzle; Surai, P.F., Surai, A., Eds.; Wageningen Academic Publishers: Wageningen, The Netherlands, 2023; pp. 151–177. [Google Scholar]
- Austermann, J.; Roth, J.; Barczyk-Kahlert, K. The good and the bad: Monocytes’ and macrophages’ diverse functions in inflammation. Cells 2022, 11, 1979. [Google Scholar] [CrossRef]
- Juráňová, J.; Aury-Landas, J.; Boumediene, K.; Baugé, C.; Biedermann, D.; Ulrichová, J.; Franková, J. Modulation of skin inflammatory response by active components of silymarin. Molecules 2019, 24, 123. [Google Scholar] [CrossRef] [PubMed]
- Bijak, M. Silybin, a major bioactive component of milk thistle (Silybum marianum L. Gaernt.)—Chemistry, bioavailability, and metabolism. Molecules 2017, 22, 1942. [Google Scholar] [CrossRef]
- Křen, V.; Valentová, K. Silybin and its congeners: From traditional medicine to molecular effects. Nat. Prod. Rep. 2022, 39, 1264–1281. [Google Scholar] [CrossRef]
- Tvrdý, V.; Pourová, J.; Jirkovský, E.; Křen, V.; Valentová, K.; Mladěnka, P. Systematic review of pharmacokinetics and potential pharmacokinetic interactions of flavonolignans from silymarin. Med. Res. Rev. 2021, 41, 2195–2246. [Google Scholar] [CrossRef]
- Wang, X.; Zhang, Z.; Wu, S.C. Health benefits of Silybum marianum: Phytochemistry, pharmacology, and applications. J. Agric. Food Chem. 2020, 68, 11644–11664. [Google Scholar] [CrossRef]
- Aziz, M.; Saeed, F.; Ahmad, N.; Ahmad, A.; Afzaal, M.; Hussain, S.; Mohamed, A.A.; Alamri, M.S.; Anjum, F.M. Biochemical profile of milk thistle (Silybum marianum L.) with special reference to silymarin content. Food Sci. Nutr. 2020, 9, 244–250. [Google Scholar] [CrossRef]
- Martinelli, T.; Fulvio, F.; Pietrella, M.; Focacci, M.; Lauria, M.; Paris, R. In Silybum marianum Italian wild populations the variability of silymarin profiles results from the combination of only two stable chemotypes. Fitoterapia 2021, 148, 104797. [Google Scholar] [CrossRef]
- Devi, K.P. Milk thistle (Silybum marianum). In Nonvitamin and Nonmineral Nutritional Supplements; Seyed, N., Silva, A., Eds.; Elsevier: Amsterdam, The Netherlands; Academic Press: London, UK, 2019; pp. 321–325. [Google Scholar]
- Abenavoli, L.; Izzo, A.A.; Milić, N.; Cicala, C.; Santini, A.; Capasso, R. Milk thistle (Silybum marianum): A concise overview on its chemistry, pharmacological, and nutraceutical uses in liver diseases. Phytother. Res. 2018, 32, 2202–2213. [Google Scholar] [CrossRef]
- Fallah, M.; Davoodvandi, A.; Nikmanzar, S.; Aghili, S.; Mirazimi, S.; Aschner, M.; Rashidian, A.; Hamblin, M.R.; Chamanara, M.; Naghsh, N.; et al. Silymarin (milk thistle extract) as a therapeutic agent in gastrointestinal cancer. Biomed. Pharmacother. 2021, 142, 112024. [Google Scholar] [CrossRef]
- Křen, V. Chirality matters: Biological activity of optically pure silybin and Its congeners. Int. J. Mol. Sci. 2021, 22, 7885. [Google Scholar] [CrossRef]
- Kim, N.C.; Graf, T.N.; Sparacino, C.M.; Wani, M.C.; Wall, M.E. Complete isolation and characterization of silybins and isosilybins from milk thistle (Silybum marianum). Org. Biomol. Chem. 2003, 1, 1684–1689. [Google Scholar] [CrossRef]
- Degboé, Y.; Poupot, R.; Poupot, M. Repolarization of unbalanced macrophages: Unmet medical need in chronic inflammation and cancer. Int. J. Mol. Sci. 2022, 23, 1496. [Google Scholar] [CrossRef]
- Rendra, E.; Riabov, V.; Mossel, D.M.; Sevastyanova, T.; Harmsen, M.C.; Kzhyshkowska, J. Reactive oxygen species (ROS) in macrophage activation and function in diabetes. Immunobiology 2019, 224, 242–253. [Google Scholar] [CrossRef]
- Kapellos, T.S.; Iqbal, A.J. Epigenetic control of macrophage polarisation and soluble mediator gene expression during inflammation. Mediat. Inflamm. 2016, 2016, 6591703. [Google Scholar] [CrossRef]
- Kim, E.J.; Lee, M.Y.; Jeon, Y.J. Silymarin inhibits morphological changes in LPS-stimulated macrophages by blocking NF-κB pathway. Korean J. Physiol. Pharmacol. 2015, 19, 211–218. [Google Scholar] [CrossRef]
- Lu, C.P.; Huang, C.Y.; Wang, S.H.; Chiu, C.H.; Li, L.H.; Hua, K.F.; Wu, T.H. Improvement of hyperglycemia in a murine model of insulin resistance and high glucose- and inflammasome-mediated IL-1β expressions in macrophages by silymarin. Chem. Biol. Interact. 2018, 290, 12–18. [Google Scholar] [CrossRef]
- Chen, J.; Li, D.L.; Xie, L.N.; Ma, Y.R.; Wu, P.P.; Li, C.; Liu, W.F.; Zhang, K.; Zhou, R.P.; Xu, X.T.; et al. Synergistic anti-inflammatory effects of silibinin and thymol combination on LPS-induced RAW264.7 cells by inhibition of NF-κB and MAPK activation. Phytomedicine 2020, 78, 153309. [Google Scholar] [CrossRef]
- Zheng, Y.; Chen, J.; Wu, X.; Zhang, X.; Hu, C.; Kang, Y.; Lin, J.; Li, J.; Huang, Y.; Zhang, X.; et al. Enhanced anti-inflammatory effects of silibinin and capsaicin combination in lipopolysaccharide-induced RAW264.7 cells by inhibiting NF-κB and MAPK activation. Front. Chem. 2022, 10, 934541. [Google Scholar] [CrossRef]
- Huang, R.Y.; Chang, H.Y.; Chih, S.M.; Dyke, T.V.; Cheng, C.D.; Sung, C.E.; Weng, P.W.; Shieh, Y.S.; Cheng, W.C. Silibinin alleviates inflammation-induced bone loss by modulating biological interaction between human gingival fibroblasts and monocytes. J. Periodontol. 2023, 94, 905–918. [Google Scholar] [CrossRef]
- Mi, X.J.; Le, H.M.; Lee, S.; Park, H.R.; Kim, Y.J. Silymarin-Functionalized Selenium Nanoparticles Prevent LPS-Induced Inflammatory Response in RAW264.7 Cells through Downregulation of the PI3K/Akt/NF-κB Pathway. ACS Omega 2022, 7, 42723–42732. [Google Scholar] [CrossRef]
- Liu, Z.; Sun, M.; Wang, Y.; Zhang, L.; Zhao, H.; Zhao, M. Silymarin attenuated paraquat-induced cytotoxicity in macrophage by regulating Trx/TXNIP complex, inhibiting NLRP3 inflammasome activation and apoptosis. Toxicol. Vitr. 2018, 46, 265–272. [Google Scholar] [CrossRef]
- Zhang, B.; Xu, D.; She, L.; Wang, Z.; Yang, N.; Sun, R.; Zhang, Y.; Yan, C.; Wei, Q.; Aa, J.; et al. Silybin inhibits NLRP3 inflammasome assembly through the NAD+/SIRT2 pathway in mice with nonalcoholic fatty liver disease. FASEB J. 2018, 32, 757–767. [Google Scholar] [CrossRef]
- Bittencourt, M.; Rodrigues, R.P.; Kitagawa, R.R.; Gonçalves, R. The gastroprotective potential of silibinin against Helicobacter pylori infection and gastric tumor cells. Life Sci. 2020, 256, 117977. [Google Scholar] [CrossRef]
- Azadpour, M.; Farajollahi, M.M.; Dariushnejad, H.; Varzi, A.M.; Varezardi, A.; Barati, M. Effects of synthetic silymarin-PLGA nanoparticles on M2 polarization and inflammatory cytokines in LPS-treated murine peritoneal macrophages. Iran. J. Basic Med. Sci. 2021, 24, 1446–1454. [Google Scholar]
- Xu, W.; Sun, Y.; Wang, J.; Wang, B.; Xu, F.; Xie, Z.; Wang, Y. Controlled release of silibinin in GelMA hydrogels inhibits inflammation by inducing M2-type macrophage polarization and promotes vascularization in vitro. RSC Adv. 2022, 12, 13192–13202. [Google Scholar] [CrossRef]
- Bannwart, C.F.; Nakaira-Takahagi, E.; Golim, M.A.; de Medeiros, L.T.; Romão, M.; Weel, I.C.; Peraçoli, M.T. Downregulation of nuclear factor-kappa B (NF-kappaB) pathway by silibinin in human monocytes challenged with Paracoccidioides brasiliensis. Life Sci. 2010, 86, 880–886. [Google Scholar] [CrossRef]
- Giorgi, V.S.; Bannwart-Castro, C.F.; Peracoli, J.C.; Peracoli, M.T. Silibinin modulates NF-κB pathway and proinflammatory cytokines production by mononuclear cells of preeclamptic women. Pregnancy Hypertens. 2012, 2, 275–276. [Google Scholar] [CrossRef]
- Kim, B.R.; Seo, H.S.; Ku, J.M.; Kim, G.J.; Jeon, C.Y.; Park, J.H.; Jang, B.H.; Park, S.J.; Shin, Y.C.; Ko, S.G. Silibinin inhibits the production of pro-inflammatory cytokines through inhibition of NF-κB signaling pathway in HMC-1 human mast cells. Inflamm. Res. 2013, 62, 941–950. [Google Scholar] [CrossRef]
- Choi, Y.H.; Yan, G.H. Silibinin attenuates mast cell-mediated anaphylaxis-like reactions. Biol. Pharm. Bull. 2009, 32, 868–875. [Google Scholar] [CrossRef]
- Fordham, J.B.; Naqvi, A.R.; Nares, S. Leukocyte production of inflammatory mediators is inhibited by the antioxidants phloretin, silymarin, hesperetin, and resveratrol. Mediat. Inflamm. 2014, 2014, 938712. [Google Scholar] [CrossRef]
- Lovelace, E.S.; Wagoner, J.; MacDonald, J.; Bammler, T.; Bruckner, J.; Brownell, J.; Beyer, R.P.; Zink, E.M.; Kim, Y.M.; Kyle, J.E.; et al. Silymarin suppresses cellular inflammation by inducing reparative stress signaling. J. Nat. Prod. 2015, 78, 1990–2000. [Google Scholar] [CrossRef]
- Lovelace, E.S.; Maurice, N.J.; Miller, H.W.; Slichter, C.K.; Harrington, R.; Magaret, A.; Prlic, M.; De Rosa, S.; Polyak, S.J. Silymarin suppresses basal and stimulus-induced activation, exhaustion, differentiation, and inflammatory markers in primary human immune cells. PLoS ONE 2017, 12, e0171139. [Google Scholar] [CrossRef]
- Gugliandolo, E.; Crupi, R.; Biondi, V.; Licata, P.; Cuzzocrea, S.; Passantino, A. Protective effect of silibinin on lipopolysaccharide-induced inflammatory responses in equine peripheral blood mononuclear cells, an in vitro study. Animals 2020, 10, 2022. [Google Scholar] [CrossRef]
- Gomes, V.J.; Nunes, P.R.; Matias, M.L.; Ribeiro, V.R.; Devides, A.C.; Bannwart-Castro, C.F.; Romagnoli, G.G.; Peraçoli, J.C.; Peraçoli, M.; Romao-Veiga, M. Silibinin induces in vitro M2-like phenotype polarization in monocytes from preeclamptic women. Int. Immunopharmacol. 2020, 89, 107062. [Google Scholar] [CrossRef]
- Dupuis, M.L.; Conti, F.; Maselli, A.; Pagano, M.T.; Ruggieri, A.; Anticoli, S.; Fragale, A.; Gabriele, L.; Gagliardi, M.C.; Sanchez, M.; et al. The natural agonist of estrogen receptor β Silibinin plays an immunosuppressive role representing a potential therapeutic tool in rheumatoid arthritis. Front. Immunol. 2018, 9, 1903. [Google Scholar] [CrossRef]
- Li, D.; Hu, J.; Wang, T.; Zhang, X.; Liu, L.; Wang, H.; Wu, Y.; Xu, D.; Wen, F. Silymarin attenuates cigarette smoke extract-induced inflammation via simultaneous inhibition of autophagy and ERK/p38 MAPK pathway in human bronchial epithelial cells. Sci. Rep. 2016, 6, 37751. [Google Scholar] [CrossRef]
- Miethe, C.; Nix, H.; Martin, R.; Hernandez, A.R.; Price, R.S. Silibinin reduces the impact of obesity on invasive liver cancer. Nutr. Cancer 2017, 69, 1272–1280. [Google Scholar] [CrossRef]
- Lim, R.; Morwood, C.J.; Lim, J.O.; Shin, N.R.; Seo, Y.S.; Nam, H.H.; Ko, J.W.; Jung, T.Y.; Lee, S.J.; Kim, H.J.; et al. Silibinin attenuates silica dioxide nanoparticles-induced inflammation by suppressing TXNIP/MAPKs/AP-1 signaling. Cells 2020, 9, 678. [Google Scholar] [CrossRef]
- Speciale, A.; Muscarà, C.; Molonia, M.S.; Cimino, F.; Saija, A.; Giofrè, S.V. Silibinin as potential tool against SARS-CoV-2: In silico spike receptor-binding domain and main protease molecular docking analysis, and in vitro endothelial protective effects. Phytother. Res. 2021, 35, 4616–4625. [Google Scholar] [CrossRef]
- Xu, S.; Jiang, X.; Liu, Y.; Jiang, X.; Che, L.; Lin, Y.; Zhuo, Y.; Feng, B.; Fang, Z.; Hua, L.; et al. Silibinin Alleviates Lipopolysaccharide Induced Inflammation in Porcine Mammary Epithelial Cells via mTOR/NF-κB Signaling Pathway. Mol. Nutr. Food Res. 2023, 67, e2200715. [Google Scholar] [CrossRef]
- Tewari-Singh, N.; Jain, A.K.; Inturi, S.; Agarwal, C.; White, C.W.; Agarwal, R. Silibinin attenuates sulfur mustard analog-induced skin injury by targeting multiple pathways connecting oxidative stress and inflammation. PLoS ONE 2012, 7, e46149. [Google Scholar] [CrossRef]
- Frankova, J.; Juranova, J.; Biedermann, D.; Ulrichova, J. Influence of silymarin components on keratinocytes and 3D reconstructed epidermis. Toxicol. Vitr. 2021, 74, 105162. [Google Scholar] [CrossRef]
- Zheng, W.; Feng, Z.; Lou, Y.; Chen, C.; Zhang, C.; Tao, Z.; Li, H.; Cheng, L.; Ying, X. Silibinin protects against osteoarthritis through inhibiting the inflammatory response and cartilage matrix degradation in vitro and in vivo. Oncotarget 2017, 8, 99649–99665. [Google Scholar] [CrossRef]
- Fernandes, C.; Veiga, M.R.; Peracoli, M.; Zambuzzi, W.F. Modulatory effects of silibinin in cell behavior during osteogenic phenotype. J. Cell. Biochem. 2019, 120, 13413–13425. [Google Scholar] [CrossRef]
- Tong, W.W.; Zhang, C.; Hong, T.; Liu, D.H.; Wang, C.; Li, J.; He, X.K.; Xu, W.D. Silibinin alleviates inflammation and induces apoptosis in human rheumatoid arthritis fibroblast-like synoviocytes and has a therapeutic effect on arthritis in rats. Sci. Rep. 2018, 8, 3241. [Google Scholar] [CrossRef] [PubMed]
- Sharifi, R.; Pasalar, P.; Kamalinejad, M.; Dehpour, A.R.; Tavangar, S.M.; Paknejad, M.; Mehrabani Natanzi, M.; Nourbakhsh, M.; Ahmadi Ashtiani, H.R.; Akbari, M.; et al. The effect of silymarin (Silybum marianum) on human skin fibroblasts in an in vitro wound healing model. Pharm. Biol. 2013, 51, 298–303. [Google Scholar] [CrossRef]
- Dobiasová, S.; Řehořová, K.; Kučerová, D.; Biedermann, D.; Káňová, K.; Petrásková, L.; Koucká, K.; Václavíková, R.; Valentová, K.; Ruml, T.; et al. Multidrug resistance modulation activity of silybin derivatives and their anti-inflammatory Potential. Antioxidants 2020, 9, 455. [Google Scholar] [CrossRef] [PubMed]
- Raina, K.; Agarwal, C.; Agarwal, R. Effect of silibinin in human colorectal cancer cells: Targeting the activation of NF-κB signaling. Mol. Carcinog. 2013, 52, 195–206. [Google Scholar] [CrossRef] [PubMed]
- Zhang, B.; Wang, B.; Cao, S.; Wang, Y.; Wu, D. Silybin attenuates LPS-induced lung injury in mice by inhibiting NF-κB signaling and NLRP3 activation. Int. J. Mol. Med. 2017, 39, 1111–1118. [Google Scholar] [CrossRef] [PubMed]
- Matias, M.L.; Gomes, V.J.; Romao-Veiga, M.; Ribeiro, V.R.; Nunes, P.R.; Romagnoli, G.G.; Peracoli, J.C.; Peracoli, M. Silibinin downregulates the NF-κB pathway and NLRP1/NLRP3 inflammasomes in monocytes from pregnant women with preeclampsia. Molecules 2019, 24, 1548. [Google Scholar] [CrossRef] [PubMed]
- Cho, K.; Lee, H.G.; Piao, J.Y.; Kim, S.J.; Na, H.K.; Surh, Y.J. Protective effects of silibinin on Helicobacter pylori-induced gastritis: NF-κB and STAT3 as potential targets. J. Cancer Prev. 2021, 26, 118–127. [Google Scholar] [CrossRef]
- Faixová, D.; Ratvaj, M.; Maruščáková, I.C.; Hrčková, G.; Karaffová, V.; Faixová, Z.; Mudroňová, D. Silybin Showed Higher Cytotoxic, Antiproliferative, and Anti-Inflammatory Activities in the CaCo Cancer Cell Line while Retaining Viability and Proliferation in Normal Intestinal IPEC-1 Cells. Life 2023, 13, 492. [Google Scholar] [CrossRef]
- Yan, L.; Zhou, J.; Yuan, L.; Ye, J.; Zhao, X.; Ren, G.; Chen, H. Silibinin alleviates intestinal inflammation via inhibiting JNK signaling in Drosophila. Front. Pharmacol. 2023, 14, 1246960. [Google Scholar] [CrossRef]
- Yassin, N.; AbouZid, S.F.; El-Kalaawy, A.M.; Ali, T.M.; Elesawy, B.H.; Ahmed, O.M. Tackling of renal carcinogenesis in Wistar rats by Silybum marianum total extract, silymarin, and silibinin via modulation of oxidative stress, apoptosis, Nrf2, PPARγ, NF-κB, and PI3K/Akt signaling pathways. Oxid. Med. Cell. Longev. 2021, 2021, 7665169. [Google Scholar] [CrossRef]
- Shen, Y.; Zhao, H.; Wang, Z.; Guan, W.; Kang, X.; Tai, X.; Sun, Y. Silibinin declines blue light-induced apoptosis and inflammation through MEK/ERK/CREB of retinal ganglion cells. Artif. Cells Nanomed. Biotechnol. 2019, 47, 4059–4065. [Google Scholar] [CrossRef] [PubMed]
- Chen, Y.H.; Lin, H.; Wang, Q.; Hou, J.W.; Mao, Z.J.; Li, Y.G. Protective role of silibinin against myocardial ischemia/reperfusion injury-induced cardiac dysfunction. Int. J. Biol. Sci. 2020, 16, 1972–1988. [Google Scholar] [CrossRef] [PubMed]
- Meng, D.; Wang, Y.; Liu, T. Protective effects of silibinin on LPS-induced inflammation in human periodontal ligament cells. Front. Chem. 2022, 10, 1019663. [Google Scholar] [CrossRef] [PubMed]
- Li, X.; Zhou, R.; Han, Y.; Zeng, J.; Shi, L.; Mao, Y.; Sun, X.; Ji, Y.; Zhang, X.; Chen, Y.; et al. Silibinin Attenuates Experimental Periodontitis by Downregulation of Inflammation and Oxidative Stress. Oxid. Med. Cell. Longev. 2023, 2023, 5617800. [Google Scholar] [CrossRef]
- Saliou, C.; Valacchi, G.; Rimbach, G. Assessing bioflavonoids as regulators of NF-kappa B activity and inflammatory gene expression in mammalian cells. Methods Enzymol. 2001, 335, 380–387. [Google Scholar] [PubMed]
- Podder, B.; Kim, Y.S.; Zerin, T.; Song, H.Y. Antioxidant effect of silymarin on paraquat-induced human lung adenocarcinoma A549 cell line. Food Chem. Toxicol. 2012, 50, 3206–3214. [Google Scholar] [CrossRef] [PubMed]
- Zhao, X.; Wang, H.; Yang, Y.; Gou, Y.; Wang, Z.; Yang, D.; Li, C. Protective effects of silymarin against D-Gal/LPS-induced organ damage and Inflammation in mice. Drug Des. Devel. Ther. 2021, 15, 1903–1914. [Google Scholar] [CrossRef] [PubMed]
- Schümann, J.; Prockl, J.; Kiemer, A.K.; Vollmar, A.M.; Bang, R.; Tiegs, G. Silibinin protects mice from T cell-dependent liver injury. J. Hepatol. 2003, 39, 333–340. [Google Scholar] [CrossRef]
- Thibaut, R.; Gage, M.C.; Pineda-Torra, I.; Chabrier, G.; Venteclef, N.; Alzaid, F. Liver macrophages and inflammation in physiology and physiopathology of non-alcoholic fatty liver disease. FEBS J. 2022, 289, 3024–3057. [Google Scholar] [CrossRef]
- Itoh, M.; Ogawa, Y.; Suganami, T. Chronic inflammation as a molecular basis of nonalcoholic steatohepatitis: Role of macrophages and fibroblasts in the liver. Nagoya J. Med. Sci. 2020, 82, 391–397. [Google Scholar]
- Salamone, F.; Galvano, F.; Marino, A.; Paternostro, C.; Tibullo, D.; Bucchieri, F.; Mangiameli, A.; Parola, M.; Bugianesi, E.; Volti, G.L. Silibinin improves hepatic and myocardial injury in mice with nonalcoholic steatohepatitis. Dig. Liver Dis. 2012, 44, 334–342. [Google Scholar] [CrossRef] [PubMed]
- Marin, V.; Gazzin, S.; Gambaro, S.E.; Dal Ben, M.; Calligaris, S.; Anese, M.; Raseni, A.; Avellini, C.; Giraudi, P.J.; Tiribelli, C.; et al. Effects of oral administration of silymarin in a juvenile murine model of non-alcoholic steatohepatitis. Nutrients 2017, 9, 1006. [Google Scholar] [CrossRef] [PubMed]
- Lee, S.J.; Nam, M.J.; Lee, D.E.; Park, J.W.; Kang, B.S.; Lee, D.S.; Lee, H.S.; Kwon, O.S. Silibinin ameliorates O-GlcNAcylation and inflammation in a mouse model of nonalcoholic steatohepatitis. Int. J. Mol. Sci. 2018, 19, 2165. [Google Scholar] [CrossRef] [PubMed]
- Ou, Q.; Weng, Y.; Wang, S.; Zhao, Y.; Zhang, F.; Zhou, J.; Wu, X. Silybin alleviates hepatic steatosis and fibrosis in NASH mice by inhibiting oxidative stress and involvement with the Nf-κB pathway. Dig. Dis. Sci. 2018, 63, 3398–3408. [Google Scholar] [CrossRef] [PubMed]
- Zhang, R.; Xu, D.; Zhang, Y.; Wang, R.; Yang, N.; Lou, Y.; Zhao, H.; Aa, J.; Wang, G.; Xie, Y. Silybin restored CYP3A expression through the Sirtuin 2/Nuclear Factor κ-B pathway in mouse nonalcoholic fatty liver disease. Drug Metab. Dispos. 2021, 49, 770–779. [Google Scholar] [CrossRef] [PubMed]
- Gu, M.; Zhao, P.; Huang, J.; Zhao, Y.; Wang, Y.; Li, Y.; Li, Y.; Fan, S.; Ma, Y.M.; Tong, Q.; et al. Silymarin ameliorates metabolic dysfunction associated with diet-Induced obesity via activation of Farnesyl X receptor. Front. Pharmacol. 2016, 7, 345. [Google Scholar] [CrossRef]
- Guo, Y.; Wang, S.; Wang, Y.; Zhu, T. Silymarin improved diet-induced liver damage and insulin resistance by decreasing inflammation in mice. Pharm. Biol. 2016, 54, 2995–3000. [Google Scholar] [CrossRef]
- Alsaggar, M.; Bdour, S.; Ababneh, Q.; El-Elimat, T.; Qinna, N.; Alzoubi, K.H. Silibinin attenuates adipose tissue inflammation and reverses obesity and its complications in diet-induced obesity model in mice. BMC Pharmacol. Toxicol. 2020, 21, 8. [Google Scholar] [CrossRef]
- Shen, H.H.; Alex, R.; Bellner, L.; Raffaele, M.; Licari, M.; Vanella, L.; Stec, D.E.; Abraham, N.G. Milk thistle seed cold press oil attenuates markers of the metabolic syndrome in a mouse model of dietary-induced obesity. J. Food Biochem. 2020, 44, e13522. [Google Scholar] [CrossRef]
- Li, S.; Duan, F.; Li, S.; Lu, B. Administration of Silymarin in NAFLD/NASH: A systematic review and meta-analysis. Ann. Hepatol. 2023, in press. [Google Scholar] [CrossRef]
- Luster, M.I.; Simeonova, P.P.; Gallucci, R.M.; Matheson, J.M.; Yucesoy, B. Immunotoxicology: Role of inflammation in chemical-induced hepatoto-xicity. Int. J. Immunopharmacol. 2000, 22, 1143–1147. [Google Scholar] [CrossRef] [PubMed]
- Clichici, S.; Olteanu, D.; Filip, A.; Nagy, A.L.; Oros, A.; Mircea, P.A. Beneficial effects of silymarin after the discontinuation of CCl4-induced liver fibrosis. J. Med. Food 2016, 19, 789–797. [Google Scholar] [CrossRef] [PubMed]
- Sokar, S.S.; El-Sayad, M.E.; Ghoneim, M.E.; Shebl, A.M. Combination of Sitagliptin and silymarin ameliorates liver fibrosis induced by carbon tetrachloride in rats. Biomed. Pharmacother. 2017, 89, 98–107. [Google Scholar] [CrossRef]
- Al-Rasheed, N.; Faddah, L.; Al-Rasheed, N.; Bassiouni, Y.A.; Hasan, I.H.; Mahmoud, A.M.; Mohamad, R.A.; Yacoub, H.I. Protective effects of silymarin, alone or in combination with chlorogenic acid and/or melatonin, against carbon tetrachloride-induced hepatotoxicity. Pharmacogn. Mag. 2016, 12, S337–S345. [Google Scholar] [PubMed]
- Zhao, X.A.; Chen, G.M.; Liu, Y.; Chen, Y.X.; Wu, H.Y.; Chen, J.; Xiong, Y.L.; Tian, C.; Wang, G.Y.; Jia, B.; et al. Inhibitory effect of silymarin on CCl4-induced liver fibrosis by reducing Ly6Chi monocytes infiltration. Int. J. Clin. Exp. Pathol. 2017, 10, 11941–11951. [Google Scholar]
- El-Kot, S.M.; Wanas, W.; Hafez, A.M.; Mahmoud, N.A.; Tolba, A.M.; Younis, A.H.; Sayed, G.E.; Abdelwahab, H.E. Effect of silymarin on the relative gene expressions of some inflammatory cytokines in the liver of CCl4-intoxicated male rats. Sci. Rep. 2023, 13, 15245. [Google Scholar] [CrossRef]
- Ye, Z.; Zhang, X.; Huang, Q.; Zhang, W.; Ye, M. Synergistic hepatoprotective effect of combined administration of Lachnum polysaccharide with silymarin. Bioorg. Med. Chem. Lett. 2021, 46, 128159. [Google Scholar] [CrossRef]
- Ahmad, I.; Shukla, S.; Kumar, A.; Singh, B.K.; Kumar, V.; Chauhan, A.K.; Singh, D.; Pandey, H.P.; Singh, C. Biochemical and molecular mechanisms of N-acetyl cysteine and silymarin-mediated protection against maneb- and paraquat-induced hepatotoxicity in rats. Chem. Biol. Interact. 2013, 201, 9–18. [Google Scholar] [CrossRef]
- Razavi-Azarkhiavi, K.; Ali-Omrani, M.; Solgi, R.; Bagheri, P.; Haji-Noormohammadi, M.; Amani, N.; Sepand, M.R. Silymarin alleviates bleomycin-induced pulmonary toxicity and lipid peroxidation in mice. Pharm. Biol. 2014, 52, 1267–1271. [Google Scholar] [CrossRef]
- Liu, W.; Li, Y.; Zheng, X.; Zhang, K.; Du, Z. Potent inhibitory effect of silibinin from milk thistle on skin inflammation stimuli by 12-O-tetradecanoylphorbol-13-acetate. Food Funct. 2015, 6, 3712–3719. [Google Scholar] [CrossRef]
- Zaulet, M.; Kevorkian, S.; Dinescu, S.; Cotoraci, C.; Suciu, M.; Herman, H.; Buburuzan, L.; Badulescu, L.; Ardelean, A.; Hermenean, A. Protective effects of silymarin against bisphenol A-induced hepatotoxicity in mouse liver. Exp. Ther. Med. 2017, 13, 821–828. [Google Scholar] [CrossRef] [PubMed]
- Wang, L.; Huang, Q.H.; Li, Y.X.; Huang, Y.F.; Xie, J.H.; Xu, L.Q.; Dou, Y.X.; Su, Z.R.; Zeng, H.F.; Chen, J.N. Protective effects of silymarin on triptolide-induced acute hepatotoxicity in rats. Mol. Med. Rep. 2018, 17, 789–800. [Google Scholar] [CrossRef] [PubMed]
- Papackova, Z.; Heczkova, M.; Dankova, H.; Sticova, E.; Lodererova, A.; Bartonova, L.; Poruba, M.; Cahova, M. Silymarin prevents acetaminophen-induced hepatotoxicity in mice. PLoS ONE 2018, 13, e0191353. [Google Scholar] [CrossRef] [PubMed]
- Vangaveti, S.; Das, P.; Kumar, V.L. Metformin and silymarin afford protection in cyclosporine A induced hepatorenal toxicity in rat by modulating redox status and inflammation. J. Biochem. Mol. Toxicol. 2021, 35, e22614. [Google Scholar] [CrossRef] [PubMed]
- Yardım, A.; Kucukler, S.; Özdemir, S.; Çomaklı, S.; Caglayan, C.; Kandemir, F.M.; Çelik, H. Silymarin alleviates docetaxel-induced central and peripheral neurotoxicity by reducing oxidative stress, inflammation and apoptosis in rats. Gene 2021, 769, 145239. [Google Scholar] [CrossRef]
- Hussein, R.M.; Sawy, D.M.; Kandeil, M.A.; Farghaly, H.S. Chlorogenic acid, quercetin, coenzyme Q10 and silymarin modulate Keap1-Nrf2/heme oxygenase-1 signaling in thioacetamide-induced acute liver toxicity. Life Sci. 2021, 277, 119460. [Google Scholar] [CrossRef] [PubMed]
- Saxena, N.; Dhaked, R.K.; Nagar, D.P. Silibinin ameliorates abrin induced hepatotoxicity by attenuating oxidative stress, inflammation and inhibiting Fas pathway. Environ. Toxicol. Pharmacol. 2022, 93, 103868. [Google Scholar] [CrossRef]
- Abbas, N.; Awad, M.M.; Nafea, O.E. Silymarin in combination with chlorogenic acid protects against hepatotoxicity induced by doxorubicin in rats: Possible role of adenosine monophosphate-activated protein kinase pathway. Toxicol. Res. 2020, 9, 771–777. [Google Scholar] [CrossRef]
- Singh, G.; Mittra, N.; Singh, C. Tempol and silymarin rescue from zinc-induced degeneration of dopaminergic neurons through modulation of oxidative stress and inflammation. Mol. Cell. Biochem. 2023, 478, 1705–1718. [Google Scholar] [CrossRef]
- Jin, Y.; Zhao, X.; Zhang, H.; Li, Q.; Lu, G.; Zhao, X. Modulatory effect of silymarin on pulmonary vascular dysfunction through HIF-1α-iNOS following rat lung ischemia-reperfusion injury. Exp. Ther. Med. 2016, 12, 1135–1140. [Google Scholar] [CrossRef]
- Moghaddam, A.H.; Mokhtari Sangdehi, S.R.; Ranjbar, M.; Hasantabar, V. Preventive effect of silymarin-loaded chitosan nanoparticles against global cerebral ischemia/reperfusion injury in rats. Eur. J. Pharmacol. 2020, 877, 173066. [Google Scholar] [CrossRef] [PubMed]
- Akbari-Kordkheyli, V.; Abbaszadeh-Goudarzi, K.; Nejati-Laskokalayeh, M.; Zarpou, S.; Khonakdar-Tarsi, A. The protective effects of silymarin on ischemia-reperfusion injuries: A mechanistic review. Iran. J. Basic. Med. Sci. 2019, 22, 968–976. [Google Scholar] [PubMed]
- Zarpou, S.; Mosavi, H.; Bagheri, A.; Malekzadeh Shafaroudi, M.; Khonakdar-Tarsi, A. NF-κB and NLRP3 gene expression changes during warm hepatic ischemia-reperfusion in rats with and without silibinin. Gastroenterol. Hepatol. Bed Bench. 2021, 14, 267–275. [Google Scholar] [PubMed]
- Pasala, P.K.; Uppara, R.K.; Rudrapal, M.; Zothantluanga, J.H.; Umar, A.K. Silybin phytosome attenuates cerebral ischemia-reperfusion injury in rats by suppressing oxidative stress and reducing inflammatory response: In vivo and in silico approaches. J. Biochem. Mol. Toxicol. 2022, 36, e23073. [Google Scholar] [CrossRef]
- Kyriakopoulos, G.; Lambropoulou, M.; Valsami, G.; Kostomitsopoulos, N.; Konstandi, O.; Anagnostopoulos, K.; Tsalikidis, C.; Oikonomou, P.; Simopoulos, K.; Tsaroucha, A.K. Pro-inflammatory cytokines/chemokines, TNF-α, IL-6 and MCP-1, as biomarkers for the nephro- and pneumoprotective effect of silibinin after hepatic ischemia/reperfusion: Confirmation by immunohistochemistry and qRT-PCR. Basic Clin. Pharmacol. Toxicol. 2022, 130, 457–467. [Google Scholar] [CrossRef]
- Jin, G.; Bai, D.; Yin, S.; Yang, Z.; Zou, D.; Zhang, Z.; Li, X.; Sun, Y.; Zhu, Q. Silibinin rescues learning and memory deficits by attenuating microglia activation and preventing neuroinflammatory reactions in SAMP8 mice. Neurosci. Lett. 2016, 629, 256–261. [Google Scholar] [CrossRef]
- Kumar, J.; Park, K.C.; Awasthi, A.; Prasad, B. Silymarin extends lifespan and reduces proteotoxicity in C. elegans Alzheimer’s model. CNS Neurol. Disord. Drug Targets 2015, 14, 295–302. [Google Scholar] [CrossRef] [PubMed]
- Sarubbo, F.; Ramis, M.R.; Kienzer, C.; Aparicio, S.; Esteban, S.; Miralles, A.; and Moranta, D. Chronic silymarin, quercetin and naringenin treatments increase monoamines synthesis and hippocampal Sirt1 levels improving cognition in aged rats. J. Neuroimmune. Pharmacol. 2018, 13, 24–38. [Google Scholar] [CrossRef]
- Vargas-Mendoza, N.; Ángeles-Valencia, M.; Madrigal-Santillán, E.O.; Morales-Martínez, M.; Tirado-Lule, J.M.; Solano-Urrusquieta, A.; Madrigal-Bujaidar, E.; Álvarez-González, I.; Fregoso-Aguilar, T.; Morales-González, Á.; et al. Effect of silymarin supplementation on physical performance, muscle and myocardium histological changes, bodyweight, and food consumption in rats subjected to regular exercise training. Int. J. Mol. Sci. 2020, 21, 7724. [Google Scholar] [CrossRef]
- Vargas-Mendoza, N.; Angeles-Valencia, M.; Morales-González, Á.; Morales-Martínez, M.; Madrigal-Bujaidar, E.; Álvarez-González, I.; Fregoso-Aguilar, T.; Delgado-Olivares, L.; Madrigal-Santillán, E.O.; Morales-González, J.A. Effect of silymarin supplementation in lung and liver histological modifications during exercise training in a rodent model. J. Funct. Morphol. Kinesiol. 2021, 6, 72. [Google Scholar] [CrossRef]
- Aghaei, F.; Wong, A.; Zargani, M.; Sarshin, A.; Feizolahi, F.; Derakhshan, Z.; Hashemi, M.; Arabzadeh, E. Effects of swimming exercise combined with silymarin and vitamin C supplementation on hepatic inflammation, oxidative stress, and histopathology in elderly rats with high-fat diet-induced liver damage. Nutrition 2023, 115, 112167. [Google Scholar] [CrossRef] [PubMed]
- Kim, S.H.; Oh, D.S.; Oh, J.Y.; Son, T.G.; Yuk, D.Y.; Jung, Y.S. Silymarin prevents restraint stress-induced acute liver injury by ameliorating oxidative stress and reducing inflammatory response. Molecules 2016, 21, 443. [Google Scholar] [CrossRef] [PubMed]
- Zhu, Z.; Sun, G. Silymarin mitigates lung impairments in a rat model of acute respiratory distress syndrome. Inflammopharmacology 2018, 26, 747–754. [Google Scholar] [CrossRef] [PubMed]
- Garikapati, D.R.; Shaik, P.B.; Penchalaiah, H. Evaluate neuroprotective effect of silibinin using chronic unpredictable stress (cus) model. Int. J. Physiol. Pathophysiol. Pharmacol. 2018, 10, 184–191. [Google Scholar] [PubMed]
- Son, Y.; Lee, H.J.; Rho, J.K.; Chung, S.Y.; Lee, C.G.; Yang, K.; Kim, S.H.; Lee, M.; Shin, I.S.; Kim, J.S. The ameliorative effect of silibinin against radiation-induced lung injury: Protection of normal tissue without decreasing therapeutic efficacy in lung cancer. BMC Pulm. Med. 2015, 15, 68. [Google Scholar] [CrossRef] [PubMed]
- Rigby, C.M.; Roy, S.; Deep, G.; Guillermo-Lagae, R.; Jain, A.K.; Dhar, D.; Orlicky, D.J.; Agarwal, C.; Agarwal, R. Role of p53 in silibinin-mediated inhibition of ultraviolet B radiation-induced DNA damage, inflammation and skin carcino-genesis. Carcinogenesis 2017, 38, 40–50. [Google Scholar] [CrossRef] [PubMed]
- Ali Karami, M.; Sharif Makhmalzadeh, B.; Pooranian, M.; Rezai, A. Preparation and optimization of silibinin-loaded chitosan-fucoidan hydrogel: An in vivo evaluation of skin protection against UVB. Pharm. Dev. Technol. 2021, 26, 209–219. [Google Scholar] [CrossRef]
- Cha, S.R.; Jang, J.; Park, S.M.; Ryu, S.M.; Cho, S.J.; Yang, S.R. Cigarette Smoke-Induced Respiratory Response: Insights into Cellular Processes and Biomarkers. Antioxidants 2023, 12, 1210. [Google Scholar] [CrossRef]
- Ko, J.W.; Shin, N.R.; Park, S.H.; Lee, I.C.; Ryu, J.M.; Kim, H.J.; Cho, Y.K.; Kim, J.C.; Shin, I.S. Silibinin inhibits the fibrotic responses induced by cigarette smoke via suppression of TGF-β1/Smad 2/3 signaling. Food Chem. Toxicol. 2017, 106, 424–429. [Google Scholar] [CrossRef]
- Li, D.; Xu, D.; Wang, T.; Shen, Y.; Guo, S.; Zhang, X.; Guo, L.; Li, X.; Liu, L.; Wen, F. Silymarin attenuates airway inflammation induced by cigarette smoke in mice. Inflammation 2015, 38, 871–878. [Google Scholar] [CrossRef]
- Park, J.W.; Shin, N.R.; Shin, I.S.; Kwon, O.K.; Kim, J.S.; Oh, S.R.; Kim, J.H.; Ahn, K.S. Silibinin inhibits neutrophilic inflammation and mucus secretion induced by cigarette smoke via suppression of ERK-SP1 pathway. Phytother. Res. 2016, 30, 1926–1936. [Google Scholar] [CrossRef] [PubMed]
- Ahmed, R.F.; Moussa, R.A.; Eldemerdash, R.S.; Zakaria, M.M.; Abdel-Gaber, S.A. Ameliorative effects of silymarin on HCl-induced acute lung injury in rats; role of the Nrf-2/HO-1 pathway. Iran. J. Basic. Med. Sci. 2019, 22, 1483–1492. [Google Scholar] [PubMed]
- Hanafy, N.; El-Kemary, M.A. Silymarin/curcumin loaded albumin nanoparticles coated by chitosan as muco-inhalable delivery system observing anti-inflammatory and anti COVID-19 characterizations in oleic acid triggered lung injury and in vitro COVID-19 experiment. Int. J. Biol. Macromol. 2022, 198, 101–110. [Google Scholar] [CrossRef] [PubMed]
- Kosari-Nasab, M.; Shokouhi, G.; Ghorbanihaghjo, A.; Abbasi, M.M.; Salari, A.A. Anxiolytic- and antidepressant-like effects of silymarin compared to diazepam and fluoxetine in a mouse model of mild traumatic brain injury. Toxicol. Appl. Pharmacol. 2018, 338, 159–173. [Google Scholar] [CrossRef] [PubMed]
- Shokouhi, G.; Kosari-Nasab, M.; Salari, A.A. Silymarin sex-dependently improves cognitive functions and alters TNF-α, BDNF, and glutamate in the hippocampus of mice with mild traumatic brain injury. Life Sci. 2020, 257, 118049. [Google Scholar] [CrossRef]
- Clichici, S.; David, L.; Moldovan, B.; Baldea, I.; Olteanu, D.; Filip, M.; Nagy, A.; Luca, V.; Crivii, C.; Mircea, P.; et al. Hepatoprotective effects of silymarin coated gold nanoparticles in experimental cholestasis. Mater. Sci. Eng. C Mater. Biol. Appl. 2020, 115, 111117. [Google Scholar] [CrossRef]
- Nazemian, F.; Karimi, G.; Moatamedi, M.; Charkazi, S.; Shamsara, J.; Mohammadpour, A.H. Effect of silymarin administration on TNF-α serum concentration in peritoneal dialysis patients. Phytother. Res. 2010, 24, 1654–1657. [Google Scholar] [CrossRef]
- Atawia, R.T.; Mosli, H.H.; Tadros, M.G.; Khalifa, A.E.; Mosli, H.A.; Abdel-Naim, A.B. Modulatory effect of silymarin on inflammatory mediators in experimentally induced benign prostatic hyperplasia: Emphasis on PTEN, HIF-1α, and NF-κB. Naunyn Schmiedebergs Arch. Pharmacol. 2014, 387, 1131–1140. [Google Scholar] [CrossRef]
- Mazhari, S.; Razi, M.; Sadrkhanlou, R. Silymarin and celecoxib ameliorate experimental varicocele-induced pathogenesis: Evidences for oxidative stress and inflammation inhibition. Int. Urol. Nephrol. 2018, 50, 1039–1052. [Google Scholar] [CrossRef]
- Mady, F.M.; Essa, H.; El-Ammawi, T.; Abdelkader, H.; Hussein, A.K. Formulation and clinical evaluation of silymarin pluronic-lecithin organogels for treatment of atopic dermatitis. Drug Des. Devel. Ther. 2016, 10, 1101–1110. [Google Scholar]
- Rigon, C.; Marchiori, M.; da Silva Jardim, F.; Pegoraro, N.S.; Chaves, P.; Velho, M.C.; Beck, R.; Ourique, A.F.; Sari, M.; Oliveira, S.M.; et al. Hydrogel containing silibinin nanocapsules presents effective anti-inflammatory action in a model of irritant contact dermatitis in mice. Eur. J. Pharm. Sci. 2019, 137, 104969. [Google Scholar] [CrossRef] [PubMed]
- Choi, Y.H.; Jin, G.Y.; Guo, H.S.; Piao, H.M.; Li, L.C.; Li, G.Z.; Lin, Z.H.; Yan, G.H. Silibinin attenuates allergic airway inflammation in mice. Biochem. Biophys. Res. Commun. 2012, 427, 450–455. [Google Scholar] [CrossRef] [PubMed]
- Min, K.; Yoon, W.K.; Kim, S.K.; Kim, B.H. Immunosuppressive effect of silibinin in experimental autoimmune encephalomyelitis. Arch. Pharm. Res. 2007, 30, 1265–1272. [Google Scholar] [CrossRef] [PubMed]
- Toklu, H.Z.; Tunali Akbay, T.; Velioglu-Ogunc, A.; Ercan, F.; Gedik, N.; Keyer-Uysal, M.; Sener, G. Silymarin, the antioxidant component of Silybum marianum, prevents sepsis-induced acute lung and brain injury. J. Surg. Res. 2008, 145, 214–222. [Google Scholar] [CrossRef] [PubMed]
- Nasab, E.M.; Athari, S.M.; Ghafarzade, S.; Nasab, A.M.; Athari, S.S. Immunomodulatory effects of two silymarin isomers in a Balb/c mouse model of allergic asthma. Allergol. Immunopathol. 2020, 48, 646–653. [Google Scholar] [CrossRef]
- Navabi, F.; Shaygannejad, V.; Abbasirad, F.; Vaez, E.; Hosseininasab, F.; Kazemi, M.; Mirmosayyeb, O.; Alsahebfosoul, F.; Esmaeil, N. Immunoregulatory effects of silymarin on proliferation and activation of Th1 cells isolated from newly diagnosed and IFN-ß1b-treated MS patients. Inflammation 2019, 42, 54–63. [Google Scholar] [CrossRef]
- Yang, H.L.; Shi, X.W. Silybin alleviates experimental autoimmune encephalomyelitis by suppressing dendritic cell activation and Th17 cell differentiation. Front. Neurol. 2021, 12, 659678. [Google Scholar] [CrossRef]
- Ferraz, A.C.; Almeida, L.T.; da Silva Caetano, C.C.; da Silva Menegatto, M.B.; Souza Lima, R.L.; de Senna, J.; de Oliveira Cardoso, J.M.; Perucci, L.O.; Talvani, A.; Geraldo de Lima, W.; et al. Hepatoprotective, antioxidant, anti-inflammatory, and antiviral activities of silymarin against mayaro virus infection. Antivir. Res. 2021, 194, 105168. [Google Scholar] [CrossRef]
- Stolf, A.M.; Cardoso, C.C.; Acco, A. 2017. Effects of silymarin on diabetes mellitus complications: A review. Phytother. Res. 2017, 31, 366–374. [Google Scholar] [CrossRef]
- Chu, C.; Li, D.; Zhang, S.; Ikejima, T.; Jia, Y.; Wang, D.; Xu, F. Role of silibinin in the management of diabetes mellitus and its complications. Arch. Pharm. Res. 2018, 41, 785–796. [Google Scholar] [CrossRef]
- Lempesis, I.G.; Georgakopoulou, V.E. Physiopathological mechanisms related to inflammation in obesity and type 2 diabetes mellitus. World J. Exp. Med. 2023, 13, 7–16. [Google Scholar] [CrossRef] [PubMed]
- Stolf, A.M.; Campos Cardoso, C.; Morais, H.; Alves de Souza, C.E.; Lomba, L.A.; Brandt, A.P.; Agnes, J.P.; Collere, F.C.; Galindo, C.M.; Corso, C.R.; et al. Effects of silymarin on angiogenesis and oxidative stress in streptozotocin-induced diabetes in mice. Biomed. Pharmacother. 2018, 108, 232–243. [Google Scholar] [CrossRef] [PubMed]
- Chen, Y.; Chen, L.; Yang, T. Silymarin nanoliposomes attenuate renal injury on diabetic nephropathy rats via co-suppressing TGF-β/Smad and JAK2/STAT3/SOCS1 pathway. Life Sci. 2021, 271, 119197. [Google Scholar] [CrossRef] [PubMed]
- Mohammadi, H.; Manouchehri, H.; Changizi, R.; Bootorabi, F.; Khorramizadeh, M.R. Concurrent metformin and silibinin therapy in diabetes: Assessments in zebrafish (Danio rerio) animal model. J. Diabetes Metab. Disord. 2020, 19, 1233–1244. [Google Scholar] [CrossRef] [PubMed]
- Zhang, J.; Shi, Q.; Hu, Y.; Li, X. Silibinin augments the effect of clopidogrel on atherosclerosis in diabetic ApoE deficiency mice. Clin. Hemorheol. Microcirc. 2022, 80, 353–361. [Google Scholar] [CrossRef] [PubMed]
- Wang, C.; Wang, Z.; Zhang, X.; Zhang, X.; Dong, L.; Xing, Y.; Li, Y.; Liu, Z.; Chen, L.; Qiao, H.; et al. Protection by silibinin against experimental ischemic stroke: Up-regulated pAkt, pmTOR, HIF-1α and Bcl-2, down-regulated Bax, NF-κB expression. Neurosci. Lett. 2012, 529, 45–50. [Google Scholar] [CrossRef]
- Li, Z.; Wang, H.; Yin, Y. Peripheral inflammation is a potential etiological factor in Alzheimer’s disease. Rev. Neurosci. 2023, in press. [Google Scholar] [CrossRef]
- Wiatrak, B.; Jawień, P.; Szeląg, A.; Jęśkowiak-Kossakowska, I. Does Inflammation Play a Major Role in the Pathogenesis of Alzheimer’s Disease? Neuromolecular Med. 2023, 25, 330–335. [Google Scholar] [CrossRef]
- Standaert, D.G.; Harms, A.S.; Childers, G.M.; Webster, J.M. Disease mechanisms as subtypes: Inflammation in Parkinson disease and related disorders. Handb. Clin. Neurol. 2023, 193, 95–106. [Google Scholar]
- Williams, G.P.; Schonhoff, A.M.; Sette, A.; Lindestam Arlehamn, C.S. Central and Peripheral Inflammation: Connecting the Immune Responses of Parkinson’s Disease. J. Park. Dis. 2022, 12, S129–S136. [Google Scholar] [CrossRef]
- Jung, U.J.; Jeon, M.T.; Choi, M.S.; Kim, S.R. Silibinin attenuates MPP+-induced neurotoxicity in the substantia nigra in vivo. J. Med. Food 2014, 17, 599–605. [Google Scholar] [CrossRef] [PubMed]
- Huo, Q.; Shi, Y.; Qi, Y.; Huang, L.; Sui, H.; Zhao, L. Biomimetic silibinin-loaded macrophage-derived exosomes induce dual inhibition of Aβ aggregation and astrocyte activation to alleviate cognitive impairment in a model of Alzheimer’s disease. Mater. Sci. Eng. C Mater. Biol. Appl. 2021, 129, 112365. [Google Scholar] [CrossRef] [PubMed]
- Leem, E.; Oh, Y.S.; Shin, W.H.; Jin, B.K.; Jeong, J.Y.; Shin, M.; Kim, D.W.; Jang, J.H.; Kim, H.J.; Ha, C.M.; et al. Effects of silibinin against prothrombin Kringle-2-induced neurotoxicity in the nigrostriatal dopaminergic system In vivo. J. Med. Food 2019, 22, 277–285. [Google Scholar] [CrossRef]
- Ramírez-Carreto, R.J.; Zaldívar-Machorro, V.J.; Pérez-Ramírez, D.J.; Rodríguez-López, B.E.; Meza, C.; García, E.; Santamaría, A.; Chavarría, A. Oral Administration of Silybin Protects Against MPTP-Induced Neurotoxicity by Reducing Pro-inflammatory Cytokines and Preserving BDNF Levels in Mice. Mol. Neurobiol. 2023, in press. [Google Scholar] [CrossRef] [PubMed]
- Karim, A.; Anwar, F.; Saleem, U.; Fatima, S.; Ismail, T.; Obaidullah, A.J.; Khayat, R.O.; Alqahtani, M.J.; Alsharif, I.; Khan, H.; et al. Administration of α-lipoic acid and silymarin attenuates aggression by modulating endocrine, oxidative stress and inflammatory pathways in mice. Metab. Brain Dis. 2023, 38, 2255–2267. [Google Scholar] [CrossRef]
- Habotta, O.; Ateya, A.; Saleh, R.M.; El-Ashry, E.S. Thiamethoxam evoked neural oxido-inflammatory stress in male rats through modulation of Nrf2/NF-kB/iNOS signaling and inflammatory cytokines: Neuroprotective effect of Silymarin. Neurotoxicology 2023, 96, 28–36. [Google Scholar] [CrossRef] [PubMed]
- Haddadi, R.; Eyvari-Brooshghalan, S.; Makhdoomi, S.; Fadaiie, A.; Komaki, A.; Daneshvar, A. Neuroprotective effects of silymarin in 3-nitropropionic acid-induced neurotoxicity in male mice: Improving behavioral deficits by attenuating oxidative stress and neuroinflammation. Naunyn Schmiedebergs Arch. Pharmacol. 2023, in press. [Google Scholar] [CrossRef]
- Herrock, O.; Deer, E.; LaMarca, B. Setting a stage: Inflammation during preeclampsia and postpartum. Front. Physiol. 2023, 14, 1130116. [Google Scholar] [CrossRef]
- Alston, M.C.; Redman, L.M.; Sones, J.L. An Overview of Obesity, Cholesterol, and Systemic Inflammation in Preeclampsia. Nutrients 2022, 14, 2087. [Google Scholar] [CrossRef]
- Giorgi, V.S.; Peracoli, M.T.; Peracoli, J.C.; Witkin, S.S.; Bannwart-Castro, C.F. Silibinin modulates the NF-κB pathway and pro-inflammatory cytokine production by mononuclear cells from preeclamptic women. J. Reprod. Immunol. 2012, 95, 67–72. [Google Scholar] [CrossRef]
- Cristofalo, R.; Bannwart-Castro, C.F.; Magalhães, C.G.; Borges, V.T.; Peraçoli, J.C.; Witkin, S.S.; Peraçoli, M.T. Silibinin attenuates oxidative metabolism and cytokine production by monocytes from preeclamptic women. Free Rad. Res. 2013, 47, 268–275. [Google Scholar] [CrossRef]
- Lim, R.; Morwood, C.J.; Barker, G.; Lappas, M. Effect of silibinin in reducing inflammatory pathways in in vitro and in vivo models of infection-induced preterm birth. PLoS ONE 2014, 9, e92505. [Google Scholar] [CrossRef] [PubMed]
- Ribeiro, V.R.; Romao-Veiga, M.; Nunes, P.R.; de Oliveira, L.; Romagnoli, G.G.; Peracoli, J.C.; Peracoli, M. Silibinin downregulates the expression of the Th1 and Th17 profiles by modulation of STATs and transcription factors in pregnant women with preeclampsia. Int. Immunopharmacol. 2022, 109, 108807. [Google Scholar] [CrossRef] [PubMed]
- Gluba-Brzózka, A.; Franczyk, B.; Rysz-Górzyńska, M.; Rokicki, R.; Koziarska-Rościszewska, M.; Rysz, J. Pathomechanisms of Immunological Disturbances in β-Thalassemia. Int. J. Mol. Sci. 2021, 22, 9677. [Google Scholar] [CrossRef] [PubMed]
- Balouchi, S.; Gharagozloo, M.; Esmaeil, N.; Mirmoghtadaei, M.; Moayedi, B. Serum levels of TGFβ, IL-10, IL-17, and IL-23 cytokines in β-thalassemia major patients: The impact of silymarin therapy. Immunopharmacol. Immunotoxicol. 2014, 36, 271–274. [Google Scholar] [CrossRef] [PubMed]
- Alidoost, F.; Gharagozloo, M.; Bagherpour, B.; Jafarian, A.; Sajjadi, S.E.; Hourfar, H.; Moayedi, B. Effects of silymarin on the proliferation and glutathione levels of peripheral blood mononuclear cells from beta-thalassemia major patients. International Immunopharmacology 2006, 6, 1305–1310. [Google Scholar] [CrossRef] [PubMed]
- Darvishi-Khezri, H.; Kosaryan, M.; Karami, H.; Salehifar, E.; Mahdavi, M.; Alipour, A.; Aliasgharian, A. Can use of silymarin improve inflammatory status in patients with β-Thalassemia Major? A crossover, randomized controlled trial. Complement. Med. Res. 2021, 28, 123–130. [Google Scholar] [CrossRef]
- Uçmak, F.; Ekin, N.; İbiloğlu, İ.; Arslan, S.; Kaplan, İ.; Şenateş, E. Prophylactic administration of silybin ameliorates L-Arginine-induced acute pancreatitis. Med. Sci. Monit. 2016, 22, 3641–3646. [Google Scholar] [CrossRef]
- Song, J.; Pan, W.; Sun, Y.; Han, J.; Shi, W.; Liao, W. Aspergillus fumigatus-induced early inflammatory response in pulmonary microvascular endothelial cells: Role of p38 MAPK and inhibition by silibinin. Int. Immunopharmacol. 2017, 49, 195–202. [Google Scholar] [CrossRef]
- Ali, S.A.; Saifi, M.A.; Godugu, C.; Talla, V. Silibinin alleviates silica-induced pulmonary fibrosis: Potential role in modulating inflammation and epithelial-mesenchymal transition. Phytother. Res. 2021, 35, 5290–5304. [Google Scholar] [CrossRef]
- Hussain, S.A.; Jassim, N.A.; Numan, I.T.; Al-Khalifa, I.I.; Abdullah, T.A. Anti-inflammatory activity of silymarin in patients with knee osteoarthritis. A comparative study with piroxicam and meloxicam. Saudi Med. J. 2009, 30, 98–103. [Google Scholar] [PubMed]
- Arafa Keshk, W.; Zahran, S.M.; Katary, M.A.; Abd-Elaziz Ali, D. Modulatory effect of silymarin on nuclear factor-erythroid-2-related factor 2 regulated redox status, nuclear factor-κB mediated inflammation and apoptosis in experimental gastric ulcer. Chem. Biol. Interact. 2017, 273, 266–272. [Google Scholar] [CrossRef] [PubMed]
- Esmaily, H.; Vaziri-Bami, A.; Miroliaee, A.E.; Baeeri, M.; Abdollahi, M. The correlation between NF-κB inhibition and disease activity by coadministration of silibinin and ursodeoxycholic acid in experimental colitis. Fundam. Clin. Pharmacol. 2011, 25, 723–733. [Google Scholar] [CrossRef] [PubMed]
- Safarpour, S.; Safarpour, S.; Moghadamnia, A.A.; Kazemi, S.; Ebrahimpour, A.; Shirafkan, F. The protective effects of silymarin nanoemulsion on 5-fluorouracil-induced gastrointestinal toxicity in rats. Saudi Pharm. J. 2023, 31, 101672. [Google Scholar] [CrossRef] [PubMed]
- Akhtar, M.; Chen, Y.; Ma, Z.; Zhang, X.; Shi, D.; Khan, J.A.; Liu, H. Gut microbiota-derived short chain fatty acids are potential mediators in gut inflammation. Anim. Nutr. 2021, 8, 350–360. [Google Scholar] [CrossRef] [PubMed]
- Oryan, A.; Tabatabaei Naeini, A.; Moshiri, A.; Mohammadalipour, A.; Tabandeh, M.R. Modulation of cutaneous wound healing by silymarin in rats. J. Wound Care 2012, 21, 457–464. [Google Scholar] [CrossRef]
- Sharifi, R.; Rastegar, H.; Kamalinejad, M.; Dehpour, A.R.; Tavangar, S.M.; Paknejad, M.; Mehrabani Natanzi, M.; Ghannadian, N.; Akbari, M.; Pasalar, P. Effect of topical application of silymarin (Silybum marianum) on excision wound healing in albino rats. Acta Med. Iran. 2012, 50, 583–588. [Google Scholar]
- Mahmoodi-Nesheli, M.; Alizadeh, S.; Solhi, H.; Mohseni, J.; Mahmoodi-Nesheli, M. Adjuvant effect of oral silymarin on patients’ wound healing process caused by thermal injuries. Casp. J. Intern. Med. 2018, 9, 341–346. [Google Scholar]
- Liu, X.; Xu, Q.; Long, X.; Liu, W.; Zhao, Y.; Hayashi, T.; Hattori, S.; Fujisaki, H.; Ogura, T.; Tashiro, S.I.; et al. Silibinin-induced autophagy mediated by PPARα-sirt1-AMPK pathway participated in the regulation of type I collagen-enhanced migration in murine 3T3-L1 preadipocytes. Mol. Cell. Biochem. 2019, 450, 1–23. [Google Scholar] [CrossRef]
- Soleimani, H.; Behfar, M.; Hobbenaghi, R. Tenogenic effects of silymarin following experimental Achilles tendon transection in rats. Vet. Res. Forum 2021, 12, 69–75. [Google Scholar]
- Gupta, S.C.; Tyagi, A.K.; Deshmukh-Taskar, P.; Hinojosa, M.; Prasad, S.; Aggarwal, B.B. Downregulation of tumor necrosis factor and other proinflammatory biomarkers by polyphenols. Arch. Biochem. Biophys. 2014, 559, 91–99. [Google Scholar] [CrossRef] [PubMed]
- Stevenson, D.E.; Hurst, R.D. Polyphenolic phytochemicals–just antioxidants or much more? Cell. Mol. Life Sci. 2007, 64, 2900–2916. [Google Scholar] [CrossRef] [PubMed]
- Manna, S.K.; Mukhopadhyay, A.; Van, N.T.; Aggarwal, B.B. Silymarin suppresses TNF-induced activation of NF-κB, c-Jun N-terminal kinase, and apoptosis. J. Immunol. 1999, 163, 6800–6809. [Google Scholar] [CrossRef] [PubMed]
- Gharagozloo, M.; Velardi, E.; Bruscoli, S.; Agostini, M.; Di Sante, M.; Donato, V.; Amirghofran, Z.; Riccardi, C. Silymarin suppress CD4+ T cell activation and proliferation: Effects on NF-kappaB activity and IL-2 production. Pharm. Res. 2010, 61, 405–409. [Google Scholar] [CrossRef] [PubMed]
- Oskoueian, E.; Abdullah, N.; Idrus, Z.; Ebrahimi, M.; Goh, Y.M.; Shakeri, M.; Oskoueian, A. Palm kernel cake extract exerts hepatoprotective activity in heat-induced oxidative stress in chicken hepatocytes. BMC Complement. Altern. Med. 2014, 14, 368. [Google Scholar] [CrossRef] [PubMed]
- Kang, D.Y.; Sp, N.; Do Park, K.; Lee, H.K.; Song, K.D.; Yang, Y.M. Silibinin inhibits in vitro ketosis by regulating HMGCS2 and NF-κB: Elucidation of signaling molecule relationship under ketotic conditions. In vitro cellular & developmental biology. Animal 2019, 55, 368–375. [Google Scholar]
- Xie, Y.; Feng, S.L.; Mai, C.T.; Zheng, Y.F.; Wang, H.; Liu, Z.Q.; Zhou, H.; Liu, L. Suppression of up-regulated LXRα by silybin ameliorates experimental rheumatoid arthritis and abnormal lipid metabolism. Phytomedicine 2021, 80, 153339. [Google Scholar] [CrossRef]
- Baldini, F.; Portincasa, P.; Grasselli, E.; Damonte, G.; Salis, A.; Bonomo, M.; Florio, M.; Serale, N.; Voci, A.; Gena, P.; et al. Aquaporin-9 is involved in the lipid-lowering activity of the nutraceutical silybin on hepatocytes through modulation of autophagy and lipid droplets composition. Biochim. Biophys. Acta Mol. Cell. Biol. Lipids 2020, 1865, 158586. [Google Scholar] [CrossRef]
- Tian, M.Y.; Fan, J.H.; Zhuang, Z.W.; Dai, F.; Wang, C.Y.; Hou, H.T.; Ma, Y.Z. Effects of silymarin on p65 NF-κB, p38 MAPK and CYP450 in LPS-induced hoof dermal inflammatory cells of dairy cows. BMC Vet. Res. 2019, 15, 127. [Google Scholar] [CrossRef]
- Prack Mc Cormick, B.; Langle, Y.; Belgorosky, D.; Vanzulli, S.; Balarino, N.; Sandes, E.; Eiján, A.M. Flavonoid silybin improves the response to radiotherapy in invasive bladder cancer. J. Cell. Biochem. 2018, 119, 5402–5412. [Google Scholar] [CrossRef]
- Liu, X.; Xu, Q.; Liu, W.; Yao, G.; Zhao, Y.; Xu, F.; Hayashi, T.; Fujisaki, H.; Hattori, S.; Tashiro, S.I.; et al. Enhanced migration of murine fibroblast-like 3T3-L1 preadipocytes on type I collagen-coated dish is reversed by silibinin treatment. Mol. Cell. Biochem. 2018, 441, 35–62. [Google Scholar] [CrossRef]
- Henriet, E.; Abdallah, F.; Laurent, Y.; Guimpied, C.; Clement, E.; Simon, M.; Pichon, C.; Baril, P. Targeting TGF-β1/miR-21 Pathway in Keratinocytes Reveals Protective Effects of Silymarin on Imiquimod-Induced Psoriasis Mouse Model. JID Innov. 2022, 3, 100175. [Google Scholar] [CrossRef] [PubMed]
- Wang, Q.; Zou, L.; Liu, W.; Hao, W.; Tashiro, S.I.; Onodera, S.; Ikejima, T. Inhibiting NF-κB activation and ROS production are involved in the mechanism of silibinin’s protection against D-galactose-induced senescence. Pharmacol. Biochem. Behav. 2011, 98, 140–149. [Google Scholar] [CrossRef] [PubMed]
- Prabu, S.M.; Muthumani, M. Silibinin ameliorates arsenic induced nephrotoxicity by abrogation of oxidative stress, inflammation and apoptosis in rats. Mol. Biol. Rep. 2012, 39, 11201–11216. [Google Scholar] [CrossRef] [PubMed]
- Salamone, F.; Galvano, F.; Cappello, F.; Mangiameli, A.; Barbagallo, I.; Li Volti, G. Silibinin modulates lipid homeostasis and inhibits nuclear factor kappa B activation in experimental nonalcoholic steatohepatitis. Transl. Res. 2012, 59, 477–486. [Google Scholar] [CrossRef]
- Li, C.C.; Hsiang, C.Y.; Wu, S.L.; Ho, T.Y. Identification of novel mechanisms of silymarin on the carbon tetrachloride-induced liver fibrosis in mice by nuclear factor-κB bioluminescent imaging-guided transcriptomic analysis. Food Chem. Toxicol. 2012, 50, 1568–1575. [Google Scholar] [CrossRef]
- Liu, B.N.; Han, B.X.; Liu, F. Neuroprotective effect of pAkt and HIF-1α on ischemia rats. Asian Pac. J. Trop. Med. 2014, 7, 221–225. [Google Scholar] [CrossRef]
- Aristatile, B.; Al-Assaf, A.H.; Pugalendi, K.V. Carvacrol suppresses the expression of inflammatory marker genes in D-galactosamine-hepatotoxic rats. Asian Pac. J. Trop. Med. 2013, 6, 205–211. [Google Scholar] [CrossRef]
- Salama, S.M.; Abdulla, M.A.; Al Rashdi, A.S.; Hadi, A.H.A. Mechanism of hepatoprotective effect of Boesenbergia rotunda in thioacetamide-induced liver damage in rats. Evid. Based Complement. Alternat. Med. 2013, 2013, 157456. [Google Scholar] [CrossRef]
- Abhilash, P.A.; Harikrishnan, R.; Indira, M. Ascorbic acid is superior to silymarin in the recovery of ethanol-induced inflammatory reactions in hepatocytes of guinea pigs. J. Physiol. Biochem. 2013, 69, 785–798. [Google Scholar] [CrossRef]
- Abhilash, P.A.; Harikrishnan, R.; Indira, M. Ascorbic acid suppresses endotoxemia and NF-κB signaling cascade in alcoholic liver fibrosis in guinea pigs: A mechanistic approach. Toxicol. Appl. Pharmacol. 2014, 274, 215–224. [Google Scholar] [CrossRef] [PubMed]
- Sharma, S.; Le Guillou, D.; Chen, J.Y. Cellular stress in the pathogenesis of nonalcoholic steatohepatitis and liver fibrosis. Nat. Rev. Gastroenterol. Hepatol. 2023, 20, 662–678. [Google Scholar] [CrossRef] [PubMed]
- Liu, K.; Zhou, S.; Liu, J.; Wang, Y.; Zhu, F.; Liu, M. Silibinin attenuates high-fat diet-induced renal fibrosis of diabetic nephropathy. Drug Des. Devel. Ther. 2019, 13, 3117–3126. [Google Scholar] [CrossRef] [PubMed]
- Ali, N.M.; Mahmoud, A.A.A.; Mahmoud, M.F.; El Fayoumi, H.M. Glycyrrhizic acid and silymarin alleviate the neurotoxic effects of aluminum in rats challenged with fructose-induced insulin resistance: Possible role of toll-like receptor 4 pathway. Drug Chem. Toxicol. 2019, 42, 210–219. [Google Scholar] [CrossRef] [PubMed]
- El-Marasy, S.A.; Abd-Elsalam, R.M.; Ahmed-Farid, O.A. Ameliorative Effect of Silymarin on Scopolamine-induced Dementia in Rats. Open Access Maced. J. Med. Sci. 2018, 6, 1215–1224. [Google Scholar] [CrossRef]
- Yuan, R.; Fan, H.; Cheng, S.; Gao, W.; Xu, X.; Lv, S.; Ye, M.; Wu, M.; Zhu, X.; Zhang, Y. Silymarin prevents NLRP3 inflammasome activation and protects against intracerebral hemorrhage. Biomed. Pharmacother. 2017, 93, 308–315. [Google Scholar] [CrossRef]
- Tian, L.; Li, W.; Wang, T. Therapeutic effects of silibinin on LPS-induced acute lung injury by inhibiting NLRP3 and NF-κB signaling pathways. Microb. Pathog. 2017, 108, 104–108. [Google Scholar] [CrossRef]
- Marcolino Assis-Júnior, E.; Melo, A.T.; Pereira, V.B.M.; Wong, D.V.T.; Sousa, N.R.P.; Oliveira, C.M.G.; Malveira, L.R.C.; Moreira, L.S.; Souza, M.H.L.P.; Almeida, P.R.C.; et al. Dual effect of silymarin on experimental non-alcoholic steatohepatitis induced by irinotecan. Toxicol. Appl. Pharmacol. 2017, 327, 71–79. [Google Scholar] [CrossRef]
- Liu, P.; Chen, W.; Kang, Y.; Wang, C.; Wang, X.; Liu, W.; Hayashi, T.; Qiu, Z.; Mizuno, K.; Hattori, S.; et al. Silibinin ameliorates STING-mediated neuroinflammation via downregulation of ferroptotic damage in a sporadic Alzheimer’s disease model. Arch. Biochem. Biophys. 2023, 744, 109691. [Google Scholar] [CrossRef]
- Karimi, R.; Bakhshi, A.; Dayati, P.; Abazari, O.; Shahidi, M.; Savaee, M.; Kafi, E.; Rahmanian, M.; Naghib, S.M. Silymarin reduces retinal microvascular damage in streptozotocin-induced diabetic rats. Sci. Rep. 2022, 12, 15872. [Google Scholar] [CrossRef]
- Menzel, A.; Samouda, H.; Dohet, F.; Loap, S.; Ellulu, M.S.; Bohn, T. Common and novel markers for measuring inflammation and oxidative stress Ex Vivo in research and clinical practice-Which to use regarding disease outcomes? Antioxidants 2021, 10, 414. [Google Scholar] [CrossRef] [PubMed]
- Cardoso Dal Pont, G.; Farnell, M.; Farnell, Y.; Kogut, M.H. Dietary factors as triggers of low-grade chronic intestinal inflammation in poultry. Microorganisms 2020, 8, 139. [Google Scholar] [CrossRef]
- Kogut, M.H.; Genovese, K.; Swaggerty, C.L.; He, H.; Broom, L. Inflammatory phenotypes in the intestine of poultry: Not all inflammation is created equal. Poult. Sci. 2018, 97, 2339–2346. [Google Scholar] [CrossRef] [PubMed]
- Ducatelle, R.; Goossens, E.; Eeckhaut, V.; Van Immerseel, F. Poultry gut health and beyond. Anim. Nutr. 2023, 13, 240–248. [Google Scholar] [CrossRef] [PubMed]
- Tellez-Isaias, G.; Eisenreich, W.; Petrone-Garcia, V.M.; Hernandez-Velasco, X.; Castellanos-Huerta, I.; Tellez, G., Jr.; Latorre, J.D.; Bottje, W.G.; Senas-Cuesta, R.; Coles, M.E.; et al. Effects of chronic stress and intestinal inflammation on commercial poultry health and performance: A review. Ger. J. Vet. Res. 2023, 3, 38–57. [Google Scholar] [CrossRef]
- Gusev, E.; Zhuravleva, Y. Inflammation: A new look at an old problem. Int. J. Mol. Sci. 2022, 23, 4596. [Google Scholar] [CrossRef]
- Ralli, T.; Tripathi, T.; Kalaiselvan, V.; Tiwari, R.; Aeri, V.; Kohli, K. Silymarin as a Phyto-pharmaceutical: Isolation, Simultaneous Quantification of four Biomarkers and in-silico Anti-inflammatory Activity. Chin. J. Anal. Chem. 2023, 51, 100174. [Google Scholar] [CrossRef]
- Yu, C.; Wang, D.; Yang, Z.; Wang, T. Pharmacological effects of polyphenol phytochemicals on the intestinal inflammation via targeting TLR4/NF-κB signaling pathway. Int. J. Mol. Sci. 2022, 23, 6939. [Google Scholar] [CrossRef]
- DuBreuil, D.M.; Lai, X.; Zhu, K.; Chahyadinata, G.; Perner, C.; Chiang, B.M.; Battenberg, A.; Sokol, C.L.; Wainger, B.J. Phenotypic screen identifies the natural product silymarin as a novel anti-inflammatory analgesic. Mol. Pain 2023, 19, 17448069221148351. [Google Scholar] [CrossRef]
- Ghodousi, M.; Karbasforooshan, H.; Arabi, L.; Elyasi, S. Silymarin as a preventive or therapeutic measure for chemotherapy and radiotherapy-induced adverse reactions: A comprehensive review of preclinical and clinical data. Eur. J. Clin. Pharm. 2023, 79, 5–38. [Google Scholar] [CrossRef]
- Gür, F.M.; Bilgiç, S. Silymarin, an antioxidant flavonoid, protects the liver from the toxicity of the anticancer drug paclitaxel. Tissue Cell 2023, 83, 102158. [Google Scholar] [CrossRef] [PubMed]




{kind=link}
{kind=link}
{kind=link}
| Category | Proinflammatory Factors | |
|---|---|---|
| Human | Poultry | |
| Physical factors | Radiation, UV, hyperthermia, hypothermia, trauma | Hyperthermia, hypothermia, trauma, increased stocking density |
| Chemical factors | Asbestos, heavy metals, organic toxicants, dust, lipopolysaccharides | Heavy metals, mycotoxins, ammonia, CO, dust |
| Biological factors | Bacterial infection, viral infection, fungal infection | Bacterial infection, viral infection, fungal infection |
| Unhealthy lifestyle | Smoking, alcohol, high-calorie diet, stress, sedentary lifestyle | Restricted movement (cage housing), nutrient deficiency |
| Chronic diseases | Obesity, diabetes, hyperglycaemia | Chronic Respiratory Disease |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Surai, P.F.; Surai, A.; Earle-Payne, K. Silymarin and Inflammation: Food for Thoughts. Antioxidants 2024, 13, 98. https://doi.org/10.3390/antiox13010098
Surai PF, Surai A, Earle-Payne K. Silymarin and Inflammation: Food for Thoughts. Antioxidants. 2024; 13(1):98. https://doi.org/10.3390/antiox13010098
Chicago/Turabian StyleSurai, Peter F., Anton Surai, and Katie Earle-Payne. 2024. "Silymarin and Inflammation: Food for Thoughts" Antioxidants 13, no. 1: 98. https://doi.org/10.3390/antiox13010098
APA StyleSurai, P. F., Surai, A., & Earle-Payne, K. (2024). Silymarin and Inflammation: Food for Thoughts. Antioxidants, 13(1), 98. https://doi.org/10.3390/antiox13010098