

Molecular Link between Glo-1 Expression and Markers of Hyperglycemia and Oxidative Stress in Vascular Complications of Type 2 Diabetes Mellitus

Abstract
:1. Introduction
2. Materials and Methods
2.1. In Vitro Model of Dicarbonyl Stress
2.1.1. Cell Culture
2.1.2. siRNA-Mediated Knockdown of Glo-1 Expression
2.1.3. Spectrophotometric Determination of Glo-1 Activity
2.2. In Vivo Model of Dicarbonyl Stress
2.3. Gene Expression
2.4. Data Analysis
3. Results
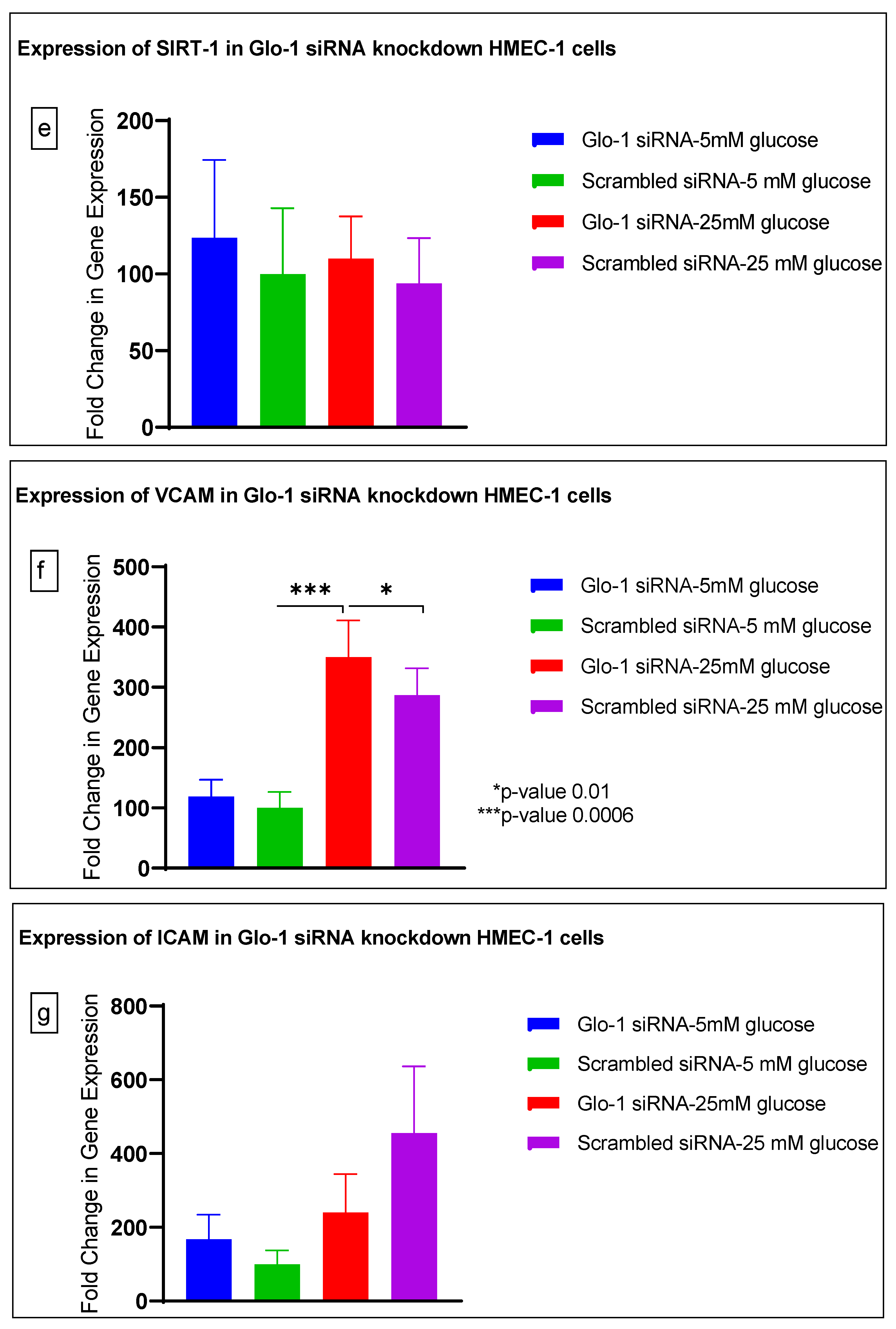
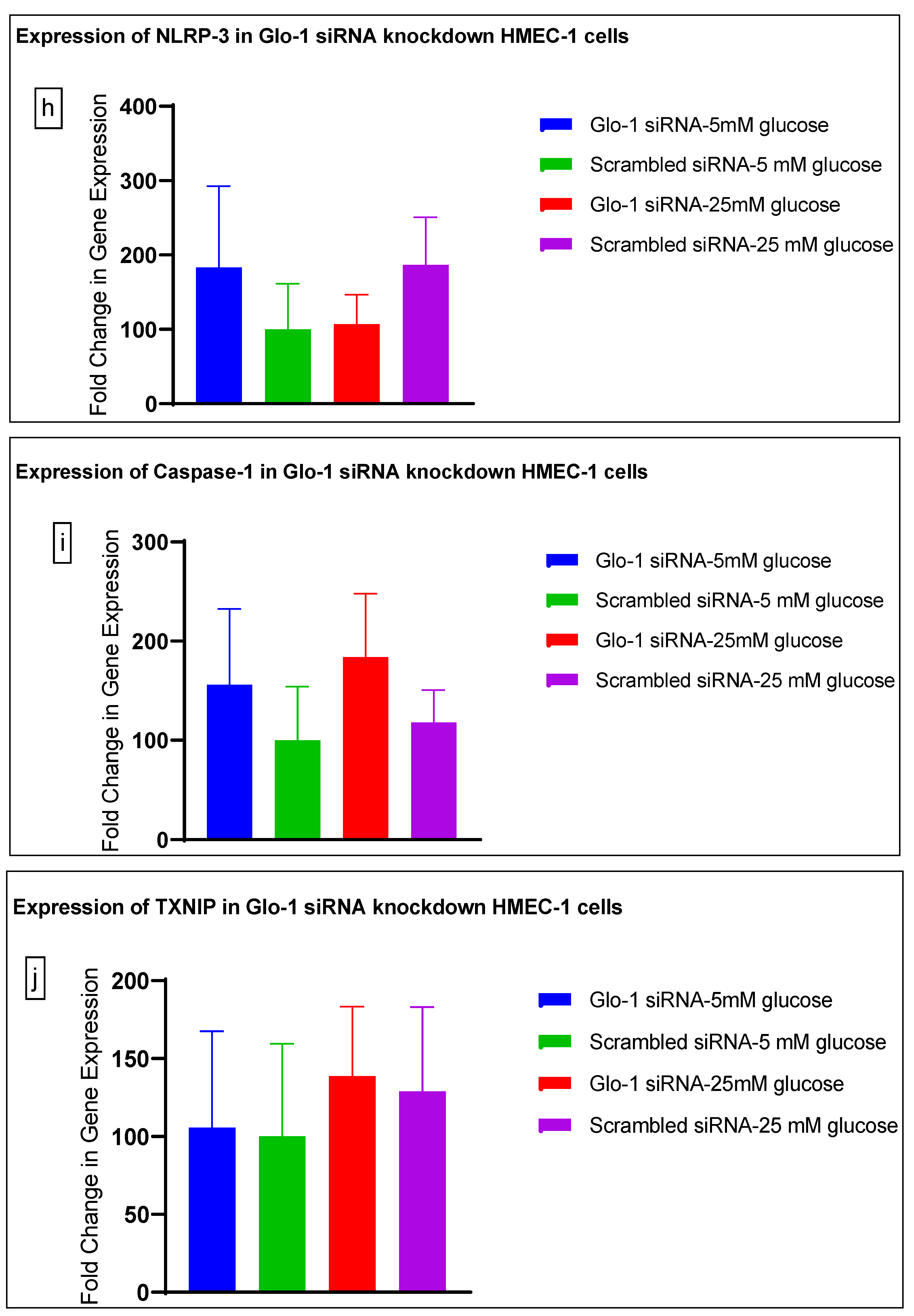
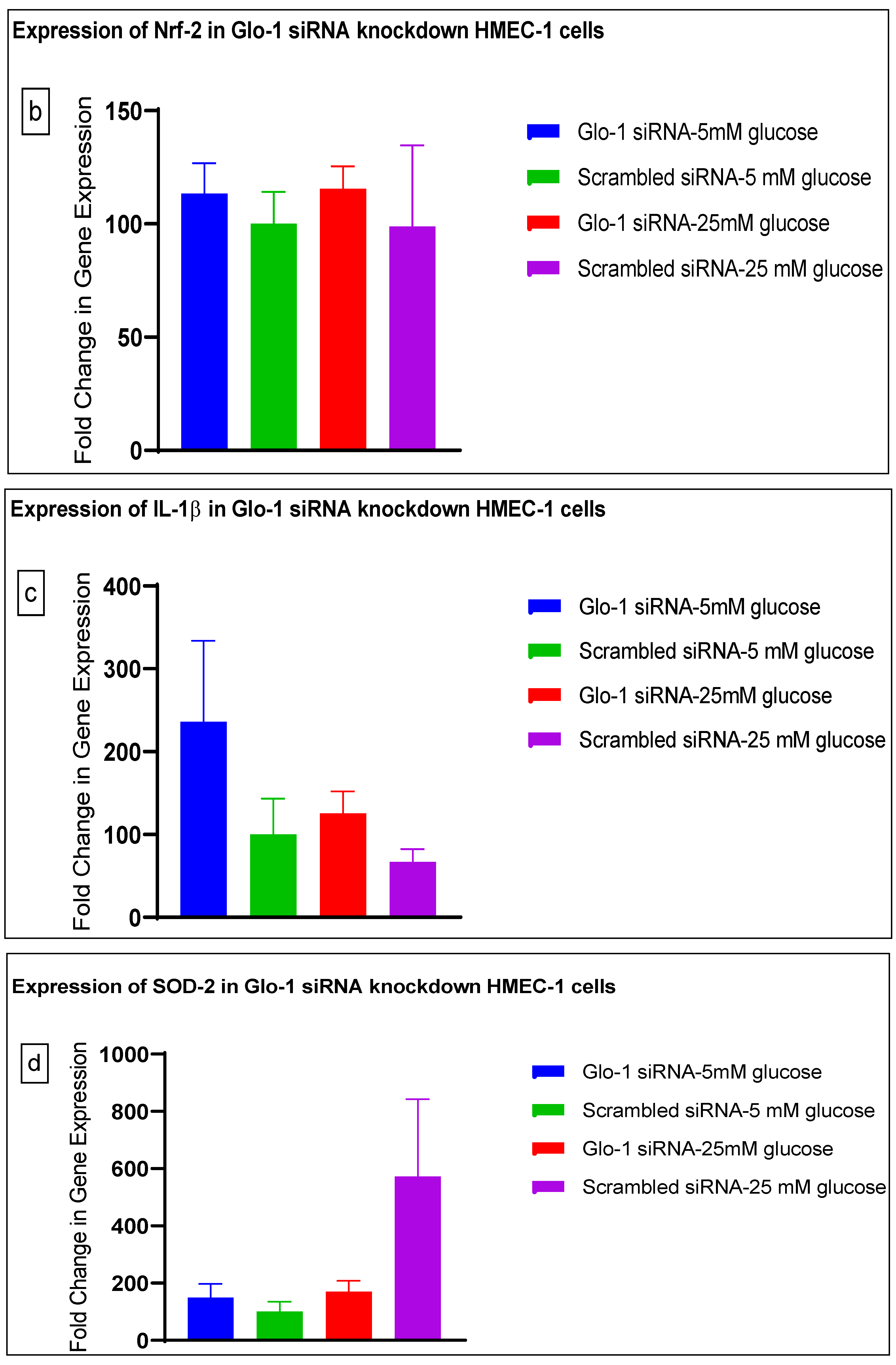
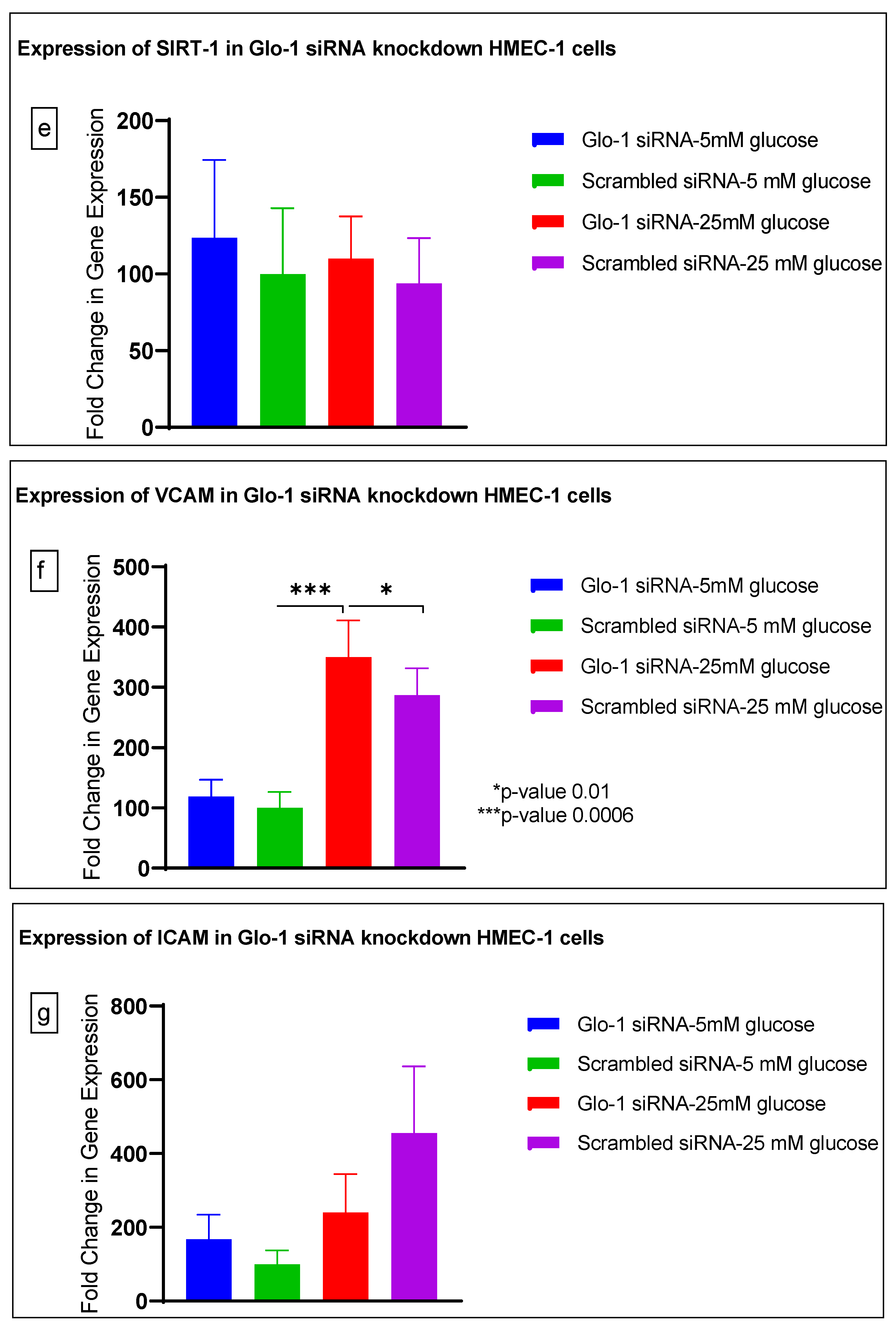
3.1. Effect of siRNA-Mediated Downregulation of Glo-1 on Markers of Hyperglycemia and Oxidative Stress
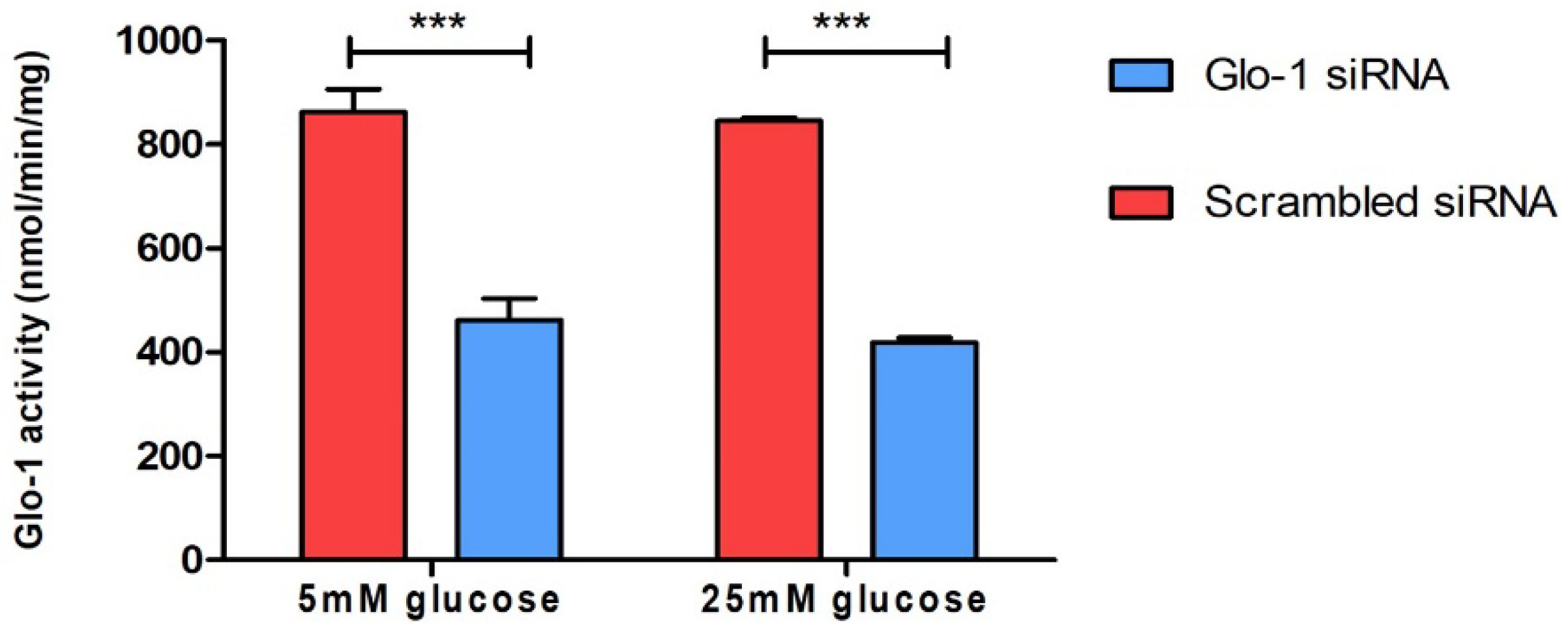
3.1.1. Spectrophotometric Determination of Glyoxalase 1 Activity
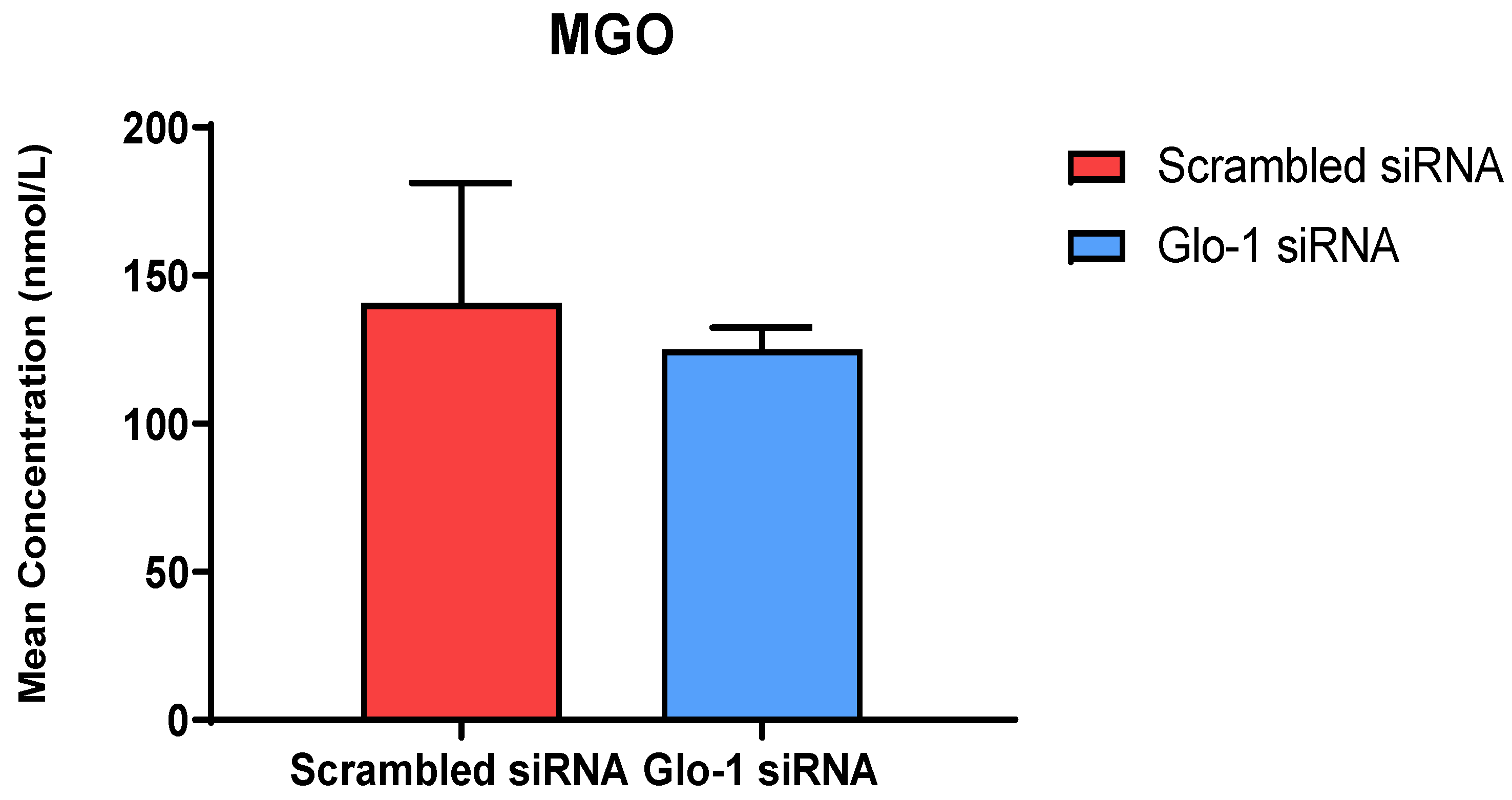
3.1.2. Quantification of MGO and Its Derived Adducts
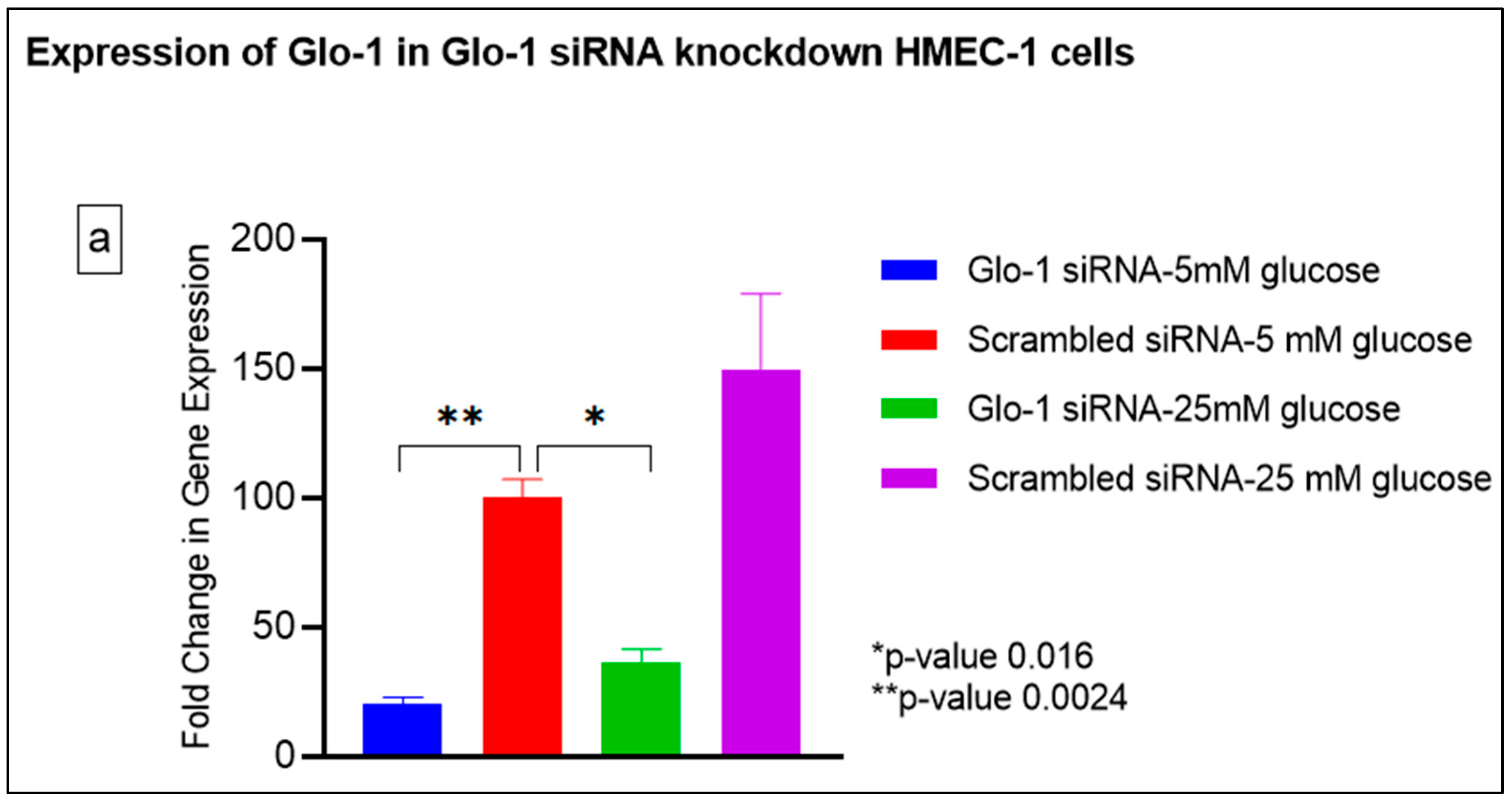
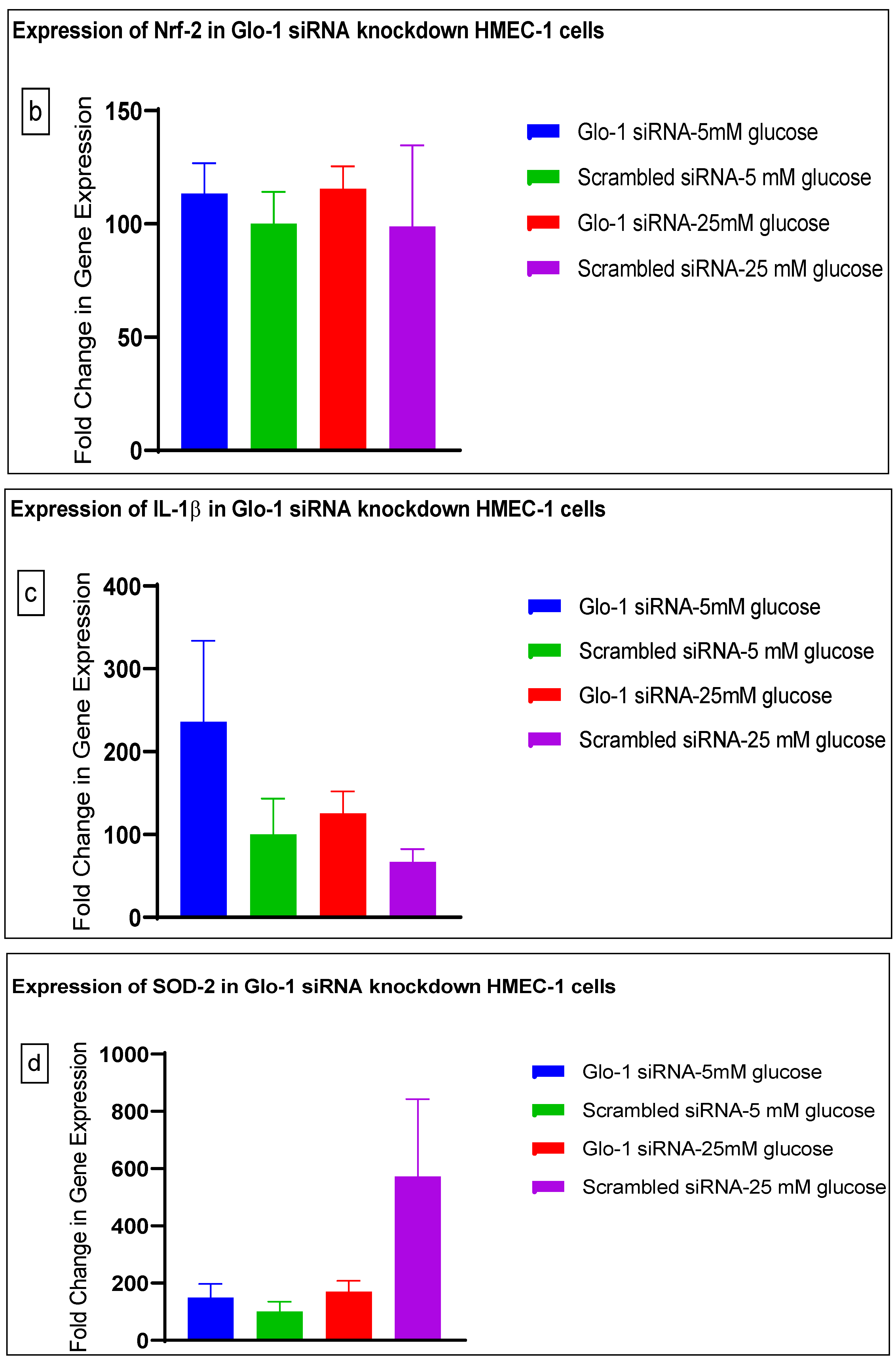
3.1.3. Quantitative Gene Expression of Markers of Hyperglycemia and Oxidative Stress
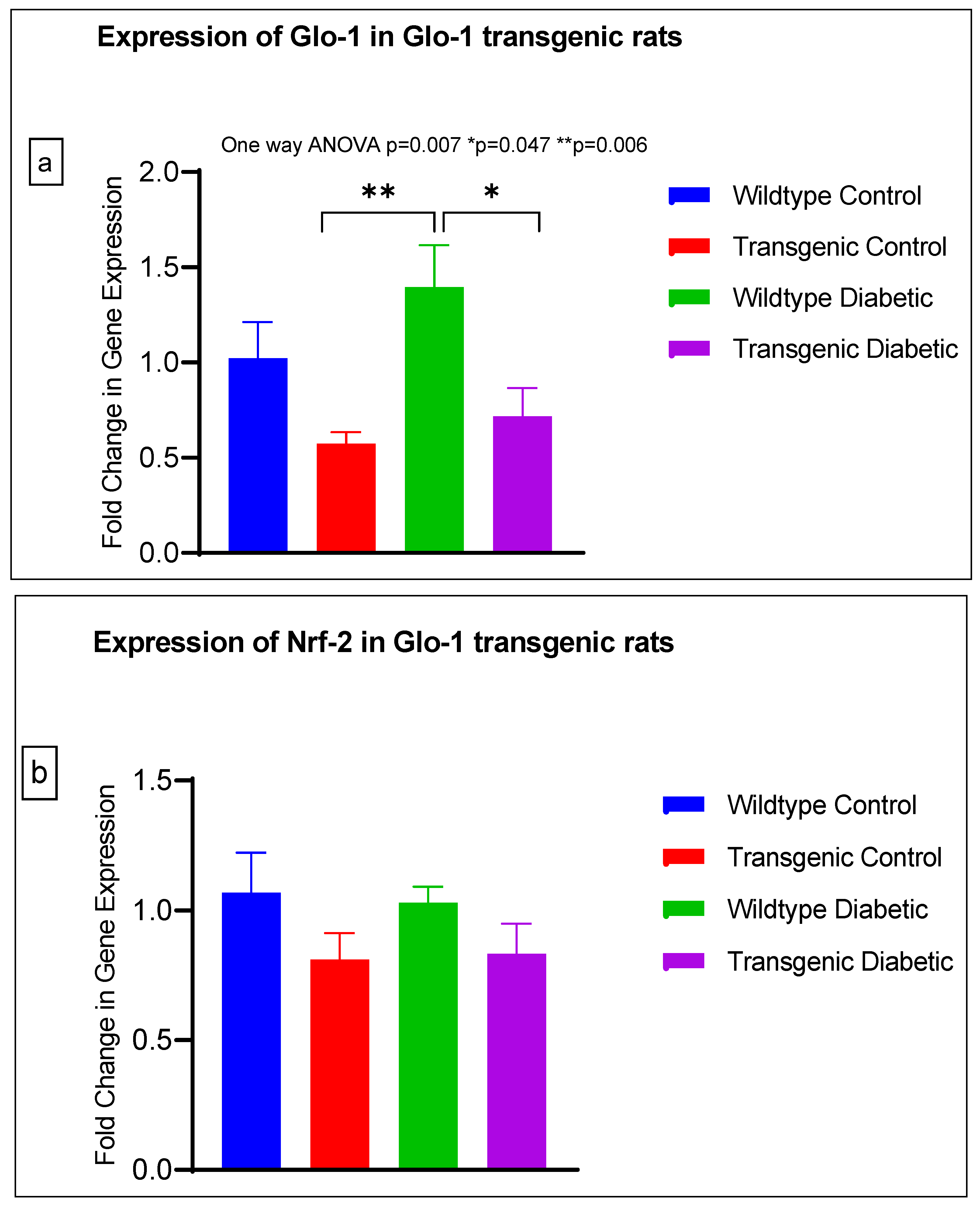
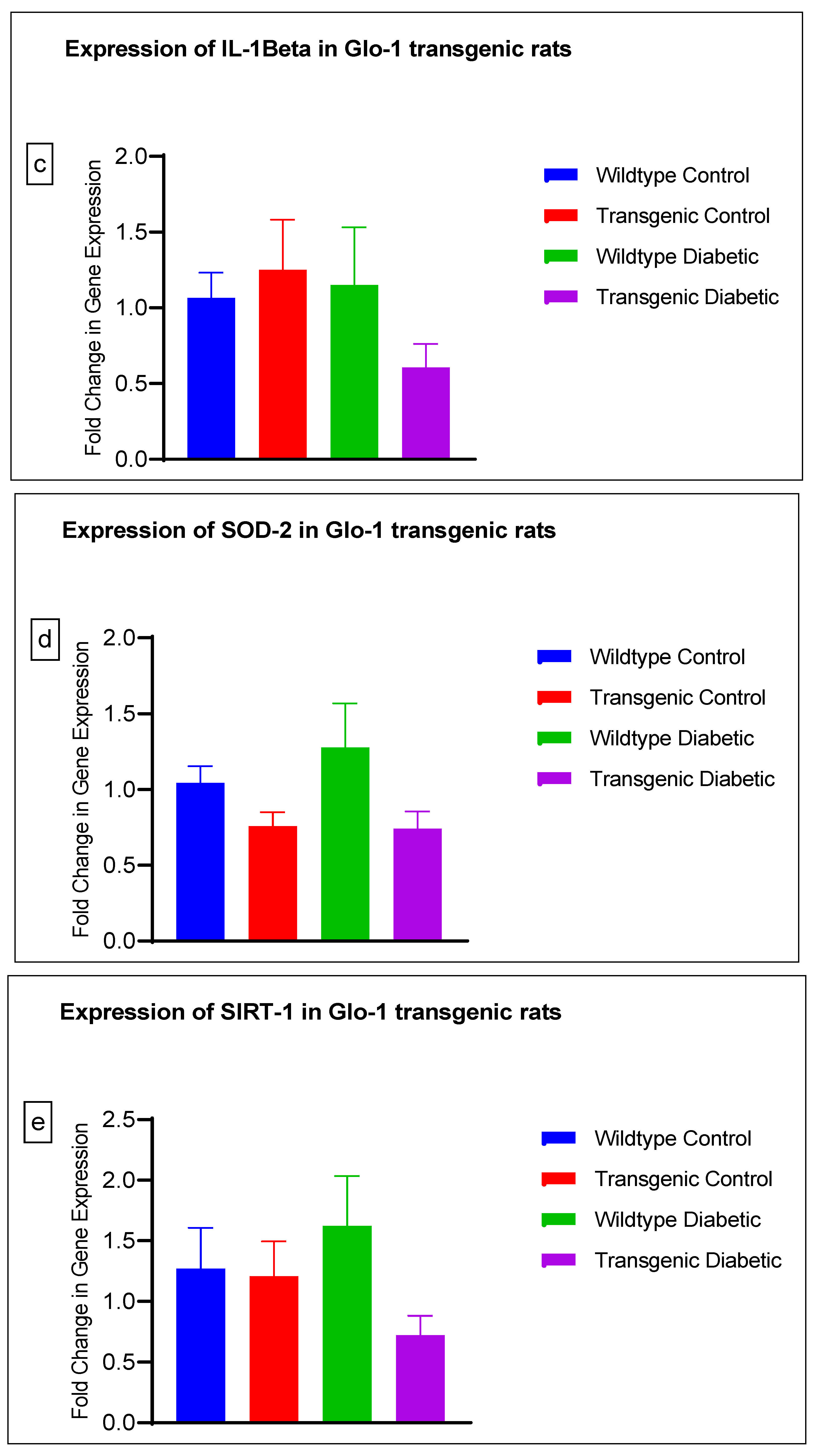
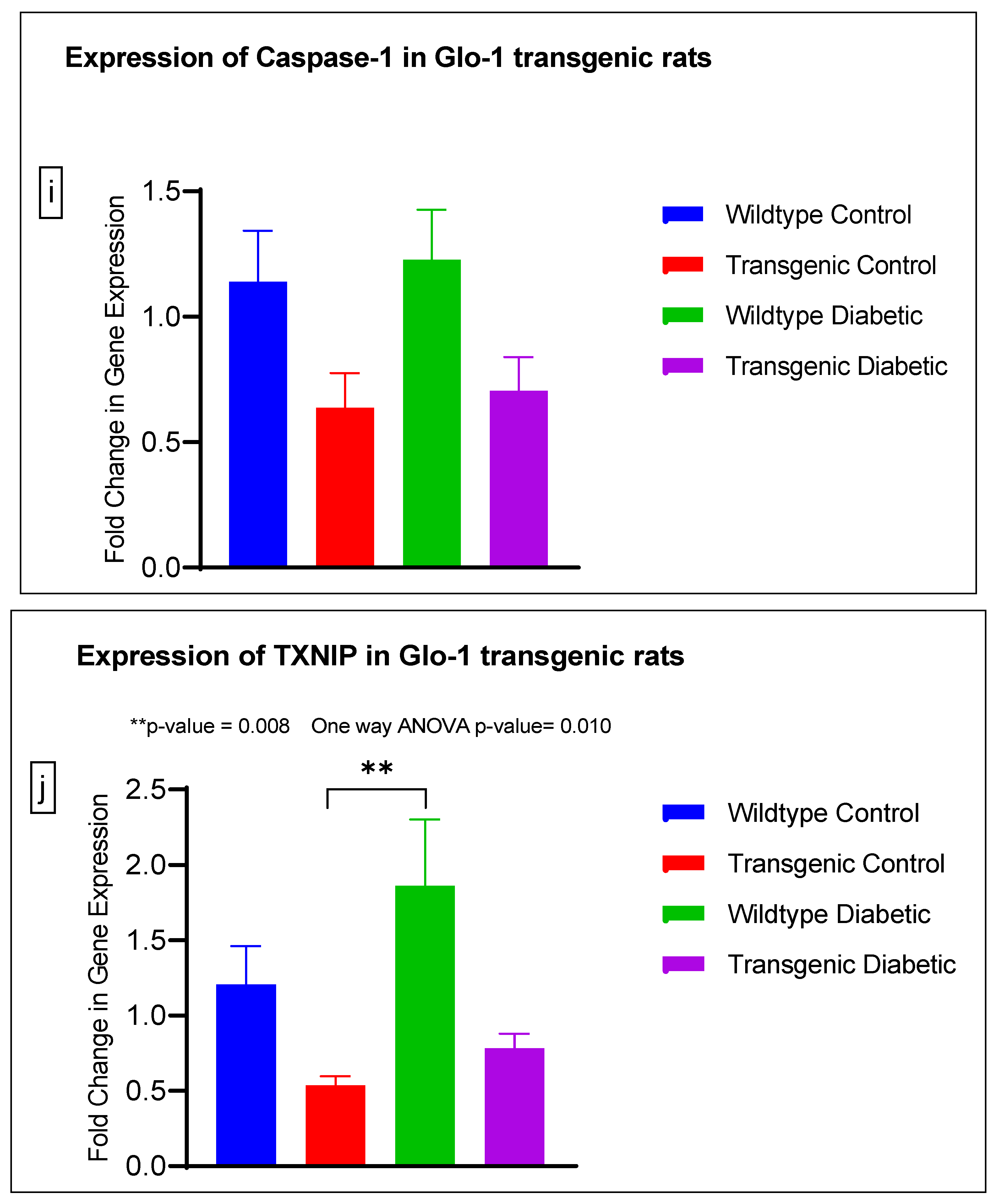
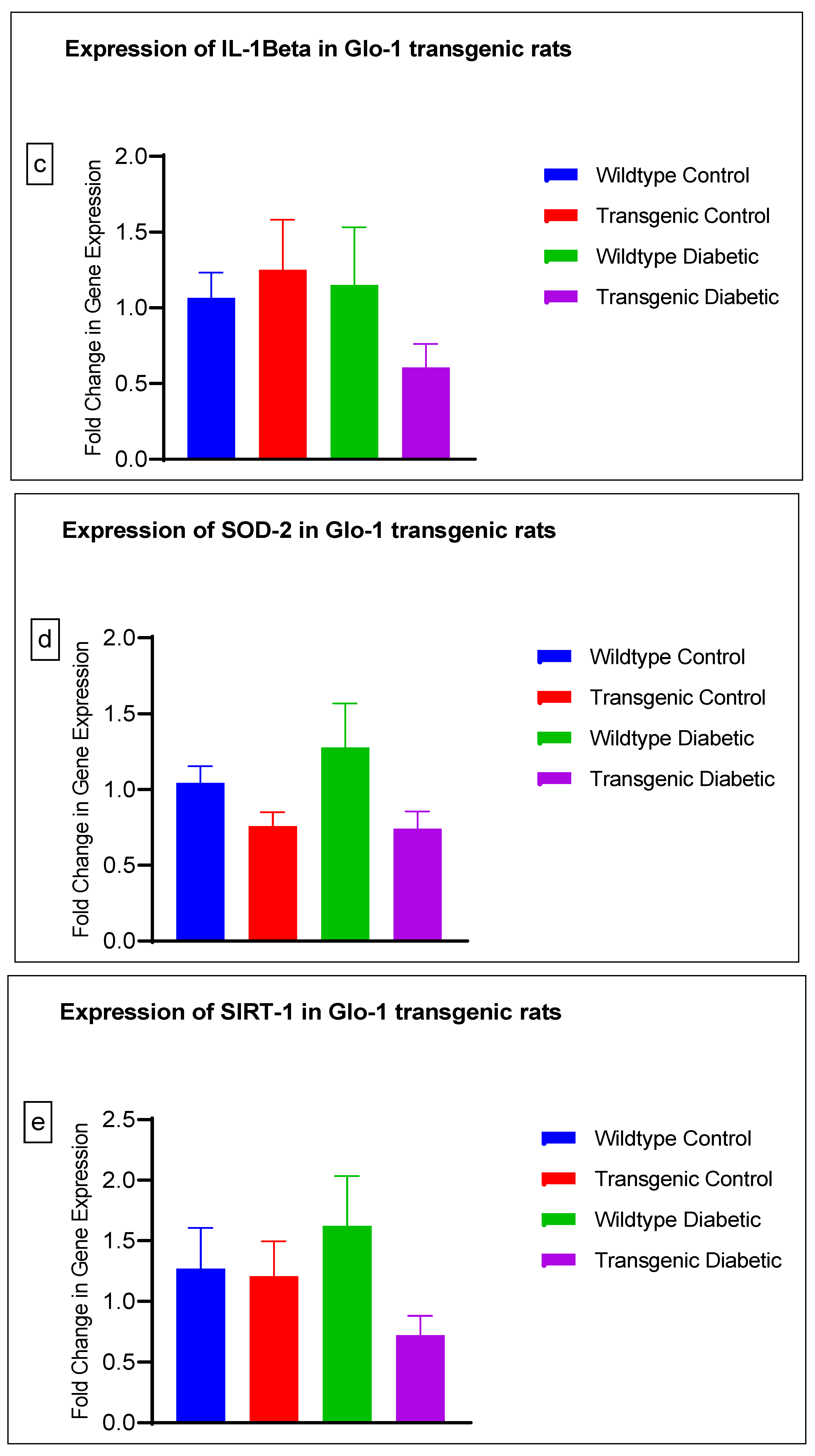
3.2. Effect of Upregulation of Glo-1 Expression on Markers of Hyperglycemia and Oxidative Stress
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Peiró, C.; Lorenzo, Ó.; Carraro, R.; Sánchez-Ferrer, C.F. IL-1β Inhibition in Cardiovascular Complications Associated to Diabetes Mellitus. Front. Pharmacol. 2017, 8, 363. [Google Scholar] [CrossRef] [PubMed]
- Schalkwijk, C.G.; Stehouwer, C.D.A. Methylglyoxal, a Highly Reactive Dicarbonyl Compound, in Diabetes, Its Vascular Complications, and Other Age-Related Diseases. Physiol. Rev. 2020, 100, 407–461. [Google Scholar] [CrossRef] [PubMed]
- Skapare, E.; Konrade, I.; Liepinsh, E.; Makrecka, M.; Zvejniece, L.; Svalbe, B.; Vilskersts, R.; Dambrova, M. Glyoxalase 1 and Glyoxalase 2 Activities in Blood and Neuronal Tissue Samples from Experimental Animal Models of Obesity and Type 2 Diabetes Mellitus. J. Physiol. Sci. 2012, 62, 469–478. [Google Scholar] [CrossRef] [PubMed]
- Gora, I.M.; Ciechanowska, A.; Ladyzynski, P. NLRP3 Inflammasome at the Interface of Inflammation, Endothelial Dysfunction, and Type 2 Diabetes. Cells 2021, 10, 314. [Google Scholar] [CrossRef] [PubMed]
- Irshad, Z.; Xue, M.; Ashour, A.; Larkin, J.R.; Thornalley, P.J.; Rabbani, N. Activation of the Unfolded Protein Response in High Glucose Treated Endothelial Cells Is Mediated by Methylglyoxal. Sci. Rep. 2019, 9, 7889. [Google Scholar] [CrossRef]
- Ott, C.; Jacobs, K.; Haucke, E.; Santos, A.N.; Grune, T.; Simm, A. Role of Advanced Glycation End Products in Cellular Signaling. Redox Biol. 2014, 2, 411–429. [Google Scholar] [CrossRef]
- Syed, N.A.; Bhatti, A.; John, P. Molecular Dynamics Simulations and Bioinformatics’ Analysis of Deleterious Missense Single Nucleotide Polymorphisms in Glyoxalase-1 Gene. J. Biomol. Struct. Dyn. 2023. Online ahead of print. [Google Scholar] [CrossRef]
- Maher, P.; Dargusch, R.; Ehren, J.L.; Okada, S.; Sharma, K.; Schubert, D. Fisetin Lowers Methylglyoxal Dependent Protein Glycation and Limits the Complications of Diabetes. PLoS ONE 2011, 6, e21226. [Google Scholar] [CrossRef]
- Rabbani, N.; Xue, M.; Thornalley, P.J. Chapter 36—Dicarbonyl Stress and the Glyoxalase System. In Oxidative Stress Eustress and Distress; Sies, H.B.T.-O.S., Ed.; Academic Press: Cambridge, MA, USA, 2020; pp. 759–777. ISBN 978-0-12-818606-0. [Google Scholar]
- Nigro, C.; Leone, A.; Fiory, F.; Prevenzano, I.; Nicolò, A.; Mirra, P.; Beguinot, F.; Miele, C. Dicarbonyl Stress at the Crossroads of Healthy and Unhealthy Aging. Cells 2019, 8, 749. [Google Scholar] [CrossRef]
- Chapple, S.J.; Siow, R.C.M.; Mann, G.E. Crosstalk between Nrf2 and the Proteasome: Therapeutic Potential of Nrf2 Inducers in Vascular Disease and Aging. Int. J. Biochem. Cell Biol. 2012, 44, 1315–1320. [Google Scholar] [CrossRef]
- Uruno, A.; Yagishita, Y.; Yamamoto, M. The Keap1–Nrf2 System and Diabetes Mellitus. Arch. Biochem. Biophys. 2015, 566, 76–84. [Google Scholar] [CrossRef] [PubMed]
- Kang, L.; Dai, C.; Lustig, M.E.; Bonner, J.S.; Mayes, W.H.; Mokshagundam, S.; James, F.D.; Thompson, C.S.; Lin, C.-T.; Perry, C.G.; et al. Heterozygous SOD2 Deletion Impairs Glucose-Stimulated Insulin Secretion, but Not Insulin Action, in High-Fat–Fed Mice. Diabetes 2014, 63, 3699–3710. [Google Scholar] [CrossRef]
- Brownlee, M. Biochemistry and Molecular Cell Biology of Diabetic Complications. Nature 2001, 414, 813–820. [Google Scholar] [CrossRef]
- Di Emidio, G.; Santini, S.J.; D’Alessandro, A.M.; Vetuschi, A.; Sferra, R.; Artini, P.G.; Carta, G.; Falone, S.; Amicarelli, F.; Tatone, C. SIRT1 Participates in the Response to Methylglyoxal-Dependent Glycative Stress in Mouse Oocytes and Ovary. Biochim. Biophys. Acta Mol. Basis Dis. 2019, 1865, 1389–1401. [Google Scholar] [CrossRef] [PubMed]
- Yang, Q.; Liu, R.; Yu, Q.; Bi, Y.; Liu, G. Metabolic Regulation of Inflammasomes in Inflammation. Immunology 2019, 157, 95–109. [Google Scholar] [CrossRef] [PubMed]
- Han, Y.; Xu, X.; Tang, C.; Gao, P.; Chen, X.; Xiong, X.; Yang, M.; Yang, S.; Zhu, X.; Yuan, S.; et al. Reactive Oxygen Species Promote Tubular Injury in Diabetic Nephropathy: The Role of the Mitochondrial Ros-Txnip-Nlrp3 Biological Axis. Redox Biol. 2018, 16, 32–46. [Google Scholar] [CrossRef]
- Pan, M.; Zhang, F.; Qu, K.; Liu, C.; Zhang, J. TXNIP: A Double-Edged Sword in Disease and Therapeutic Outlook. Oxidative Med. Cell. Longev. 2022, 2022, 7805115. [Google Scholar] [CrossRef]
- Xu, J.; Cai, S.; Zhao, J.; Xu, K.; Ji, H.; Wu, C.; Xiao, J.; Wu, Y. Advances in the Relationship between Pyroptosis and Diabetic Neuropathy. Front. Cell Dev. Biol. 2021, 9, 753660. [Google Scholar] [CrossRef]
- Molla, M.D.; Akalu, Y.; Geto, Z.; Dagnew, B.; Ayelign, B.; Shibabaw, T. Role of Caspase-1 in the Pathogenesis of Inflammatory-Associated Chronic Noncommunicable Diseases. J. Inflamm. Res. 2020, 13, 749–764. [Google Scholar] [CrossRef]
- Dinarello, C.A.; Donath, M.Y.; Mandrup-Poulsen, T. Role of IL-1β in Type 2 Diabetes. Curr. Opin. Endocrinol. Diabetes Obes. 2010, 17, 314–321. [Google Scholar] [CrossRef]
- Wang, J.; Shen, X.; Liu, J.; Chen, W.; Wu, F.; Wu, W.; Meng, Z.; Zhu, M.; Miao, C. High Glucose Mediates NLRP3 Inflammasome Activation via Upregulation of ELF3 Expression. Cell Death Dis. 2020, 11, 383. [Google Scholar] [CrossRef] [PubMed]
- Forbes, J.M.; Cooper, M.E. Mechanisms of Diabetic Complications. Physiol. Rev. 2013, 93, 137–188. [Google Scholar] [CrossRef] [PubMed]
- McLellan, A.C.; Phillips, S.A.; Thornalley, P.J. The Assay of SD-Lactoylglutathione in Biological Systems. Anal. Biochem. 1993, 211, 37–43. [Google Scholar] [CrossRef]
- Eelen, G.; de Zeeuw, P.; Treps, L.; Harjes, U.; Wong, B.W.; Carmeliet, P. Endothelial Cell Metabolism. Physiol. Rev. 2018, 98, 3–58. [Google Scholar] [CrossRef] [PubMed]
- Souza-Smith, F.M.; Katz, P.S.; Trask, A.J.; Stewart, J.A., Jr.; Lord, K.C.; Varner, K.J.; Vassallo, D.V.; Lucchesi, P.A. Mesenteric Resistance Arteries in Type 2 Diabetic Db/Db Mice Undergo Outward Remodeling. PLoS ONE 2011, 6, e23337. [Google Scholar] [CrossRef] [PubMed]
- Dafre, A.L.; Goldberg, J.; Wang, T.; Spiegel, D.A.; Maher, P. Methylglyoxal, the Foe and Friend of Glyoxalase and Trx/TrxR Systems in HT22 Nerve Cells. Free Radic. Biol. Med. 2015, 89, 8–19. [Google Scholar] [CrossRef] [PubMed]
- Thornalley, P.J.; Rabbani, N. Detection of Oxidized and Glycated Proteins in Clinical Samples Using Mass Spectrometry—A User’s Perspective. Biochim. Biophys. Acta Gen. Subj. 2014, 1840, 818–829. [Google Scholar] [CrossRef]
- Allaman, I.; Bélanger, M.; Magistretti, P.J. Methylglyoxal, the Dark Side of Glycolysis. Front. Neurosci. 2015, 9, 23. [Google Scholar] [CrossRef]
- Kalapos, M.P. The Tandem of Free Radicals and Methylglyoxal. Chem. Biol. Interact. 2008, 171, 251. [Google Scholar] [CrossRef]
- Lai, S.W.T.; Lopez Gonzalez, E.D.J.; Zoukari, T.; Ki, P.; Shuck, S.C. Methylglyoxal and Its Adducts: Induction, Repair, and Association with Disease. Chem. Res. Toxicol. 2022, 35, 1720–1746. [Google Scholar] [CrossRef]
- Rabbani, N.; Xue, M.; Thornalley, P.J. Dicarbonyls and Glyoxalase in Disease Mechanisms and Clinical Therapeutics. Glycoconj. J. 2016, 33, 513–525. [Google Scholar] [CrossRef] [PubMed]
- Nigro, C.; Leone, A.; Raciti, G.A.; Longo, M.; Mirra, P.; Formisano, P.; Beguinot, F.; Miele, C. Methylglyoxal-Glyoxalase 1 Balance: The Root of Vascular Damage. Int. J. Mol. Sci. 2017, 18, 188. [Google Scholar] [CrossRef] [PubMed]
- Blum, A.; Pastukh, N.; Socea, D.; Jabaly, H. Levels of Adhesion Molecules in Peripheral Blood Correlat with Stages of Diabetic Retinopathy and May Serve as Bio Markers for Microvascular Complications. Cytokine 2018, 106, 76–79. [Google Scholar] [CrossRef]
- Tousoulis, D.; Papageorgiou, N.; Androulakis, E.; Siasos, G.; Latsios, G.; Tentolouris, K.; Stefanadis, C. Diabetes Mellitus-Associated Vascular Impairment. J. Am. Coll. Cardiol. 2013, 62, 667–676. [Google Scholar] [CrossRef] [PubMed]
- Stratmann, B.; Engelbrecht, B.; Espelage, B.C.; Klusmeier, N.; Tiemann, J.; Gawlowski, T.; Mattern, Y.; Eisenacher, M.; Meyer, H.E.; Rabbani, N.; et al. Glyoxalase 1-Knockdown in Human Aortic Endothelial Cells—Effect on the Proteome and Endothelial Function Estimates. Sci. Rep. 2016, 6, 37737. [Google Scholar] [CrossRef]
- Gustavsson, C.; Agardh, C.-D.; Zetterqvist, A.V.; Nilsson, J.; Agardh, E.; Gomez, M.F. Vascular Cellular Adhesion Molecule-1 (VCAM-1) Expression in Mice Retinal Vessels Is Affected by Both Hyperglycemia and Hyperlipidemia. PLoS ONE 2010, 5, e12699. [Google Scholar] [CrossRef]
- Hegazy, G.A.; Awan, Z.; Hashem, E.; Al-Ama, N.; Abunaji, A.B. Levels of Soluble Cell Adhesion Molecules in Type 2 Diabetes Mellitus Patients with Macrovascular Complications. J. Int. Med. Res. 2020, 48, 0300060519893858. [Google Scholar] [CrossRef]
- Liu, J.-J.; Yeoh, L.Y.; Sum, C.F.; Tavintharan, S.; Ng, X.W.; Liu, S.; Lee, S.B.M.; Tang, W.E.; Lim, S.C. Vascular Cell Adhesion Molecule-1, but Not Intercellular Adhesion Molecule-1, Is Associated with Diabetic Kidney Disease in Asians with Type 2 Diabetes. J. Diabetes Its Complicat. 2015, 29, 707–712. [Google Scholar] [CrossRef]
- Brouwers, O.; Niessen, P.M.G.; Miyata, T.; Østergaard, J.A.; Flyvbjerg, A.; Peutz-Kootstra, C.J.; Sieber, J.; Mundel, P.H.; Brownlee, M.; Janssen, B.J.A.; et al. Glyoxalase-1 Overexpression Reduces Endothelial Dysfunction and Attenuates Early Renal Impairment in a Rat Model of Diabetes. Diabetologia 2014, 57, 224–235. [Google Scholar] [CrossRef]
- Domingues, A.; Jolibois, J.; Marquet de Rougé, P.; Nivet-Antoine, V. The Emerging Role of TXNIP in Ischemic and Cardiovascular Diseases; A Novel Marker and Therapeutic Target. Int. J. Mol. Sci. 2021, 22, 1693. [Google Scholar] [CrossRef]
- Zhou, R.; Tardivel, A.; Thorens, B.; Choi, I.; Tschopp, J. Thioredoxin-Interacting Protein Links Oxidative Stress to Inflammasome Activation. Nat. Immunol. 2010, 11, 136–140. [Google Scholar] [CrossRef] [PubMed]
- Li, X.; Kover, K.L.; Heruth, D.P.; Watkins, D.J.; Guo, Y.; Moore, W.V.; He, L.G.; Zang, M.; Clements, M.A.; Yan, Y. Thioredoxin-Interacting Protein Promotes High-Glucose-Induced Macrovascular Endothelial Dysfunction. Biochem. Biophys. Res. Commun. 2017, 493, 291–297. [Google Scholar] [CrossRef] [PubMed]


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Gene | Organism | Forward Primer (5′ to 3′) | Reverse Primer (5′ to 3′) |
|---|---|---|---|
| Glo-1 | Human | GGTTTGAAGAACTGGGAGTCAAA | ATCCAGTAGCCATCAGGATCTTG |
| Nrf-2 | Human | TCATGATGGACTTGGAGCTG | CATACTCTTTCCGTCGCTGA |
| IL-1β | Human | AATCTGTACCTGTCCTGCGTGTT | TGGGTAATTTTTGGGATCTACACTCT |
| SOD-2 | Human | CAGGACCCACTGCAAGGAA | CGTGCTCCCACACATCAATC |
| SIRT-1 | Human | CCGCGGATAGGTCCATATACT | AACAATCTGCCACAGCGTCA |
| VCAM-1 | Human | ACATGGAATTCGAACCCAAA | TGTATCTCTGGGGGCAACAT |
| ICAM-1 | Human | CCTTCCTCACCGTGTACTGG | AGCGTAGGGTAAGGTTCTTGC |
| NLRP-3 | Human | GATCTTCGCTGCGATCAACA | GGGATTCGAAACACGTGCATTA |
| Caspase-1 | Human | GCCTGTTCCTGTGATGTGGAG | TGCCCACAGACATTCATACAGTTTC |
| TXNIP | Human | TGGTGGATGTCAATACCCCT | ATTGGCAAGGTAAGTGTGGC |
| β2-Microglobulin | Human | GGCTATCCAGCGTACTCCAAA | CGGCAGGCATACTCATCTTTTT |
| Cyclophilin | Human | TTCCTGCTTTCACAGAATTATTCC | GCCACCAGTGCCATTATGG |
| Glo-1 | Rat | GAAGATGACGAGACGCAGAGTTAC | CAGGATCTTGAACGAACGCCAGAC |
| Nrf-2 | Rat | ACTCCCAGGTTGCCCACAT | CATGGTCATCTACAAATGGGAATG |
| IL-1β | Rat | CTGCAGCTGGAGAGTGTGGAT | CACTTTGCTCTTGACTTCTATCTTGTTG |
| SOD-2 | Rat | CAGGACCCACTGCAAGGAA | CGTGCTCCCACACATCAATC |
| SIRT-1 | Rat | CCGCGGATAGGTCCATATACT | AACAATCTGCCACAGCGTCA |
| VCAM-1 | Rat | AGTGTGAATCGAAAACCGAAGTC | AATGGCGGGTATTACCAAGGA |
| ICAM-1 | Rat | GCTGCGCTGTGTTTTGGA | GGATGGGAGCTGAAAAGTTGTAGA |
| NLRP-3 | Rat | AAGCTGTCCTCAGTCAGAGGCAAT | ATGACTTTCTTGGAGCCAGGGACA |
| Caspase-1 | Rat | AGGAGGGAATATGTGGG | AACCTTGGGCTTGTCTT |
| TXNIP | Rat | CAGACCTCCAAGACCACGACTG | CATCCGCAGCCAATGAACAGAG |
| β2-Microglobulin | Rat | TGGCCGTCGTGCTTGCCATT | TCTCCGGTGGATGGCGAGAGT |
| Cyclophilin | Rat | TTCCTCCTTTCACAGAATTATTCCA | CCATTATGGCGTGTGAAGTCA |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Syed, N.A.; Bhatti, A.; John, P. Molecular Link between Glo-1 Expression and Markers of Hyperglycemia and Oxidative Stress in Vascular Complications of Type 2 Diabetes Mellitus. Antioxidants 2023, 12, 1663. https://doi.org/10.3390/antiox12091663
Syed NA, Bhatti A, John P. Molecular Link between Glo-1 Expression and Markers of Hyperglycemia and Oxidative Stress in Vascular Complications of Type 2 Diabetes Mellitus. Antioxidants. 2023; 12(9):1663. https://doi.org/10.3390/antiox12091663
Chicago/Turabian StyleSyed, Nida Ali, Attya Bhatti, and Peter John. 2023. "Molecular Link between Glo-1 Expression and Markers of Hyperglycemia and Oxidative Stress in Vascular Complications of Type 2 Diabetes Mellitus" Antioxidants 12, no. 9: 1663. https://doi.org/10.3390/antiox12091663
APA StyleSyed, N. A., Bhatti, A., & John, P. (2023). Molecular Link between Glo-1 Expression and Markers of Hyperglycemia and Oxidative Stress in Vascular Complications of Type 2 Diabetes Mellitus. Antioxidants, 12(9), 1663. https://doi.org/10.3390/antiox12091663