

Integrated Multi-Omics Analysis for Inferring Molecular Players in Inclusion Body Myositis

 , , , ,
, , , ,
Abstract
:
1. Introduction
2. Materials and Methods
2.1. Study Design
2.2. Sample Collection
2.3. Aβ1–42 Detection
2.4. Metabolite Quantification
2.5. Glutathione Peroxidase (GPX) Activity Assay
2.6. RNA Extraction and mRNA Library Preparation and Sequencing
2.7. RNA-Seq Data Processing and Analysis
2.8. Statistical Analysis
3. Results
3.1. Clinical and Epidemiological Data
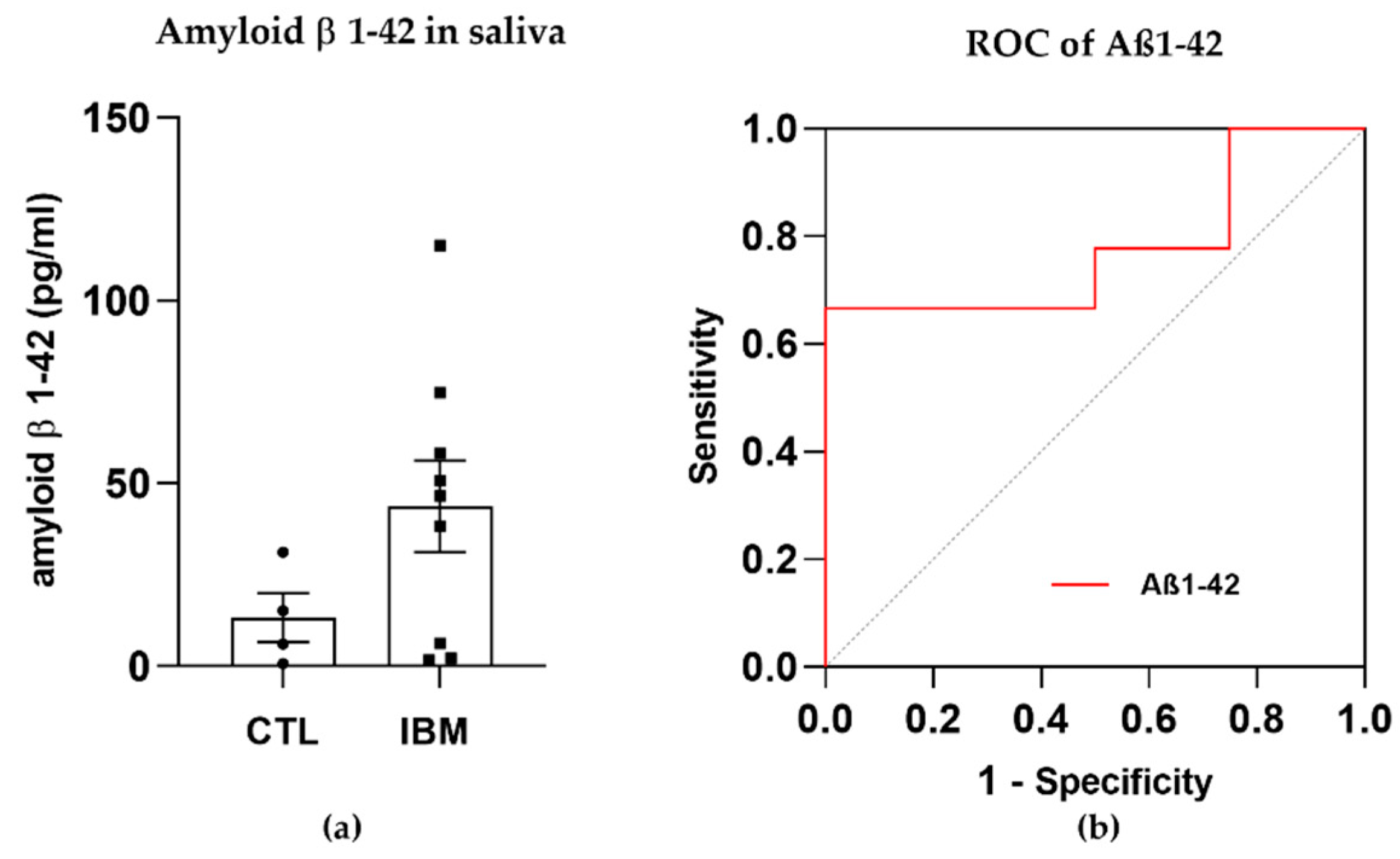
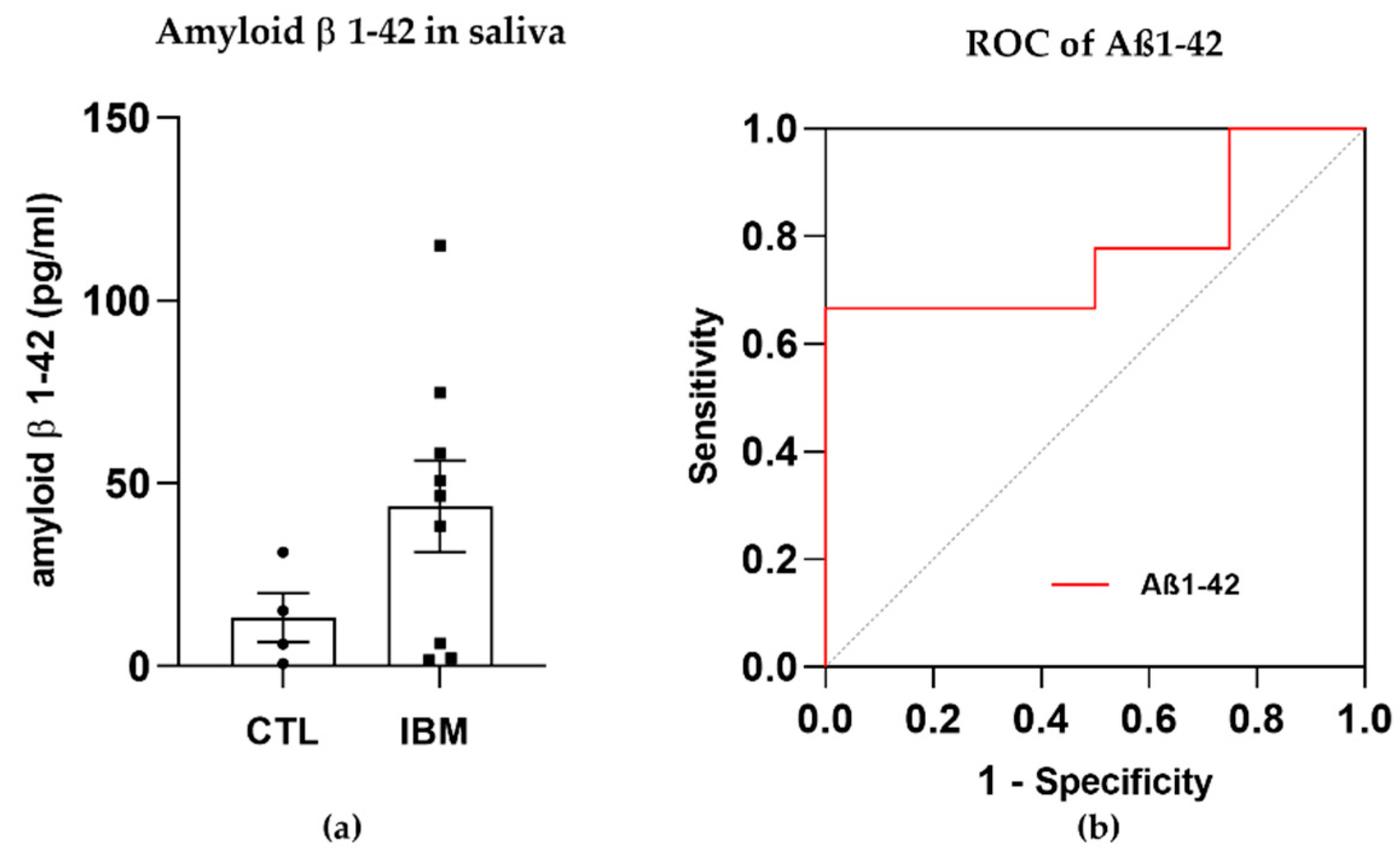
3.2. Degeneration Markers: Aβ1–42 Concentration Was Higher in IBM Samples
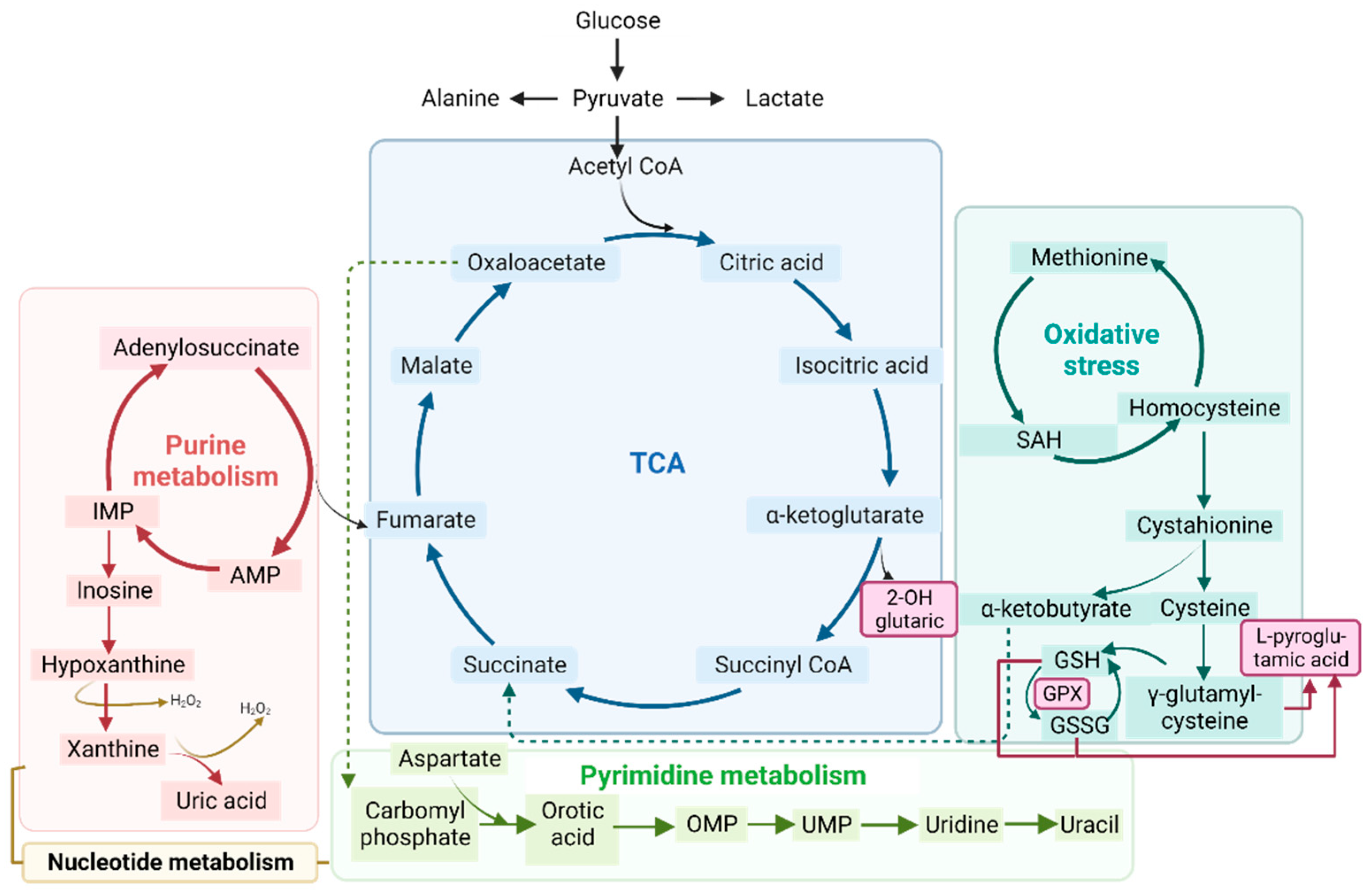
3.3. Mitochondrial Alterations
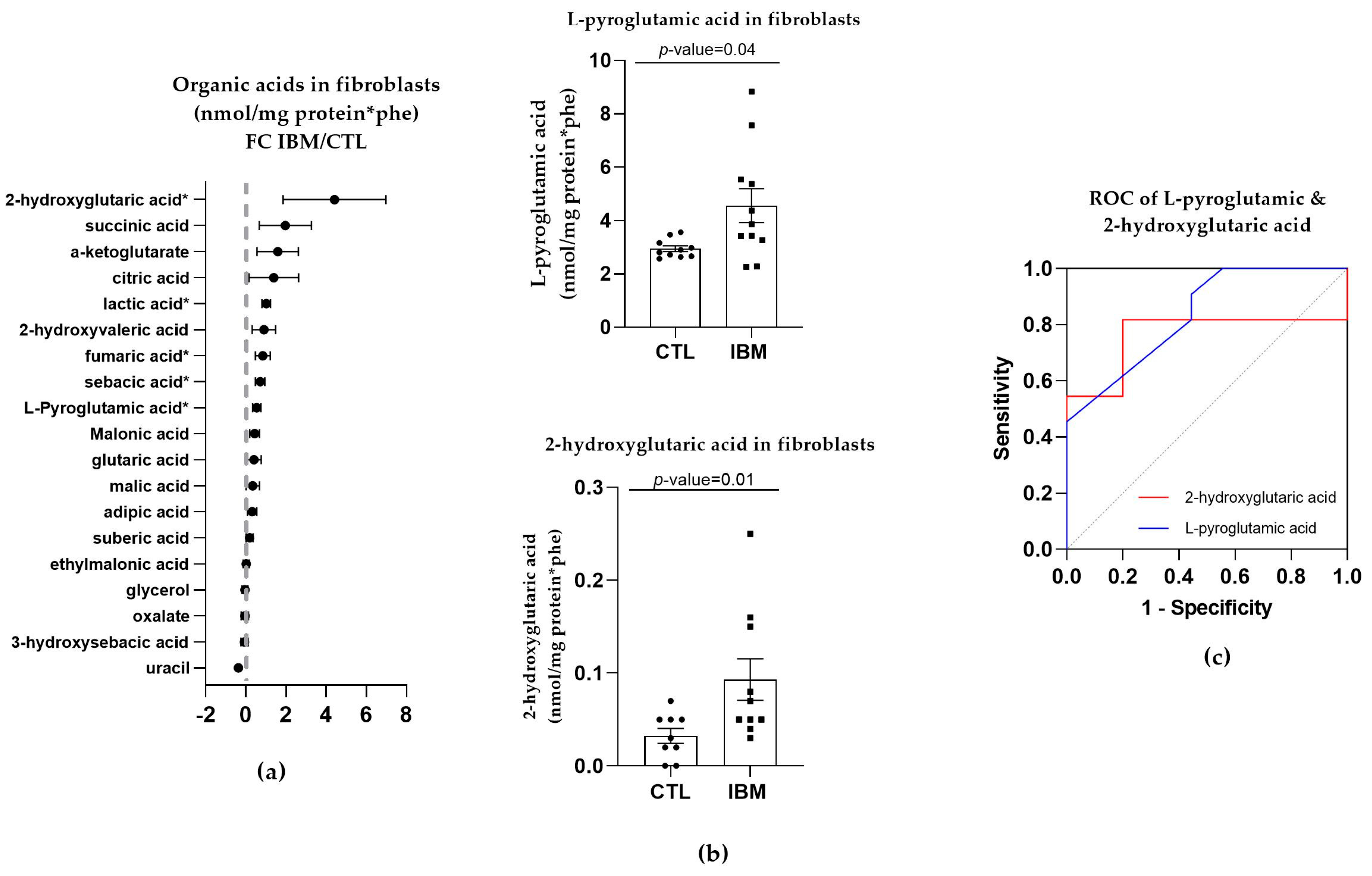
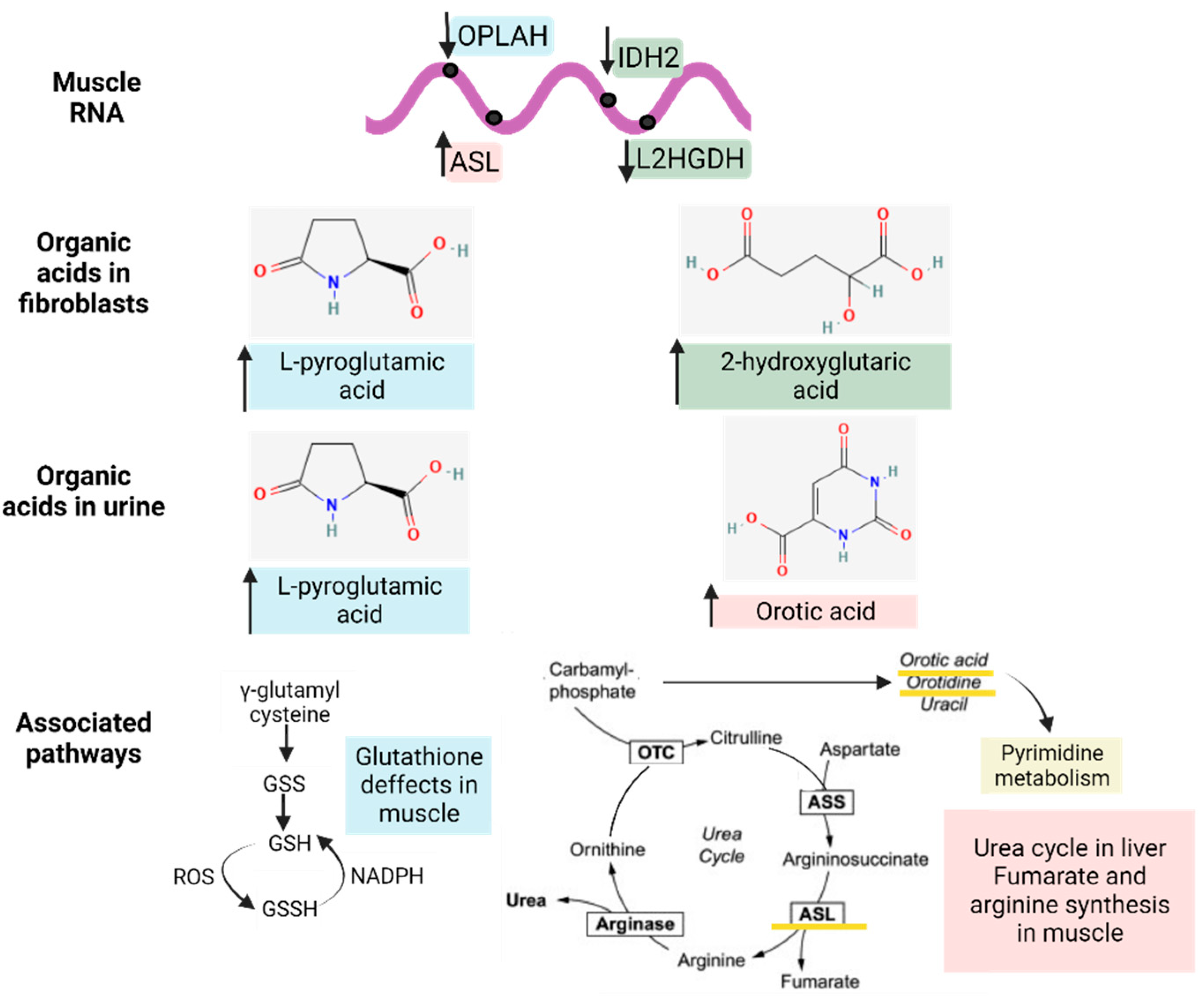
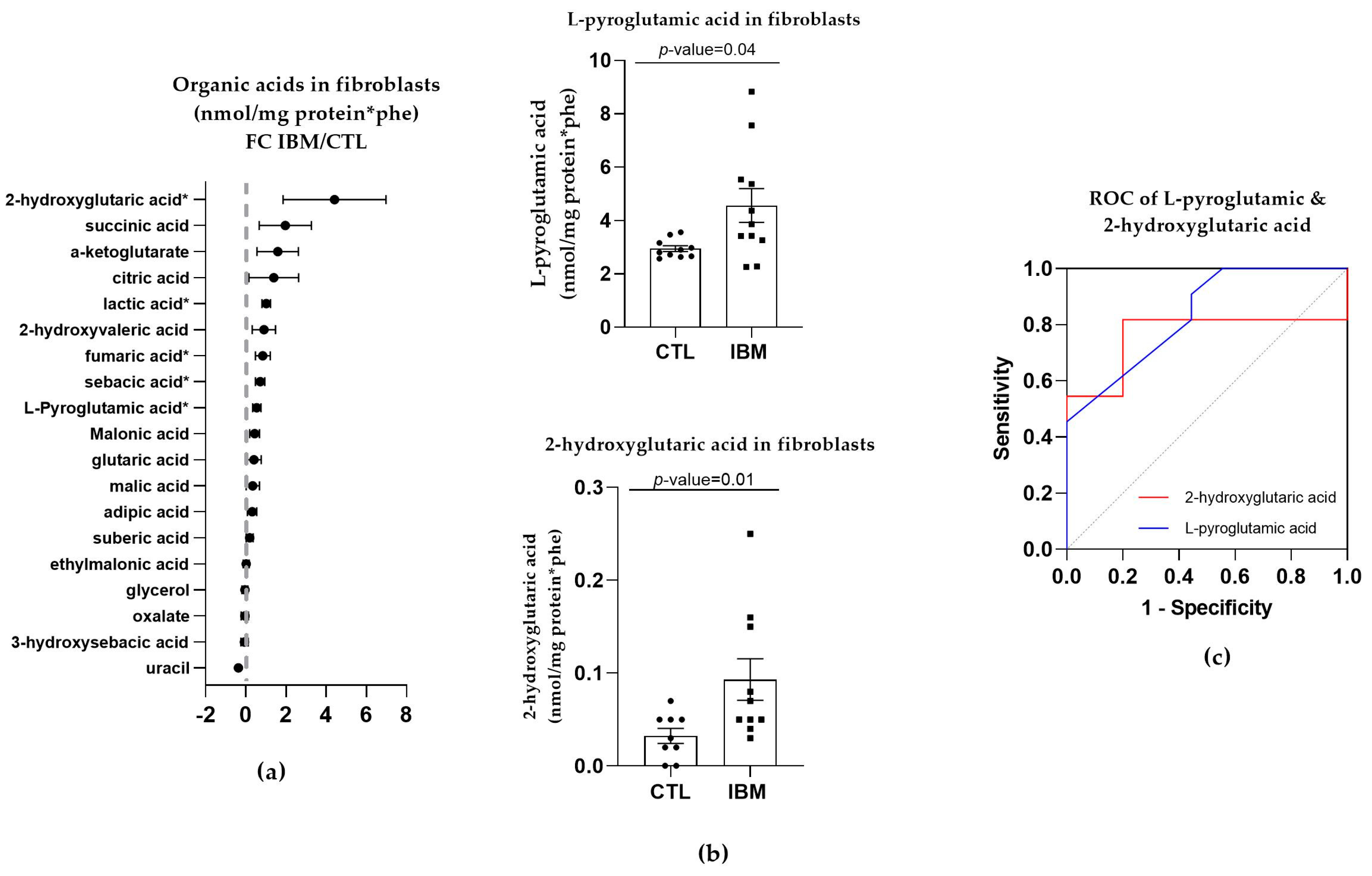
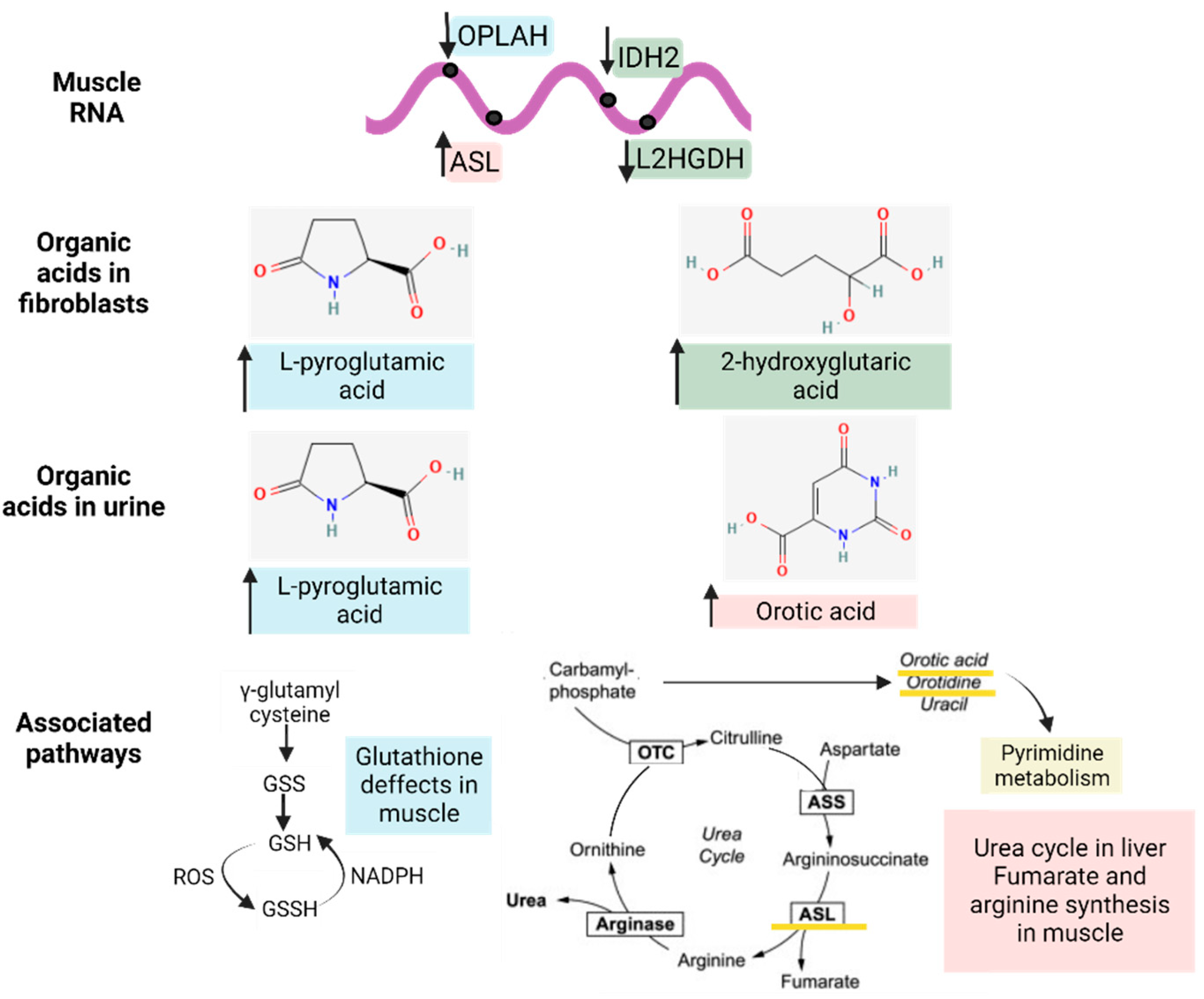
3.3.1. Organic Acid Profiles Increased in IBM Fibroblasts
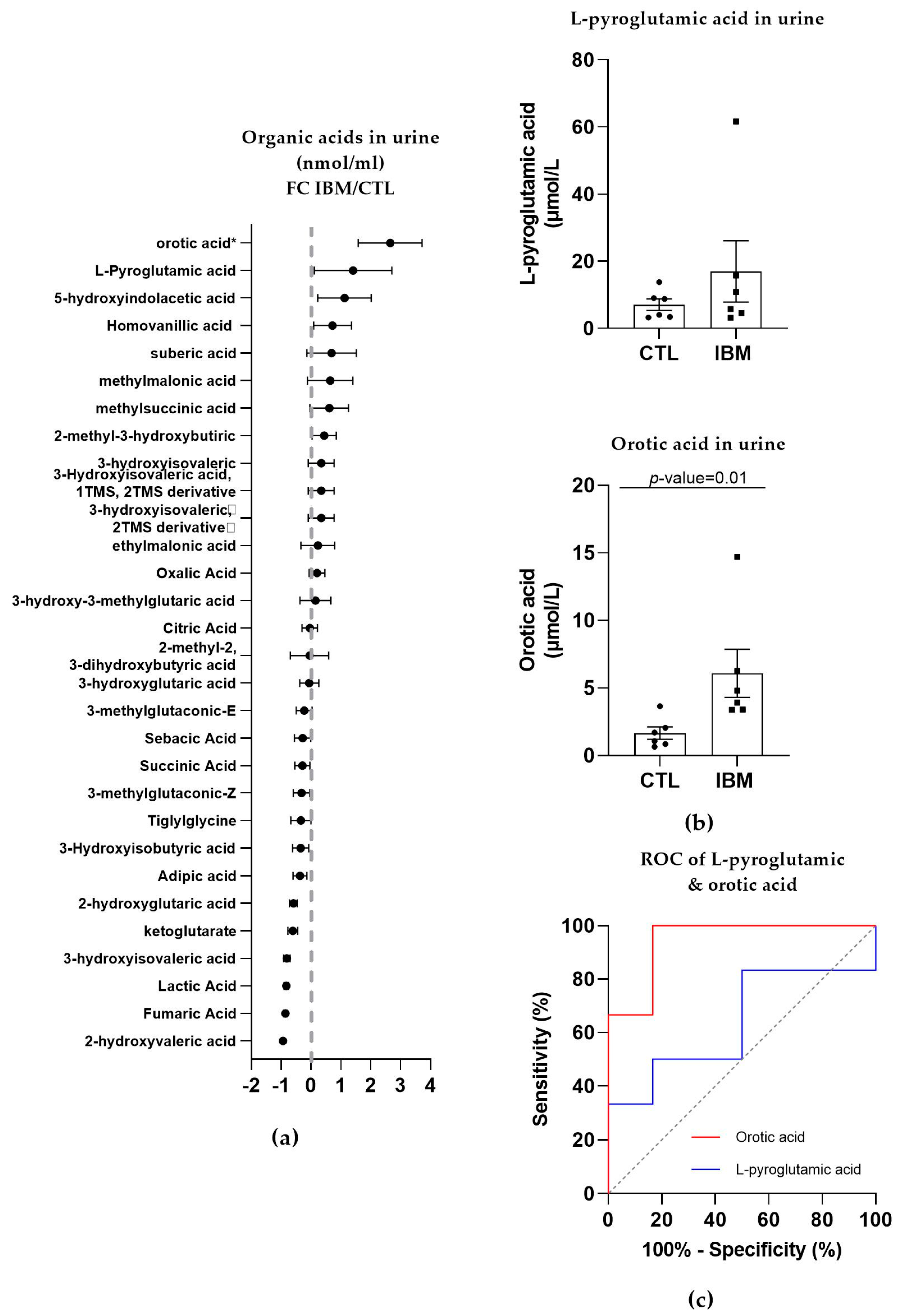
3.3.2. Organic Acid Profiling in Urine Reveals Altered Metabolism in IBM
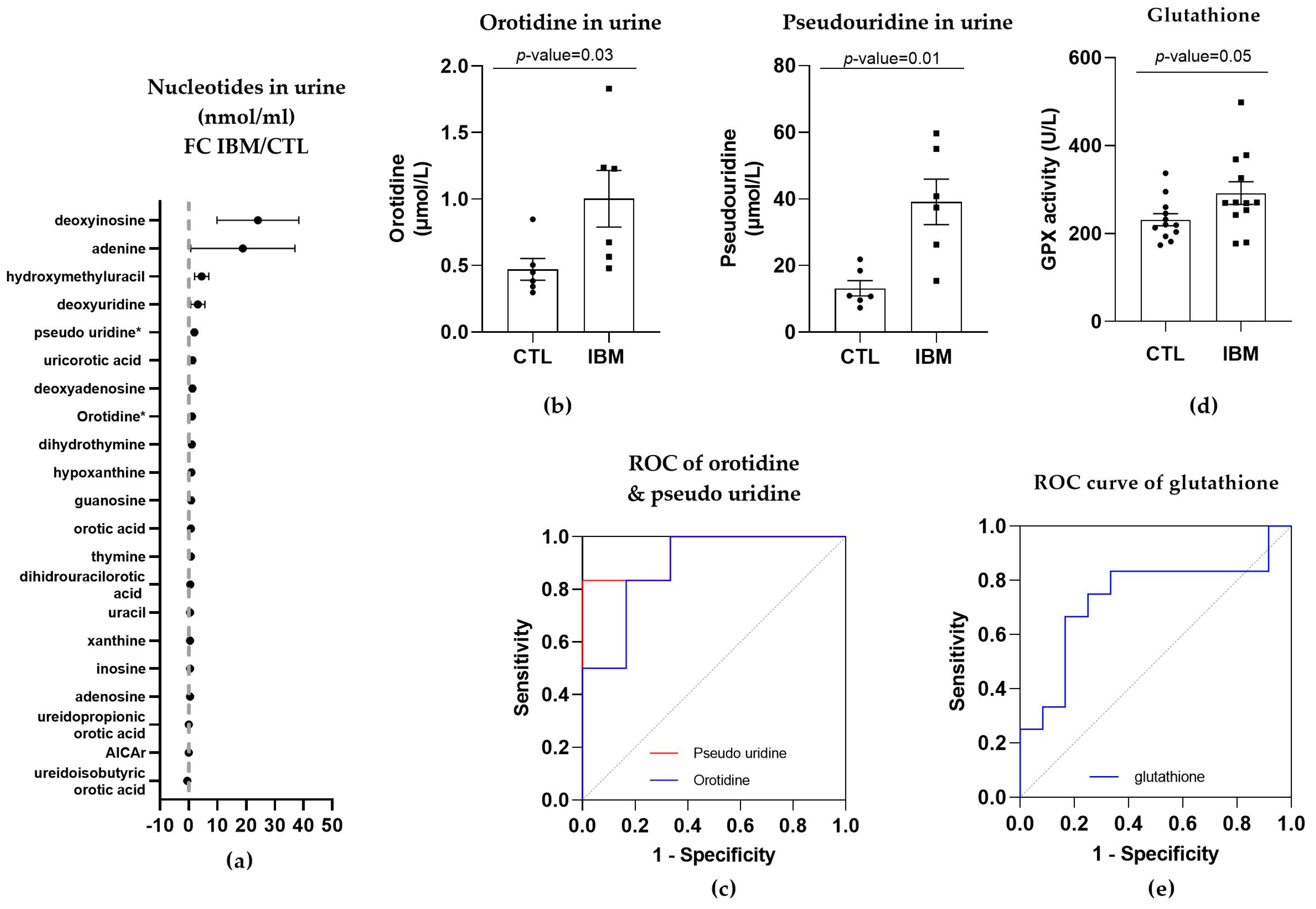
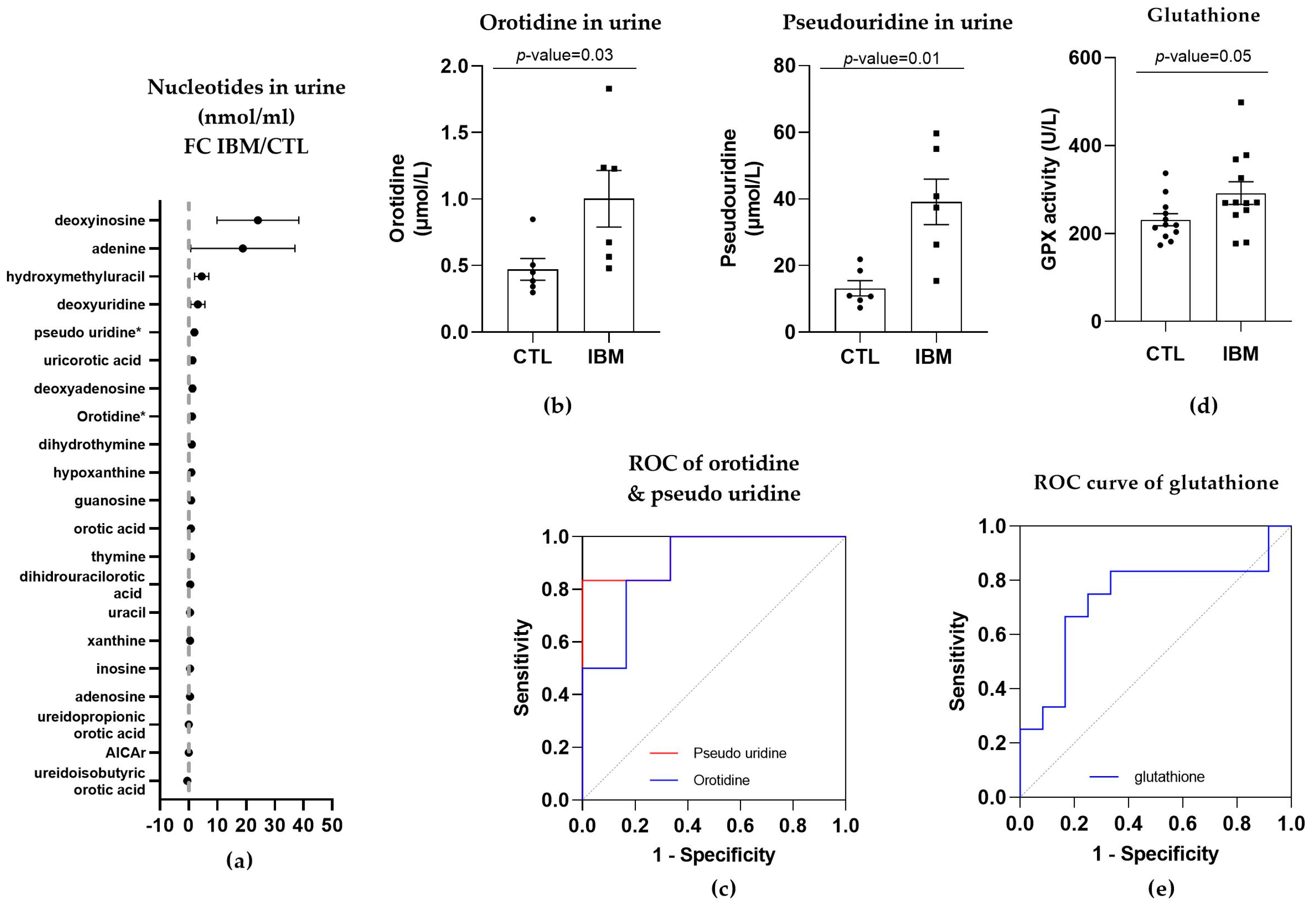
3.3.3. Nucleotide Synthesis Precursor Levels Were Increased in the Urine of IBM Patients
3.3.4. GPX Activity Levels in Plasma
3.3.5. Metabolic-Upstream-Related Genes in the IBM Muscle Transcriptome
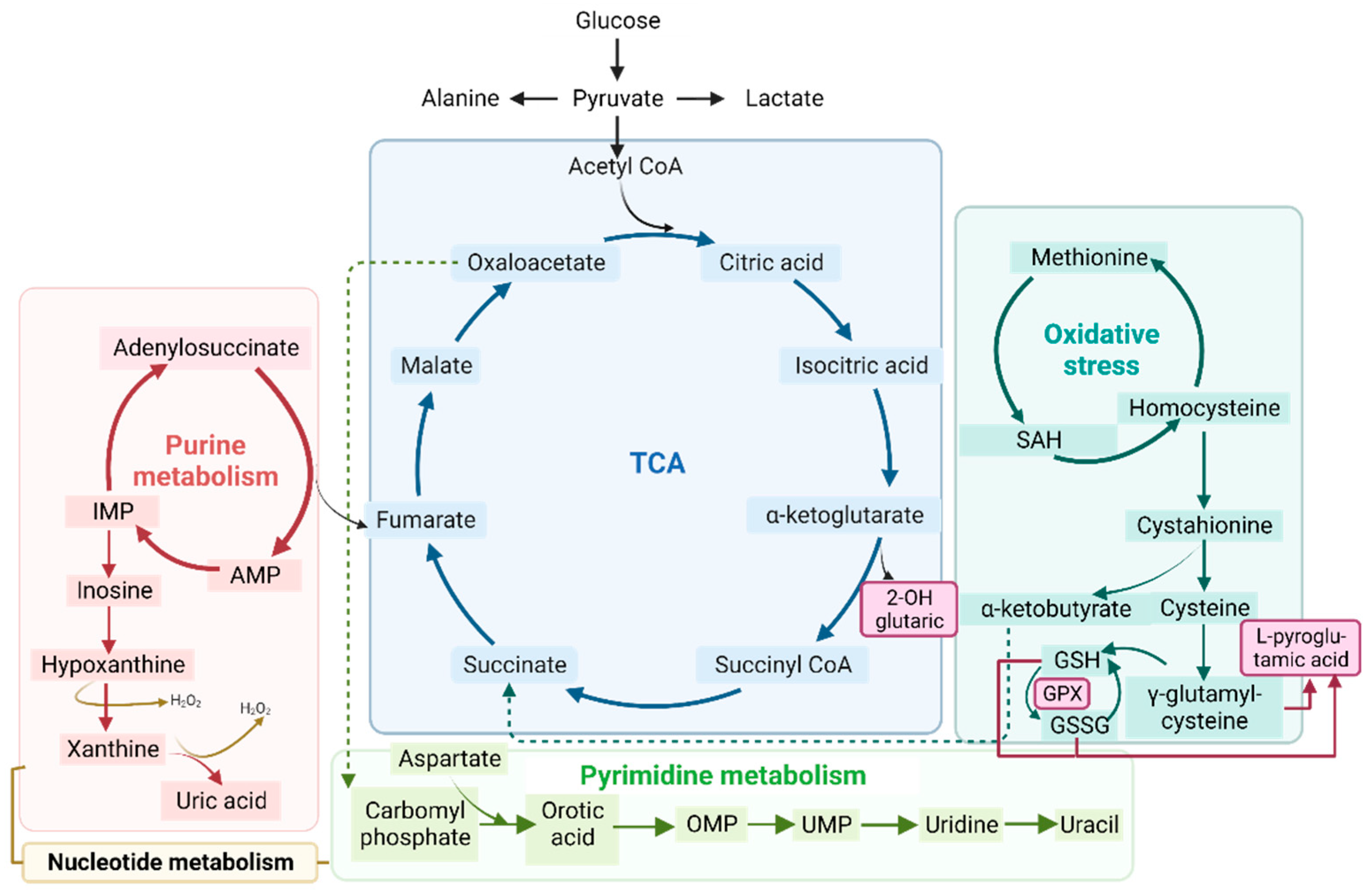
3.4. Gene–Metabolites Interaction: Potential Interactome Framework for IBM Samples
4. Discussion
5. Conclusions
- Metabolic dysregulation in IBM is present outside the target tissue (muscle), as seen in the altered organic acids in fibroblasts and urine;
- The multi-omics profiling of patients’ samples allows for the evaluation of disease-associated phenotypes, constituting an untargeted approach enabling the potential detection of novel molecular players;
- The detection of L-pyroglutamic and orotic acids in urine displayed an outstanding biomarker signature, with 100% sensitivity and specificity;
- The validation of potential biomarkers in non-invasive samples like urine may eventually aid in the screening of patients’ disease progression and treatment efficacy.
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Nagy, S.; Khan, A.; Machado, P.M.; Houlden, H. Inclusion Body Myositis: From Genetics to Clinical Trials. J. Neurol. 2023, 270, 1787–1797. [Google Scholar] [CrossRef] [PubMed]
- Callan, A.; Capkun, G.; Vasanthaprasad, V.; Freitas, R.; Needham, M. A Systematic Review and Meta-Analysis of Prevalence Studies of Sporadic Inclusion Body Myositis. J. Neuromuscul. Dis. 2017, 4, 127–137. [Google Scholar] [CrossRef] [PubMed]
- Greenberg, S.A. Inclusion Body Myositis: Clinical Features and Pathogenesis. Nat. Rev. Rheumatol. 2019, 15, 257–272. [Google Scholar] [CrossRef]
- Naddaf, E.; Shelly, S.; Mandrekar, J.; Chamberlain, A.M.; Hoffman, E.M.; Ernste, F.C.; Liewluck, T. Survival and Associated Comorbidities in Inclusion Body Myositis. Rheumatology 2022, 61, 2016–2024. [Google Scholar] [CrossRef]
- Buzkova, J.; Nikkanen, J.; Ahola, S.; Hakonen, A.H.; Sevastianova, K.; Hovinen, T.; Yki-Järvinen, H.; Pietiläinen, K.H.; Lönnqvist, T.; Velagapudi, V.; et al. Metabolomes of Mitochondrial Diseases and Inclusion Body Myositis Patients: Treatment Targets and Biomarkers. EMBO Mol. Med. 2018, 10, 9091. [Google Scholar] [CrossRef]
- Hedberg-Oldfors, C.; Lindgren, U.; Basu, S.; Visuttijai, K.; Lindberg, C.; Falkenberg, M.; Larsson Lekholm, E.; Oldfors, A. Mitochondrial DNA Variants in Inclusion Body Myositis Characterized by Deep Sequencing. Brain Pathol. 2021, 31, e12931. [Google Scholar] [CrossRef] [PubMed]
- Catalan-Garcia, M.; Garrabou, G.; Moren, C.; Guitart-Mampel, M.; Hernando, A.; Diaz-Ramos, A.; Gonzalez-Casacuberta, I.; Juarez, D.-L.; Bano, M.; Enrich-Bengoa, J.; et al. Mitochondrial DNA Disturbances and Deregulated Expression of Oxidative Phosphorylation and Mitochondrial Fusion Proteins in Sporadic Inclusion Body Myositis. Clin. Sci. 2016, 130, 1741–1751. [Google Scholar] [CrossRef] [PubMed]
- Greenberg, S.A.; Pinkus, J.L.; Kong, S.W.; Baecher-Allan, C.; Amato, A.A.; Dorfman, D.M. Highly Differentiated Cytotoxic T Cells in Inclusion Body Myositis. Brain 2019, 142, 2590–2604. [Google Scholar] [CrossRef]
- Greenberg, S.A. Pathogenesis of Inclusion Body Myositis. Curr. Opin. Rheumatol. 2020, 32, 542–547. [Google Scholar] [CrossRef]
- Singh, R.J. Glutathione: A Marker and Antioxidant for Aging. J. Lab. Clin. Med. 2002, 140, 1. [Google Scholar] [CrossRef] [PubMed]
- Wu, J.H.; Batist, G. Glutathione and Glutathione Analogues; Therapeutic Potentials. Biochim. Biophys. Acta (BBA)—Gen. Subj. 2013, 1830, 3350–3353. [Google Scholar] [CrossRef] [PubMed]
- Forman, H.J.; Zhang, H.; Rinna, A. Glutathione: Overview of Its Protective Roles, Measurement, and Biosynthesis. Mol. Asp. Med. 2009, 30, 1–12. [Google Scholar] [CrossRef] [PubMed]
- Cantó-Santos, J.; Valls-Roca, L.; Tobías, E.; García-García, F.J.; Guitart-Mampel, M.; Esteve-Codina, A.; Martín-Mur, B.; Casado, M.; Artuch, R.; Solsona-Vilarrasa, E.; et al. Unravelling Inclusion Body Myositis Using a Patient-derived Fibroblast Model. J. Cachexia Sarcopenia Muscle 2023, 14, 964–977. [Google Scholar] [CrossRef] [PubMed]
- Amici, D.R.; Pinal-Fernandez, I.; Christopher-Stine, L.; Mammen, A.L.; Mendillo, M.L. A Network of Core and Subtype-Specific Gene Expression Programs in Myositis. Acta Neuropathol. 2021, 142, 887–898. [Google Scholar] [CrossRef] [PubMed]
- Britson, K.A.; Ling, J.P.; Braunstein, K.E.; Montagne, J.M.; Kastenschmidt, J.M.; Wilson, A.; Ikenaga, C.; Tsao, W.; Pinal-Fernandez, I.; Russell, K.A.; et al. Loss of TDP-43 Function and Rimmed Vacuoles Persist after T Cell Depletion in a Xenograft Model of Sporadic Inclusion Body Myositis. Sci. Transl. Med. 2022, 14, 9196. [Google Scholar] [CrossRef]
- De Paepe, B. Sporadic Inclusion Body Myositis: An Acquired Mitochondrial Disease with Extras. Biomolecules 2019, 9, 15. [Google Scholar] [CrossRef]
- Hasin, Y.; Seldin, M.; Lusis, A. Multi-Omics Approaches to Disease. Genome Biol. 2017, 18, 83. [Google Scholar] [CrossRef]
- MR, R. 188th ENMC International Workshop: Inclusion Body Myositis, 2–4 December 2011, Naarden, The Netherlands. Neuromuscul. Disord. 2013, 23, 1044–1055. [Google Scholar] [CrossRef]
- Bermejo-Pareja, F.; Antequera, D.; Vargas, T.; Molina, J.A.; Carro, E. Saliva Levels of Abeta1–42 as Potential Biomarker of Alzheimer’s Disease: A Pilot Study. BMC Neurol. 2010, 10, 108. [Google Scholar] [CrossRef]
- Lee, M.; Guo, J.P.; Kennedy, K.; Mcgeer, E.G.; McGeer, P.L. A Method for Diagnosing Alzheimer’s Disease Based on Salivary Amyloid-β Protein 42 Levels. J. Alzheimer’s Disease 2017, 55, 1175–1182. [Google Scholar] [CrossRef]
- Sabbagh, M.N.; Shi, J.; Lee, M.; Arnold, L.; Al-Hasan, Y.; Heim, J.; McGeer, P. Salivary Beta Amyloid Protein Levels Are Detectable and Differentiate Patients with Alzheimer’s Disease Dementia from Normal Controls: Preliminary Findings. BMC Neurol. 2018, 18, 155. [Google Scholar] [CrossRef] [PubMed]
- Catalán, M.; Selva-O’Callaghan, A.; Grau, J.M. Diagnosis and Classification of Sporadic Inclusion Body Myositis (SIBM). Autoimmun. Rev. 2014, 13, 363–366. [Google Scholar] [CrossRef]
- Selva-O’callaghan, A.; Pinal-Fernandez, I.; Trallero-Araguás, E.; Milisenda, J.C.; Grau-Junyent, M.; Mammen, A.L. Classification and Management of Adult Inflammatory Myopathies. Lancet Neurol. 2018, 17, 816–828. [Google Scholar] [CrossRef] [PubMed]
- Hoffmann, G.F.; Feyh, P. Organic Acid Analysis. In Physician’s Guide to the Laboratory Diagnosis of Metabolic Diseases; Springer: Berlin/Heidelberg, Germany, 2003; pp. 27–44. [Google Scholar]
- Dobin, A.; Davis, C.A.; Schlesinger, F.; Drenkow, J.; Zaleski, C.; Jha, S.; Batut, P.; Chaisson, M.; Gingeras, T.R. STAR: Ultrafast Universal RNA-Seq Aligner. Bioinformatics 2013, 29, 15–21. [Google Scholar] [CrossRef]
- Li, B.; Dewey, C.N. RSEM: Accurate Transcript Quantification from RNA-Seq Data with or without a Reference Genome. BMC Bioinform. 2011, 12, 323. [Google Scholar] [CrossRef] [PubMed]
- Love, M.I.; Huber, W.; Anders, S. Moderated Estimation of Fold Change and Dispersion for RNA-Seq Data with DESeq2. Genome Biol. 2014, 15, 550. [Google Scholar] [CrossRef] [PubMed]
- Rath, S.; Sharma, R.; Gupta, R.; Ast, T.; Chan, C.; Durham, T.J.; Goodman, R.P.; Grabarek, Z.; Haas, M.E.; Hung, W.H.W.; et al. MitoCarta3.0: An Updated Mitochondrial Proteome Now with Sub-Organelle Localization and Pathway Annotations. Nucleic Acids Res. 2021, 49, D1541–D1547. [Google Scholar] [CrossRef]
- Metaboanalyst. Available online: https://www.metaboanalyst.ca/ (accessed on 10 March 2023).
- Snedden, A.M.; Kellett, K.A.B.; Lilleker, J.B.; Hooper, N.M.; Chinoy, H. The Role of Protein Aggregation in the Pathogenesis of Inclusion Body Myositis. Clin. Exp. Rheumatol. 2022, 40, 414–424. [Google Scholar] [CrossRef]
- Catalán-García, M.; Garrabou, G.; Morén, C.; Guitart-Mampel, M.; Gonzalez-Casacuberta, I.; Hernando, A.; Gallego-Escuredo, J.M.; Yubero, D.; Villarroya, F.; Montero, R.; et al. BACE-1, PS-1 and SAPPβ Levels Are Increased in Plasma from Sporadic Inclusion Body Myositis Patients: Surrogate Biomarkers among Inflammatory Myopathies. Mol. Med. 2016, 21, 817–823. [Google Scholar] [CrossRef]
- Lane, A.N.; Fan, T.W.M. Regulation of Mammalian Nucleotide Metabolism and Biosynthesis. Nucleic Acids Res. 2015, 43, 2466–2485. [Google Scholar] [CrossRef]
- Cantó-Santos, J.; Grau-Junyent, J.M.; Garrabou, G. The Impact of Mitochondrial Deficiencies in Neuromuscular Diseases. Antioxidants 2020, 9, 964. [Google Scholar] [CrossRef] [PubMed]
- Orphanet. Available online: https://www.orpha.net/ (accessed on 10 March 2023).
- Calpena, E.; Casado, M.; Martínez-Rubio, D.; Nascimento, A.; Colomer, J.; Gargallo, E.; García-Cazorla, A.; Palau, F.; Artuch, R.; Espinós, C. 5-Oxoprolinuria in Heterozygous Patients for 5-Oxoprolinase (OPLAH) Missense Changes. JIMD Rep. 2012, 7, 123–128. [Google Scholar] [CrossRef] [PubMed]
- Benveniste, O.; Stenzel, W.; Hilton-Jones, D.; Sandri, M.; Boyer, O.; van Engelen, B.G.M. Amyloid Deposits and Inflammatory Infiltrates in Sporadic Inclusion Body Myositis: The Inflammatory Egg Comes before the Degenerative Chicken. Acta Neuropathol. 2015, 3, 611–624. [Google Scholar] [CrossRef]
- Goyal, N.A.; Coulis, G.; Duarte, J.; Farahat, P.K.; Mannaa, A.H.; Cauchii, J.; Irani, T.; Araujo, N.; Wang, L.; Wencel, M.; et al. Immunophenotyping of Inclusion Body Myositis Blood T and NK Cells. Neurology 2022, 98, e1374–e1383. [Google Scholar] [CrossRef] [PubMed]
- Perez-Rosendahl, M.; Mozaffar, T. Inclusion Body Myositis: Evolving Concepts. Curr. Opin. Neurol. 2022, 35, 604–610. [Google Scholar] [CrossRef] [PubMed]
- Tawara, N.; Yamashita, S.; Zhang, X.; Korogi, M.; Zhang, Z.; Doki, T.; Matsuo, Y.; Nakane, S.; Maeda, Y.; Sugie, K.; et al. Pathomechanisms of Anti–Cytosolic 5′-Nucleotidase 1A Autoantibodies in Sporadic Inclusion Body Myositis. Ann. Neurol. 2017, 81, 512–525. [Google Scholar] [CrossRef]
- Ikenaga, C.; Date, H.; Kanagawa, M.; Mitsui, J.; Ishiura, H.; Yoshimura, J.; Pinal-Fernandez, I.; Mammen, A.L.; Lloyd, T.E.; Tsuji, S.; et al. Muscle Transcriptomics Shows Overexpression of Cadherin 1 in Inclusion Body Myositis. Ann. Neurol. 2022, 91, 317–328. [Google Scholar] [CrossRef]
- Pinal-Fernandez, I.; Casal-Dominguez, M.; Derfoul, A.; Pak, K.; Plotz, P.; Miller, F.W.; Milisenda, J.C.; Grau-Junyent, J.M.; Selva-O’Callaghan, A.; Paik, J.; et al. Identification of Distinctive Interferon Gene Signatures in Different Types of Myositis. Neurology 2019, 93, e1193–e1204. [Google Scholar] [CrossRef]
- Liss, D.B.; Paden, M.S.; Schwarz, E.S.; Mullins, M.E. What Is the Clinical Significance of 5-Oxoproline (Pyroglutamic Acid) in High Anion Gap Metabolic Acidosis Following Paracetamol (Acetaminophen) Exposure? Clin. Toxicol. 2013, 51, 817–827. [Google Scholar] [CrossRef]
- L-Pyroglutamic Acid. Available online: https://pubchem.ncbi.nlm.nih.gov/compound/L-Pyroglutamic-acid (accessed on 24 June 2023).
- Orotic Acid. Available online: https://pubchem.ncbi.nlm.nih.gov/ (accessed on 12 May 2023).


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Sample Type | Summary | Num. of Samples (n) | Male/Female Ratio | Mean Age (Years) |
|---|---|---|---|---|
| Fibroblasts | IBM | 14 | 0.6 | 66.9 ± 3.6 |
| CTL | 12 | 0.4 | 57.4 ± 4.6 | |
| Urine | IBM | 6 | 0.5 | 70.8 ± 3.7 |
| CTL | 6 | 0.5 | 71.2 ± 3.4 | |
| Muscle | IBM | 5 | 0.40 | 62.6 ± 6.4 |
| CTL | 6 | 0.67 | 44.6 ± 4.7 | |
| Saliva | IBM | 9 | 0.44 | 67.2 ± 4.7 |
| CTL | 5 | 0.6 | 57.0 ± 5.9 | |
| Plasma | IBM | 12 | 0.6 | 63.7 ± 3.5 |
| CTL | 12 | 0.6 | 75.3 ± 0.9 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Cantó-Santos, J.; Valls-Roca, L.; Tobías, E.; Oliva, C.; García-García, F.J.; Guitart-Mampel, M.; Andújar-Sánchez, F.; Esteve-Codina, A.; Martín-Mur, B.; Padrosa, J.; et al. Integrated Multi-Omics Analysis for Inferring Molecular Players in Inclusion Body Myositis. Antioxidants 2023, 12, 1639. https://doi.org/10.3390/antiox12081639
Cantó-Santos J, Valls-Roca L, Tobías E, Oliva C, García-García FJ, Guitart-Mampel M, Andújar-Sánchez F, Esteve-Codina A, Martín-Mur B, Padrosa J, et al. Integrated Multi-Omics Analysis for Inferring Molecular Players in Inclusion Body Myositis. Antioxidants. 2023; 12(8):1639. https://doi.org/10.3390/antiox12081639
Chicago/Turabian StyleCantó-Santos, Judith, Laura Valls-Roca, Ester Tobías, Clara Oliva, Francesc Josep García-García, Mariona Guitart-Mampel, Félix Andújar-Sánchez, Anna Esteve-Codina, Beatriz Martín-Mur, Joan Padrosa, and et al. 2023. "Integrated Multi-Omics Analysis for Inferring Molecular Players in Inclusion Body Myositis" Antioxidants 12, no. 8: 1639. https://doi.org/10.3390/antiox12081639
APA StyleCantó-Santos, J., Valls-Roca, L., Tobías, E., Oliva, C., García-García, F. J., Guitart-Mampel, M., Andújar-Sánchez, F., Esteve-Codina, A., Martín-Mur, B., Padrosa, J., Aránega, R., Moreno-Lozano, P. J., Milisenda, J. C., Artuch, R., Grau-Junyent, J. M., & Garrabou, G. (2023). Integrated Multi-Omics Analysis for Inferring Molecular Players in Inclusion Body Myositis. Antioxidants, 12(8), 1639. https://doi.org/10.3390/antiox12081639