
Unraveling Obesity: Transgenerational Inheritance, Treatment Side Effects, Flavonoids, Mechanisms, Microbiota, Redox Balance, and Bioavailability—A Narrative Review


 ,
,  , , and
, , and
Abstract
1. Introduction
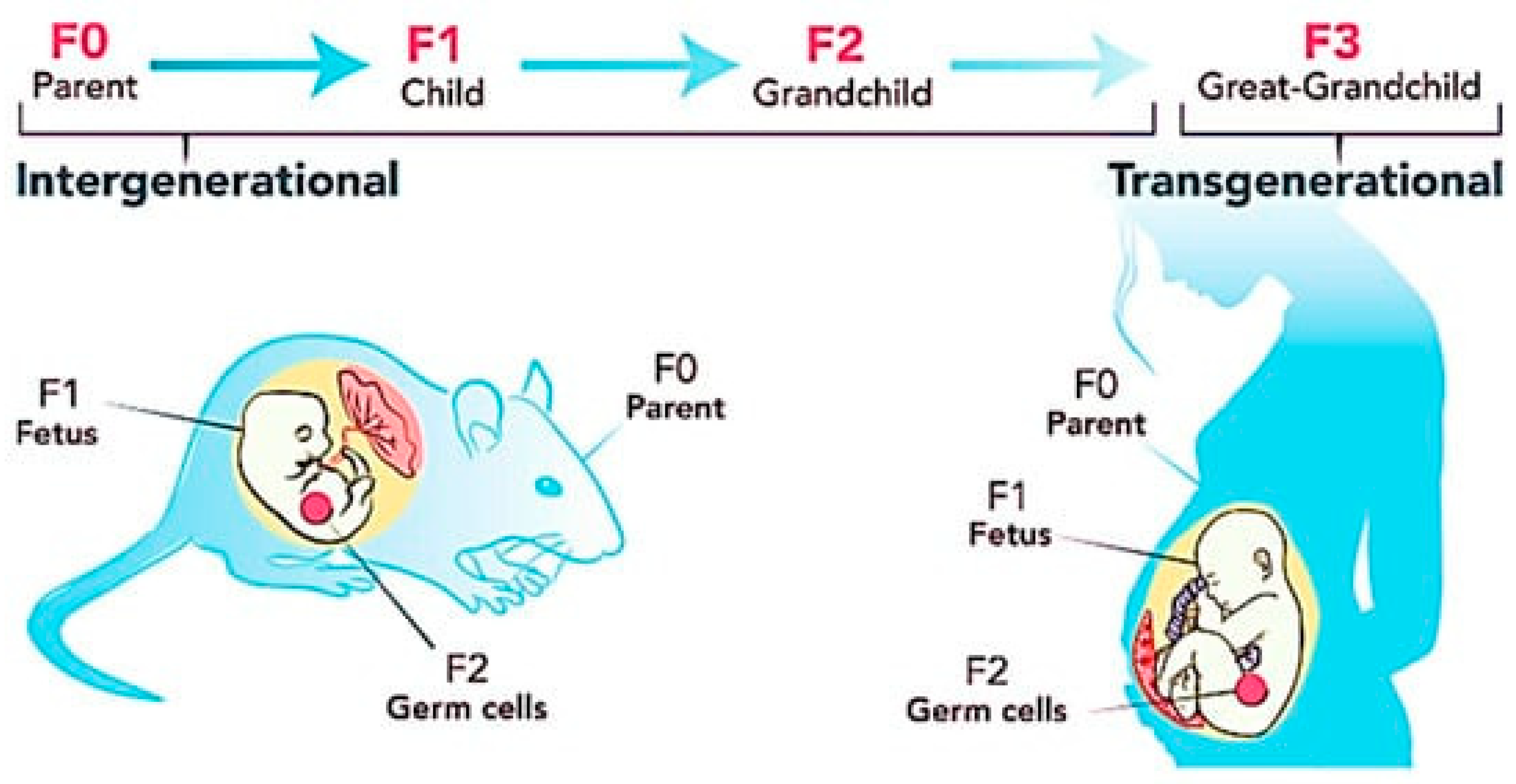
2. Transgenerational Inheritance of Obesity
3. Current Available Treatment and Its Side Effects
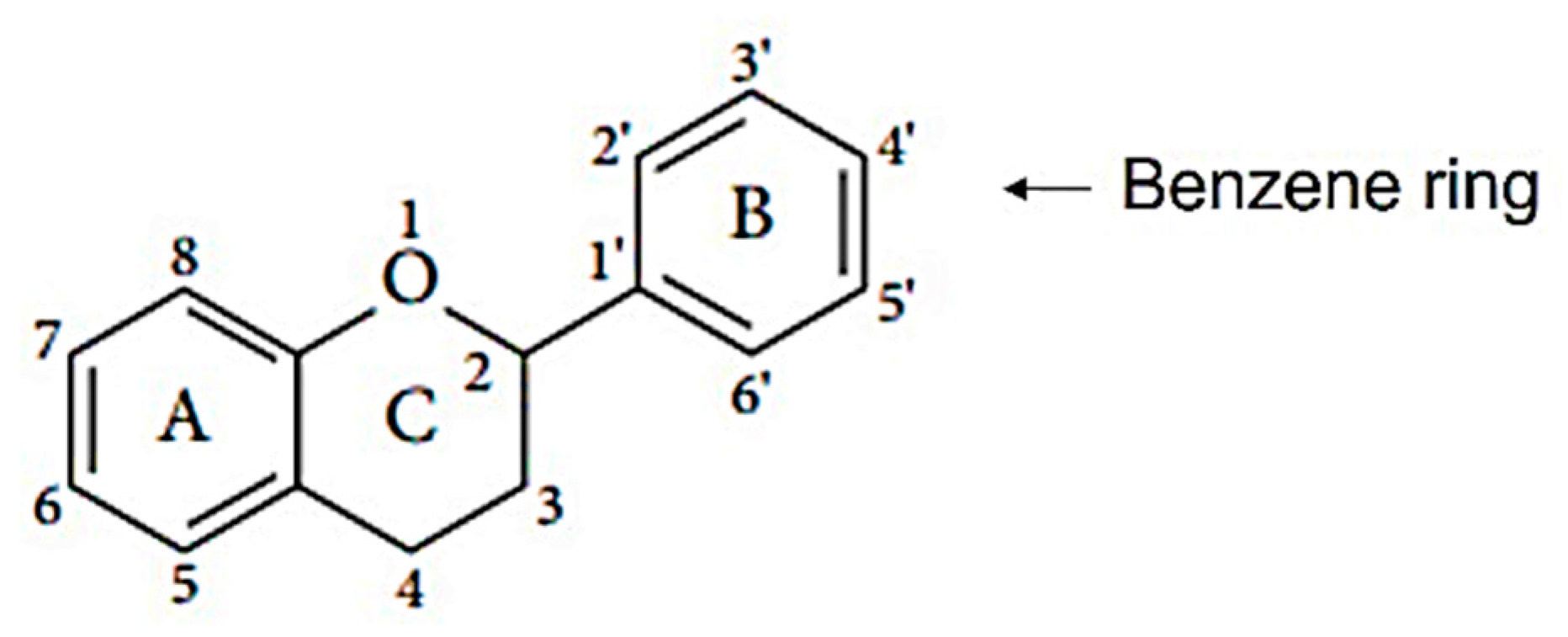
4. Flavonoids and Their Sources

{kind=link}
{kind=link}
{kind=link}
| Classification | Structure | Sources | Examples of Compounds |
|---|---|---|---|
| Flavonols [46,47] |  | Apples, tea, wine, broccoli, black tea, olive oil | Kaempferol, quercetin, dihydroquercetin, prunetin, and rutin |
| Flavanols [46] |  | Coco, dark chocolate, blackberry, green tee, apple, wine, kiwi | Catechin, gallocatechin, and epicatechin, |
| Flavanones [46,54] |  | Citrus fruit, orange, tomato, lemon | Eriodictyol, naringenin, hesperetin, isosakuranetin, and their glycosides |
| Flavones [46,54,55] |  | Legumes, cereal, angiosperms, mint, ginkgo biloba, thyme | Apingenin, Luteolin flavone, and their glycosides |
| Isoflavones [46] |  | lentils, fava beans, soybeans | Genistein, daidzein, glycitein, equol, biochanin A, and formononetein |
| Anthocyanidins [56,57] |  | Berries, currants, grapes, blue-colored leafy vegetables, grains, cranberries | Malvidin, cyanidin, delphinidin, pelargonidin, and petunidin |
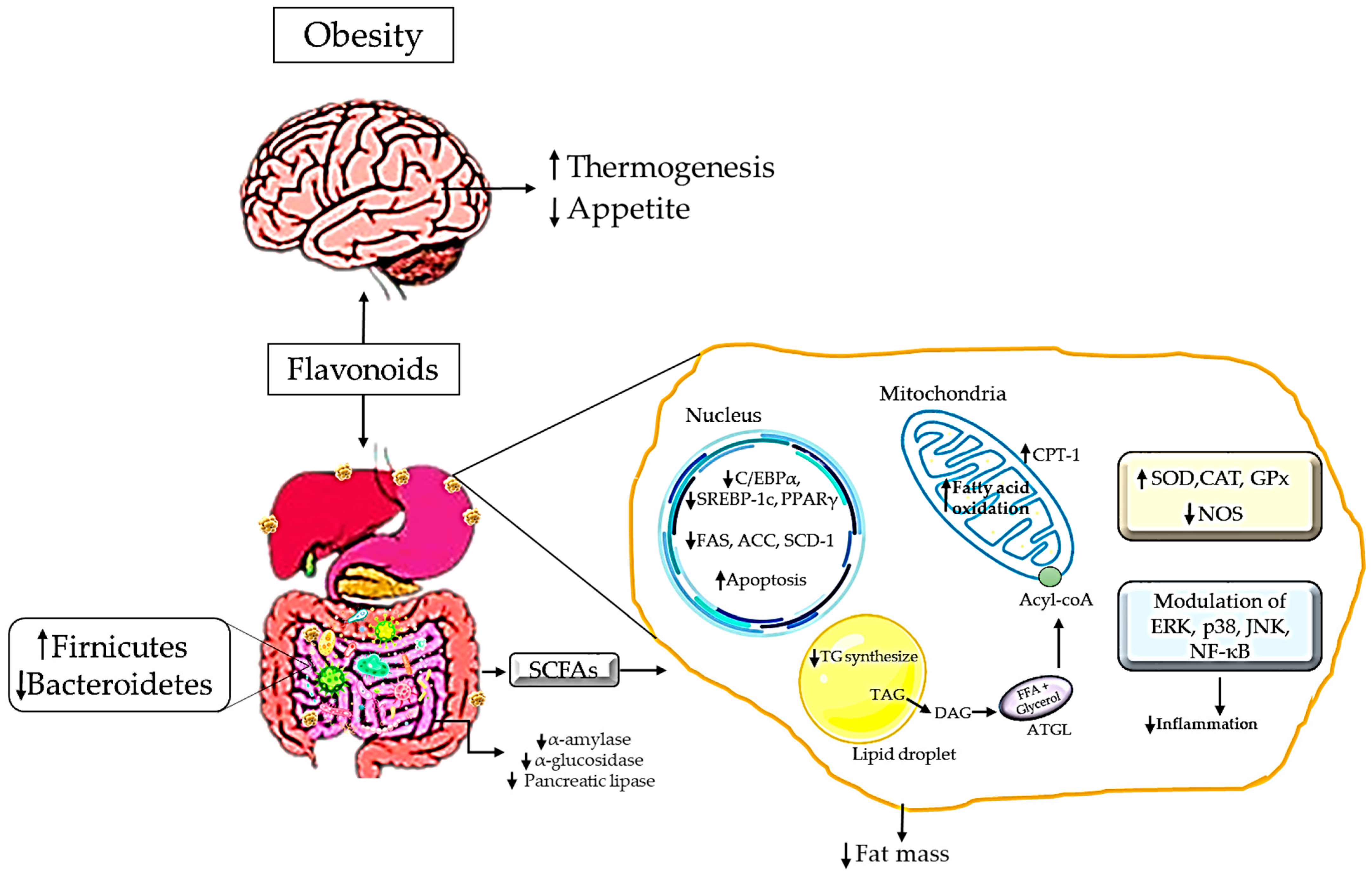
5. Mechanism of Action of Flavonoids as a Natural Antioxidant in Managing Obesity
| Author | Flavonoids | Animal | Dosage and Duration | Mode of Intervention | Findings |
|---|---|---|---|---|---|
| Lu et al., 2013 [108] | Citrange Fruit Extracts | 6-week-old female C57BL/6 mice | 1% w/w for 8 weeks | Diet |
|
| Xu et al., 2013 [109] | Luteolin | 5-week-old C57BL/6 mice | 0.002% or 0.01% for 6 weeks | Diet |
|
| Dong et al., 2014 [110] | Quercetin | 4-week-old C57BL/6 mice | 0.1% w/w for 12 weeks | Diet |
|
| Tan et al., 2014 [111] | Fortunella margarita fruit extract | 12-week-old female C57BL/6 mice | 1% w/w for 8 weeks | Diet |
|
| Assini et al., 2015 [112] | Naringenin | 8–12 weeks of male Fgf21−/− C57BL6/J mice | 3% w/w for 4 weeks | Diet |
|
| Zang et al., 2015 [113] | Kaempferol glycosides | 6-week-old male C57BL/6J mice | 0.15% for 92 days | Diet |
|
| Bao et al., 2016 [114] | Decussatin | 6-week-old male Wistar rats | 20 mg/kg for 12 weeks | Diet |
|
| Pan et al., 2016 [115] | Lychee extracts, green tea polyphenols, and citrus polymethoxyflavones | 4-week-old male C57BL/6J mice | 0.1 or 0.5% for 16 weeks | Diet |
|
| Ke et al., 2017 [116] | Naringenin | 20-week-old C57BL/6J mice | 1 or 3% for 13 days | Diet |
|
| Kwon et al., 2017 [117] | Flavonoid glycosides extract from Seabuckthorn leaves | 4-week-old male C57BL/6J mice | 0.04% for 12 weeks | Diet |
|
| Burke et al., 2018 [118] | Naringenin, Nobiletin | 10–12-week-old Ldlr−/− mice | 0.3 or 3% for 12 weeks | Diet |
|
| Sheng et al., 2019 [119] | Mulberry leaves | 4-week-old C57BL/6J mice | 20% for 13 weeks | Diet |
|
| Bian et al., 2021 [120] | Kaempferol | 8-week-old male C57BL/6J mice | 0.1% for 16 weeks | Diet |
|
| Miranda et al., 2016 [121] | Xanthohumol | 8-week-old male C57BL/6J mice, | 0.033 or 0.066% for 12 weeks | Pellet |
|
| Nascimento et al., 2013 [122] | Camu–camu pulp | Male wistar rats | 25 mL for 12 weeks | Ingestion |
|
| Alam et al., 2013 [123] | Naringin | 9–10-week-old male wistar rats | 100 mg/kg/day for 8 weeks | Oral gavage |
|
| You et al., 2013 [124] | Nelumbo nucifera | 4-week-old Sprague Dawley rats | 400 mg kg/day for 7 weeks | Oral gavage |
|
| Yoshida et al., 2014 [125] | Naringenin | 7-week-old male C57BL/6J mice | 100 mg/kg/day for 1, 7, and 14 days | Oral gavage |
|
| Baselga-Escudero et al., 2015 [126] | Proanthocyanidins | 6-week-old female Wistar rats | 5, 25, or 50 mg/kg for 3 weeks | Oral gavage |
|
| Miyata et al., 2015 [127] | Xanthohumol | 6-week-old male C57BL/6J mice | 75 or 150 mg/kg/day for 3 days | Oral gavage |
|
| Qin et al., 2015 [128] | 3-O-[(E)-4-(4-cyanophenyl)-2-oxobut-3-en-1-yl] kaempferol | 4-week-old Male C57BL/6 mice | 2, 5 or 50 mg/kg for 4 weeks | Oral gavage |
|
| Liu et al., 2017 [129] | Flavonoid rich extract of paulownia fortunei flowers | Male ICR mice | 50 and 100 mg/kg for 8 weeks | Oral gavage |
|
| Gentile et al., 2018 [130] | Apigenin | 6-week-old male C57BL/6 mice | 1, 10 or 30 mg/kg/day for 8 weeks | Oral gavage |
|
| Bai et al., 2019 [131] | Quzhou Fructus Aurantii extract | 8-week-old male C57BL/6J mice | 300 mg/kg for 12 weeks | Oral gavage |
|
| Feng et al., 2016 [67] | Apigenin | 3 to 4-week-old male C57BL/6J mice | 10, 30, or 50 mg/kg for 21 days | Intraperitoneal Injection |
|
| Varshney et al., 2019 [132] | Quercetin, rutin, kaempferol, and myricetin | 7-week-old male C57BL/6 mice | 25 mg/kg/day for 4 weeks | Intraperitoneal injection |
|
| Romero-Juárez et al., 2021 [133] | Kaempferol | 8-week-old C57Bl/6J male mice | 0.5 mg/kg/day for 40 days | Intraperitoneal injection |
|
| Yang et al., 2016 [134] | Flavonoid-enriched extract from Hippophae rhamnoides | 8-week-old C57BL/6 mice | 100 or 300 mg/kg for 9 weeks | Intragastric administration |
|
| Dai et al., 2018 [135] | Baicalin | Mice | 400 mg/kg/day for 12 weeks | Intragastric administration |
|
| Shen et al., 2019 [136] | Neohesperidin, hesperidin, and naringin | 4-week-old male C57BL/6 mice | 50, 100, or 200 mg/kg/ day for 12 weeks | Intragastric gavage |
|
| Su et al., 2019 [137] | Apigenin | C57BL/6 male mice | 15 or 30 mg/kg/day for 13 days | Subcutaneous injection |
|
6. Flavonoids in the Modulation of Gut Microbiota in Obesity
7. Flavonoids in Redox Modulation in Obesity
8. Bioavailability of Flavonoids
9. Conclusions
Author Contributions
Funding
Conflicts of Interest
References
- Kong, L.; Norstedt, G.; Schalling, M.; Gissler, M.; Lavebratt, C. The risk of offspring psychiatric disorders in the setting of maternal obesity and diabetes. Pediatrics 2018, 142, e20180776. [Google Scholar] [CrossRef]
- Slack, E.; Brandon, H.; Heslehurst, N. Obesity and Pregnancy. In Practical Guide to Obesity Medicine; Weaver, J.U., Ed.; Elsevier: Newcastle upon Tyne, UK, 2018; pp. 143–151. ISBN 978-0-323-48559-3. [Google Scholar]
- Al Beloushi, M.; Doshani, A.; Konje, J.C. Obesity, insulin resistance, and placental dysfunction-fetal growth. In Obesity and Obstetrics; Mahmood, T.A., Arulkumaran, S., Chervenak, F.A., Eds.; Elsevier: Doha, Qatar, 2020; pp. 191–197. ISBN 978-0-12-817921-5. [Google Scholar]
- King, S.E.; Skinner, M.K. Epigenetic Transgenerational Inheritance of Obesity Susceptibility. Trends Endocrinol. Metab. 2020, 31, 494. [Google Scholar] [CrossRef] [PubMed]
- Stirrat, L.I.; Reynolds, R.M. Effects of maternal obesity on early and long-term outcomes for offspring. Res. Rep. Neonatol. 2014, 4, 43–53. [Google Scholar] [CrossRef][Green Version]
- Şanlı, E.; Kabaran, S. Maternal Obesity, Maternal Overnutrition and Fetal Programming: Effects of Epigenetic Mechanisms on the Development of Metabolic Disorders. Curr. Genomics 2019, 20, 427. [Google Scholar] [CrossRef]
- Mina, T.H.; Lahti, M.; Drake, A.J.; Denison, F.C.; Räikkönen, K.; Norman, J.E.; Reynolds, R.M. Prenatal exposure to maternal very severe obesity is associated with impaired neurodevelopment and executive functioning in children. Pediatr. Res. 2017, 82, 47–54. [Google Scholar] [CrossRef] [PubMed]
- Rivera, H.M.; Christiansen, K.J.; Sullivan, E.L. The role of maternal obesity in the risk of neuropsychiatric disorders. Front. Neurosci. 2015, 9, 194. [Google Scholar] [CrossRef]
- Gillman, M.W. Interrupting Intergenerational Cycles of Maternal Obesity. Nestle. Nutr. Inst. Work. Ser. 2016, 85, 59–69. [Google Scholar] [CrossRef]
- Dunn, G.A.; Bale, T.L. Maternal high-fat diet effects on third-generation female body size via the paternal lineage. Endocrinology 2010, 152, 2228–2236. [Google Scholar] [CrossRef]
- Sarker, G.; Litwan, K.; Kastli, R.; Peleg-Raibstein, D. Maternal overnutrition during critical developmental periods leads to different health adversities in the offspring: Relevance of obesity, addiction and schizophrenia. Sci. Rep. 2019, 9, 17322. [Google Scholar] [CrossRef]
- Peral-Sanchez, I.; Hojeij, B.; Ojeda, D.A.; Steegers-Theunissen, R.P.M.; Willaime-Morawek, S. Epigenetics in the Uterine Environment: How Maternal Diet and ART May Influence the Epigenome in the Offspring with Long-Term Health Consequences. Genes 2022, 13, 31. [Google Scholar] [CrossRef]
- Chen, Z.; Zuo, X.; He, D.; Ding, S.; Xu, F.; Yang, H.; Jin, X.; Fan, Y.; Ying, L.; Tian, C.; et al. Long-term exposure to a “safe” dose of bisphenol A reduced protein acetylation in adult rat testes. Sci. Rep. 2017, 9, 40337. [Google Scholar] [CrossRef]
- Manikkam, M.; Tracey, R.; Guerrero-Bosagna, C.; Skinner, M.K. Plastics Derived Endocrine Disruptors (BPA, DEHP and DBP) Induce Epigenetic Transgenerational Inheritance of Obesity, Reproductive Disease and Sperm Epimutations. PLoS ONE 2013, 8, e55387. [Google Scholar] [CrossRef]
- Naomi, R.; Yazid, M.D.; Bahari, H.; Keong, Y.Y.; Rajandram, R.; Embong, H.; Teoh, S.H.; Halim, S.; Othman, F. Bisphenol A (BPA) Leading to Obesity and Cardiovascular Complications: A Compilation of Current In Vivo Study. Int. J. Mol. Sci. 2022, 23, 2969. [Google Scholar] [CrossRef]
- Motawi, T.K.; Shaker, O.G.; Ismail, M.F.; Sayed, N.H. Peroxisome Proliferator-Activated Receptor Gamma in Obesity and Colorectal Cancer: The Role of Epigenetics. Sci. Rep. 2017, 7, 10714. [Google Scholar] [CrossRef]
- Yan, F.; Shen, N.; Pang, J.X.; Zhang, Y.W.; Rao, E.Y.; Bode, A.M.; Al-Kali, A.; Zhang, D.E.; Litzow, M.R.; Li, B.; et al. Fatty acid-binding protein FABP4 mechanistically links obesity with aggressive AML by enhancing aberrant DNA methylation in AML cells. Leukemia 2016, 31, 1434–1442. [Google Scholar] [CrossRef]
- Ruiz, R.; Jideonwo, V.; Ahn, M.; Surendran, S.; Tagliabracci, V.S.; Hou, Y.; Gamble, A.; Kerner, J.; Irimia-Dominguez, J.M.; Puchowicz, M.A.; et al. Sterol regulatory element-binding protein-1 (SREBP-1) is required to regulate glycogen synthesis and gluconeogenic gene expression in mouse liver. J. Biol. Chem. 2014, 289, 5510–5517. [Google Scholar] [CrossRef] [PubMed]
- Marraudino, M.; Bo, E.; Carlini, E.; Farinetti, A.; Ponti, G.; Zanella, I.; Di Lorenzo, D.; Panzica, G.C.; Gotti, S. Hypothalamic Expression of Neuropeptide Y (NPY) and Pro-OpioMelanoCortin (POMC) in Adult Male Mice Is Affected by Chronic Exposure to Endocrine Disruptors. Metabolites 2021, 11, 368. [Google Scholar] [CrossRef] [PubMed]
- Candler, T.; Kühnen, P.; Prentice, A.M.; Silver, M. Epigenetic regulation of POMC; implications for nutritional programming, obesity and metabolic disease. Front. Neuroendocrinol. 2019, 54, 100773. [Google Scholar] [CrossRef] [PubMed]
- Parlee, S.D.; MacDougald, O.A. Maternal Nutrition and Risk of Obesity in Offspring: The Trojan Horse of Developmental Plasticity. Biochim. Biophys. Acta 2013, 1842, 495. [Google Scholar] [CrossRef]
- Guo, J.; Wu, J.; He, Q.; Zhang, M.; Li, H.; Liu, Y. The Potential Role of PPARs in the Fetal Origins of. Cells 2022, 11, 3474. [Google Scholar] [CrossRef] [PubMed]
- Dubois, V.; Eeckhoute, J.; Lefebvre, P.; Staels, B. Distinct but complementary contributions of PPAR isotypes to energy homeostasis. J. Clin. Investig. 2017, 127, 1202–1214. [Google Scholar] [CrossRef] [PubMed]
- Girousse, A.; Tavernier, G.; Valle, C.; Moro, C.; Mejhert, N.; Dinel, A.L.; Houssier, M.; Roussel, B.; Besse-Patin, A.; Combes, M.; et al. Partial Inhibition of Adipose Tissue Lipolysis Improves Glucose Metabolism and Insulin Sensitivity Without Alteration of Fat Mass. PLoS Biol. 2013, 11, e1001485. [Google Scholar] [CrossRef]
- Catalano, P.M.; Nizielski, S.E.; Shao, J.; Preston, L.; Qiao, L.; Friedman, J.E.; Patrick, M.; Nizielski, S.E.; Preston, L.; Qiao, L.; et al. Downregulated IRS-1 and PPARγ in obese women with gestational diabetes: Relationship to FFA during pregnancy. Am. J. Physiol. Endocrinol. Metab. 2001, 80262, 522–533. [Google Scholar] [CrossRef]
- Liao, W.; Nguyen, M.T.A.; Yoshizaki, T.; Favelyukis, S.; Patsouris, D.; Imamura, T.; Verma, I.M.; Olefsky, J.M. Suppression of PPARγ attenuates insulin-stimulated glucose uptake by affecting both GLUT1 and GLUT4 in 3T3-L1 adipocytes. Am. J. Physiol. Endocrinol. Metab. 2006, 293, 219–227. [Google Scholar] [CrossRef]
- Joseph, R.; Poschmann, J.; Sukarieh, R.; Too, P.G.; Julien, S.G.; Xu, F.; The, A.L.; Holbrook, J.D.; Ng, K.L.; Chong, Y.S.; et al. ACSL1 is associated with fetal programming of insulin sensitivity and cellular lipid content. Mol. Endocrinol. 2015, 29, 909–920. [Google Scholar] [CrossRef]
- McDonnell, E.; Crown, S.B.; Fox, D.B.; Kitir, B.; Ilkayeva, O.R.; Olsen, C.A.; Grimsrud, P.A.; Hirschey, M.D. Lipids Reprogram Metabolism to Become a Major Carbon Source for Histone Acetylation. Cell Rep. 2016, 17, 1463–1472. [Google Scholar] [CrossRef]
- Alexander, D.C.; Corman, T.; Mendoza, M.; Glass, A.; Belity, T.; Wu, R.; Campbell, R.R.; Han, J.; Keiser, A.A.; Winkler, J.; et al. Targeting acetyl-CoA metabolism attenuates the formation of fear memories through reduced activity-dependent histone acetylation. Proc. Natl. Acad. Sci. USA 2022, 119, e2114758119. [Google Scholar] [CrossRef]
- James, W.P.T.; Caterson, I.D.; Coutinho, W.; Finer, N.; Van Gaal, L.F.; Maggioni, A.P.; Torp-Pedersen, C.; Sharma, A.M.; Shepherd, G.M.; Rode, R.A.; et al. Effect of sibutramine on cardiovascular outcomes in overweight and obese subjects. N. Engl. J. Med. 2010, 363, 905–917. [Google Scholar] [CrossRef] [PubMed]
- Sharretts, J.; Galescu, O.; Gomatam, S.; Andraca-Carrera, E.; Hampp, C.; Yanoff, L. Cancer Risk Associated with Lorcaserin-The FDA’s Review of the CAMELLIA-TIMI 61 Trial. N. Engl. J. Med. 2020, 383, 1000–1002. [Google Scholar] [CrossRef] [PubMed]
- Onge, E.S.; Miller, S.A.; Motycka, C. Liraglutide (Saxenda®) as a Treatment for Obesity. Food Nutr. Sci. 2016, 07, 227–235. [Google Scholar] [CrossRef]
- Drew, B.S.; Dixon, A.F.; Dixon, J.B. Obesity management: Update on orlistat. Vasc. Health Risk Manag. 2007, 3, 817–821. [Google Scholar] [PubMed]
- Tonstad, S.; Rössner, S.; Rissanen, A.; Astrup, A. Medical management of obesity in Scandinavia 2016. Obes. Med. 2016, 1, 38–44. [Google Scholar] [CrossRef]
- DiNicolantonio, J.J.; Chatterjee, S.; O’Keefe, J.H.; Meier, P. Lorcaserin for the treatment of obesity? A closer look at its side effects. Open Heart 2014, 1, e000173. [Google Scholar] [CrossRef] [PubMed]
- Shin, J.H.; Gadde, K.M. Clinical utility of phentermine/topiramate (QsymiaTM) combination for the treatment of obesity. Diabetes Metab. Syndr. Obes. 2013, 6, 131–139. [Google Scholar] [CrossRef]
- Pocai, A. Action and therapeutic potential of oxyntomodulin. Mol. Metab. 2013, 3, 251. [Google Scholar] [CrossRef]
- Kim, D.D.; Krishnarajah, J.; Lillioja, S.; de Looze, F.; Marjason, J.; Proietto, J.; Shakib, S.; Stuckey, B.G.A.; Vath, J.E.; Hughes, T.E. Efficacy and safety of beloranib for weight loss in obese adults: A randomized controlled trial. Diabetes Obes. Metab. 2015, 17, 566–572. [Google Scholar] [CrossRef]
- Biobaku, F.; Ghanim, H.; Monte, S.V.; Caruana, J.A.; Dandona, P. Bariatric Surgery: Remission of Inflammation, Cardiometabolic Benefits, and Common Adverse Effects. J. Endocr. Soc. 2020, 4, bvaa049. [Google Scholar] [CrossRef]
- Eid, I.; Birch, D.W.; Sharma, A.M.; Sherman, V.; Shahzeer, K. Complications associated with adjustable gastric banding for morbid obesity: A surgeon’s guide. Can. J. Surg. 2011, 54, 66. [Google Scholar] [CrossRef]
- Higa, K.D.; Boone, K.B.; Ho, T. Complications of the laparoscopic Roux-en-Y gastric bypass: 1, 040 Patients—What have we learned? Obes. Surg. 2000, 10, 509–513. [Google Scholar] [CrossRef]
- Himpens, J.; Dobbeleir, J.; Peeters, G. Long-term results of laparoscopic sleeve gastrectomy for obesity. Ann. Surg. 2010, 252, 319–324. [Google Scholar] [CrossRef]
- Marceau, P.; Hould, F.S.; Simard, S.; Lebel, S.; Bourque, R.A.; Potvin, M.; Biron, S. Biliopancreatic diversion with duodenal switch. World J. Surg. 1998, 22, 947–954. [Google Scholar] [CrossRef] [PubMed]
- Al-momen, A.; Ireland, M.; El-mogy, I. Intragastric Balloon for Obesity: A Retrospective Evaluation of Tolerance and Efficacy. Obes. Surg. 2005, 15, 101–105. [Google Scholar] [CrossRef] [PubMed]
- Chen, L.; Cao, H.; Huang, Q.; Xiao, J.; Teng, H. Absorption, metabolism and bioavailability of flavonoids: A review. Crit. Rev. Food Sci. Nutr. 2021, 62, 7730–7742. [Google Scholar] [CrossRef]
- Ruiz-Cruz, S.; Chaparro-Hernández, S.; Hernández-Ruiz, K.L.; Cira-Chávez, L.A.; Estrada-Alvarado, M.I.; GassosOrtega, L.E.; de Ornelas-Paz, J.J.; Mata, M.A.L.; Ruiz-Cruz, S.; Chaparro-Hernández, S.; et al. Flavonoids: Important Biocompounds in Food. In Flavonoids—From Biosynthesis to Human Health; Justino, G.C., Ed.; IntechOpen: London, UK, 2017; pp. 353–369. ISBN 978-953-51-3424-4. [Google Scholar]
- Roy, A.; Khan, A.; Ahmad, I.; Alghamdi, S.; Rajab, B.S.; Babalghith, A.O.; Alshahrani, M.Y.; Islam, S.; Islam, M.R. Flavonoids a Bioactive Compound from Medicinal Plants and Its Therapeutic Applications. BioMed Res. Int. 2022, 2022, 5445291. [Google Scholar] [CrossRef] [PubMed]
- Kutaiba Ibrahim, A.; Mohamed Abdalkarim, M. Flavonoids: Chemistry, Biochemistry and Antioxidant activity. J. Pharm. Res. 2012, 5, 4013–4020. [Google Scholar]
- Lodhi, S.; Kori, M.L. Structure—Activity Relationship and Therapeutic Benefits of Flavonoids in the Management of Diabetes and Associated Disorders. Pharm. Chem. J. 2021, 54, 1106–1125. [Google Scholar] [CrossRef]
- Zhang, Q.; Yang, W.; Liu, J.; Liu, H.; Lv, Z.; Zhang, C.; Chen, D.; Jiao, Z. Identification of Six Flavonoids as Novel Cellular Antioxidants and Their Structure-Activity Relationship. Oxid. Med. Cell. Longev. 2020, 2020, 4150897. [Google Scholar] [CrossRef]
- Song, D.; Cheng, L.; Zhang, X.; Wu, Z.; Zheng, X. The modulatory effect and the mechanism of flavonoids on obesity. J. Food Biochem. 2019, 43, e12954. [Google Scholar] [CrossRef]
- Chen, L.; Teng, H.; Xie, Z.; Cao, H.; Cheang, W.S.; Skalicka-Woniak, K.; Georgiev, M.I.; Xiao, J. Modifications of dietary flavonoids towards improved bioactivity: An update on structure–activity relationship. Crit. Rev. Food Sci. Nutr. 2016, 58, 513–527. [Google Scholar] [CrossRef]
- Mehmood, S.; Maqsood, M.; Mahtab, N.; Khan, M.I.; Sahar, A.; Zaib, S.; Gul, S. Epigallocatechin gallate: Phytochemistry, bioavailability, utilization challenges, and strategies. J. Food Biochem. 2022, 46, e14189. [Google Scholar] [CrossRef]
- Tajammal, A.; Siddiqa, A.; Irfan, A.; Azam, M.; Hafeez, H.; Munawar, M.A.; Basra, M.A.R. Antioxidant, molecular docking and computational investigation of new flavonoids. J. Mol. Struct. 2021, 1254, 132189. [Google Scholar] [CrossRef]
- Li, R.; Wang, Q.; Peng, H.; Zhao, G.; Zhang, D.; Li, Z. Exploring the effect of microwave treatment on phenolic flavonoids, antioxidant capacity, and phenolic in vitro bioaccessibility of sorghum. Int. J. Food Sci. Technol. 2022, 57, 2510–2522. [Google Scholar] [CrossRef]
- Di Lorenzo, C.; Colombo, F.; Biella, S.; Stockley, C.; Restani, P. Polyphenols and Human Health: The Role of Bioavailability. Nutrients 2021, 13, 273. [Google Scholar] [CrossRef]
- Mu, J.; Klosek, A.; Brennan, M.A.; Chen, Z.; Hui, X.; Wu, G.; Brennan, C.S. The effects of blackcurrant powder (Ribes nigrum) supplementation on pasting properties, physicochemical properties, and nutritive values of starch derived from mung bean (Vigna radiata L.) and pea (Pisum sativum L.). Int. J. Food Sci. Technol. 2021, 56, 4408–4416. [Google Scholar] [CrossRef]
- Bionaz, M.; Chen, S.; Khan, M.J.; Loor, J.J. Functional role of PPARs in ruminants: Potential targets for fine-tuning metabolism during growth and lactation. PPAR Res. 2013, 2013, 684159. [Google Scholar] [CrossRef]
- Loor, J.J. Nutrigenomics in livestock: Potential role in physiological regulation and practical applications. Anim. Prod. Sci. 2022, 62, 901–912. [Google Scholar] [CrossRef]
- Wu, T.T.L.; Tong, Y.C.; Chen, I.H.; Niu, H.S.; Li, Y.; Cheng, J.T. Induction of apoptosis in prostate cancer by ginsenoside Rh2. Oncotarget 2018, 9, 11109–11118. [Google Scholar] [CrossRef]
- Silva, O.A.; Ribeiro-Filho, H.V.; Avelino, T.M.; Tittanegro, T.H.; Figueira, A.C.M.; Rabelo, L.A.; Pitta, I.d.R.; Lahlou, S.; Duarte, G.P. GQ-130, a novel analogue of thiazolidinedione, improves obesity-induced metabolic alterations in rats: Evidence for the involvement of PPARβ/δ pathway. Clin. Exp. Pharmacol. Physiol. 2020, 47, 798–808. [Google Scholar] [CrossRef]
- Ament, Z.; Masoodi, M.; Griffin, J.L. Applications of metabolomics for understanding the action of peroxisome proliferator-activated receptors (PPARs) in diabetes, obesity and cancer. Genome Med. 2012, 4, 32. [Google Scholar] [CrossRef] [PubMed]
- Stienstra, R.; Duval, C.; Müller, M.; Kersten, S. PPARs, Obesity, and Inflammation. PPAR Res. 2007, 2007, 95974. [Google Scholar] [CrossRef]
- Zhang, Y.; Dallner, O.S.; Nakadai, T.; Fayzikhodjaeva, G.; Lu, Y.H.; Lazar, M.A.; Roeder, R.G.; Friedman, J.M. A noncanonical PPARγ/RXRα-binding sequence regulates leptin expression in response to changes in adipose tissue mass. Proc. Natl. Acad. Sci. USA 2018, 115, E6039–E6047. [Google Scholar] [CrossRef]
- Giorgino, F.; Leonardini, A.; Laviola, L.; Perrini, S.; Natalicchio, A. Cross-Talk between PPARγ and Insulin Signaling and Modulation of Insulin Sensitivity. PPAR Res. 2009, 2009, 818945. [Google Scholar] [CrossRef]
- Teixeira, C.; Sousa, A.P.; Santos, I.; Rocha, A.C.; Alencastre, I.; Pereira, A.C.; Martins-Mendes, D.; Barata, P.; Baylina, P.; Fernandes, R. Enhanced 3T3-L1 Differentiation into Adipocytes by Pioglitazone Pharmacological Activation of Peroxisome Proliferator Activated Receptor-Gamma (PPAR-γ). Biology 2022, 11, 806. [Google Scholar] [CrossRef]
- Feng, X.; Weng, D.; Zhou, F.; Owen, Y.D.; Qin, H.; Zhao, J.; Wen, Y.; Huang, Y.; Chen, J.; Fu, H.; et al. Activation of PPARγ by a Natural Flavonoid Modulator, Apigenin Ameliorates Obesity-Related Inflammation Via Regulation of Macrophage Polarization. EBioMedicine 2016, 9, 61–76. [Google Scholar] [CrossRef] [PubMed]
- Baker, R.G.; Hayden, M.S.; Ghosh, S. NF-κB, inflammation, and metabolic disease. Cell Metab. 2010, 13, 11–22. [Google Scholar] [CrossRef] [PubMed]
- Xu, Y.; Li, X.; Wang, H. Protective Roles of Apigenin Against Cardiometabolic Diseases: A Systematic Review. Front. Nutr. 2022, 9, 875826. [Google Scholar] [CrossRef]
- Williamson, G.; Kay, C.D.; Crozier, A. The Bioavailability, Transport, and Bioactivity of Dietary Flavonoids: A Review from a Historical Perspective. Compr. Rev. Food Sci. Food Saf. 2018, 17, 1054–1112. [Google Scholar] [CrossRef]
- Itoh, K.; Hikage, F.; Ida, Y.; Ohguro, H. Prostaglandin F2α agonists negatively modulate the size of 3d organoids from primary human orbital fibroblasts. Investig. Ophthalmol. Vis. Sci. 2020, 61, 13. [Google Scholar] [CrossRef]
- Sui, G.G.; Xiao, H.B.; Lu, X.Y.; Sun, Z.L. Naringin Activates AMPK Resulting in Altered Expression of SREBPs, PCSK9, and LDLR to Reduce Body Weight in Obese C57BL/6J Mice. J. Agric. Food Chem. 2018, 66, 8983–8990. [Google Scholar] [CrossRef]
- Pan, Y.; Cao, S.; Tang, J.; Arroyo, J.P.; Terker, A.S.; Wang, Y.; Niu, A.; Fan, X.; Wang, S.; Zhang, Y.; et al. Cyclooxygenase-2 in adipose tissue macrophages limits adipose tissue dysfunction in obese mice. J. Clin. Investig. 2022, 132, e152391. [Google Scholar] [CrossRef]
- Kim, B.S.; Pallua, N.; Bernhagen, J.; Bucala, R. The macrophage migration inhibitory factor protein superfamily in obesity and wound repair. Exp. Mol. Med. 2015, 47, e161. [Google Scholar] [CrossRef]
- Cha, M.H.; Kim, I.C.; Lee, B.H.; Yoon, Y. Baicalein inhibits adipocyte differentiation by enhancing COX-2 expression. J. Med. Food 2006, 9, 145–153. [Google Scholar] [CrossRef] [PubMed]
- Liu, T.; Zhang, L.; Joo, D.; Sun, S.C. NF-κB signaling in inflammation. Signal Transduct. Target. Ther. 2017, 2, e17023. [Google Scholar] [CrossRef] [PubMed]
- Wang, C.; Zhang, X.; Luo, L.; Luo, Y.; Wu, D.; Spilca, D.; Le, Q.; Yang, X.; Alvarez, K.; Hines, W.C.; et al. COX-2 Deficiency Promotes White Adipogenesis via PGE2-Mediated Paracrine Mechanism and Exacerbates Diet-Induced Obesity. Cells 2022, 11, 1819. [Google Scholar] [CrossRef]
- Chan, P.C.; Liao, M.T.; Hsieh, P.S. The dualistic effect of COX-2-mediated signaling in obesity and insulin resistance. Int. J. Mol. Sci. 2019, 20, 3115. [Google Scholar] [CrossRef]
- Hoang, M.H.; Jia, Y.; Mok, B.; Jun, H.J.; Hwang, K.Y.; Lee, S.J. Kaempferol ameliorates symptoms of metabolic syndrome by regulating activities of liver X receptor-β. J. Nutr. Biochem. 2015, 26, 868–875. [Google Scholar] [CrossRef]
- Gathercole, L.L.; Morgan, S.A.; Tomlinson, J.W. Hormonal Regulation of Lipogenesis. In Vitamins and Hormones; Litwack, G., Ed.; Academic Press: Birmingham, UK, 2013; Volume 91, pp. 1–27. [Google Scholar]
- Hussain, T.; Tan, B.; Murtaza, G.; Liu, G.; Rahu, N.; Saleem Kalhoro, M.; Hussain Kalhoro, D.; Adebowale, T.O.; Usman Mazhar, M.; ur Rehman, Z.; et al. Flavonoids and type 2 diabetes: Evidence of efficacy in clinical and animal studies and delivery strategies to enhance their therapeutic efficacy. Pharmacol. Res. 2020, 152, 104629. [Google Scholar] [CrossRef] [PubMed]
- Brobst, D.F. Pancreatic Function. In Clinical Biochemistry of Domestic Animals; Kaneko, J.J., Ed.; Academic Press: Davis, CA, USA, 1980; pp. 259–281. ISBN 978-0-12-396350-5. [Google Scholar]
- Xiao, C.; Stahel, P.; Lewis, G.F. Regulation of Chylomicron Secretion: Focus on Post-Assembly Mechanisms. Cell. Mol. Gastroenterol. Hepatol. 2018, 7, 487–501. [Google Scholar] [CrossRef] [PubMed]
- Hasim, H.; Faridah, D.N.; Qomaliyah, E.N.; Afandi, F.A. Pancreatic Lipase Inhibition Activity in Lipid Absorption Using Traditional Plants: A Systematic Review and Meta-Analysis. Indones. J. Chem. 2023, 23, 568–582. [Google Scholar] [CrossRef]
- Seo, Y.S.; Kang, O.H.; Kim, S.B.; Mun, S.H.; Kang, D.H.; Yang, D.W.; Choi, J.G.; Lee, Y.M.; Kang, D.K.; Lee, H.S.; et al. Quercetin prevents adipogenesis by regulation of transcriptional factors and lipases in OP9 cells. Int. J. Mol. Med. 2015, 35, 1779–1785. [Google Scholar] [CrossRef]
- Kim, S.-J.; Tang, T.; Abbott, M.; Viscarra, J.A.; Wang, Y.; Sul, H.S. AMPK Phosphorylates Desnutrin/ATGL and Hormone-Sensitive Lipase To Regulate Lipolysis and Fatty Acid Oxidation within Adipose Tissue. Mol. Cell. Biol. 2016, 36, 1961–1976. [Google Scholar] [CrossRef]
- Alves-Bezerra, M.; Cohen, D.E. Triglyceride metabolism in the liver. Compr. Physiol. 2017, 8, 8. [Google Scholar] [CrossRef]
- Zhou, J.F.; Wang, W.J.; Yin, Z.P.; Zheng, G.D.; Chen, J.G.; Li, J.E.; Chen, L.L.; Zhang, Q.F. Quercetin is a promising pancreatic lipase inhibitor in reducing fat absorption in vivo. Food Biosci. 2021, 43, 101248. [Google Scholar] [CrossRef]
- Kumar, S.; Alagawadi, K.R. Anti-obesity effects of galangin, a pancreatic lipase inhibitor in cafeteria diet fed female rats. Pharm. Biol. 2012, 51, 607–613. [Google Scholar] [CrossRef] [PubMed]
- Gómez-Zorita, S.; Lasa, A.; Abendaño, N.; Fernández-Quintela, A.; Mosqueda-Solís, A.; Garcia-Sobreviela, M.P.; Arbonés-Mainar, J.M.; Portillo, M.P. Phenolic compounds apigenin, hesperidin and kaempferol reduce in vitro lipid accumulation in human adipocytes. J. Transl. Med. 2017, 15, 237. [Google Scholar] [CrossRef] [PubMed]
- Garcia, D.; Shaw, R.J. AMPK: Mechanisms of cellular energy sensing and restoration of metabolic balance. Mol. Cell 2017, 66, 800. [Google Scholar] [CrossRef] [PubMed]
- Kola, B.; Grossman, A.B.; Korbonits, M. The Role of AMP-Activated Protein Kinase in Obesity. Obe. Metab. 2008, 36, 198–211. [Google Scholar]
- Vijayakumar, R.S.; Lin, Y.; Shia, K.S.; Yeh, Y.N.; Hsieh, W.P.; Hsiao, W.C.; Chang, C.P.; Chao, Y.S.; Hung, M.S. Induction of fatty acid oxidation resists weight gain, ameliorates hepatic steatosis and reduces cardiometabolic risk factors. Int. J. Obes. 2011, 36, 999–1006. [Google Scholar] [CrossRef]
- Wu, C.; Kang, J.E.; Peng, L.J.; Li, H.; Khan, S.A.; Hillard, C.J.; Okar, D.A.; Lange, A.J. Enhancing hepatic glycolysis reduces obesity: Differential effects on lipogenesis depend on site of glycolytic modulation. Cell Metab. 2005, 2, 131–140. [Google Scholar] [CrossRef]
- Sandoval, V.; Sanz-Lamora, H.; Arias, G.; Marrero, P.F.; Haro, D.; Relat, J. Metabolic Impact of Flavonoids Consumption in Obesity: From Central to Peripheral. Nutrients 2020, 12, 2393. [Google Scholar] [CrossRef]
- Rajan, P.; Natraj, P.; Ranaweera, S.S.; Dayarathne, L.A.; Lee, Y.J.; Han, C.H. Anti-adipogenic effect of the flavonoids through the activation of AMPK in palmitate (PA)-treated HepG2 cells. J. Vet. Sci. 2022, 23, e4. [Google Scholar] [CrossRef] [PubMed]
- Kou, G.; Hu, Y.; Jiang, Z.; Li, Z.; Li, P.; Song, H.; Chen, Q.; Zhou, Z.; Lyu, Q. Citrus aurantium L. polymethoxyflavones promote thermogenesis of brown and white adipose tissue in high-fat diet induced C57BL/6J mice. J. Funct. Foods 2020, 67, 103860. [Google Scholar] [CrossRef]
- Arias, N.; Picó, C.; Teresa Macarulla, M.; Oliver, P.; Miranda, J.; Palou, A.; Portillo, M.P. A combination of resveratrol and quercetin induces browning in white adipose tissue of rats fed an obesogenic diet. Obesity 2017, 25, 111–121. [Google Scholar] [CrossRef] [PubMed]
- Wang, Z.; Yu, X.; Chen, Y. Recruitment of Thermogenic Fat: Trigger of Fat Burning. Front. Endocrinol. 2021, 12, 696505. [Google Scholar] [CrossRef] [PubMed]
- Zhang, X.; Li, X.; Fang, H.; Guo, F.; Li, F.; Chen, A.; Huang, S. Flavonoids as inducers of white adipose tissue browning and thermogenesis: Signalling pathways and molecular triggers. Nutr. Metab. 2019, 16, 47. [Google Scholar] [CrossRef]
- Lee, S.H.; Kim, B.; Oh, M.J.; Yoon, J.; Kim, H.Y.; Lee, K.J.; Lee, J.D.; Choi, K.Y. Persicaria hydropiper (L.) Spach and its Flavonoid Components, Isoquercitrin and Isorhamnetin, Activate the Wnt/β-catenin Pathway and Inhibit Adipocyte Differentiation of 3T3-L1 Cells. Phyther. Res. 2011, 25, 1629–1635. [Google Scholar] [CrossRef]
- Kennell, J.A.; MacDougald, O.A. Wnt signaling inhibits adipogenesis through beta-catenin-dependent and -independent mechanisms. J. Biol. Chem. 2005, 280, 24004–24010. [Google Scholar] [CrossRef]
- Liu, J.; Farmer, S.R. Regulating the balance between peroxisome proliferator-activated receptor gamma and beta-catenin signaling during adipogenesis. A glycogen synthase kinase 3beta phosphorylation-defective mutant of beta-catenin inhibits expression of a subset of adipogenic genes. J. Biol. Chem. 2004, 279, 45020–45027. [Google Scholar] [CrossRef]
- Al Shukor, N.; Ravallec, R.; Van Camp, J.; Raes, K.; Smagghe, G. Flavonoids stimulate cholecystokinin peptide secretion from the enteroendocrine STC-1 cells. Fitoterapia 2016, 113, 128–131. [Google Scholar] [CrossRef]
- Zhang, X.; Tang, N.; Qi, J.; Wang, S.; Hao, J.; Wu, Y.; Chen, H.; Tian, Z.; Wang, B.; Chen, D.; et al. CCK reduces the food intake mainly through CCK1R in Siberian sturgeon (Acipenser baerii Brandt). Sci. Rep. 2017, 7, 12413. [Google Scholar] [CrossRef]
- Cremonini, E.; Daveri, E.; Mastaloudis, A.; Oteiza, P.I. (–)-Epicatechin and Anthocyanins Modulate GLP-1 Metabolism: Evidence from C57BL/6J Mice and GLUTag Cells. J. Nutr. 2021, 151, 1506. [Google Scholar] [CrossRef]
- Shah, M.; Vella, A. Effects of GLP-1 on appetite and weight. Rev. Endocr. Metab. Disord. 2014, 15, 187. [Google Scholar] [CrossRef]
- Lu, Y.; Xi, W.; Ding, X.; Fan, S.; Zhang, Y.; Jiang, D.; Li, Y.; Huang, C.; Zhou, Z. Citrange Fruit Extracts Alleviate Obesity-Associated Metabolic Disorder in High-Fat Diet-Induced Obese C57BL/6 Mouse. Int. J. Mol. Sci. 2013, 14, 23736–23750. [Google Scholar] [CrossRef] [PubMed]
- Xu, N.; Zhang, L.; Dong, J.; Zhang, X.; Chen, Y.G.; Bao, B.; Liu, J. Low-dose diet supplement of a natural flavonoid, luteolin, ameliorates diet-induced obesity and insulin resistance in mice. Mol. Nutr. Food Res. 2013, 58, 1258–1268. [Google Scholar] [CrossRef]
- Dong, J.; Zhang, X.; Zhang, L.; Bian, H.X.; Xu, N.; Bao, B.; Liu, J. Quercetin reduces obesity-associated ATM infiltration and inflammation in mice: A mechanism including AMPKα1/SIRT. J. Lipid Res. 2014, 55, 363–374. [Google Scholar] [CrossRef]
- Tan, S.; Li, M.; Ding, X.; Fan, S.; Guo, L.; Gu, M.; Zhang, Y.; Feng, L.; Jiang, D.; Li, Y.; et al. Effects of Fortunella margarita fruit extract on metabolic disorders in high-fat diet-induced obese C57BL/6 mice. PLoS ONE 2014, 9, e93510. [Google Scholar] [CrossRef] [PubMed]
- Assini, J.M.; Mulvihill, E.E.; Burke, A.C.; Sutherland, B.G.; Telford, D.E.; Chhoker, S.S.; Sawyez, C.G.; Drangova, M.; Adams, A.C.; Kharitonenkov, A.; et al. Naringenin Prevents Obesity, Hepatic Steatosis, and Glucose Intolerance in Male Mice Independent of Fibroblast Growth Factor 21. Endocrinology 2015, 156, 2087–2102. [Google Scholar] [CrossRef] [PubMed]
- Zang, Y.; Zhang, L.; Igarashi, K.; Yu, C. The anti-obesity and anti-diabetic effects of kaempferol glycosides from unripe soybean leaves in high-fat-diet mice. Food Funct. 2015, 6, 834–841. [Google Scholar] [CrossRef]
- Bao, L.; Hu, L.; Zhang, Y.; Wang, Y. Hypolipidemic effects of flavonoids extracted from Lomatogonium rotatum. Exp. Ther. Med. 2016, 11, 1417–1424. [Google Scholar] [CrossRef]
- Pan, M.H.; Yang, G.; Li, S.; Li, M.Y.; Tsai, M.L.; Wu, J.C.; Badmaev, V.; Ho, C.T.; Lai, C.S. Combination of citrus polymethoxyflavones, green tea polyphenols, and Lychee extracts suppresses obesity and hepatic steatosis in high-fat diet induced obese mice. Mol. Nutr. Food Res. 2016, 61, 1601104. [Google Scholar] [CrossRef]
- Ke, J.Y.; Banh, T.; Hsiao, Y.H.; Cole, R.M.; Straka, S.R.; Yee, L.D.; Belury, M.A. Citrus flavonoid naringenin reduces mammary tumor cell viability, adipose mass, and adipose inflammation in obese ovariectomized mice. Mol. Nutr. Food Res. 2017, 61, 1600934. [Google Scholar] [CrossRef]
- Kwon, E.Y.; Lee, J.; Kim, Y.J.; Do, A.; Choi, J.Y.; Cho, S.J.; Jung, U.J.; Lee, M.K.; Park, Y.B.; Choi, M.S. Seabuckthorn leaves extract and flavonoid glycosides extract from seabuckthorn leaves ameliorates adiposity, hepatic steatosis, insulin resistance, and inflammation in diet-induced obesity. Nutrients 2017, 9, 569. [Google Scholar] [CrossRef] [PubMed]
- Burke, A.C.; Sutherland, B.G.; Telford, D.E.; Morrow, M.R.; Sawyez, C.G.; Edwards, J.Y.; Drangova, M.; Huff, M.W. Intervention with citrus flavonoids reverses obesity and improves metabolic syndrome and atherosclerosis in obese Ldlr/mice. J. Lipid Res. 2018, 59, 1714–1728. [Google Scholar] [CrossRef] [PubMed]
- Sheng, Y.; Liu, J.; Zheng, S.; Liang, F.; Luo, Y.; Huang, K.; Xu, W.; He, X. Mulberry leaves ameliorate obesity through enhancing brown adipose tissue activity and modulating gut microbiota. Food Funct. 2019, 10, 4771–4781. [Google Scholar] [CrossRef]
- Bian, Y.; Lei, J.; Zhong, J.; Wang, B.; Wan, Y.; Li, J.; Liao, C.; He, Y.; Liu, Z.; Ito, K.; et al. Kaempferol reduces obesity, prevents intestinal inflammation, and modulates gut microbiota in high-fat diet mice. J. Nutr. Biochem. 2021, 99, 108840. [Google Scholar] [CrossRef]
- Miranda, C.L.; Elias, V.D.; Hay, J.J.; Choi, J.; Reed, R.L.; Stevens, J.F. Xanthohumol improves dysfunctional glucose and lipid metabolism in diet-induced obese C57BL/6J mice. Arch. Biochem. Biophys. 2016, 599, 22–30. [Google Scholar] [CrossRef] [PubMed]
- Nascimento, O.V.; Boleti, A.P.A.; Yuyama, L.K.O.; Lima, E.S. Effects of diet supplementation with Camu-camu (Myrciaria dubiaHBK McVaugh) fruit in a rat model of diet-induced obesity. An. Acad. Bras. Ciências 2013, 85, 355–363. [Google Scholar] [CrossRef]
- Alam, M.A.; Kauter, K.; Brown, L. Naringin Improves Diet-Induced Cardiovascular Dysfunction and Obesity in High Carbohydrate, High Fat Diet-Fed Rats. Nutrients 2013, 5, 637–650. [Google Scholar] [CrossRef]
- You, J.S.; Lee, Y.J.; Kim, K.S.; Kim, S.H.; Chang, K.J. Anti-obesity and hypolipidaemic effects of Nelumbo nucifera seed ethanol extract in human pre-adipocytes and rats fed a high-fat diet. J. Sci. Food Agric. 2013, 94, 568–575. [Google Scholar] [CrossRef]
- Yoshida, H.; Watanabe, H.; Ishida, A.; Watanabe, W.; Narumi, K.; Atsumi, T.; Sugita, C.; Kurokawa, M. Naringenin suppresses macrophage infiltration into adipose tissue in an early phase of high-fat diet-induced obesity. Biochem. Biophys. Res. Commun. 2014, 454, 95–101. [Google Scholar] [CrossRef]
- Baselga-Escudero, L.; Pascual-Serrano, A.; Ribas-Latre, A.; Casanova, E.; Salvadó, M.J.; Arola, L.; Arola-Arnal, A.; Bladé, C. Long-term supplementation with a low dose of proanthocyanidins normalized liver miR-33a and miR-122 levels in high-fat diet–induced obese rats. Nutr. Res. 2015, 35, 337–345. [Google Scholar] [CrossRef] [PubMed]
- Miyata, S.; Inoue, J.; Shimizu, M.; Sato, R. Xanthohumol improves diet-induced obesity and fatty liver by suppressing Sterol Regulatory Element-binding Protein (SREBP) activation. J. Biol. Chem. 2015, 290, 20565–20579. [Google Scholar] [CrossRef] [PubMed]
- Qin, N.; Chen, Y.; Jin, M.N.; Zhang, C.; Qiao, W.; Yue, X.L.; Duan, H.Q.; Niu, W.Y. Anti-obesity and anti-diabetic effects of flavonoid derivative (Fla-CN) via microRNA in high fat diet induced obesity mice. Eur. J. Pharm. Sci. 2015, 82, 52–63. [Google Scholar] [CrossRef] [PubMed]
- Liu, C.; Ma, J.; Sun, J.; Cheng, C.; Feng, Z.; Jiang, H.; Yang, W. Flavonoid-Rich Extract of Paulownia fortunei Flowers Attenuates Diet-Induced Hyperlipidemia, Hepatic Steatosis and Insulin Resistance in Obesity Mice by AMPK Pathway. Nutrients 2017, 9, 959. [Google Scholar] [CrossRef]
- Gentile, D.; Fornai, M.; Colucci, R.; Pellegrini, C.; Tirotta, E.; Benvenuti, L.; Segnani, C.; Ippolito, C.; Duranti, E.; Virdis, A.; et al. The flavonoid compound apigenin prevents colonic inflammation and motor dysfunctions associated with high fat diet-induced obesity. PLoS ONE 2018, 13, e0195502. [Google Scholar] [CrossRef] [PubMed]
- Bai, Y.F.; Wang, S.W.; Wang, X.X.; Weng, Y.Y.; Fan, X.Y.; Sheng, H.; Zhu, X.T.; Lou, L.J.; Zhang, F. The flavonoid-rich Quzhou Fructus Aurantii extract modulates gut microbiota and prevents obesity in high-fat diet-fed mice. Nutr. Diabetes 2019, 9, 30. [Google Scholar] [CrossRef]
- Varshney, R.; Mishra, R.; Das, N.; Sircar, D.; Roy, P. A comparative analysis of various flavonoids in the regulation of obesity and diabetes: An in vitro and in vivo study. J. Funct. Foods 2019, 59, 194–205. [Google Scholar] [CrossRef]
- Romero-Juárez, P.A.; Visco, D.B.; Manhães-de-Castro, R.; Urquiza-Martínez, M.V.; Saavedra, L.M.; González-Vargas, M.C.; Mercado-Camargo, R.; Aquino, J.d.S.; Toscano, A.E.; Torner, L.; et al. Dietary flavonoid kaempferol reduces obesity-associated hypothalamic microglia activation and promotes body weight loss in mice with obesity. Nutr. Neurosci. 2021, 26, 25–39. [Google Scholar] [CrossRef]
- Yang, X.; Wang, Q.; Pang, Z.R.; Pan, M.R.; Zhang, W. Flavonoid-enriched extract from Hippophae rhamnoides seed reduces high fat diet induced obesity, hypertriglyceridemia, and hepatic triglyceride accumulation in C57BL/6 mice. Pharm. Biol. 2016, 55, 1207–1214. [Google Scholar] [CrossRef]
- Dai, J.; Liang, K.; Zhao, S.; Jia, W.; Liu, Y.; Wu, H.; Lv, J.; Cao, C.; Chen, T.; Zhuang, S.; et al. Chemoproteomics reveals baicalin activates hepatic CPT1 to ameliorate diet-induced obesity and hepatic steatosis. Biochemistry 2018, 115, E5896–E5905. [Google Scholar] [CrossRef]
- Shen, C.Y.; Wan, L.; Wang, T.X.; Jiang, J.G. Citrus aurantium L. var. amara Engl. inhibited lipid accumulation in 3T3-L1 cells and Caenorhabditis elegans and prevented obesity in high-fat diet-fed mice. Pharmacol. Res. 2019, 147, 104347. [Google Scholar] [CrossRef]
- Su, T.; Huang, C.; Yang, C.; Jiang, T.; Su, J.; Chen, M.; Fatima, S.; Gong, R.; Hu, X.; Bian, Z.; et al. Apigenin inhibits STAT3/CD36 signaling axis and reduces visceral obesity. Pharmacol. Res. 2019, 152, 104586. [Google Scholar] [CrossRef] [PubMed]
- Shahi, S.K.; Ghimire, S.; Lehman, P.; Mangalam, A.K. Obesity induced gut dysbiosis contributes to disease severity in an animal model of multiple sclerosis. Front. Immunol. 2022, 13, 4874. [Google Scholar] [CrossRef]
- Probiotics, N.; Breton, J.; Galmiche, M. Dysbiotic gut bacteria in obesity: An overview of the metabolic mechanisms and therapeutic perspectives of next-generation probiotics. Microorganisms 2022, 10, 452. [Google Scholar] [CrossRef]
- Mishra, S.P.; Wang, B.; Jain, S.; Ding, J.; Rejeski, J.; Furdui, C.M.; Kitzman, D.W.; Taraphder, S.; Brechot, C.; Kumar, A.; et al. A mechanism by which gut microbiota elevates permeability and inflammation in obese/diabetic mice and human gut. Gut Microbiota 2022. [Google Scholar] [CrossRef]
- Chen, L.; Chen, R.; Wang, H.; Liang, F. Mechanisms Linking Inflammation to Insulin Resistance. Int. J. Endocrinol. 2015, 2015, 508409. [Google Scholar] [CrossRef]
- Kolb, H.; Stumvoll, M.; Kramer, W.; Kempf, K.; Martin, S. Insulin translates unfavourable lifestyle into obesity. BMC Med. 2018, 16, 232. [Google Scholar] [CrossRef] [PubMed]
- Lee, H.C.; Jenner, A.M.; Low, C.S.; Lee, Y.K. Effect of tea phenolics and their aromatic fecal bacterial metabolites on intestinal microbiota. Res. Microbiol. 2006, 157, 876–884. [Google Scholar] [CrossRef]
- Collins, B.; Hoffman, J.; Martinez, K.; Grace, M.; Lila, M.A.; Cockrell, C.; Nadimpalli, A.; Chang, E.; Chuang, C.C.; Zhong, W.; et al. A polyphenol-rich fraction obtained from table grapes decreases adiposity, insulin resistance and markers of inflammation and impacts gut microbiota in high-fat-fed mice. J. Nutr. Biochem. 2015, 31, 150–165. [Google Scholar] [CrossRef]
- Zhao, L.; Zhang, Q.; Ma, W.; Tian, F.; Shen, H.; Zhou, M. A combination of quercetin and resveratrol reduces obesity in high-fat diet-fed rats by modulation of gut microbiota. Food Funct. 2017, 8, 4644–4656. [Google Scholar] [CrossRef]
- Yi, R.; Tan, F.; Zhou, X.; Mu, J.; Li, L.; Du, X. Effects of Lactobacillus fermentum CQPC04 on Lipid Reduction in C57BL / 6J Mice Activation of Bacteria. Front. Microbiol. 2020, 11, 573586. [Google Scholar] [CrossRef] [PubMed]
- Etxeberria, U.; Arias, N.; Boqué, N.; Macarulla, M.T.; Portillo, M.P.; Martínez, J.A.; Milagro, F.I. ScienceDirect Reshaping faecal gut microbiota composition by the intake of trans-resveratrol and quercetin in high-fat sucrose diet-fed rats. J. Nutr. Biochem. 2015, 26, 651–660. [Google Scholar] [CrossRef] [PubMed]
- Noratto, G.D.; Garcia-mazcorro, J.F.; Markel, M.; Martino, H.S.; Steiner, M.; Byrne, D.; Suchodolski, J.S.; Mertens-talcott, S.U.; Minamoto, Y. Carbohydrate-Free Peach (Prunus persica) and Plum (Prunus domestica) Juice Affects Fecal Microbial Ecology in an Obese Animal Model. PLoS ONE 2014, 9, e101723. [Google Scholar] [CrossRef] [PubMed]
- Nakatsu, C.H.; Armstrong, A.; Clavijo, A.P.; Martin, B.R.; Barnes, S.; Weaver, C.M. Fecal bacterial community changes associated with isoflavone metabolites in postmenopausal women after soy bar consumption. PLoS ONE 2014, 9, e108924. [Google Scholar] [CrossRef]
- Radulovic, K.; Normand, S.; Rehman, A.; Chatagnon, J.; Delacre, M.; Waldschmitt, N.; Poulin, L.F.; Iovanna, J.; Ryffel, B.; Rosenstiel, P.; et al. ARTICLES A dietary flavone confers communicable protection against colitis through NLRP6 signaling independently of inflammasome activation. Mucosal Immunol. 2017, 11, 811–819. [Google Scholar] [CrossRef]
- Neri-Numa, I.A.; Cazarin, C.B.B.; Ruiz, A.L.T.G.; Paulino, B.N.; Molina, G.; Pastore, G.M. Targeting flavonoids on modulation of metabolic syndrome. J. Funct. Foods 2020, 73, 104132. [Google Scholar] [CrossRef]
- Manna, P.; Jain, S.K. Obesity, Oxidative Stress, Adipose Tissue Dysfunction, and the Associated Health Risks: Causes and Therapeutic Strategies. Metab. Syndr. Relat. Disord. 2015, 13, 444. [Google Scholar] [CrossRef]
- Pérez-Torres, I.; Castrejón-Téllez, V.; Soto, M.E.; Rubio-Ruiz, M.E.; Manzano-Pech, L.; Guarner-Lans, V. Oxidative Stress, Plant Natural Antioxidants, and Obesity. Int. J. Mol. Sci. 2021, 22, 1786. [Google Scholar] [CrossRef]
- Marseglia, L.; Manti, S.; D’Angelo, G.; Nicotera, A.; Parisi, E.; Di Rosa, G.; Gitto, E.; Arrigo, T. Oxidative Stress in Obesity: A Critical Component in Human Diseases. Int. J. Mol. Sci. 2015, 16, 400. [Google Scholar] [CrossRef]
- Furukawa, S.; Fujita, T.; Shimabukuro, M.; Iwaki, M.; Yamada, Y.; Nakajima, Y.; Nakayama, O.; Makishima, M.; Matsuda, M.; Shimomura, I. Increased oxidative stress in obesity and its impact on metabolic syndrome. J. Clin. Investig. 2004, 114, 1752–1761. [Google Scholar] [CrossRef]
- Savini, I.; Catani, M.V.; Evangelista, D.; Gasperi, V.; Avigliano, L. Obesity-associated oxidative stress: Strategies finalized to improve redox state. Int. J. Mol. Sci. 2013, 14, 10497–10538. [Google Scholar] [CrossRef]
- Zhou, Y.; Li, H.; Xia, N. The Interplay Between Adipose Tissue and Vasculature: Role of Oxidative Stress in Obesity. Front. Cardiovasc. Med. 2021, 8, 131. [Google Scholar] [CrossRef] [PubMed]
- Macedo, R.C.; Bondan, E.F.; Otton, R. Redox status on different regions of the central nervous system of obese and lean rats treated with green tea extract. Nutr. Neurosci. 2017, 22, 119–131. [Google Scholar] [CrossRef] [PubMed]
- Gaballah, H.H.; El-Horany, H.E.; Helal, D.S. Mitigative effects of the bioactive flavonol fisetin on high-fat/high-sucrose induced nonalcoholic fatty liver disease in rats. J. Cell. Biochem. 2018, 120, 12762–12774. [Google Scholar] [CrossRef]
- Zeng, X.; Yang, J.; Hu, O.; Huang, J.; Ran, L.; Chen, M.; Zhang, Y.; Zhou, X.; Zhu, J.; Zhang, Q.; et al. Dihydromyricetin Ameliorates Nonalcoholic Fatty Liver Disease by Improving Mitochondrial Respiratory Capacity and Redox Homeostasis Through Modulation of SIRT3 Signaling. Antioxid. Redox Signal. 2017, 30, 7172. [Google Scholar] [CrossRef] [PubMed]
- Manach, C.; Williamson, G.; Morand, C.; Scalbert, A.; Rémésy, C. Bioavailability and bioefficacy of polyphenols in humans. I. Review of 97 bioavailability studies. Am. J. Clin. Nutr. 2005, 81, 230S–242S. [Google Scholar] [CrossRef]
- Akhlaghi, M.; Foshati, S. Bioavailability and Metabolism of Flavonoids: A Review. Int. J. Nutr. Sci. 2017, 2, 180–184. [Google Scholar]
- Naeem, A.; Ming, Y.; Pengyi, H.; Jie, K.Y.; Yali, L.; Haiyan, Z.; Shuai, X.; Wenjing, L.; Ling, W.; Xia, Z.M.; et al. The fate of flavonoids after oral administration: A comprehensive overview of its bioavailability. Crit. Rev. Food Sci. Nutr. 2021, 62, 6169–6186. [Google Scholar] [CrossRef]
- Kawabata, K.; Yoshioka, Y.; Terao, J. Role of Intestinal Microbiota in the Bioavailability and Physiological Functions of Dietary Polyphenols. Molecules 2019, 24, 370. [Google Scholar] [CrossRef]
- Al-Ishaq, R.K.; Liskova, A.; Kubatka, P.; Büsselberg, D. Enzymatic Metabolism of Flavonoids by Gut Microbiota and Its Impact on Gastrointestinal Cancer. Cancers 2021, 13, 3934. [Google Scholar] [CrossRef]
- Rahaman, S.T.; Mondal, S. Flavonoids: A vital resource in healthcare and medicine. Pharm. Pharmacol. Int. J. 2020, 8, 91–104. [Google Scholar] [CrossRef]
- Slámová, K.; Kapešová, J.; Valentová, K. “Sweet Flavonoids”: Glycosidase-Catalyzed Modifications. Int. J. Mol. Sci. 2018, 19, 2126. [Google Scholar] [CrossRef] [PubMed]
- Kumar, S.; Pandey, A.K. Chemistry and biological activities of flavonoids: An overview. Sci. World J. 2013, 2013, 162750. [Google Scholar] [CrossRef] [PubMed]



| Pharmacotherapy | |
|---|---|
| Types | Side Effects |
| Liraglutide (Saxenda®) [32] | Abdominal pain, constipation, vomiting, dyspepsia, fatigue, dizziness |
| Orlistat [33] | Steatorrhoea, bloating, oily spot, fecal urgency, fecal incontinence |
| Naltrexone/bupropion (Mysimba®) [34] | Increased risk of epileptic fits, nausea |
| Lorcaserin (Belviq®) [35] | Mitral regurgitation, depression, pulmonary hypertension |
| Phentermine/topiramate (Qsymia™) [36] | Paresthesia, dry mouth, dysgeusia, insomnia, dizziness, constipation |
| Oxyntomodulin [37] | Nausea, vomiting, diarrhea, acute pancreatitis, decreased blood pressure, increased risk of certain types of cancer |
| Beloranib [38] | Insomnia, decreased of mean systolic blood pressure |
| Surgical intervention | |
| Bariatric surgery [39] | Loss of lean body mass and muscle, micronutrient deficiency |
| Laparoscopic adjustable gastric banding [40] | Pouch enlargement, band slip, band erosion, intra-abdominal infection, port breakage |
| Roux-en-Y gastric bypass [41] | Stenosis at gastrojejunostomy, internal hernia, gallstones |
| Sleeve gastrectomy [42] | Midstomach stenosis, weight re-gain |
| Biliopancreatic diversion with a duodenal switch [43] | Bone pain, diarrhea, vomiting |
| Non-surgical intervention | |
| Intragastric balloon [44] | Vomiting, hypokalemia, renal insufficiency, abdominal pain, gastroesophageal reflux |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Naomi, R.; Teoh, S.H.; Halim, S.; Embong, H.; Hasain, Z.; Bahari, H.; Kumar, J. Unraveling Obesity: Transgenerational Inheritance, Treatment Side Effects, Flavonoids, Mechanisms, Microbiota, Redox Balance, and Bioavailability—A Narrative Review. Antioxidants 2023, 12, 1549. https://doi.org/10.3390/antiox12081549
Naomi R, Teoh SH, Halim S, Embong H, Hasain Z, Bahari H, Kumar J. Unraveling Obesity: Transgenerational Inheritance, Treatment Side Effects, Flavonoids, Mechanisms, Microbiota, Redox Balance, and Bioavailability—A Narrative Review. Antioxidants. 2023; 12(8):1549. https://doi.org/10.3390/antiox12081549
Chicago/Turabian StyleNaomi, Ruth, Soo Huat Teoh, Shariff Halim, Hashim Embong, Zubaidah Hasain, Hasnah Bahari, and Jaya Kumar. 2023. "Unraveling Obesity: Transgenerational Inheritance, Treatment Side Effects, Flavonoids, Mechanisms, Microbiota, Redox Balance, and Bioavailability—A Narrative Review" Antioxidants 12, no. 8: 1549. https://doi.org/10.3390/antiox12081549
APA StyleNaomi, R., Teoh, S. H., Halim, S., Embong, H., Hasain, Z., Bahari, H., & Kumar, J. (2023). Unraveling Obesity: Transgenerational Inheritance, Treatment Side Effects, Flavonoids, Mechanisms, Microbiota, Redox Balance, and Bioavailability—A Narrative Review. Antioxidants, 12(8), 1549. https://doi.org/10.3390/antiox12081549