
Effects of Prometryn Exposure on Hepatopancreas Oxidative Stress and Intestinal Flora in Eriocheir sinensis (Crustacea: Decapoda)

 ,
,
Abstract
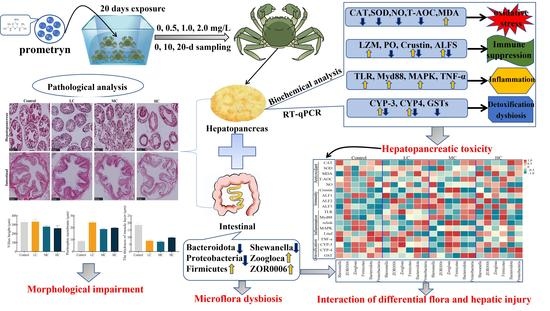
1. Introduction
2. Materials and Methods
2.1. Experimental Reagents and Animals
2.2. Experimental Design and Sample Collection
2.3. Biochemical Parameters Analysis
2.4. Histopathological Observation of Hepatopancreas and Intestinal
2.5. RT-qPCR Analysis
2.6. 16s Sequencing
2.7. Statistics and Analysis
3. Results
3.1. Effect of Prometryn Exposure on Hepatopancreas and Plasma Oxidative Stress Parameters
3.2. Effect of Prometryn Exposure on Hepatopancreas and Plasma Immune Parameters
3.3. Effect of Prometryn Exposure on Hepatopancreas Inflammatory Response and Detoxification Parameters
3.4. Effect of Prometryn on Morphological Impairment of the Hepatopancreas and Intestine
3.5. Effect of Prometryn on the Intestinal Microbiota
3.5.1. Community Structures of the Intestinal Microbiota
3.5.2. Analysis of Significant Differences in Intestinal Flora
3.5.3. Analysis of Significant Differences in the Phenotypic Abundance among the Intestinal Flora
3.6. Correlation Analysis of Parameters Related to Differential Intestinal Flora and Hepatopancreatic Injury
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Bottoni, P.; Grenni, P.; Lucentini, L.; Caracciolo, A.B. Terbuthylazine and other triazines in Italian water resources. Microchem. J. 2013, 107, 136–142. [Google Scholar] [CrossRef]
- Giardi, M.T.; Piletska, E.V.; Guzzella, L.; Pozzoni, F. The problem of herbicide water monitoring in Europe. In Biotechnological Applications of Photosynthetic Proteins: Biochips, Biosensors and Biodevices; Springer: Boston, MA, USA, 2006; pp. 116–129. [Google Scholar]
- Chong, R.S.-M. Pesticides toxicosis. Aquaculture Pathophysiology; Academic Press: Cambridge, MA, USA, 2022; pp. 783–797. [Google Scholar]
- Wang, R.; Zhang, S.; Cai, M.; Zhu, J.; Xu, Z.; Zheng, H.; Xiao, K.; Wang, F. Screening triazine herbicides in drinking water in the Yangtze River Delta, China: Occurrence and health risk. J. Hazard. Mater. Adv. 2023, 10, 100277. [Google Scholar] [CrossRef]
- Sule, R.; Gomes, A. Effects of prometryn on cardiac and liver cell lines and mouse tissues. FASEB J. 2022, 36. [Google Scholar] [CrossRef]
- Ru, S.; Wang, Y.; Zhang, X.; Yang, L. Toxic effects of triazine herbicides on aquatic animals and their degradation method. J. Ocean Univ. China Nat. Sci. Ed. 2022, 52, 1–12. [Google Scholar]
- Yang, Y.; Li, S.; Wang, Z.; Ren, Y.; Mu, Y.; Zhang, X.; Van den Brink, P.J.; Sun, H.; Song, Y.; Cheng, B. Acute toxicity, bioaccumulation and elimination of prometryn in tilapia (Oreochromis niloticus). Chemosphere 2022, 300, 134565. [Google Scholar] [CrossRef] [PubMed]
- Konstantinou, I.K.; Hela, D.G.; Albanis, T.A. The status of pesticide pollution in surface waters (rivers and lakes) of Greece. Part I. Review on occurrence and levels. Environ. Pollut. 2006, 141, 555–570. [Google Scholar] [CrossRef]
- Zhang, W.; Fan, G.; Meng, X.; Wang, T.; Chen, X.; Mang, Q. Bioaccumulation and elimination of prometryn in hard clam Meretrix meretrix. Anim. Husb. Feed Sci. 2018, 10, 231–235. [Google Scholar]
- Xie, H.; Chen, J.; Huang, Y.; Zhang, R.; Chen, C.-E.; Li, X.; Kadokami, K. Screening of 484 trace organic contaminants in coastal waters around the Liaodong Peninsula, China: Occurrence, distribution, and ecological risk. Environ. Pollut. 2020, 267, 115436. [Google Scholar] [CrossRef]
- Zhang, R.; Du, J.; Dong, X.; Huang, Y.; Xie, H.; Chen, J.; Li, X.; Kadokami, K. Occurrence and ecological risks of 156 pharmaceuticals and 296 pesticides in seawater from mariculture areas of Northeast China. Sci. Total Environ. 2021, 792, 148375. [Google Scholar] [CrossRef] [PubMed]
- Fu, X.; Liu, Q.; Xu, Y.; Meng, D.; Song, Y. Safety assessment of prometryn on human health and aquatic environment. Chin. Agric. Sci. Bull. 2015, 31, 49–57. [Google Scholar]
- Liu, Y.; Zhang, H.; Kong, C.; Gu, R.; Xi, Y.; Yang, G.; Zhang, K.; Shen, X. Pollution characteristics and risk assessment of pesticides and veterinary drugs in aquaculture environment and non-drugs fishery inputs in Shanghai, China. J. Agro-Environ. Sci. 2020, 41, 2055–2063. [Google Scholar]
- Fishery Administration Bureau of the Ministry of Agriculture and Villages; National Aquatic Products Technology Extension Station; China Society of Fisheries. China Fishery Statistical Yearbook; Chinese Agricultural Press: Beijing, China, 2022; p. 24.
- Barbee, G.C.; Stout, M.J. Comparative acute toxicity of neonicotinoid and pyrethroid insecticides to non-target crayfish (Procambarus clarkii) associated with rice-crayfish crop rotations. Pest Manag. Sci. 2009, 65, 1250–1256. [Google Scholar] [CrossRef] [PubMed]
- Hong, Y.; Yang, X.; Yan, G.; Huang, Y.; Zuo, F.; Shen, Y.; Ding, Y.; Cheng, Y. Effects of glyphosate on immune responses and haemocyte DNA damage of Chinese mitten crab, Eriocheir sinensis. Fish Shellfish Immunol. 2017, 71, 19–27. [Google Scholar] [CrossRef] [PubMed]
- Li, K. Study on the Toxic Effects of Atrazine and Chlorpyrifos on the Chinese Mitten Crab, Eriocheir sinensis. Ph.D. Thesis, East China Normal University, Shanghai, China, 2005. [Google Scholar]
- Wang, M.; Lv, J.; Deng, H.; Liu, Q.; Liang, S. Occurrence and removal of triazine herbicides during wastewater treatment processes and their environmental impact on aquatic life. Int. J. Environ. Res. Public Health 2022, 19, 4557. [Google Scholar] [CrossRef] [PubMed]
- Min, N.; Park, H.; Hong, T.; An, G.; Song, G.; Lim, W. Developmental toxicity of prometryn induces mitochondrial dysfunction, oxidative stress, and failure of organogenesis in zebrafish (Danio rerio). J. Hazard. Mater. 2023, 443, 130202. [Google Scholar] [CrossRef]
- Stara, A.; Kristan, J.; Zuskova, E.; Velisek, J. Effect of long-term exposure to prometryne on oxidative stress and antioxidant response in common carp (Cyprinus carpio L.). Pestic. Biochem. Physiol. 2013, 105, 18–23. [Google Scholar] [CrossRef]
- Zeng, F.; Wu, L.; Ren, X.; Xu, B.; Cui, S.; Li, M.; Chen, W.; Han, Y.; Ren, T. Effects of long-term prometryn exposure on antioxidative status, intestinal morphology, and microbiota in sea cucumber (Apostichopus japonicus). Comp. Biochem. Physiol. Part C Toxicol. Pharmacol. 2021, 250, 109187. [Google Scholar] [CrossRef]
- Stará, A.; Kouba, A.; Velíšek, J. Effect of long-term exposure to prometryne on oxidative stress and antioxidant response in red swamp crayfish (Procambarus clarkii). BioMed Res. Int. 2014, 2014, 680131. [Google Scholar] [CrossRef]
- Li, X.; Han, T.; Zheng, S.; Wu, G. Nutrition and functions of amino acids in aquatic crustaceans. Adv. Exp. Med. Biol. 2021, 1285, 169–198. [Google Scholar] [CrossRef]
- Li, M.; Ma, C.; Zhu, P.; Yang, Y.; Lei, A.; Chen, X.; Liang, W.; Chen, M.; Xiong, J.; Li, C. A new crustin is involved in the innate immune response of shrimp Litopenaeus vannamei. Fish Shellfish Immunol. 2019, 94, 398–406. [Google Scholar] [CrossRef]
- Li, C.; Zhao, J.; Song, L.; Mu, C.; Zhang, H.; Gai, Y.; Qiu, L.; Yu, Y.; Ni, D.; Xing, K. Molecular cloning, genomic organization and functional analysis of an anti-lipopolysaccharide factor from Chinese mitten crab Eriocheir sinensis. Dev. Comp. Immunol. 2008, 32, 784–794. [Google Scholar] [CrossRef] [PubMed]
- Fu, L.; Zhou, G.; Pan, J.; Li, Y.; Lu, Q.; Zhou, J.; Li, X. Effects of astragalus polysaccharides on antioxidant abilities and non-specific immune responses of Chinese mitten crab, Eriocheir sinensis. Aquacult. Int. 2017, 25, 1333–1343. [Google Scholar] [CrossRef]
- Liu, F.; Qu, Y.; Geng, C.; Wang, A.; Zhang, J.; Chen, K.; Liu, B.; Tian, H.; Yang, W.; Yu, Y. Effects of hesperidin on the growth performance, antioxidant capacity, immune responses and disease resistance of red swamp crayfish (Procambarus clarkii). Fish Shellfish Immunol. 2020, 99, 154–166. [Google Scholar] [CrossRef]
- Hong, Y.; Huang, Y.; Wu, S.; Yang, X.; Dong, Y.; Xu, D.; Huang, Z. Effects of imidacloprid on the oxidative stress, detoxification and gut microbiota of Chinese mitten crab, Eriocheir sinensis. Sci. Total Environ. 2020, 729, 138276. [Google Scholar] [CrossRef] [PubMed]
- Feng, W.; Su, S.; Song, C.; Yu, F.; Zhou, J.; Li, J.; Jia, R.; Xu, P.; Tang, Y. Effects of copper exposure on oxidative stress, apoptosis, endoplasmic reticulum stress, autophagy and immune response in different tissues of Chinese mitten crab (Eriocheir sinensis). Antioxidants 2022, 11, 2029. [Google Scholar] [CrossRef]
- Frova, C. Glutathione transferases in the genomics era: New insights and perspectives. Biomol. Eng. 2006, 23, 149–169. [Google Scholar] [CrossRef]
- Ringel, Y.; Maharshak, N. Intestinal microbiota and immune function in the pathogenesis of irritable bowel syndrome. Am. J. Physiol. 2013, 305, G529–G541. [Google Scholar] [CrossRef] [PubMed]
- Rescigno, M. Intestinal microbiota and its effects on the immune system. Cell. Microbiol. 2014, 16, 1004–1013. [Google Scholar] [CrossRef]
- Zeng, F. Effects of Long-Term Prometryn Exposure on Growth Performance, Antioxidative Status, and Morphology in Sea Cucumber (Apostichopus japonicus). Master’s Thesis, Dalian Ocean University, Dalian, China, 2022. [Google Scholar]
- Huang, P.; Du, J.; Cao, L.; Gao, J.; Li, Q.; Sun, Y.; Shao, N.; Zhang, Y.; Xu, G. Effects of prometryn on oxidative stress, immune response and apoptosis in the hepatopancreas of Eriocheir sinensis (Crustacea: Decapoda). Ecotoxicol. Environ. Saf. 2023, 262, 115159. [Google Scholar] [CrossRef]
- Livak, K.J.; Schmittgen, T.D. Analysis of relative gene expression data using real-time quantitative PCR and the 2−ΔΔCT method. Methods 2001, 25, 402–408. [Google Scholar] [CrossRef]
- Bolyen, E.; Rideout, J.R.; Dillon, M.R.; Bokulich, N.A.; Abnet, C.; Al-Ghalith, G.A.; Alexander, H.; Alm, E.J.; Arumugam, M.; Asnicar, F.; et al. Reproducible, interactive, scalable and extensible microbiome data science using QIIME 2. Nat. Biotechnol. 2018, 37, 852–857. [Google Scholar] [CrossRef] [PubMed]
- Ighodaro, O.M.; Akinloye, O.A. First line defence antioxidants-superoxide dismutase (SOD), catalase (CAT) and glutathione peroxidase (GPX): Their fundamental role in the entire antioxidant defence grid. Alex. J. Med. 2018, 54, 287–293. [Google Scholar] [CrossRef]
- Hong, Y.; Huang, Y.; Huang, Z. Oxidative stress, immunological response, and heat shock proteins induction in the Chinese mitten crab, Eriocheir sinensis following avermectin exposure. Environ. Toxicol. 2019, 35, 213–222. [Google Scholar] [CrossRef] [PubMed]
- Valenzuela, A. The biological significance of malondialdehyde determination in the assessment of tissue oxidative stress. Life Sci. 1991, 48, 301–309. [Google Scholar] [CrossRef]
- Wink, D.A.; Katrina, M.M.; Michael, G.E.; Ryzard, M.P.; Sandra, J.H.; Carol, C.; Michael, V.; Martin, F.; Mathew, B.G. Mechanisms of the antioxidant effects of nitric oxide. Antioxid. Redox Signal. 2004, 3, 203–213. [Google Scholar] [CrossRef]
- Zhu, S.; Long, X.; Turchini, G.M.; Deng, D.; Cheng, Y.; Wu, X. Towards defining optimal dietary protein levels for male and female sub-adult Chinese mitten crab, Eriocheir sinensis reared in earthen ponds: Performances, nutrient composition and metabolism, antioxidant capacity and immunity. Aquaculture 2021, 536, 736442. [Google Scholar] [CrossRef]
- Ma, R.; Zhou, G.; Feng, D.; Fang, W.; Chen, T.; Hu, K. Transcriptome analysis of Penaeus vannamei hepatopancreas reveals differences in toxicity mechanisms between phoxim and prometryne. Fish Shellfish Immunol. 2020, 105, 274–285. [Google Scholar] [CrossRef]
- Le Moullac, G.; Haffner, P. Environmental factors affecting immune responses in crustacea. Aquaculture 2000, 191, 121–131. [Google Scholar] [CrossRef]
- Liu, Z.; Yu, P.; Cai, M.; Wu, D.; Zhang, M.; Chen, M.; Zhao, Y. Effects of microplastics on the innate immunity and intestinal microflora of juvenile Eriocheir sinensis. Sci. Total Environ. 2019, 685, 836–846. [Google Scholar] [CrossRef]
- Zhou, X.; Xiao, C.; Zhu, F. The mechanism of semicarbazide (SEM) exposure causes immunosuppression and reduces disease resistance of marine crustacean species, Scylla paramamosain. Aquaculture 2022, 558, 738404. [Google Scholar] [CrossRef]
- Yue, F.; Pan, L.; Miao, J.; Zhang, L.; Li, J. Molecular cloning, characterization and mRNA expression of two antibacterial peptides: Crustin and anti-lipopolysaccharide factor in swimming crab Portunus trituberculatus. Comp. Biochem. Physiol. Part B Biochem. Mol. Biol. 2010, 156, 77–85. [Google Scholar] [CrossRef] [PubMed]
- Yang, Y.; Yu, Q.; Zhang, C.; Wang, X.; He, L.; Huang, Y.; Li, E.; Qin, J.; Chen, L. Acute thiamethoxam exposure induces hepatotoxicity and neurotoxicity in juvenile Chinese mitten crab (Eriocheir sinensis). Ecotoxicol. Environ. Saf. 2023, 249, 114399. [Google Scholar] [CrossRef] [PubMed]
- Yang, H.; Jiang, Y.; Lu, K.; Xiong, H.; Zhang, Y.; Wei, W. Herbicide atrazine exposure induce oxidative stress, immune dysfunction and WSSV proliferation in red swamp crayfish Procambarus clarkii. Chemosphere 2021, 283, 131227. [Google Scholar] [CrossRef]
- Takano, T.; Kondo, H.; Hirono, I.; Endo, M.; Saito-Taki, T.; Aoki, T. Toll-like receptors in teleosts. In Diseases in Asian Aquaculture VII; Asian Fisheries Society: Puchong, Malaysia, 2011; pp. 197–208. [Google Scholar]
- Liu, T.; Zhang, L.; Joo, D.; Sun, S.-C. NF-κB signaling in inflammation. Signal Transduct. Target. Ther. 2017, 2, 17023. [Google Scholar] [PubMed]
- Xu, P.; Derynck, R. Direct activation of TACE-mediated ectodomain shedding by p38 MAP kinase regulates EGF receptor-dependent cell proliferation. Mol. Cell 2010, 37, 551–566. [Google Scholar] [CrossRef]
- Quivy, V.; Van Lint, C. Regulation at multiple levels of NF-κB-mediated transactivation by protein acetylation. Biochem. Pharmacol. 2004, 68, 1221–1229. [Google Scholar] [CrossRef]
- Li, F.; Wang, L.; Zhang, H.; Zheng, P.; Zhao, J.; Qiu, L.; Zhang, Y.; Song, L. Molecular cloning and expression of a relish gene in Chinese mitten crab Eriocheir sinensis. Int. J. Immunogenet. 2010, 37, 499–508. [Google Scholar] [CrossRef]
- Zhang, C.; He, J.; Wang, X.; Su, R.; Huang, Q.; Qiao, F.; Qin, C.; Qin, J.; Chen, L. Dietary gamma-aminobutyric acid (GABA) improves non-specific immunity and alleviates lipopolysaccharide (lps)-induced immune overresponse in juvenile Chinese mitten crab (Eriocheir sinensis). Fish Shellfish Immunol. 2022, 124, 480–489. [Google Scholar] [CrossRef]
- Wang, X.; Yao, Q.; Zhang, D.; Lei, X.; Wang, S.; Wan, J.; Liu, H.; Chen, Y.; Zhao, Y.; Wang, G.; et al. Effects of acute salinity stress on osmoregulation, antioxidant capacity and physiological metabolism of female Chinese mitten crabs (Eriocheir sinensis). Aquaculture 2022, 552, 737989. [Google Scholar] [CrossRef]
- Bu, X.; Song, Y.; Pan, J.; Wang, X.; Qin, C.; Jia, Y.; Du, Z.; Qin, J.G.; Chen, L. Toxicity of long-term copper exposure on Chinese mitten crab (Eriocheir sinensis) and mitigation of its adverse impact by myo-inositol. Aquaculture 2022, 547, 737511. [Google Scholar] [CrossRef]
- Samanta, P.; Pal, S.; Mukherjee, A.K.; Ghosh, A.R. Biochemical effects of glyphosate based herbicide, excel mera 71 on enzyme activities of acetylcholinesterase (Ache), lipid peroxidation (LPO), catalase (CAT), glutathione-s-transferase (GST) and protein content on teleostean fishes. Ecotoxicol. Environ. Saf. 2014, 107, 120–125. [Google Scholar] [CrossRef] [PubMed]
- Patil, A. Thiamethoxam induced histopathological alterations in hepatopancreas of freshwater bivalve Parreysia cylindrica. J. Entomol. Zool. Stud. 2019, 7, 1236–1240. [Google Scholar]
- Zhao, Q.; Shi, F.; Zhu, L. Prometryn and humic acid induce cytochrome P450 1A expression in Danio rerio (zebrafish). Ecotoxicol. Environ. Saf. 2017, 135, 40–47. [Google Scholar] [CrossRef]
- Lee, B.-Y.; Choi, B.-S.; Kim, M.-S.; Park, J.C.; Jeong, C.-B.; Han, J.; Lee, J.-S. The genome of the freshwater water flea Daphnia magna: A potential use for freshwater molecular ecotoxicology. Aquat. Toxicol. 2019, 210, 69–84. [Google Scholar] [CrossRef]
- Wang, C.; Wang, X.; Xiao, S.; Bu, X.; Lin, Z.; Qi, C.; Qin, J.G.; Chen, L. T-2 toxin in the diet suppresses growth and induces immunotoxicity in juvenile Chinese mitten crab (Eriocheir sinensis). Fish Shellfish Immunol. 2020, 97, 593–601. [Google Scholar] [CrossRef]
- Jajte, J.; Grzegorczyk, J.; Zmyślony, M.; Rajkowska, E. Effect of 7 mT static magnetic field and iron ions on rat lymphocytes: Apoptosis, necrosis and free radical processes. Bioelectrochemistry 2002, 57, 107–111. [Google Scholar] [CrossRef] [PubMed]
- Wang, Q.; Xu, Z.; Wang, Y.; Huo, G.; Zhang, X.; Li, J.; Hua, C.; Li, S.; Zhou, F. Transcriptomics analysis of the toxicological impact of enrofloxacin in an aquatic environment on the Chinese mitten crab (Eriocheir sinensis). Int. J. Environ. Res. Public Health 2023, 20, 1836. [Google Scholar] [CrossRef]
- Yang, E.; Zhang, J.; Yang, L.; Amenyogbe, E.; Wang, W.; Huang, J.; Chen, G. Effects of hypoxia stress on digestive enzyme activities, intestinal structure and the expression of tight junction proteins coding genes in juvenile cobia (Rachycentron canadum). Aquacult. Res. 2021, 52, 5630–5641. [Google Scholar] [CrossRef]
- Guo, X.; Qian, Z.; Pan, Q.; Hu, Y.; Mei, W.; Xing, X.; Yin, S.; Ji, J.; Zhang, K. Effects of florfenicol on intestinal histology, apoptosis and gut microbiota of Chinese mitten crab (Eriocheir sinensis). Int. J. Mol. Sci. 2023, 24, 4412. [Google Scholar] [CrossRef]
- Wang, C.; Wang, X.; Huang, Y.; Bu, X.; Xiao, S.; Qin, C.; Qiao, F.; Qin, J.G.; Chen, L. Effects of dietary T-2 toxin on gut health and gut microbiota composition of the juvenile Chinese mitten crab (Eriocheir sinensis). Fish Shellfish Immunol. 2020, 106, 574–582. [Google Scholar] [CrossRef]
- Han, F.; Qi, C.; Lin, Z.; Jia, Y.; Yang, L.; Lu, J.; Wang, X.; Li, E.; Qin, J.G.; Chen, L. Improvement of dietary N-acetylcysteine on growth inhibition and intestinal damage induced by β-conglycinin in juvenile Chinese mitten crabs (Eriocheir sinensis). Aquaculture 2020, 514, 734504. [Google Scholar] [CrossRef]
- Cai, C.; Wu, P.; Ye, Y.; Song, L.; Hooft, J.; Yang, C.; Kong, L.; Chen, Q.; Wang, Y. Assessment of the feasibility of including high levels of oilseed meals in the diets of juvenile Chinese mitten crabs (Eriocheir sinensis): Effects on growth, non-specific immunity, hepatopancreatic function, and intestinal morphology. Anim. Feed Sci. Technol. 2014, 196, 117–127. [Google Scholar] [CrossRef]
- Huang, Y.; Hong, Y.; Yin, H.; Yan, G.; Huang, Q.; Li, Z.; Huang, Z. Imidacloprid induces locomotion impairment of the freshwater crayfish, Procambarus clarkii via neurotoxicity and oxidative stress in digestive system. Aquat. Toxicol. 2021, 238, 105913. [Google Scholar] [CrossRef]
- Mo, B.; Li, J.; Liao, G.; Wang, L.; Fan, L. Toxic effects of glyphosate on histopathology and intestinal microflora of juvenile Litopenaeus vannamei. Aquat. Toxicol. 2023, 255, 106399. [Google Scholar] [CrossRef] [PubMed]
- Yan, Q.; Li, J.; Yu, Y.; Wang, J.; He, Z.; Van Nostrand, J.D.; Kempher, M.L.; Wu, L.; Wang, Y.; Liao, L.; et al. Environmental filtering decreases with fish development for the assembly of gut microbiota. Environ. Microbiol. 2016, 18, 4739–4754. [Google Scholar] [CrossRef] [PubMed]
- Sullam, K.E.; Rubin, B.E.; Dalton, C.M.; Kilham, S.S.; Flecker, A.S.; Russell, J.A. Divergence across diet, time and populations rules out parallel evolution in the gut microbiomes of Trinidadian guppies. ISME J. 2015, 9, 1508–1522. [Google Scholar] [CrossRef] [PubMed]
- Wang, C.; Chen, Z.; Brennan, M.A.; Wang, J.; Sun, J.; Fang, H.; Kang, M.; Brennan, C.S.; Mu, J. The effect of extruded multigrain powder on metabolism and intestinal flora of high-fat-diet induced C57BL/6J mice. Food Res. Int. 2023, 169, 112878. [Google Scholar] [CrossRef]
- Khan, I.; Huang, Z.; Liang, L.; Li, N.; Ali, Z.; Ding, L.; Hong, M.; Shi, H. Ammonia stress influences intestinal histomorphology, immune status and microbiota of Chinese striped-neck turtle (Mauremys sinensis). Ecotoxicol. Environ. Saf. 2021, 222, 112471. [Google Scholar] [CrossRef] [PubMed]
- Chen, Y.; Chiu, C.; Hung, S.; Huang, W.; Lee, Y.; Liu, J.; Huang, Y.; Chen, T.; Chuang, H. Gnotobiotic mice inoculated with Firmicutes, but not Bacteroidetes, deteriorate nonalcoholic fatty liver disease severity by modulating hepatic lipid metabolism. Nutr. Res. 2019, 69, 20–29. [Google Scholar] [CrossRef]
- Zhan, M.; Xi, C.; Gong, J.; Zhu, M.; Shui, Y.; Xu, Z.; Xu, G.; Shen, H. 16S rRNA gene sequencing analysis reveals an imbalance in the intestinal flora of Eriocheir sinensis with hepatopancreatic necrosis disease. Comp. Biochem. Physiol. Part D Genom. Proteom. 2022, 42, 100988. [Google Scholar] [CrossRef]
- Cottrell, M.T.; Kirchman, D.L. Natural assemblages of marine proteobacteria and members of the Cytophaga-Flavobacter cluster consuming low- and high-molecular-weight dissolved organic matter. Appl. Environ. Microbiol. 2000, 66, 1692–1697. [Google Scholar] [CrossRef] [PubMed]
- Xie, Y.; Liu, X.; Ni, S.; Wei, H.; Chen, X.; Ismail, S.; Ahmad, S.; Lee, T. Insight into Impacts of Sewage Discharge and River Self-Purification on Microbial Dynamics and Pathogenicity in River Ecosystem; Europe PMC: London, UK, 2021. [Google Scholar]
- Gallet, A.; Halary, S.; Duval, C.; Huet, H.; Duperron, S.; Marie, B. Disruption of fish gut microbiota composition and holobiont’s metabolome during a simulated Microcystis aeruginosa (Cyanobacteria) bloom. Microbiome 2023, 11, 108. [Google Scholar] [CrossRef] [PubMed]
- Hua, Y.; Huang, W.; Wang, F.; Jing, Z.; Li, J.; Zhao, Y. Metabolites, gene expression and gut microbiota profiles suggest the putative mechanisms via which dietary creatine increases the serum taurine and g-ABA contents in Megalobrama amblycephala. Fish Physiol. Biochem. 2023, 49, 253–274. [Google Scholar] [CrossRef] [PubMed]
- Angthong, P.; Uengwetwanit, T.; Arayamethakorn, S.; Chaitongsakul, P.; Karoonuthaisiri, N.; Rungrassamee, W. Bacterial analysis in the early developmental stages of the black tiger shrimp (Penaeus monodon). Sci. Rep. 2020, 10, 4896. [Google Scholar] [CrossRef]
- Cámara-Ruiz, M.; Balebona, M.C.; Moriñigo, M.Á.; Esteban, M.Á. Probiotic shewanella putrefaciens (SpPdp11) as a fish health modulator: A review. Microorganisms 2020, 8, 1990. [Google Scholar] [CrossRef] [PubMed]
- Zhang, X.; Sun, J.; Han, Z.; Chen, F.; Lv, A.; Hu, X.; Sun, X.; Qi, H.; Guo, Y. Vibrio parahaemolyticus alters the community composition and function of intestinal microbiota in pacific white shrimp, Penaeus vannamei. Aquaculture 2021, 544, 737061. [Google Scholar] [CrossRef]










{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Taxonname | Mean Abundance | p-Value | |
|---|---|---|---|
| Bacteroidota | 0.27749 (control) | 0.14669 (HC) | 0.016 |
| Firmicutes | 0.27330 (control) | 0.43423 (HC) | 0.022 |
| Proteobacteria | 0.43711 (control) | 0.33298 (HC) | 0.049 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Huang, P.; Cao, L.; Du, J.; Gao, J.; Zhang, Y.; Sun, Y.; Li, Q.; Nie, Z.; Xu, G. Effects of Prometryn Exposure on Hepatopancreas Oxidative Stress and Intestinal Flora in Eriocheir sinensis (Crustacea: Decapoda). Antioxidants 2023, 12, 1548. https://doi.org/10.3390/antiox12081548
Huang P, Cao L, Du J, Gao J, Zhang Y, Sun Y, Li Q, Nie Z, Xu G. Effects of Prometryn Exposure on Hepatopancreas Oxidative Stress and Intestinal Flora in Eriocheir sinensis (Crustacea: Decapoda). Antioxidants. 2023; 12(8):1548. https://doi.org/10.3390/antiox12081548
Chicago/Turabian StyleHuang, Peng, Liping Cao, Jinliang Du, Jiancao Gao, Yuning Zhang, Yi Sun, Quanjie Li, Zhijuan Nie, and Gangchun Xu. 2023. "Effects of Prometryn Exposure on Hepatopancreas Oxidative Stress and Intestinal Flora in Eriocheir sinensis (Crustacea: Decapoda)" Antioxidants 12, no. 8: 1548. https://doi.org/10.3390/antiox12081548
APA StyleHuang, P., Cao, L., Du, J., Gao, J., Zhang, Y., Sun, Y., Li, Q., Nie, Z., & Xu, G. (2023). Effects of Prometryn Exposure on Hepatopancreas Oxidative Stress and Intestinal Flora in Eriocheir sinensis (Crustacea: Decapoda). Antioxidants, 12(8), 1548. https://doi.org/10.3390/antiox12081548