

The Impact of Resveratrol-Enriched Bread on Cardiac Remodeling in a Preclinical Model of Diabetes

 , , ,
, , ,  ,
,  , ,
, ,  ,
,  and
and 
Abstract
1. Introduction
2. Materials and Methods
2.1. Ethical Statement
2.2. Rodents Food Preparation
2.3. Animal Housing and Chemicals
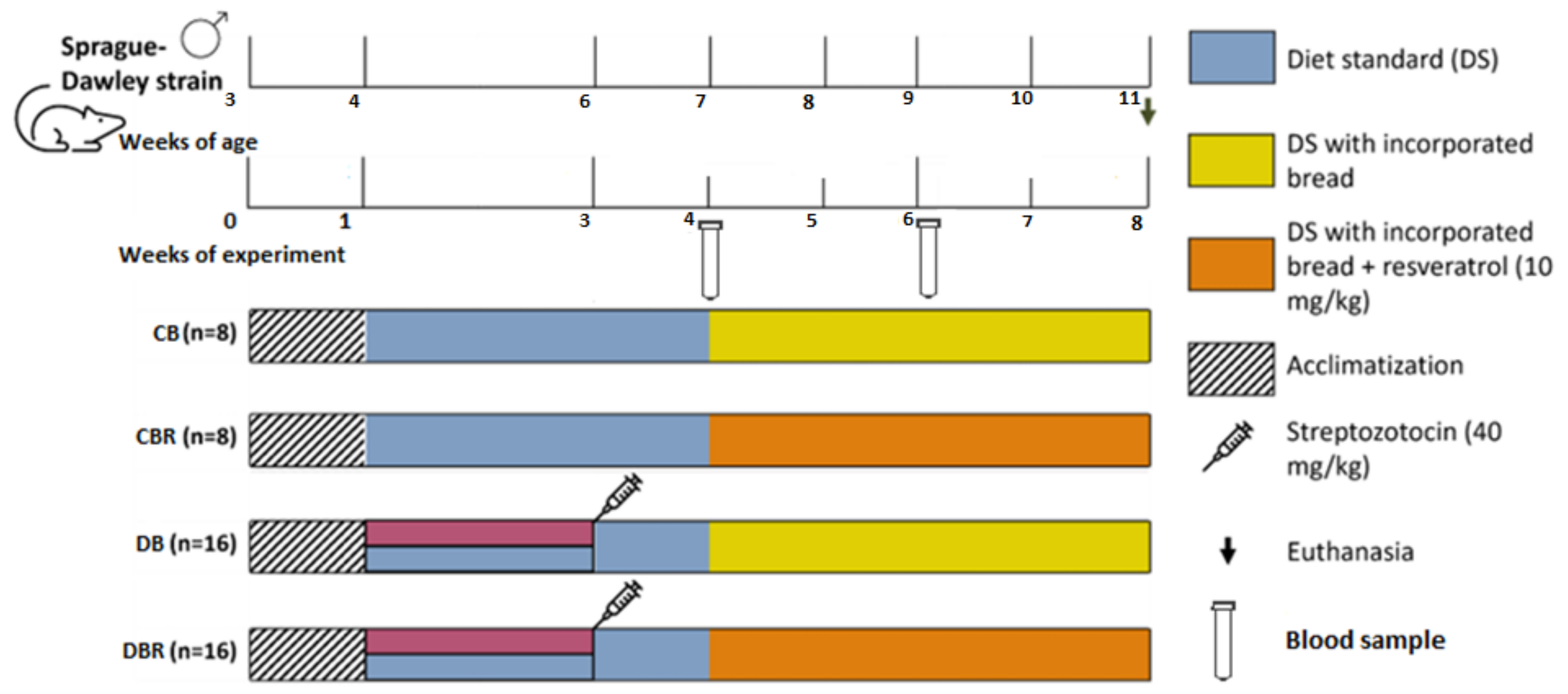
2.4. Experimental Design
2.5. Echocardiographic Study
2.6. Biochemical Analysis in Serum Samples
2.7. Histological Analysis
2.8. Cardiac Muscle Preparation for Biochemical Analysis
2.9. Immunoblotting
2.10. Citrate Synthase Activity
2.11. Statistical Analysis
3. Results
3.1. Effect of Functional Bread with RSV and Fructose Feed/STZ Injection on Anthropometric and Welfare Parameters
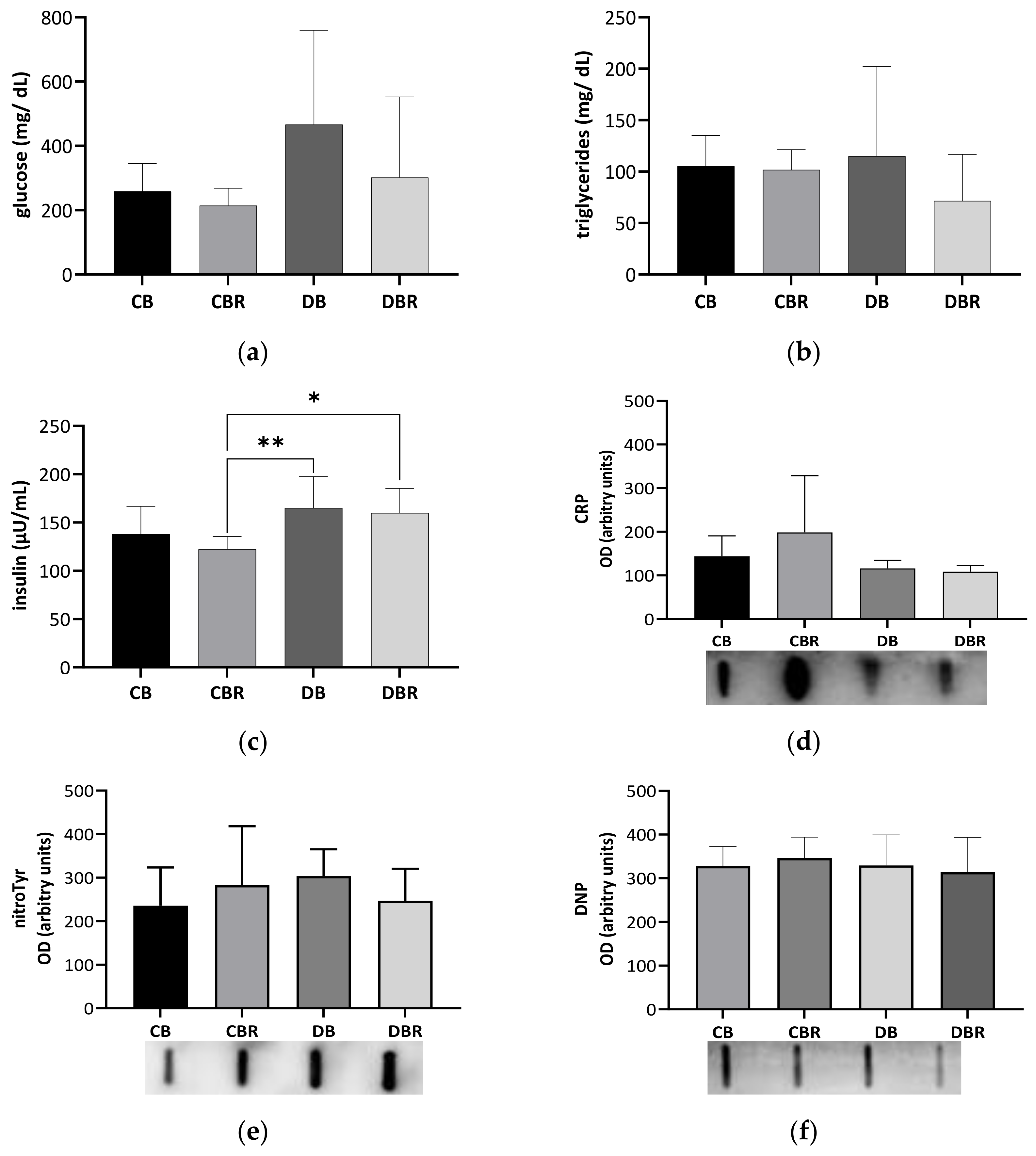
3.2. Systemic Adaptations to Fructose-Feeding plus Streptozotocin Administration and Functional Bread with RSV
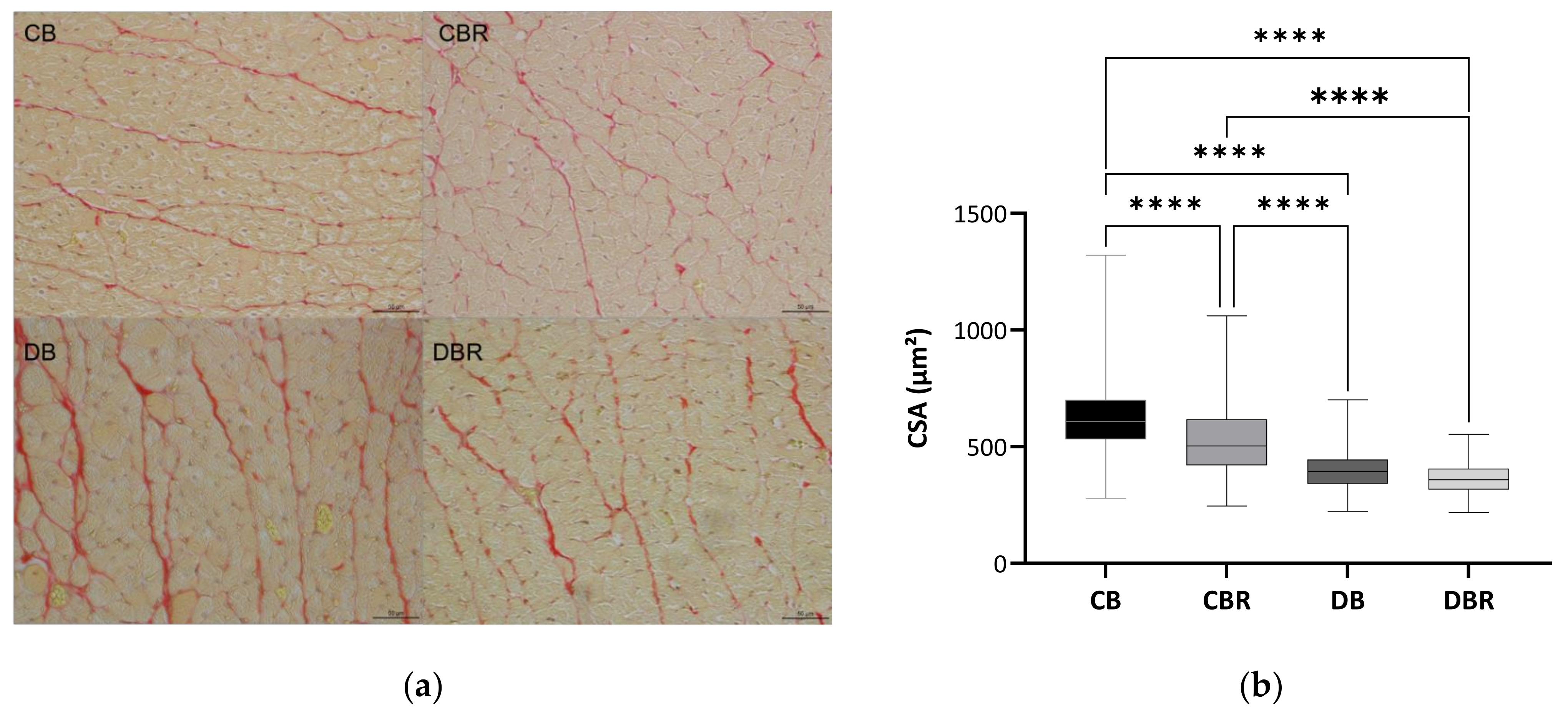
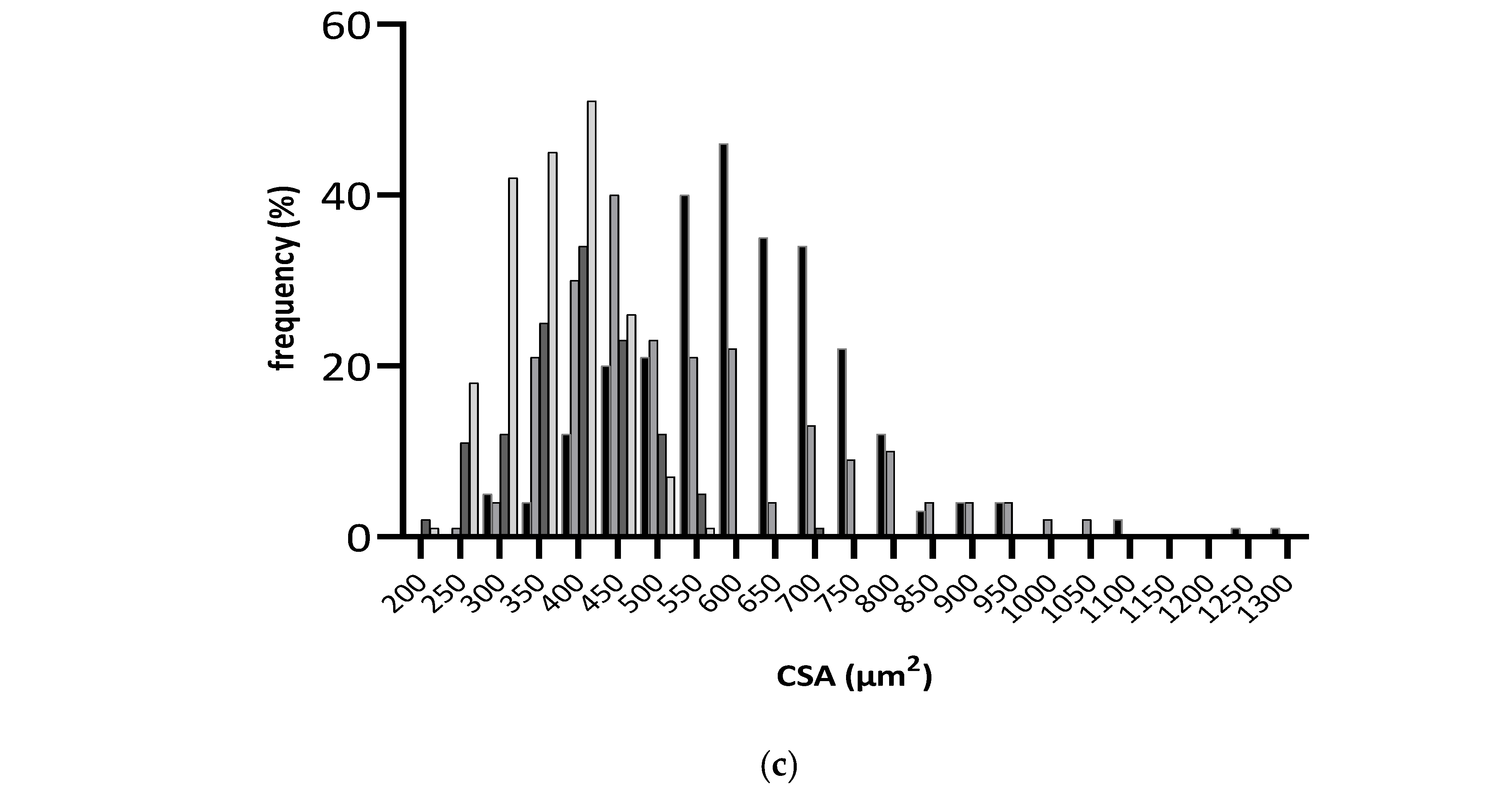
3.3. The Influence of Fructose-Feeding plus Streptozotocin Administration and Functional Bread with RSV on Cardiac Function and Morphometry
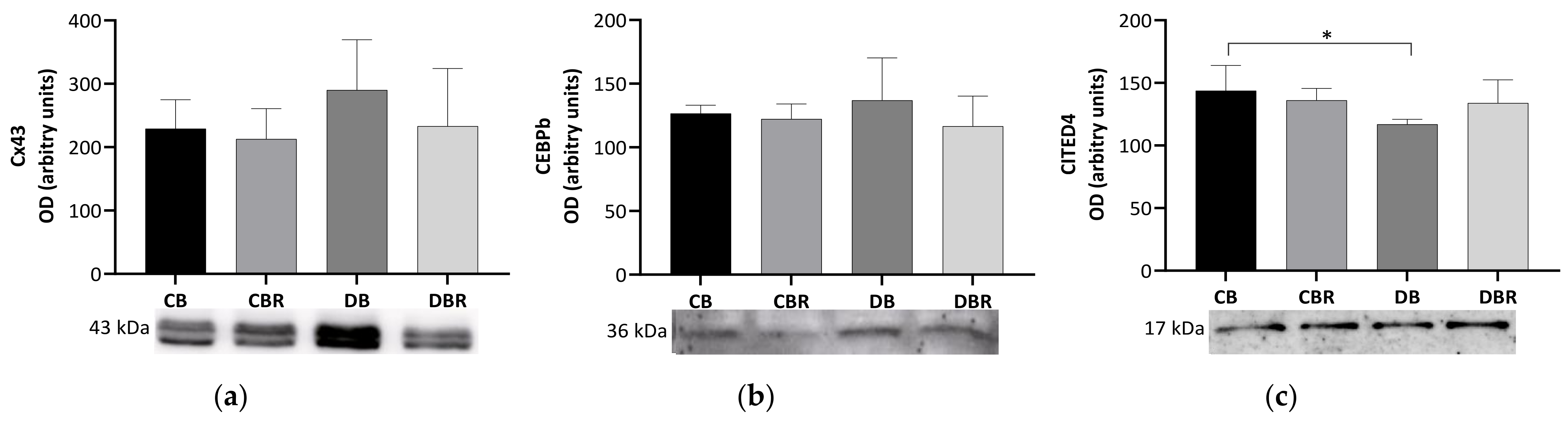
3.4. The Effect of Fructose-Feeding plus Streptozotocin Administration and Functional Bread with RSV on Molecular Markers of Cardiac Functionality and Regeneration
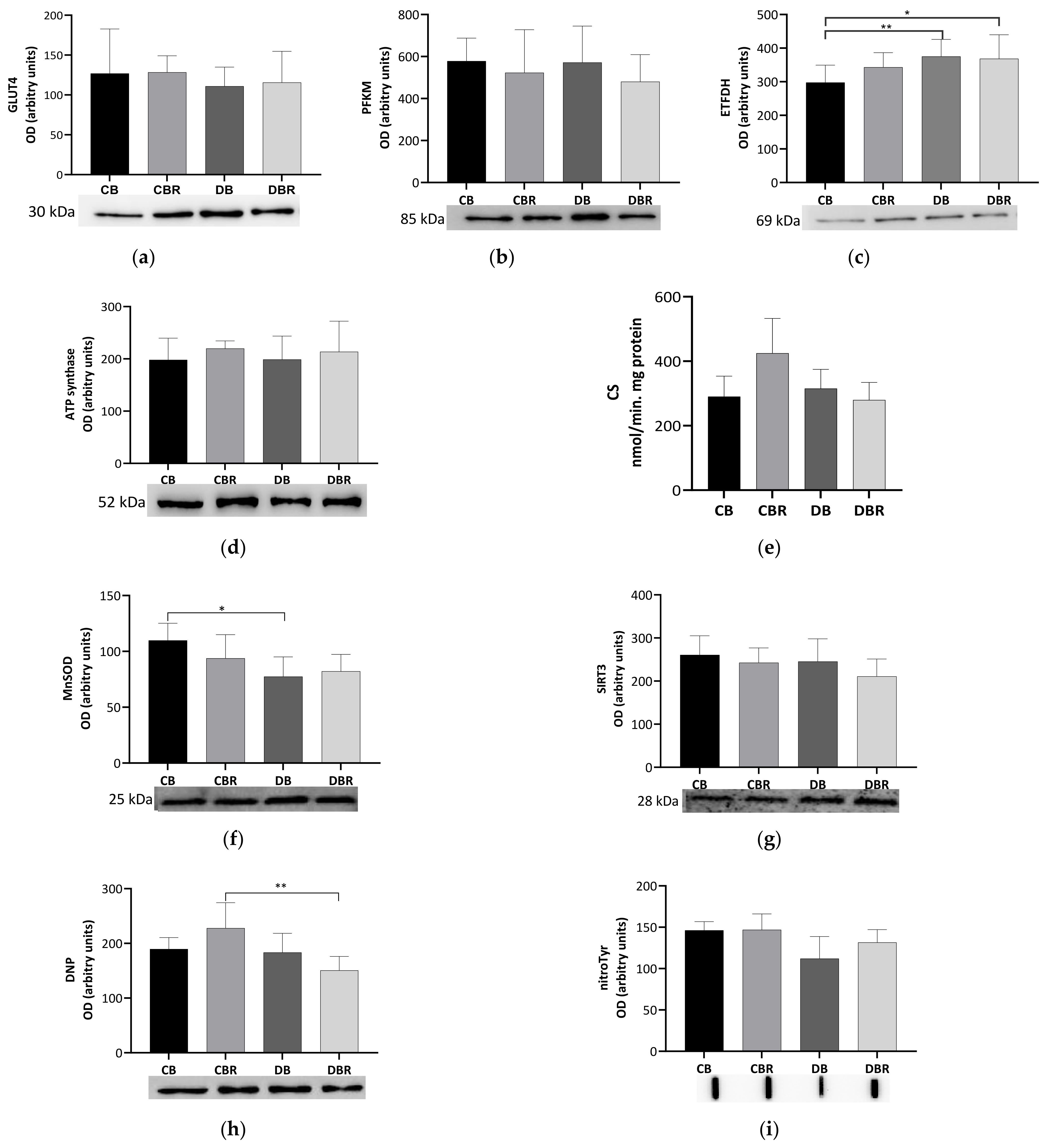
3.5. The Influence of Fructose-Feeding plus Streptozotocin Administration and Functional Bread with RSV on Cardiac Metabolism and Oxidative Stress
4. Discussion
4.1. Effect of Resveratrol-Rich Diet on Anthropometric and Systemic Parameters
4.2. Effect of Resveratrol-Rich Diet on Heart Remodeling
4.3. Limitations
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- World Health Organization. Cardiovascular Diseases. Available online: https://www.who.int/en/news-room/fact-sheets/detail/cardiovascular-diseases-(cvds) (accessed on 19 March 2022).
- Cannon, C.P. Cardiovascular Disease and Modifiable Cardiometabolic Risk Factors. Clin. Cornerstone 2007, 8, 11–28. [Google Scholar] [CrossRef] [PubMed]
- Francula-Zaninovic, S.; Nola, I.A. Management of Measurable Variable Cardiovascular Disease’ Risk Factors. Curr. Cardiol. Rev. 2018, 14, 153–163. [Google Scholar] [CrossRef] [PubMed]
- International Diabetes Federation. Diabetes Now Affects One in 10 Adults Worldwide. Available online: https://www.idf.org/news/240:diabetes-now-affects-one-in-10-adults-worldwide.html (accessed on 19 March 2022).
- Boudina, S.; Abel, E.D. Diabetic Cardiomyopathy, Causes and Effects. Rev. Endocr. Metab. Disord. 2010, 11, 31–39. [Google Scholar] [CrossRef] [PubMed]
- Mozaffarian, D. Dietary and Policy Priorities for Cardiovascular Disease, Diabetes, and Obesity. Circulation 2016, 133, 187–225. [Google Scholar] [CrossRef] [PubMed]
- Cardoso, S.M.; Pereira, O.R.; Seca, A.M.L.; Pinto, D.C.G.A.; Silva, A.M.S. Seaweeds as Preventive Agents for Cardiovascular Diseases: From Nutrients to Functional Foods. Mar. Drugs 2015, 13, 6838–6865. [Google Scholar] [CrossRef]
- Camargo-Ramos, C.M.; Correa-Bautista, J.E.; Correa-Rodríguez, M.; Ramírez-Vélez, R. Dietary Inflammatory Index and Cardiometabolic Risk Parameters in Overweight and Sedentary Subjects. Int. J. Environ. Res. Public Health 2017, 14, 1104. [Google Scholar] [CrossRef]
- Langcake, P.; Pryce, R.J. The Production of Resveratrol by Vitis Vinifera and Other Members of the Vitaceae as a Response to Infection or Injury. Physiol. Plant Pathol. 1976, 9, 77–86. [Google Scholar] [CrossRef]
- Burns, J.; Yokota, T.; Ashihara, H.; Lean, M.E.J.; Crozier, A. Plant Foods and Herbal Sources of Resveratrol. J. Agric. Food Chem. 2002, 50, 3337–3340. [Google Scholar] [CrossRef]
- European Commission. COMMISSION IMPLEMENTING REGULATION (EU) 2021/51 of 22 January 2021 Authorising a Change of the Conditions of Use of the Novel Food ‘Trans-Resveratrol’ under Regulation (EU) 2015/2283 of the European Parliament and of the Council and Amending Commission Im. Off. J. Eur. Union 2018, 2016, 48–119. [Google Scholar]
- Borriello, A.; Cucciolla, V.; Della Ragione, F.; Galletti, P. Dietary Polyphenols: Focus on Resveratrol, a Promising Agent in the Prevention of Cardiovascular Diseases and Control of Glucose Homeostasis. Nutr. Metab. Cardiovasc. Dis. 2010, 20, 618–625. [Google Scholar] [CrossRef]
- Szkudelski, T.; Szkudelska, K. Anti-Diabetic Effects of Resveratrol. Ann. N. Y. Acad. Sci. 2011, 1215, 34–39. [Google Scholar] [CrossRef]
- Sulaiman, M.; Matta, M.J.; Sunderesan, N.R.; Gupta, M.P.; Periasamy, M.; Gupta, M. Resveratrol, an Activator of SIRT1, Upregulates Sarcoplasmic Calcium ATPase and Improves Cardiac Function in Diabetic Cardiomyopathy. Am. J. Physiol. Circ. Physiol. 2010, 298, H833–H843. [Google Scholar] [CrossRef]
- Szkudelska, K.; Deniziak, M.; Hertig, I.; Wojciechowicz, T.; Tyczewska, M.; Jaroszewska, M.; Szkudelski, T. Effects of Resveratrol in Goto-Kakizaki Rat, a Model of Type 2 Diabetes. Nutrients 2019, 11, 2488. [Google Scholar] [CrossRef]
- Huang, J.-P.; Huang, S.-S.; Deng, J.-Y.; Chang, C.-C.; Day, Y.-J.; Hung, L.-M. Insulin and Resveratrol Act Synergistically, Preventing Cardiac Dysfunction in Diabetes, but the Advantage of Resveratrol in Diabetics with Acute Heart Attack Is Antagonized by Insulin. Free Radic. Biol. Med. 2010, 49, 1710–1721. [Google Scholar] [CrossRef]
- Crandall, J.P.; Oram, V.; Trandafirescu, G.; Reid, M.; Kishore, P.; Hawkins, M.; Cohen, H.W.; Barzilai, N. Pilot Study of Resveratrol in Older Adults With Impaired Glucose Tolerance. J. Gerontol. Ser. A 2012, 67, 1307–1312. [Google Scholar] [CrossRef]
- Cai, H.; Scott, E.; Kholghi, A.; Andreadi, C.; Rufini, A.; Karmokar, A.; Britton, R.G.; Horner-Glister, E.; Greaves, P.; Jawad, D.; et al. Cancer Chemoprevention: Evidence of a Nonlinear Dose Response for the Protective Effects of Resveratrol in Humans and Mice. Sci. Transl. Med. 2015, 7, 350le2. [Google Scholar] [CrossRef]
- Berman, A.Y.; Motechin, R.A.; Wiesenfeld, M.Y.; Holz, M.K. The Therapeutic Potential of Resveratrol: A Review of Clinical Trials. NPJ Precis. Oncol. 2017, 1, 35. [Google Scholar] [CrossRef]
- Hasan, M.M.; Yun, H.-K.; Kwak, E.-J.; Baek, K.-H. Preparation of Resveratrol-Enriched Grape Juice from Ultrasonication Treated Grape Fruits. Ultrason. Sonochem. 2014, 21, 729–734. [Google Scholar] [CrossRef]
- Gaudette, N.J.; Pickering, G.J. Sensory and Chemical Characteristics of Trans-Resveratrol-Fortified Wine. Aust. J. Grape Wine Res. 2011, 17, 249–257. [Google Scholar] [CrossRef]
- Silva, A.F.R.; Monteiro, M.; Resende, D.; Braga, S.S.; Coimbra, M.A.; Silva, A.M.S.; Cardoso, S.M. Inclusion Complex of Resveratrol with γ-Cyclodextrin as a Functional Ingredient for Lemon Juices. Foods 2020, 10, 16. [Google Scholar] [CrossRef]
- Silva, A.F.R.; Monteiro, M.; Nunes, R.; Baião, A.; Braga, S.S.; Sarmento, B.; Coimbra, M.A.; Silva, A.M.S.; Cardoso, S.M. Bread Enriched with Resveratrol: Influence of the Delivery Vehicles on Its Bioactivity. Food Biosci. 2022, 49, 101887. [Google Scholar] [CrossRef]
- Masood, S.; Rehman, A.U.; Bashir, S.; El Shazly, M.; Imran, M.; Khalil, P.; Ifthikar, F.; Jaffar, H.M.; Khursheed, T. Investigation of the Anti-Hyperglycemic and Antioxidant Effects of Wheat Bread Supplemented with Onion Peel Extract and Onion Powder in Diabetic Rats. J. Diabetes Metab. Disord. 2021, 20, 485–495. [Google Scholar] [CrossRef] [PubMed]
- Weng, Y.; Zhang, M.; Wang, J.; Zhang, Y. Significantly Hypoglycemic Effect of a Novel Functional Bread Rich in Mulberry Bark and Improving the Related Functions of Liver, Pancreas, and Kidney, on T2D Mice. Food Sci. Nutr. 2021, 9, 2468–2482. [Google Scholar] [CrossRef] [PubMed]
- Tan, Y.; Zhang, Z.; Zheng, C.; Wintergerst, K.A.; Keller, B.B.; Cai, L. Mechanisms of Diabetic Cardiomyopathy and Potential Therapeutic Strategies: Preclinical and Clinical Evidence. Nat. Rev. Cardiol. 2020, 17, 585–607. [Google Scholar] [CrossRef] [PubMed]
- Wilson, R.D.; Islam, M.S. Fructose-Fed Streptozotocin-Injected Rat: An Alternative Model for Type 2 Diabetes. Pharmacol. Rep. 2012, 64, 129–139. [Google Scholar] [CrossRef]
- Adoga, J.O.; Channa, M.L.; Nadar, A. Kolaviron Attenuates Cardiovascular Injury in Fructose-Streptozotocin Induced Type-2 Diabetic Male Rats by Reducing Oxidative Stress, Inflammation, and Improving Cardiovascular Risk Markers. Biomed. Pharmacother. 2021, 144, 112323. [Google Scholar] [CrossRef]
- Castelhano, J.; Ribeiro, B.; Sanches, M.; Graça, B.; Saraiva, J.; Oliveiros, B.; Neves, C.; Rodrigues, T.; Sereno, J.; Gonçalves, S.; et al. A Rat Model of Enhanced Glycation Mimics Cardiac Phenotypic Components of Human Type 2 Diabetes: A Translational Study Using MRI. J. Diabetes Complicat. 2020, 34, 107554. [Google Scholar] [CrossRef]
- Silva-Reis, R.; Faustino-Rocha, A.I.; Silva, J.; Valada, A.; Azevedo, T.; Anjos, L.; Gonçalves, L.; Pinto, M.d.L.; Ferreira, R.; Silva, A.M.S.; et al. Studying and Analyzing Humane Endpoints in the Fructose-Fed and Streptozotocin-Injected Rat Model of Diabetes. Animals 2023, 13, 1397. [Google Scholar] [CrossRef]
- Olfert, E.; Bhasin, J.; Latt, R.; Macallum, E.; McCutcheon, K.; Rainnie, D.; Schunk, M. CCAC Guidelines on: Choosing an Appropriate Endpoint in Experiments Using Animals for Research, Teaching and Testing; Canadian Council on Animal Care: Ottawa, ON, Canada, 1998; pp. 1–33. [Google Scholar]
- Cacho, J.; Sevillano, J.; De Castro, J.; Herrera, E.; Ramos, M.P. Validation of Simple Indexes to Assess Insulin Sensitivity during Pregnancy in Wistar and Sprague-Dawley Rats. Am. J. Physiol.-Endocrinol. Metab. 2008, 295, 1269–1276. [Google Scholar] [CrossRef]
- Brandão, S.R.; Reis-Mendes, A.; Domingues, P.; Duarte, J.A.; Bastos, M.L.; Carvalho, F.; Ferreira, R.; Costa, V.M. Exploring the Aging Effect of the Anticancer Drugs Doxorubicin and Mitoxantrone on Cardiac Mitochondrial Proteome Using a Murine Model. Toxicology 2021, 459, 152852. [Google Scholar] [CrossRef]
- Padrão, A.I.; Carvalho, T.; Vitorino, R.; Alves, R.M.P.; Caseiro, A.; Duarte, J.A.; Ferreira, R.; Amado, F. Impaired Protein Quality Control System Underlies Mitochondrial Dysfunction in Skeletal Muscle of Streptozotocin-Induced Diabetic Rats. Biochim. Biophys. Acta-Mol. Basis Dis. 2012, 1822, 1189–1197. [Google Scholar] [CrossRef]
- Silva, M.G.; Nunes, P.; Oliveira, P.; Ferreira, R.; Fardilha, M.; Moreira-Gonçalves, D.; Duarte, J.A.; Oliveira, M.M.; Peixoto, F. Long-Term Aerobic Training Improves Mitochondrial and Antioxidant Function in the Liver of Wistar Rats Preventing Hepatic Age-Related Function Decline. Biology 2022, 11, 1750. [Google Scholar] [CrossRef]
- Fajardo, R.J.; Karim, L.; Calley, V.I.; Bouxsein, M.L. A Review of Rodent Models of Type 2 Diabetic Skeletal Fragility. J. Bone Miner. Res. 2014, 29, 1025–1040. [Google Scholar] [CrossRef]
- Barrière, D.A.; Noll, C.; Roussy, G.; Lizotte, F.; Kessai, A.; Kirby, K.; Belleville, K.; Beaudet, N.; Longpré, J.M.; Carpentier, A.C.; et al. Combination of High-Fat/High-Fructose Diet and Low-Dose Streptozotocin to Model Long-Term Type-2 Diabetes Complications. Sci. Rep. 2018, 8, 424. [Google Scholar] [CrossRef]
- He, Q.; Su, G.; Liu, K.; Zhang, F.; Jiang, Y.; Gao, J.; Liu, L.; Jiang, Z.; Jin, M.; Xie, H. Sex-Specific Reference Intervals of Hematologic and Biochemical Analytes in Sprague-Dawley Rats Using the Nonparametric Rank Percentile Method. PLoS ONE 2017, 12, e0189837. [Google Scholar] [CrossRef]
- Maintra, A. The Endocrine System. In Robbins & Cotran–Pathologic Basis of Disease; Elsevier: Amsterdam, The Netherlands, 2015; pp. 1105–1109. ISBN 9781455726134. [Google Scholar]
- Jeyaraman, M.M.; Srisakuldee, W.; Nickel, B.E.; Kardami, E. Connexin43 Phosphorylation and Cytoprotection in the Heart. Biochim. Biophys. Acta-Biomembr. 2012, 1818, 2009–2013. [Google Scholar] [CrossRef]
- Boström, P.; Mann, N.; Wu, J.; Quintero, P.A.; Plovie, E.R.; Panáková, D.; Gupta, R.K.; Xiao, C.; MacRae, C.A.; Rosenzweig, A.; et al. C/EBPβ Controls Exercise-Induced Cardiac Growth and Protects against Pathological Cardiac Remodeling. Cell 2010, 143, 1072–1083. [Google Scholar] [CrossRef]
- Huang, G.N.; Thatcher, J.E.; McAnally, J.; Kong, Y.; Qi, X.; Tan, W.; DiMaio, J.M.; Amatruda, J.F.; Gerard, R.D.; Hill, J.A.; et al. C/EBP Transcription Factors Mediate Epicardial Activation During Heart Development and Injury. Science 2012, 338, 1599–1603. [Google Scholar] [CrossRef]
- Ding, S.; Gan, T.; Song, M.; Dai, Q.; Huang, H. C/EBPB-CITED4 in Exercised Heart. In Exercise for Cardiovascular Disease Prevention and Treatment; Xiao, J., Ed.; Advances in Experimental Medicine and Biology; Springer: Singapore, 2017; Volume 1000, pp. 247–259. ISBN 978-981-10-4303-1. [Google Scholar]
- Aerni-Flessner, L.; Abi-Jaoude, M.; Koenig, A.; Payne, M.; Hruz, P.W. GLUT4, GLUT1, and GLUT8 Are the Dominant GLUT Transcripts Expressed in the Murine Left Ventricle. Cardiovasc. Diabetol. 2012, 11, 63. [Google Scholar] [CrossRef]
- Houten, S.M.; Wanders, R.J.A. A General Introduction to the Biochemistry of Mitochondrial Fatty Acid Β-oxidation. J. Inherit. Metab. Dis. 2010, 33, 469–477. [Google Scholar] [CrossRef]
- Sun, W.; Liu, C.; Chen, Q.; Liu, N.; Yan, Y.; Liu, B. SIRT3: A New Regulator of Cardiovascular Diseases. Oxid. Med. Cell. Longev. 2018, 2018, 1–11. [Google Scholar] [CrossRef] [PubMed]
- Castro, L.; Demicheli, V.; Tórtora, V.; Radi, R. Mitochondrial Protein Tyrosine Nitration. Free Radic. Res. 2011, 45, 37–52. [Google Scholar] [CrossRef]
- Chakarova, N.; Dimova, R.; Grozeva, G.; Tankova, T. Assessment of Glucose Variability in Subjects with Prediabetes. Diabetes Res. Clin. Pract. 2019, 151, 56–64. [Google Scholar] [CrossRef]
- Palsamy, P.; Subramanian, S. Resveratrol, a Natural Phytoalexin, Normalizes Hyperglycemia in Streptozotocin-Nicotinamide Induced Experimental Diabetic Rats. Biomed. Pharmacother. 2008, 62, 598–605. [Google Scholar] [CrossRef] [PubMed]
- Baynest, H.W. Classification, Pathophysiology, Diagnosis and Management of Diabetes Mellitus. J. Diabetes Metab. 2015, 6, 1–9. [Google Scholar] [CrossRef]
- Zhang, F.; Ye, C.; Li, G.; Ding, W.; Zhou, W.; Zhu, H.; Chen, G.; Luo, T.; Guang, M.; Liu, Y.; et al. The Rat Model of Type 2 Diabetic Mellitus and Its Glycometabolism Characters. Exp. Anim. 2003, 52, 401–407. [Google Scholar] [CrossRef]
- Palsamy, P.; Subramanian, S. Ameliorative Potential of Resveratrol on Proinflammatory Cytokines, Hyperglycemia Mediated Oxidative Stress, and Pancreatic β-Cell Dysfunction in Streptozotocin-Nicotinamide-Induced Diabetic Rats. J. Cell. Physiol. 2010, 224, 423–432. [Google Scholar] [CrossRef]
- Burgeiro, A.; Cerqueira, M.; Varela-Rodríguez, B.; Nunes, S.; Neto, P.; Pereira, F.; Reis, F.; Carvalho, E. Glucose and Lipid Dysmetabolism in a Rat Model of Prediabetes Induced by a High-Sucrose Diet. Nutrients 2017, 9, 638. [Google Scholar] [CrossRef]
- Han, L.; Bittner, S.; Dong, D.; Cortez, Y.; Bittner, A.; Chan, J.; Umar, M.; Shen, W.-J.; Peterson, R.G.; Kraemer, F.B.; et al. Molecular Changes in Hepatic Metabolism in ZDSD Rats–A New Polygenic Rodent Model of Obesity, Metabolic Syndrome, and Diabetes. Biochim. Biophys. Acta-Mol. Basis Dis. 2020, 1866, 165688. [Google Scholar] [CrossRef]
- Seedevi, P.; Ramu Ganesan, A.; Moovendhan, M.; Mohan, K.; Sivasankar, P.; Loganathan, S.; Vairamani, S.; Shanmugam, A. Anti-Diabetic Activity of Crude Polysaccharide and Rhamnose-Enriched Polysaccharide from G. Lithophila on Streptozotocin (STZ)-Induced in Wistar Rats. Sci. Rep. 2020, 10, 556. [Google Scholar] [CrossRef]
- Mohan, M.; Dihoum, A.; Mordi, I.R.; Choy, A.-M.; Rena, G.; Lang, C.C. Left Ventricular Hypertrophy in Diabetic Cardiomyopathy: A Target for Intervention. Front. Cardiovasc. Med. 2021, 8, 746382. [Google Scholar] [CrossRef]
- Rospleszcz, S.; Schafnitzel, A.; Koenig, W.; Lorbeer, R.; Auweter, S.; Huth, C.; Rathmann, W.; Heier, M.; Linkohr, B.; Meisinger, C.; et al. Association of Glycemic Status and Segmental Left Ventricular Wall Thickness in Subjects without Prior Cardiovascular Disease: A Cross-Sectional Study. BMC Cardiovasc. Disord. 2018, 18, 162. [Google Scholar] [CrossRef]
- Eguchi, K.; Boden-Albala, B.; Jin, Z.; Rundek, T.; Sacco, R.L.; Homma, S.; Di Tullio, M.R. Association Between Diabetes Mellitus and Left Ventricular Hypertrophy in a Multiethnic Population. Am. J. Cardiol. 2008, 101, 1787–1791. [Google Scholar] [CrossRef]
- Brands, M.W.; Fitzgerald, S.M.; Hewitt, W.H.; Hailman, A.E. Decreased Cardiac Output at the Onset of Diabetes: Renal Mechanisms and Peripheral Vasoconstriction. Am. J. Physiol. Endocrinol. Metab. 2000, 278, E917-24. [Google Scholar] [CrossRef]
- Chen, X.-F.; Wang, J.-A.; Lin, X.-F.; Tang, L.-J.; Yu, W.-F.; Chen, H.; Xie, X.-J.; Jiang, J.-J.; Peng, X.-H. Diabetes Mellitus: Is It Protective against Aortic Root Dilatation? Cardiology 2009, 112, 138–143. [Google Scholar] [CrossRef]
- Ehl, N.F.; Kühne, M.; Brinkert, M.; Müller-Brand, J.; Zellweger, M.J. Diabetes Reduces Left Ventricular Ejection Fraction--Irrespective of Presence and Extent of Coronary Artery Disease. Eur. J. Endocrinol. 2011, 165, 945–951. [Google Scholar] [CrossRef]
- Akula, A. Biochemical, Histological and Echocardiographic Changes during Experimental Cardiomyopathy in STZ-Induced Diabetic Rats. Pharmacol. Res. 2003, 48, 429–435. [Google Scholar] [CrossRef]
- Gordan, R.; Gwathmey, J.K.; Xie, L. Autonomic and Endocrine Control of Cardiovascular Function. World J. Cardiol. 2015, 7, 204. [Google Scholar] [CrossRef]
- Hensel, K.O. Non-Ischemic Diabetic Cardiomyopathy May Initially Exhibit a Transient Subclinical Phase of Hyperdynamic Myocardial Performance. Med. Hypotheses 2016, 94, 7–10. [Google Scholar] [CrossRef]
- Patil, M.B.; Burji, N.P.A. Echocardiographic Evaluation of Diastolic Dysfunction in Asymptomatic Type 2 Diabetes Mellitus. J. Assoc. Physicians India 2012, 60, 23–26. [Google Scholar]
- Jia, G.; Whaley-Connell, A.; Sowers, J.R. Diabetic Cardiomyopathy: A Hyperglycaemia- and Insulin-Resistance-Induced Heart Disease. Diabetologia 2018, 61, 21–28. [Google Scholar] [CrossRef] [PubMed]
- Diao, J.; Wei, J.; Yan, R.; Fan, G.; Lin, L.; Chen, M. Effects of Resveratrol on Regulation on UCP2 and Cardiac Function in Diabetic Rats. J. Physiol. Biochem. 2019, 75, 39–51. [Google Scholar] [CrossRef] [PubMed]
- Kanamori, H.; Takemura, G.; Goto, K.; Tsujimoto, A.; Mikami, A.; Ogino, A.; Watanabe, T.; Morishita, K.; Okada, H.; Kawasaki, M.; et al. Autophagic Adaptations in Diabetic Cardiomyopathy Differ between Type 1 and Type 2 Diabetes. Autophagy 2015, 11, 1146–1160. [Google Scholar] [CrossRef]
- Wu, H.; Li, G.N.; Xie, J.; Li, R.; Chen, Q.H.; Chen, J.Z.; Wei, Z.H.; Kang, L.N.; Xu, B. Resveratrol Ameliorates Myocardial Fibrosis by Inhibiting ROS/ERK/TGF-β/Periostin Pathway in STZ-Induced Diabetic Mice. BMC Cardiovasc. Disord. 2016, 16, 5. [Google Scholar] [CrossRef] [PubMed]
- Sakamoto, T.; Kelly, D.P. A Case for Adaptive Cardiac Hypertrophic Remodeling Is CITED. Circ. Res. 2020, 127, 647–650. [Google Scholar] [CrossRef]
- Radosinka, J.; Kurahara, L.H.; Hirashi, K.; Viczencozva, C.; Egan, T.B.; Szeiffova, B.B.; Dosenko, V.; Navarova, J.; Obsitnik, B.; Imanaga, I.; et al. Modulation of Cardiac Connexin-43 by Omega-3 Fatty Acid Ethyl-Ester Supplementation Demonstrated in Spontaneously Diabetic Rats. Physiol. Res. 2015, 64, 795–806. [Google Scholar] [CrossRef]
- Joshi, M.S.; Mihm, M.J.; Cook, A.C.; Schanbacher, B.L.; Bauer, J.A. Alterations in Connexin 43 during Diabetic Cardiomyopathy: Competition of Tyrosine Nitration versus Phosphorylation. J. Diabetes 2015, 7, 250–259. [Google Scholar] [CrossRef]
- Karwi, Q.G.; Uddin, G.M.; Ho, K.L.; Lopaschuk, G.D. Loss of Metabolic Flexibility in the Failing Heart. Front. Cardiovasc. Med. 2018, 5, 1–19. [Google Scholar] [CrossRef]
- Mazumder, P.K.; O’Neill, B.T.; Roberts, M.W.; Buchanan, J.; Yun, U.J.; Cooksey, R.C.; Boudina, S.; Abel, E.D. Impaired Cardiac Efficiency and Increased Fatty Acid Oxidation in Insulin-Resistant Ob/Ob Mouse Hearts. Diabetes 2004, 53, 2366–2374. [Google Scholar] [CrossRef]
- Stanley, W.C.; Chandler, M.P. Energy Metabolism in the Normal and Failing Heart: Potential for Therapeutic Interventions. Heart Fail. Rev. 2002, 7, 115–130. [Google Scholar] [CrossRef]
- Verma, S.K.; Garikipati, V.N.S.; Kishore, R. Mitochondrial Dysfunction and Its Impact on Diabetic Heart. Biochim. Biophys. Acta–Mol. Basis Dis. 2017, 1863, 1098–1105. [Google Scholar] [CrossRef]
- Fadini, G.P.; Miorin, M.; Facco, M.; Bonamico, S.; Baesso, I.; Grego, F.; Menegolo, M.; de Kreutzenberg, S.V.; Tiengo, A.; Agostini, C.; et al. Circulating Endothelial Progenitor Cells Are Reduced in Peripheral Vascular Complications of Type 2 Diabetes Mellitus. J. Am. Coll. Cardiol. 2005, 45, 1449–1457. [Google Scholar] [CrossRef]
- Bagul, P.; Katare, P.; Bugga, P.; Dinda, A.; Banerjee, S.K. SIRT-3 Modulation by Resveratrol Improves Mitochondrial Oxidative Phosphorylation in Diabetic Heart through Deacetylation of TFAM. Cells 2018, 7, 235. [Google Scholar] [CrossRef]
- Chen, T.; Li, J.; Liu, J.; Li, N.; Wang, S.; Liu, H.; Zeng, M.; Zhang, Y.; Bu, P. Activation of SIRT3 by Resveratrol Ameliorates Cardiac Fibrosis and Improves Cardiac Function via the TGF-β/Smad3 Pathway. Am. J. Physiol. Circ. Physiol. 2015, 308, H424–H434. [Google Scholar] [CrossRef]
- Gumede, N.; Ngubane, P.; Khathi, A. Assessing the Risk Factors for Myocardial Infarction in Diet-Induced Prediabetes: Myocardial Tissue Changes. BMC Cardiovasc. Disord. 2022, 22, 350. [Google Scholar] [CrossRef]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Experimental Groups | ||||
|---|---|---|---|---|
| CB | CBR | DB | DBR | |
| BW (g) | 325.4 ± 41.6 ab | 343.3 ± 16.0 c | 241.9 ± 49.1 ac | 263.1 ± 69.2 b |
| HW (g) | 1.16 ± 0.04 | 1.28 ± 0.11 ab | 0.97 ± 0.17 a | 1.06 ± 0.23 b |
| Gastrocnemius (g) | 4.60 ± 0.26 ab | 4.58 ± 0.14 cd | 2.77 ± 0.91 ac | 3.22 ± 1.16 bd |
| Tibia (cm) | 4.15 ± 0.13 | 4.25 ± 0.17 a | 4.02 ± 0.19 | 3.99 ± 0.25 a |
| BW/tibia (g/cm) | 78.4 ± 9.9 a | 80.8 ± 3.8 bc | 60.0 ± 11.3 ab | 64.1 ± 14.2 ac |
| HW/BW (mg/g) | 3.50 ± 0.20 ab | 3.72 ± 0.28 | 4.05 ± 0.43 a | 4.09 ± 0.45 b |
| HW/tibia (mg/cm) | 0.284 ± 0.021 a | 0.301 ± 0.028 bc | 0.225 ± 0.024 ab | 0.254 ± 0.040 c |
| Welfare score | 0.13 ± 0.35 | 0.13 ± 0.35 | 1.13 ± 1.13 | 0.50 ± 0.82 |
| Mortality (%) | 0 | 0 | 6.25% | 0 |
| Groups of Treatment | ||||
|---|---|---|---|---|
| CB | CBR | DB | DBR | |
| IVS d (mm) | 1.12 ± 0.18 a | 1.21 ± 0.20 b | 0.968 ± 0.172 b | 1.40 ± 0.26 ab |
| LVPW d (mm) | 1.11 ± 0.18 a | 1.29 ± 0.29 b | 0.994 ± 0.159 bc | 1.51 ± 0.34 ac |
| IVS s (mm) | 1.17 ± 0.23 a | 1.17 ± 0.18 b | 0.956 ± 0.165 c | 1.47 ± 0.24 abc |
| LVPW s (mm) | 1.21 ± 0.19 a | 1.21 ± 0.24 b | 0.925 ± 0.212 ab | 1.65 ± 0.38 ab |
| LV mass (mm) | 0.498 ± 0.104 a | 0.604 ± 0.130 | 0.475 ± 0.121 b | 0.723 ± 0.250 ab |
| LV ET (s) | 82.9 ± 8.37 ab | 93.9 ± 18.3 c | 125.8 ± 18.5 ac | 108.9 ± 20.0 b |
| Ao d (cm) | 0.283 ± 0.042 | 0.299 ± 0.029 | 0.307 ± 0.027 | 0.283 ± 0.046 |
| Ao VTI (cm) | 3.94 ± 0.86 a | 5.23 ± 1.20 ab | 4.30 ± 0.82 | 3.81 ± 1.21 b |
| HR (bpm) | 321.3 ± 25.8 ab | 272.5 ± 37.9 c | 190.0 ± 36.7 ac | 217.5 ± 67.3 b |
| E (cm/s) | 0.683 ± 0.117 | 0.812 ± 0.107 ab | 0.600 ± 0.190 a | 0.575 ± 0.168 b |
| A (cm/s) | 0.427 ± 0.097 a | 0.380 ± 0.149 | 0.385 ± 0.119 | 0.308 ± 0.067 a |
| E/A | 1.64 ± 0.29 a | 2.47 ± 1.01 ab | 1.57 ± 0.21 b | 1.90 ± 0.49 |
| FS (%) | 6.69 ± 3.30 ab | 15.4 ± 7.4 a | 11.2 ± 4.76 | 13.4 ± 6.9 b |
| SV (mL) | 1.12 ± 0.30 a | 1.56 ± 0.37 ab | 1.36 ± 0.22 | 1.10 ± 0.45 b |
| CO (mL/min) | 357.6 ± 98.1 a | 420.6 ± 98.2 bc | 241.2 ± 39.9 ab | 292.4 ± 125.9 c |
| EF (%) | 20.1 ± 10.1 ab | 38.3 ± 14.7 b | 27.9 ± 11.3 | 37.6 ± 17.1 a |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Silva, A.F.R.; Silva-Reis, R.; Ferreira, R.; Oliveira, P.A.; Faustino-Rocha, A.I.; Pinto, M.d.L.; Coimbra, M.A.; Silva, A.M.S.; Cardoso, S.M. The Impact of Resveratrol-Enriched Bread on Cardiac Remodeling in a Preclinical Model of Diabetes. Antioxidants 2023, 12, 1066. https://doi.org/10.3390/antiox12051066
Silva AFR, Silva-Reis R, Ferreira R, Oliveira PA, Faustino-Rocha AI, Pinto MdL, Coimbra MA, Silva AMS, Cardoso SM. The Impact of Resveratrol-Enriched Bread on Cardiac Remodeling in a Preclinical Model of Diabetes. Antioxidants. 2023; 12(5):1066. https://doi.org/10.3390/antiox12051066
Chicago/Turabian StyleSilva, Andreia F. R., Rita Silva-Reis, Rita Ferreira, Paula A. Oliveira, Ana I. Faustino-Rocha, Maria de Lurdes Pinto, Manuel A. Coimbra, Artur M. S. Silva, and Susana M. Cardoso. 2023. "The Impact of Resveratrol-Enriched Bread on Cardiac Remodeling in a Preclinical Model of Diabetes" Antioxidants 12, no. 5: 1066. https://doi.org/10.3390/antiox12051066
APA StyleSilva, A. F. R., Silva-Reis, R., Ferreira, R., Oliveira, P. A., Faustino-Rocha, A. I., Pinto, M. d. L., Coimbra, M. A., Silva, A. M. S., & Cardoso, S. M. (2023). The Impact of Resveratrol-Enriched Bread on Cardiac Remodeling in a Preclinical Model of Diabetes. Antioxidants, 12(5), 1066. https://doi.org/10.3390/antiox12051066