
Grape Pomace as a Cardiometabolic Health-Promoting Ingredient: Activity in the Intestinal Environment

 ,
,  ,
,  , and
, and
Abstract
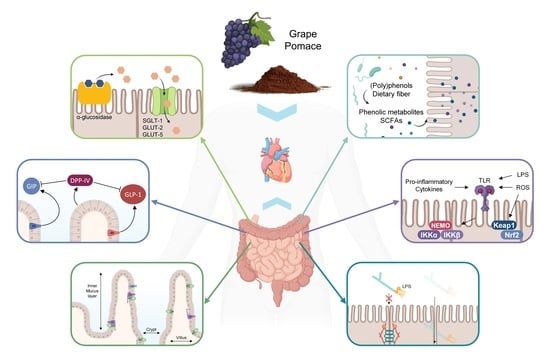
1. Introduction
2. Effects of GP on Intestinal Nutrient Digestion and Absorption
2.1. Physical Impediment on Nutrient Digestion and Absorption
2.2. Regulation of Intestinal Enzymes
2.3. Regulation of Intestinal Transporters
3. Effects of GP on Enteroendocrine Gut Hormones Release and Satiety

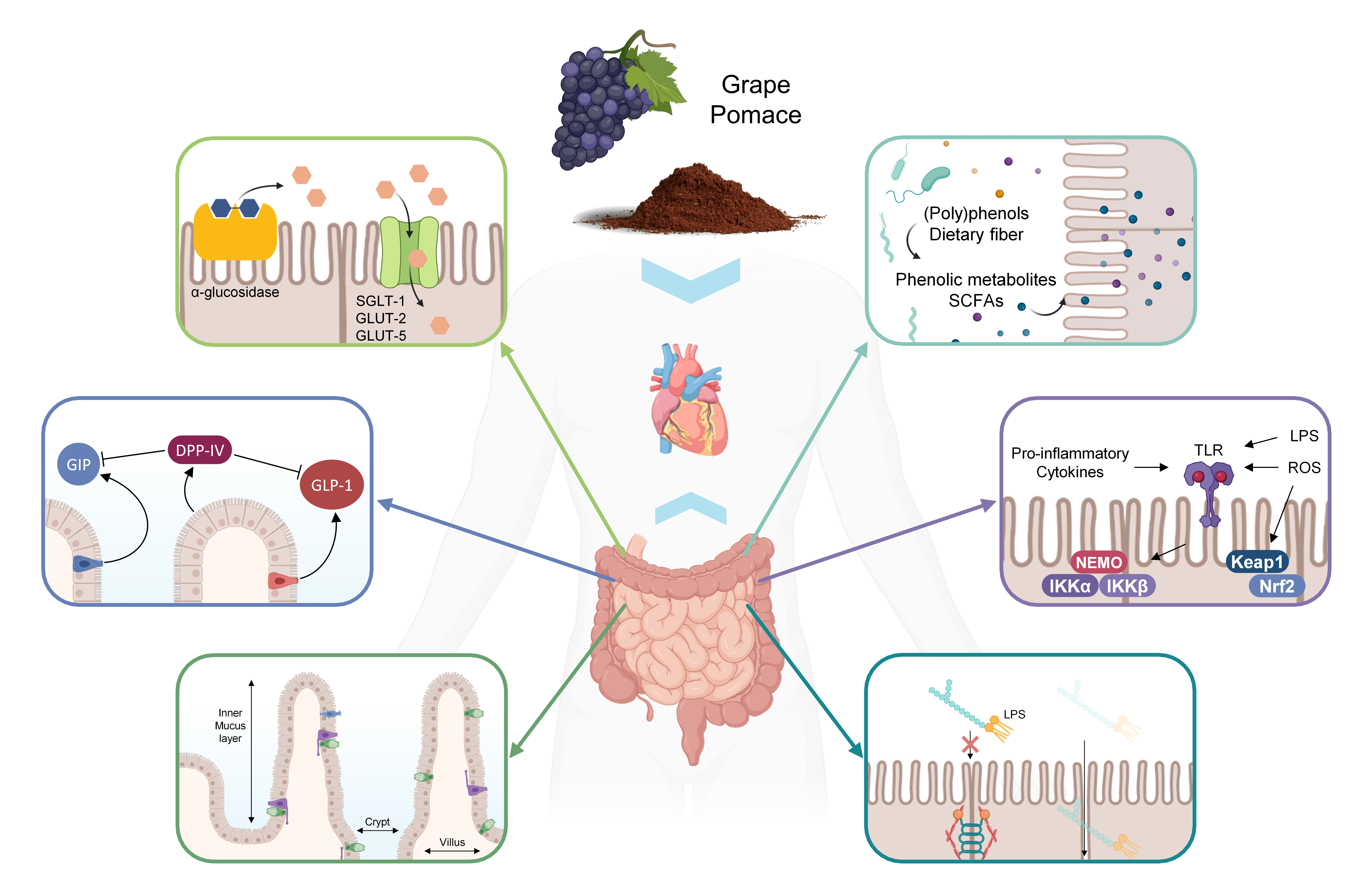
4. Effects of GP on Gut Morphology
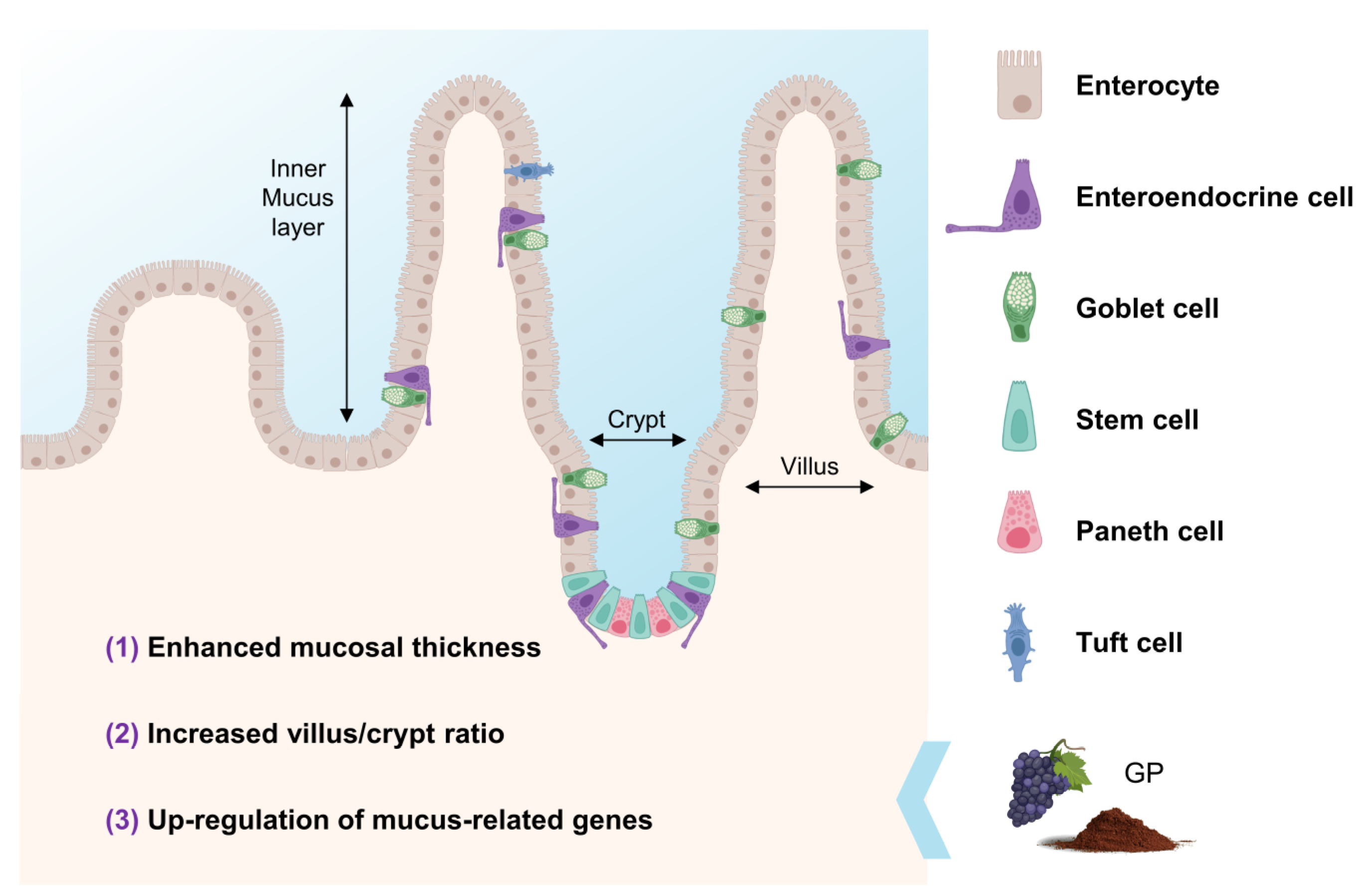
5. Effects of GP on Intestinal Barrier Integrity
6. Effects on Intestinal Inflammatory and Oxidative Status
6.1. Regulation of the NF-κB Signaling Pathway
6.2. Regulation of the Nrf2 Signaling Pathway
7. Effects of GP on the Gut Microbiome
8. Conclusions
Author Contributions
Funding
Conflicts of Interest
Abbreviations
| BMD | bacitracin methylene disalicylate |
| CCK | cholecystokinin |
| GIP | glucose-dependent insulinotropic polypeptide |
| GLP-1 | glucagon-like peptide-1 |
| GP | grape pomace |
| LPS | lipopolysaccharide |
| PYY | peptide YY |
| qPCR | quantitative real-time PCR |
| NF-κB | nuclear factor κB |
| Nrf2 | nuclear factor erythroid 2-related factor 2 |
| SCFAs | short-chain fatty acids |
References
- Bordiga, M.; Travaglia, F.; Locatelli, M. Valorisation of grape pomace: An approach that is increasingly reaching its maturity—A review. Int. J. Food Sci. Technol. 2019, 54, 933–942. [Google Scholar] [CrossRef]
- Ferrer-Gallego, R.; Silva, P. The Wine Industry By-Products: Applications for Food Industry and Health Benefits. Antioxidants 2022, 11, 2025. [Google Scholar] [CrossRef]
- Zhou, D.D.; Li, J.; Xiong, R.G.; Saimaiti, A.; Huang, S.Y.; Wu, S.X.; Yang, Z.J.; Shang, A.; Zhao, C.N.; Gan, R.Y.; et al. Bioactive Compounds, Health Benefits and Food Applications of Grape. Foods 2022, 11, 2755. [Google Scholar] [CrossRef]
- Dwyer, K.; Hosseinian, F.; Rod, M. The Market Potential of Grape Waste Alternatives. J. Food Res. 2014, 3, 91. [Google Scholar] [CrossRef]
- Yu, J.; Ahmedna, M. Functional components of grape pomace: Their composition, biological properties and potential applications. Int. J. Food Sci. Technol. 2013, 48, 221–237. [Google Scholar] [CrossRef]
- García-Lomillo, J.; González-SanJosé, M.L. Applications of Wine Pomace in the Food Industry: Approaches and Functions. Compr. Rev. Food Sci. Food Saf. 2017, 16, 3–22. [Google Scholar] [CrossRef]
- Beres, C.; Costa, G.N.S.; Cabezudo, I.; da Silva-James, N.K.; Teles, A.S.C.; Cruz, A.P.G.; Mellinger-Silva, C.; Tonon, R.V.; Cabral, L.M.C.; Freitas, S.P. Towards integral utilization of grape pomace from winemaking process: A review. Waste Manag. 2017, 68, 581–594. [Google Scholar] [CrossRef]
- Bordiga, M.; Travaglia, F.; Locatelli, M.; Arlorio, M.; Coïsson, J.D. Spent grape pomace as a still potential by-product. Int. J. Food Sci. Technol. 2015, 50, 2022–2031. [Google Scholar] [CrossRef]
- Saura-Calixto, F. Dietary fiber as a carrier of dietary antioxidants: An essential physiological function. J. Agric. Food Chem. 2011, 59, 43–49. [Google Scholar] [CrossRef]
- Gil-Sánchez, I.; Ayuda-Durán, B.; González-Manzano, S.; Santos-Buelga, C.; Cueva, C.; Martín-Cabrejas, M.A.; Sanz-Buenhombre, M.; Guadarrama, A.; Moreno-Arribas, M.V.; Bartolomé, B. Chemical characterization and in vitro colonic fermentation of grape pomace extracts. J. Sci. Food Agric. 2017, 97, 3433–3444. [Google Scholar] [CrossRef]
- Taladrid, D.; Laguna, L.; Vendrell, V.D.; Guadarrana, A.; Moreno-Arribas, M.V.; Bartolomé, B. Sensory acceptability of winery by-products as seasonings for salt replacement. Eur. Food Res. Technol. 2020, 246, 2359–2369. [Google Scholar] [CrossRef]
- Gerardi, G.; Cavia-Saiz, M.; Muñiz, P. From winery by-product to healthy product: Bioavailability, redox signaling and oxidative stress modulation by wine pomace product. Crit. Rev. Food Sci. Nutr. 2022, 62, 7427–7448. [Google Scholar] [CrossRef] [PubMed]
- Ras, R.T.; Zock, P.L.; Zebregs, Y.E.M.P.; Johnston, N.R.; Webb, D.J.; Draijer, R. Effect of polyphenol-rich grape seed extract on ambulatory blood pressure in subjects with pre- and stage I hypertension. Br. J. Nutr. 2013, 110, 2234–2241. [Google Scholar] [CrossRef] [PubMed]
- Urquiaga, I.; D’Acuña, S.; Pérez, D.; Dicenta, S.; Echeverría, G.; Rigotti, A.; Leighton, F. Wine grape pomace flour improves blood pressure, fasting glucose and protein damage in humans: A randomized controlled trial. Biol. Res. 2015, 48, 49. [Google Scholar] [CrossRef] [PubMed]
- Belcaro, G.; Ledda, A.; Hu, S.; Cesarone, M.R.; Feragalli, B.; Dugall, M. Grape seed procyanidins in pre- and mild hypertension: A registry study. Evid. Based Complement. Altern. Med. 2013, 2013, 313142. [Google Scholar] [CrossRef]
- Taladrid, D.; De Celis, M.; Belda, I.; Bartolomé, B.; Moreno-Arribas, M.V. Hypertension- and glycaemia-lowering effects of a grape-pomace-derived seasoning in high-cardiovascular risk and healthy subjects. Interplay with the gut microbiome. Food Funct. 2022, 13, 2068–2082. [Google Scholar] [CrossRef]
- Tomé-Carneiro, J.; Larrosa, M.; Yáñez-Gascón, M.J.; Dávalos, A.; Gil-Zamorano, J.; Gonzálvez, M.; García-Almagro, F.J.; Ruiz Ros, J.A.; Tomás-Barberán, F.A.; Espín, J.C.; et al. One-year supplementation with a grape extract containing resveratrol modulates inflammatory-related microRNAs and cytokines expression in peripheral blood mononuclear cells of type 2 diabetes and hypertensive patients with coronary artery disease. Pharmacol. Res. 2013, 72, 69–82. [Google Scholar] [CrossRef]
- Urquiaga, I.; Troncoso, D.; Mackenna, M.J.; Urzúa, C.; Pérez, D.; Dicenta, S.; de la Cerda, P.M.; Amigo, L.; Carreño, J.C.; Echeverría, G.; et al. The consumption of beef burgers prepared with wine grape pomace flour improves fasting glucose, plasma antioxidant levels, and oxidative damage markers in humans: A controlled trial. Nutrients 2018, 10, 1388. [Google Scholar] [CrossRef]
- Gil-Sánchez, I.; Esteban-Fernández, A.; González de Llano, D.; Sanz-Buenhombre, M.; Guadarrana, A.; Salazar, N.; Gueimonde, M.; de los Reyes-Gavilánc, C.G.; Martín Gómez, L.; García Bermejo, M.L.; et al. Supplementation with grape pomace in healthy women: Changes in biochemical parameters, gut microbiota and related metabolic biomarkers. J. Funct. Foods 2018, 45, 34–46. [Google Scholar] [CrossRef]
- Yang, C.; Han, Y.; Tian, X.; Sajid, M.; Mehmood, S.; Wang, H.; Li, H. Phenolic composition of grape pomace and its metabolism. Crit. Rev. Food Sci. Nutr. 2022, 1–17. [Google Scholar] [CrossRef]
- Chiou, Y.S.; Wu, J.C.; Huang, Q.; Shahidi, F.; Wang, Y.J.; Ho, C.T.; Pan, M.H. Metabolic and colonic microbiota transformation may enhance the bioactivities of dietary polyphenols. J. Funct. Foods 2014, 7, 3–25. [Google Scholar] [CrossRef]
- Cueva, C.; Gil-Sánchez, I.; Ayuda-Durán, B.; González-Manzano, S.; González-Paramás, A.M.; Santos-Buelga, C.; Bartolomé, B.; Victoria Moreno-Arribas, M. An integrated view of the effects of wine polyphenols and their relevant metabolites on gut and host health. Molecules 2017, 22, 99. [Google Scholar] [CrossRef]
- Lu, Y.; Zhang, Y.; Zhao, X.; Shang, C.; Xiang, M.; Li, L.; Cui, X. Microbiota-derived short-chain fatty acids: Implications for cardiovascular and metabolic disease. Front. Cardiovasc. Med. 2022, 9, 2082. [Google Scholar] [CrossRef] [PubMed]
- Campos, F.; Peixoto, A.F.; Fernandes, P.A.R.; Coimbra, M.A.; Mateus, N.; de Freitas, V.; Fernandes, I.; Fernandes, A. The antidiabetic effect of grape pomace polysaccharide-polyphenol complexes. Nutrients 2021, 13, 4495. [Google Scholar] [CrossRef]
- Zhu, F.; Du, B.; Zheng, L.; Li, J. Advance on the bioactivity and potential applications of dietary fibre from grape pomace. Food Chem. 2015, 186, 207–212. [Google Scholar] [CrossRef]
- Giuntini, E.B.; Sardá, F.A.H.; de Menezes, E.W. The Effects of Soluble Dietary Fibers on Glycemic Response: An Overview and Futures Perspectives. Foods 2022, 11, 3934. [Google Scholar] [CrossRef]
- Rocchetti, G.; Rizzi, C.; Cervini, M.; Rainero, G.; Bianchi, F.; Giuberti, G.; Lucini, L.; Simonato, B. Impact of grape pomace powder on the phenolic bioaccessibility and on in vitro starch digestibility of wheat based bread. Foods 2021, 10, 507. [Google Scholar] [CrossRef]
- Hogan, S.; Zhang, L.; Li, J.; Sun, S.; Canning, C.; Zhou, K. Antioxidant rich grape pomace extract suppresses postprandial hyperglycemia in diabetic mice by specifically inhibiting alpha-glucosidase. Nutr. Metab. 2010, 7, 71. [Google Scholar] [CrossRef]
- Asgharpour, F.; Pouramir, M.; Khalilpour, A.; Alamdari, S.A.; Rezaei, M. Antioxidant Activity and Glucose Diffusion Relationship of Traditional Medicinal Antihyperglycemic Plant Extracts. Int. J. Mol. Cell. Med. 2013, 2, 169. [Google Scholar]
- Yu, J.; Mi, Y.; Ji, S. In vitro Evaluating the Influence of Grape Seed Polyphenol Extract on the Digestibility of Macronutrients. J. Health Sci. 2016, 4, 167–176. [Google Scholar] [CrossRef]
- Kato-Schwartz, C.G.; Corrêa, R.C.G.; de Souza Lima, D.; de Sá-Nakanishi, A.B.; de Almeida Gonçalves, G.; Seixas, F.A.V.; Haminiuk, C.W.I.; Barros, L.; Ferreira, I.C.F.R.; Bracht, A.; et al. Potential anti-diabetic properties of Merlot grape pomace extract: An in vitro, in silico and in vivo study of α-amylase and α-glucosidase inhibition. Food Res. Int. 2020, 137, 109462. [Google Scholar] [CrossRef] [PubMed]
- Benítez, V.; Rebollo-Hernanz, M.; Aguilera, Y.; Bejerano, S.; Cañas, S.; Martín-Cabrejas, M.A. Extruded coffee parchment shows enhanced antioxidant, hypoglycaemic, and hypolipidemic properties by releasing phenolic compounds from the fibre matrix. Food Funct. 2021, 12, 1097–1110. [Google Scholar] [CrossRef] [PubMed]
- Adisakwattana, S.; Srichairat, S.; Ngamukote, S.; Moonrat, J.; Chanasit, C.; Tirapongporn, H.; Chanathong, B.; Mäkynen, K.; Sapwarobol, S. Lipid-Lowering mechanisms of grape seed extract (Vitis vinifera L) and its antihyperlidemic activity. Artic. J. Med. Plant Res. 2010, 4, 2113–2120. [Google Scholar]
- Jiao, R.; Zhang, Z.; Yu, H.; Huang, Y.; Chen, Z.Y. Hypocholesterolemic activity of grape seed proanthocyanidin is mediated by enhancement of bile acid excretion and up-regulation of CYP7A1. J. Nutr. Biochem. 2010, 21, 1134–1139. [Google Scholar] [CrossRef] [PubMed]
- Ngamukote, S.; Mäkynen, K.; Thilawech, T.; Adisakwattana, S. Cholesterol-lowering activity of the major polyphenols in grape seed. Molecules 2011, 16, 5054–5061. [Google Scholar] [CrossRef]
- Martinez-Gonzalez, A.I.; Díaz-Sánchez, Á.G.; De La Rosa, L.A.; Vargas-Requena, C.L.; Bustos-Jaimes, I.; Alvarez-Parrilla, E. Polyphenolic compounds and digestive enzymes: In vitro non-covalent interactions. Molecules 2017, 22, 669. [Google Scholar] [CrossRef]
- Yang, X.; Kong, F. Effects of tea polyphenols and different teas on pancreatic α-amylase activity in vitro. LWT Food Sci. Technol. 2016, 66, 232–238. [Google Scholar] [CrossRef]
- Cisneros-Yupanqui, M.; Lante, A.; Mihaylova, D.; Krastanov, A.I.; Rizzi, C. The α-Amylase and α-Glucosidase Inhibition Capacity of Grape Pomace: A Review. Food Bioprocess Technol. 2022, 16, 691–703. [Google Scholar] [CrossRef]
- Hassan, H.M. Inhibitory Effects of Red Grape Seed Extracts on Pancreatic α-amylase and Lipase. Glob. J. Biotech. Biochem. 2014, 9, 130–136. [Google Scholar]
- Jang, Y.S.; Jeong, J.M. Antioxidative Effect and Digestive Enzyme Inhibition of Grape Seed Extract (GSE). J. Korean Soc. Food Sci. Nutr. 2010, 39, 783–788. [Google Scholar] [CrossRef]
- Fernandes, A.C.F.; Martins, I.M.; Moreira, D.K.T.; Macedo, G.A. Use of agro-industrial residues as potent antioxidant, antiglycation agents, and α-amylase and pancreatic lipase inhibitory activity. J. Food Process. Preserv. 2020, 44, e14397. [Google Scholar] [CrossRef]
- Kong, F.; Su, Z.; Zhang, L.; Qin, Y.; Zhang, K. Inclusion complex of grape seeds extracts with sulfobutyl ether β-cyclodextrin: Preparation, characterization, stability and evaluation of α-glucosidase and α-amylase inhibitory effects in vitro. LWT 2019, 101, 819–826. [Google Scholar] [CrossRef]
- Kadouh, H.C.; Sun, S.; Zhu, W.; Zhou, K. α-Glucosidase inhibiting activity and bioactive compounds of six red wine grape pomace extracts. J. Funct. Foods 2016, 26, 577–584. [Google Scholar] [CrossRef] [PubMed]
- Lavelli, V.; Sri Harsha, P.S.C.; Fiori, L. Screening grape seeds recovered from winemaking by-products as sources of reducing agents and mammalian α-glucosidase and α-amylase inhibitors. Int. J. Food Sci. Technol. 2015, 50, 1182–1189. [Google Scholar] [CrossRef]
- Huamán-Castilla, N.L.; Campos, D.; García-Ríos, D.; Parada, J.; Martínez-Cifuentes, M.; Mariotti-Celis, M.S.; Pérez-Correa, J.R. Chemical properties of vitis vinifera carménère pomace extracts obtained by hot pressurized liquid extraction, and their inhibitory effect on type 2 diabetes mellitus related enzymes. Antioxidants 2021, 10, 472. [Google Scholar] [CrossRef]
- Fernández-Fernández, A.M.; Iriondo-DeHond, A.; Dellacassa, E.; Medrano-Fernandez, A.; del Castillo, M.D. Assessment of antioxidant, antidiabetic, antiobesity, and anti-inflammatory properties of a Tannat winemaking by-product. Eur. Food Res. Technol. 2019, 245, 1539–1551. [Google Scholar] [CrossRef]
- Freire, J.M.; Barroso, A.R.; de Assis, A.A.; Texeira, B.H.; Braga, J.H.G.; Oliveira, D.A.; Braga, M.A.; Marcussi, S. Enzymes modulation by dried grape pomace from the manufacture of wines and juices. Braz. J. Pharm. Sci. 2021, 56, e18467. [Google Scholar] [CrossRef]
- Uuh Narvaez, J.J.; Segura Campos, M.R. Combination therapy of bioactive compounds with acarbose: A proposal to control hyperglycemia in type 2 diabetes. J. Food Biochem. 2022, 46, e14268. [Google Scholar] [CrossRef]
- Li, N.; Liu, X.; Zhang, J.; Lang, Y.Z.; Lu, L.; Mi, J.; Cao, Y.L.; Yan, Y.M.; Ran, L.W. Preventive Effects of Anthocyanins from Lycium ruthenicum Murray in High-Fat Diet-Induced Obese Mice Are Related to the Regulation of Intestinal Microbiota and Inhibition of Pancreatic Lipase Activity. Molecules 2022, 27, 2141. [Google Scholar] [CrossRef]
- Léniz, A.; Martínez-Maqueda, D.; Fernández-Quintela, A.; Pérez-Jiménez, J.; Portillo, M.P. Potential relationship between the changes in circulating micrornas and the improvement in glycaemic control induced by grape pomace supplementation. Foods 2021, 10, 2059. [Google Scholar] [CrossRef]
- Martínez-Meza, Y.; Pérez-Jiménez, J.; Salgado-Rodríguez, L.M.; Castellanos-Jiménez, A.K.; Reynoso-Camacho, R. In Vivo Evaluation of the Cardiometabolic Potential of Grape Pomace: Effect of Applying Instant Controlled Pressure Drop. Foods 2022, 11, 3537. [Google Scholar] [CrossRef]
- Bibi, S.; Kang, Y.; Yang, G.; Zhu, M.J. Grape seed extract improves small intestinal health through suppressing inflammation and regulating alkaline phosphatase in IL-10-deficient mice. J. Funct. Foods 2016, 20, 245–252. [Google Scholar] [CrossRef]
- Zhang, L.; Carmody, R.N.; Kalariya, H.M.; Duran, R.M.; Moskal, K.; Poulev, A.; Kuhn, P.; Tveter, K.M.; Turnbaugh, P.J.; Raskin, I.; et al. Grape proanthocyanidin-induced intestinal bloom of Akkermansia muciniphila is dependent on its baseline abundance and precedes activation of host genes related to metabolic health. J. Nutr. Biochem. 2018, 56, 142–151. [Google Scholar] [CrossRef] [PubMed]
- Fiesel, A.; Gessner, D.K.; Most, E.; Eder, K. Effects of dietary polyphenol-rich plant products from grape or hop on pro-inflammatory gene expression in the intestine, nutrient digestibility and faecal microbiota of weaned pigs. BMC Vet. Res. 2014, 10, 196. [Google Scholar] [CrossRef] [PubMed]
- Fernández-Fernández, A.M.; Iriondo-Dehond, A.; Nardin, T.; Larcher, R.; Dellacassa, E.; Medrano-Fernandez, A.; Del Castillo, M.D. In Vitro Bioaccessibility of Extractable Compounds from Tannat Grape Skin Possessing Health Promoting Properties with Potential to Reduce the Risk of Diabetes. Foods 2020, 9, 1575. [Google Scholar] [CrossRef]
- Heidker, R.M.; Caiozzi, G.C.; Ricketts, M.L. Grape Seed Procyanidins and Cholestyramine Differentially Alter Bile Acid and Cholesterol Homeostatic Gene Expression in Mouse Intestine and Liver. PLoS ONE 2016, 11, e0154305. [Google Scholar] [CrossRef]
- Van Hul, M.; Geurts, L.; Plovier, H.; Druart, C.; Everard, A.; Ståhlman, M.; Rhimi, M.; Chira, K.; Teissedre, P.L.; Delzenne, N.M.; et al. Reduced obesity, diabetes, and steatosis upon cinnamon and grape pomace are associated with changes in gut microbiota and markers of gut barrier. Am. J. Physiol. Endocrinol. Metab. 2018, 314, E334–E352. [Google Scholar] [CrossRef]
- Wu, Y.; Mo, R.; Zhang, M.; Zhou, W.; Li, D. Grape Seed Proanthocyanidin Alleviates Intestinal Inflammation through Gut Microbiota-Bile Acid Crosstalk in Mice. Front. Nutr. 2022, 8, 1316. [Google Scholar] [CrossRef]
- Leifert, W.R.; Abeywardena, M.Y. Grape seed and red wine polyphenol extracts inhibit cellular cholesterol uptake, cell proliferation, and 5-lipoxygenase activity. Nutr. Res. 2008, 28, 842–850. [Google Scholar] [CrossRef]
- Serrano, J.; Casanova-Martí, À.; Gual, A.; Pérez-Vendrell, A.M.; Blay, M.T.; Terra, X.; Ardévol, A.; Pinent, M. A specific dose of grape seed-derived proanthocyanidins to inhibit body weight gain limits food intake and increases energy expenditure in rats. Eur. J. Nutr. 2017, 56, 1629–1636. [Google Scholar] [CrossRef]
- Serrano, J.; Casanova-Martí, À.; Blay, M.; Terra, X.; Ardévol, A.; Pinent, M. Defining Conditions for Optimal Inhibition of Food Intake in Rats by a Grape-Seed Derived Proanthocyanidin Extract. Nutrients 2016, 8, 652. [Google Scholar] [CrossRef]
- Serrano, J.; Casanova-Martí, À.; Gil-Cardoso, K.; Blay, M.T.; Terra, X.; Pinent, M.; Ardévol, A. Acutely administered grape-seed proanthocyanidin extract acts as a satiating agent. Food Funct. 2016, 7, 483–490. [Google Scholar] [CrossRef] [PubMed]
- Li, H.; Parry, J.; Weeda, S.; Ren, S.; Castonguay, T.W.; Guo, T.L. Grape Pomace Aqueous Extract (GPE) Prevents High Fat Diet-Induced Diabetes and Attenuates Systemic Inflammation. Food Nutr. Sci. 2016, 7, 647–660. [Google Scholar] [CrossRef]
- Casanova-Martí, À.; Serrano, J.; Portune, K.J.; Sanz, Y.; Blay, M.T.; Terra, X.; Ardévol, A.; Pinent, M. Grape seed proanthocyanidins influence gut microbiota and enteroendocrine secretions in female rats. Food Funct. 2018, 9, 1672–1682. [Google Scholar] [CrossRef] [PubMed]
- González-Abuín, N.; Martínez-Micaelo, N.; Margalef, M.; Blay, M.; Arola-Arnal, A.; Muguerza, B.; Ardévol, A.; Pinent, M. A grape seed extract increases active glucagon-like peptide-1 levels after an oral glucose load in rats. Food Funct. 2014, 5, 2357–2364. [Google Scholar] [CrossRef]
- González-Abuín, N.; Martínez-Micaelo, N.; Blay, M.; Ardévol, A.; Pinent, M. Grape-seed procyanidins prevent the cafeteria-diet-induced decrease of glucagon-like peptide-1 production. J. Agric. Food Chem. 2014, 62, 1066–1072. [Google Scholar] [CrossRef]
- González-Abuín, N.; Martínez-Micaelo, N.; Blay, M.; Green, B.D.; Pinent, M.; Ardévol, A. Grape-seed procyanidins modulate cellular membrane potential and nutrient-induced GLP-1 secretion in STC-1 cells. Am. J. Physiol. Cell Physiol. 2014, 306, 485–492. [Google Scholar] [CrossRef]
- Casanova-Martí, À.; González-Abuín, N.; Serrano, J.; Blay, M.T.; Terra, X.; Frost, G.; Pinent, M.; Ardévol, A. Long Term Exposure to a Grape Seed Proanthocyanidin Extract Enhances L-Cell Differentiation in Intestinal Organoids. Mol. Nutr. Food Res. 2020, 64, 2000303. [Google Scholar] [CrossRef]
- Casanova-Martí, À.; Serrano, J.; Blay, M.T.; Terra, X.; Ardévol, A.; Pinent, M. Acute selective bioactivity of grape seed proanthocyanidins on enteroendocrine secretions in the gastrointestinal tract. Food Nutr. Res. 2017, 61, 1321347. [Google Scholar] [CrossRef]
- Ginés, I.; Gil-Cardoso, K.; Terra, X.; Blay, M.; Pérez-Vendrell, A.M.; Pinent, M.; Ardévol, A. Grape Seed Proanthocyanidins Target the Enteroendocrine System in Cafeteria-Diet-Fed Rats. Mol. Nutr. Food Res. 2019, 63, 1800912. [Google Scholar] [CrossRef]
- Karhunen, L.J.; Juvonen, K.R.; Huotari, A.; Purhonen, A.K.; Herzig, K.H. Effect of protein, fat, carbohydrate and fibre on gastrointestinal peptide release in humans. Regul. Pept. 2008, 149, 70–78. [Google Scholar] [CrossRef] [PubMed]
- Lu, V.B.; Gribble, F.M.; Reimann, F. Nutrient-induced cellular mechanisms of gut hormone secretion. Nutrients 2021, 13, 883. [Google Scholar] [CrossRef]
- Bastings, J.J.A.J.; Venema, K.; Blaak, E.E.; Adam, T.C. Influence of the gut microbiota on satiety signaling. Trends Endocrinol. Metab. 2023, 34, 243–255. [Google Scholar] [CrossRef] [PubMed]
- González-Abuín, N.; Martínez-Micaelo, N.; Blay, M.; Pujadas, G.; Garcia-Vallvé, S.; Pinent, M.; Ardévol, A. Grape seed-derived procyanidins decrease dipeptidyl-peptidase 4 activity and expression. J. Agric. Food Chem. 2012, 60, 9055–9061. [Google Scholar] [CrossRef]
- Serrano, J.; Casanova-Martí, À.; Depoortere, I.; Blay, M.T.; Terra, X.; Pinent, M.; Ardévol, A. Subchronic treatment with grape-seed phenolics inhibits ghrelin production despite a short-term stimulation of ghrelin secretion produced by bitter-sensing flavanols. Mol. Nutr. Food Res. 2016, 60, 2554–2564. [Google Scholar] [CrossRef]
- Walton, K.D.; Mishkind, D.; Riddle, M.R.; Tabin, C.J.; Gumucio, D.L. Blueprint for an intestinal villus: Species-specific assembly required. Wiley Interdiscip. Rev. Dev. Biol. 2018, 7, e317. [Google Scholar] [CrossRef]
- Boussenna, A.; Goncalves-Mendes, N.; Joubert-Zakeyh, J.; Pereira, B.; Fraisse, D.; Vasson, M.P.; Texier, O.; Felgines, C. Impact of basal diet on dextran sodium sulphate (DSS)-induced colitis in rats. Eur. J. Nutr. 2015, 54, 1217–1227. [Google Scholar] [CrossRef]
- Wang, R.; Yu, H.; Fang, H.; Jin, Y.; Zhao, Y.; Shen, J.; Zhou, C.; Li, R.; Wang, J.; Fu, Y.; et al. Effects of dietary grape pomace on the intestinal microbiota and growth performance of weaned piglets. Arch. Anim. Nutr. 2020, 74, 296–308. [Google Scholar] [CrossRef]
- Wang, Y.H.; Yang, X.L.; Wang, L.; Cui, M.X.; Cai, Y.Q.; Li, X.L.; Wu, Y.J. Effects of proanthocyanidins from grape seed on treatment of recurrent ulcerative colitis in rats. Can. J. Physiol. Pharmacol. 2010, 88, 888–898. [Google Scholar] [CrossRef]
- Gessner, D.K.; Fiesel, A.; Most, E.; Dinges, J.; Wen, G.; Ringseis, R.; Eder, K. Supplementation of a grape seed and grape marc meal extract decreases activities of the oxidative stress-responsive transcription factors NF-κB and Nrf2 in the duodenal mucosa of pigs. Acta Vet. Scand. 2013, 55, 18. [Google Scholar] [CrossRef]
- Zhang, R.; Li, Q.; Zhang, T.; Qin, X.Z.; Zhang, J.; Zhao, J. Dietary grape pomace mediates jejunum epithelial apoptosis through antioxidative activity in ram lambs. Ital. J. Anim. Sci. 2019, 18, 1058–1066. [Google Scholar] [CrossRef]
- Lu, F.; Li, Y.; Zhou, B.; Guo, Q.; Zhang, Y. Early-life supplementation of grape polyphenol extract promotes polyphenol absorption and modulates the intestinal microbiota in association with the increase in mRNA expression of the key intestinal barrier genes. Food Funct. 2021, 12, 602–613. [Google Scholar] [CrossRef] [PubMed]
- Pistol, G.C.; Bulgaru, C.V.; Marin, D.E.; Oancea, A.G.; Taranu, I. Dietary grape seed meal bioactive compounds alleviate epithelial dysfunctions and attenuates inflammation in colon of dss-treated piglets. Foods 2021, 10, 530. [Google Scholar] [CrossRef]
- Yang, G.; Wang, H.; Kang, Y.; Zhu, M.J. Grape seed extract improves epithelial structure and suppresses inflammation in ileum of IL-10-deficient mice. Food Funct. 2014, 5, 2558–2563. [Google Scholar] [CrossRef]
- Maurer, L.H.; Cazarin, C.B.B.; Quatrin, A.; Minuzzi, N.M.; Costa, E.L.; Morari, J.; Velloso, L.A.; Leal, R.F.; Rodrigues, E.; Bochi, V.C.; et al. Grape peel powder promotes intestinal barrier homeostasis in acute TNBS-colitis: A major role for dietary fiber and fiber-bound polyphenols. Food Res. Int. 2019, 123, 425–439. [Google Scholar] [CrossRef]
- Kwon, O.; Han, T.S.; Son, M.Y. Intestinal Morphogenesis in Development, Regeneration, and Disease: The Potential Utility of Intestinal Organoids for Studying Compartmentalization of the Crypt-Villus Structure. Front. Cell Dev. Biol. 2020, 8, 1192. [Google Scholar] [CrossRef]
- Bernardi, S.; Del Bo, C.; Marino, M.; Gargari, G.; Cherubini, A.; Andrés-Lacueva, C.; Hidalgo-Liberona, N.; Peron, G.; González-Dominguez, R.; Kroon, P.; et al. Polyphenols and Intestinal Permeability: Rationale and Future Perspectives. J. Agric. Food Chem. 2020, 68, 1816–1829. [Google Scholar] [CrossRef]
- Genser, L.; Aguanno, D.; Soula, H.A.; Dong, L.; Trystram, L.; Assmann, K.; Salem, J.E.; Vaillant, J.C.; Oppert, J.M.; Laugerette, F.; et al. Increased jejunal permeability in human obesity is revealed by a lipid challenge and is linked to inflammation and type 2 diabetes. J. Pathol. 2018, 246, 217–230. [Google Scholar] [CrossRef]
- Gil-Cardoso, K.; Ginés, I.; Pinent, M.; Ardévol, A.; Arola, L.; Blay, M.; Terra, X. Chronic supplementation with dietary proanthocyanidins protects from diet-induced intestinal alterations in obese rats. Mol. Nutr. Food Res. 2017, 61, 1601039. [Google Scholar] [CrossRef]
- Nallathambi, R.; Poulev, A.; Zuk, J.B.; Raskin, I. Proanthocyanidin-rich grape seed extract reduces inflammation and oxidative stress and restores tight junction barrier function in caco-2 colon cells. Nutrients 2020, 12, 1623. [Google Scholar] [CrossRef]
- Reiche, J.; Huber, O. Post-translational modifications of tight junction transmembrane proteins and their direct effect on barrier function. Biochim. Biophys. Acta Biomembr. 2020, 1862, 183330. [Google Scholar] [CrossRef] [PubMed]
- Hidalgo-Liberona, N.; González-Domínguez, R.; Vegas, E.; Riso, P.; Del Bo, C.; Bernardi, S.; Peron, G.; Guglielmetti, S.; Gargari, G.; Kroon, P.A.; et al. Increased Intestinal Permeability in Older Subjects Impacts the Beneficial Effects of Dietary Polyphenols by Modulating Their Bioavailability. J. Agric. Food Chem. 2020, 68, 12476–12484. [Google Scholar] [CrossRef] [PubMed]
- Yang, G.; Xue, Y.; Zhang, H.; Du, M.; Zhu, M.J. Favourable effects of grape seed extract on intestinal epithelial differentiation and barrier function in IL10-deficient mice. Br. J. Nutr. 2015, 114, 15–23. [Google Scholar] [CrossRef] [PubMed]
- Cremonini, E.; Mastaloudis, A.; Hester, S.N.; Verstraeten, S.V.; Anderson, M.; Wood, S.M.; Waterhouse, A.L.; Fraga, C.G.; Oteiza, P.I. Anthocyanins inhibit tumor necrosis alpha-induced loss of Caco-2 cell barrier integrity. Food Funct. 2017, 8, 2915–2923. [Google Scholar] [CrossRef]
- Gil-Cardoso, K.; Ginés, I.; Pinent, M.; Ardévol, A.; Blay, M.; Terra, X. The co-administration of proanthocyanidins and an obesogenic diet prevents the increase in intestinal permeability and metabolic endotoxemia derived to the diet. J. Nutr. Biochem. 2018, 62, 35–42. [Google Scholar] [CrossRef]
- González-Quilen, C.; Grau-Bové, C.; Jorba-Martín, R.; Caro-Tarragó, A.; Pinent, M.; Ardévol, A.; Beltrán-Debón, R.; Terra, X.; Blay, M.T. Protective properties of grape-seed proanthocyanidins in human ex vivo acute colonic dysfunction induced by dextran sodium sulfate. Eur. J. Nutr. 2021, 60, 79–88. [Google Scholar] [CrossRef]
- Taladrid, D.; de Llano, D.G.; Zorraquín-Peña, I.; Tamargo, A.; Silva, M.; Molinero, N.; Moreno-Arribas, M.V.; Bartolomé, B. Gastrointestinal digestion of a grape pomace extract: Impact on intestinal barrier permeability and interaction with gut microbiome. Nutrients 2021, 13, 2467. [Google Scholar] [CrossRef]
- Sheng, K.; Zhang, G.; Sun, M.; He, S.; Kong, X.; Wang, J.; Zhu, F.; Zha, X.; Wang, Y. Grape seed proanthocyanidin extract ameliorates dextran sulfate sodium-induced colitis through intestinal barrier improvement, oxidative stress reduction, and inflammatory cytokines and gut microbiota modulation. Food Funct. 2020, 11, 7817–7829. [Google Scholar] [CrossRef]
- Lian, P.; Braber, S.; Garssen, J.; Wichers, H.J.; Folkerts, G.; Fink-Gremmels, J.; Varasteh, S. Beyond heat stress: Intestinal integrity disruption and mechanism-based intervention strategies. Nutrients 2020, 12, 734. [Google Scholar] [CrossRef]
- Koudoufio, M.; Feldman, F.; Ahmarani, L.; Delvin, E.; Spahis, S.; Desjardins, Y.; Levy, E. Intestinal protection by proanthocyanidins involves anti-oxidative and anti-inflammatory actions in association with an improvement of insulin sensitivity, lipid and glucose homeostasis. Sci. Rep. 2021, 11, 3878. [Google Scholar] [CrossRef]
- Shivakoti, R.; Biggs, M.L.; Djoussé, L.; Durda, P.J.; Kizer, J.R.; Psaty, B.; Reiner, A.P.; Tracy, R.P.; Siscovick, D.; Mukamal, K.J. Intake and Sources of Dietary Fiber, Inflammation, and Cardiovascular Disease in Older US Adults. JAMA Netw. Open 2022, 5, e225012. [Google Scholar] [CrossRef] [PubMed]
- Pistol, G.C.; Marin, D.E.; Rotar, M.C.; Ropota, M.; Taranu, I. Bioactive compounds from dietary whole grape seed meal improved colonic inflammation via inhibition of MAPKs and NF-kB signaling in pigs with DSS induced colitis. J. Funct. Foods 2020, 66, 103708. [Google Scholar] [CrossRef]
- Maurer, L.H.; Cazarin, C.B.B.; Quatrin, A.; Nichelle, S.M.; Minuzzi, N.M.; Teixeira, C.F.; Manica da Cruz, I.B.; Maróstica Júnior, M.R.; Emanuelli, T. Dietary fiber and fiber-bound polyphenols of grape peel powder promote GSH recycling and prevent apoptosis in the colon of rats with TNBS-induced colitis. J. Funct. Foods 2020, 64, 103644. [Google Scholar] [CrossRef]
- Wang, Y.H.; Ge, B.; Yang, X.L.; Zhai, J.; Yang, L.N.; Wang, X.X.; Liu, X.; Shi, J.C.; Wu, Y.J. Proanthocyanidins from grape seeds modulates the nuclear factor-kappa B signal transduction pathways in rats with TNBS-induced recurrent ulcerative colitis. Int. Immunopharmacol. 2011, 11, 1620–1627. [Google Scholar] [CrossRef] [PubMed]
- Kafantaris, I.; Stagos, D.; Kotsampasi, B.; Hatzis, A.; Kypriotakis, A.; Gerasopoulos, K.; Makri, S.; Goutzourelas, N.; Mitsagga, C.; Giavasis, I.; et al. Grape pomace improves performance, antioxidant status, fecal microbiota and meat quality of piglets. Animal 2018, 12, 246–255. [Google Scholar] [CrossRef]
- Christian, F.; Smith, E.L.; Carmody, R.J. The regulation of NF-kB Subunits by Phosphorylation. Cells 2016, 5, 12. [Google Scholar] [CrossRef]
- Hug, H.; Mohajeri, M.H.; La Fata, G. Toll-Like Receptors: Regulators of the Immune Response in the Human Gut. Nutrients 2018, 10, 203. [Google Scholar] [CrossRef]
- Ritchie, L.E.; Taddeo, S.S.; Weeks, B.R.; Carroll, R.J.; Dykes, L.; Rooney, L.W.; Turner, N.D. Impact of Novel Sorghum Bran Diets on DSS-Induced Colitis. Nutrients 2017, 9, 330. [Google Scholar] [CrossRef]
- Saha, R.N.; Jana, M.; Pahan, K. MAPK p38 Regulates Transcriptional Activity of NF-κB in Primary Human Astrocytes via Acetylation of p65. J. Immunol. 2007, 179, 7101–7109. [Google Scholar] [CrossRef]
- Thangaraju, M.; Cresci, G.A.; Liu, K.; Ananth, S.; Gnanaprakasam, J.P.; Browning, D.D.; Mellinger, J.D.; Smith, S.B.; Digby, G.J.; Lambert, N.A.; et al. GPFM 09A is a G-protein-coupled receptor for the bacterial fermentation product butyrate and functions as a tumor suppressor in colon. Cancer Res. 2009, 69, 2826–2832. [Google Scholar] [CrossRef]
- Sahasrabudhe, N.M.; Beukema, M.; Tian, L.; Troost, B.; Scholte, J.; Bruininx, E.; Bruggeman, G.; van den Berg, M.; Scheurink, A.; Schols, H.A.; et al. Dietary fiber pectin directly blocks toll-like receptor 2-1 and prevents doxorubicin-induced ileitis. Front. Immunol. 2018, 9, 383. [Google Scholar] [CrossRef] [PubMed]
- Shaeib, F.; Maitra, D.; Banerjee, J.; Abdulhamid, I.; Diamond, M.; Saed, G.; Pennathur, S.; Abu-Soud, H. Melatonin Prevents Myeloperoxidase Heme Destruction and the Generation of Free Iron Mediated by Self-Generated Hypochlorous Acid. Free Radic. Biol. Med. 2013, 65, S131. [Google Scholar] [CrossRef]
- Hussain, T.; Tan, B.; Yin, Y.; Blachier, F.; Tossou, M.C.B.B.; Rahu, N. Oxidative Stress and Inflammation: What Polyphenols Can Do for Us? Oxidative Med. Cell. Longev. 2016, 2016, 7432797. [Google Scholar] [CrossRef] [PubMed]
- Pisoschi, A.M.; Pop, A. The role of antioxidants in the chemistry of oxidative stress: A review. Eur. J. Med. Chem. 2015, 97, 55–74. [Google Scholar] [CrossRef]
- Rebollo-Hernanz, M.; Kusumah, J.; Bringe, N.A.; Shen, Y.; de Mejia, E.G. Peptide release, radical scavenging capacity, and antioxidant responses in intestinal cells are determined by soybean variety and gastrointestinal digestion under simulated conditions. Food Chem. 2023, 405, 134929. [Google Scholar] [CrossRef]
- Wardyn, J.D.; Ponsford, A.H.; Sanderson, C.M. Dissecting molecular cross-talk between Nrf2 and NF-κB response pathways. Biochem. Soc. Trans. 2015, 43, 621–626. [Google Scholar] [CrossRef]
- Niture, S.K.; Kaspar, J.W.; Shen, J.; Jaiswal, A.K. Nrf2 signaling and cell survival. Toxicol. Appl. Pharmacol. 2010, 244, 37–42. [Google Scholar] [CrossRef]
- Li, S.; Eguchi, N.; Lau, H.; Ichii, H. The role of the nrf2 signaling in obesity and insulin resistance. Int. J. Mol. Sci. 2020, 21, 6973. [Google Scholar] [CrossRef]
- Brody, H. The gut microbiome. Nature 2020, 577, S5. [Google Scholar] [CrossRef]
- Pascale, A.; Marchesi, N.; Govoni, S.; Coppola, A.; Gazzaruso, C. The role of gut microbiota in obesity, diabetes mellitus, and effect of metformin: New insights into old diseases. Curr. Opin. Pharmacol. 2019, 49, 1–5. [Google Scholar] [CrossRef]
- Dominianni, C.; Sinha, R.; Goedert, J.J.; Pei, Z.; Yang, L.; Hayes, R.B.; Ahn, J. Sex, body mass index, and dietary fiber intake influence the human gut microbiome. PLoS ONE 2015, 10, e0124599. [Google Scholar] [CrossRef] [PubMed]
- Dueñas, M.; Muñoz-González, I.; Cueva, C.; Jiménez-Girón, A.; Sánchez-Patán, F.; Santos-Buelga, C.; Moreno-Arribas, M.V.; Bartolomé, B. A survey of modulation of gut microbiota by dietary polyphenols. BioMed Res. Int. 2015, 2015, 850902. [Google Scholar] [CrossRef] [PubMed]
- Gil-Sánchez, I.; Cueva, C.; Sanz-Buenhombre, M.; Guadarrama, A.; Moreno-Arribas, M.V.; Bartolomé, B. Dynamic gastrointestinal digestion of grape pomace extracts: Bioaccessible phenolic metabolites and impact on human gut microbiota. J. Food Compos. Anal. 2018, 68, 41–52. [Google Scholar] [CrossRef]
- Cardona, F.; Andrés-Lacueva, C.; Tulipani, S.; Tinahones, F.J.; Queipo-Ortuño, M.I. Benefits of polyphenols on gut microbiota and implications in human health. J. Nutr. Biochem. 2013, 24, 1415–1422. [Google Scholar] [CrossRef] [PubMed]
- Koh, A.; De Vadder, F.; Kovatcheva-Datchary, P.; Bäckhed, F. From Dietary Fiber to Host Physiology: Short-Chain Fatty Acids as Key Bacterial Metabolites. Cell 2016, 165, 1332–1345. [Google Scholar] [CrossRef]
- Viveros, A.; Chamorro, S.; Pizarro, M.; Arija, I.; Centeno, C.; Brenes, A. Effects of dietary polyphenol-rich grape products on intestinal microflora and gut morphology in broiler chicks. Poult. Sci. 2011, 90, 566–578. [Google Scholar] [CrossRef]
- Erinle, T.J.; Oladokun, S.; MacIsaac, J.; Rathgeber, B.; Adewole, D. Dietary grape pomace—Effects on growth performance, intestinal health, blood parameters, and breast muscle myopathies of broiler chickens. Poult. Sci. 2022, 101, 101519. [Google Scholar] [CrossRef]
- Chacar, S.; Tarighi, M.; Fares, N.; Faivre, J.F.; Louka, N.; Maroun, R.G. Identification of phenolic compounds-rich grape pomace extracts urine metabolites and correlation with gut microbiota modulation. Antioxidants 2018, 7, 75. [Google Scholar] [CrossRef]
- Chacar, S.; Itani, T.; Hajal, J.; Saliba, Y.; Louka, N.; Faivre, J.F.; Maroun, R.; Fares, N. The Impact of Long-Term Intake of Phenolic Compounds-Rich Grape Pomace on Rat Gut Microbiota. J. Food Sci. 2018, 83, 246–251. [Google Scholar] [CrossRef]
- Lu, F.; Liu, F.; Zhou, Q.; Hu, X.; Zhang, Y. Effects of grape pomace and seed polyphenol extracts on the recovery of gut microbiota after antibiotic treatment in high-fat diet-fed mice. Food Sci. Nutr. 2019, 7, 2897–2906. [Google Scholar] [CrossRef]
- Kafantaris, I.; Kotsampasi, B.; Christodoulou, V.; Kokka, E.; Kouka, P.; Terzopoulou, Z.; Gerasopoulos, K.; Stagos, D.; Mitsagga, C.; Giavasis, I.; et al. Grape pomace improves antioxidant capacity and faecal microflora of lambs. J. Anim. Physiol. Anim. Nutr. 2017, 101, e108–e121. [Google Scholar] [CrossRef] [PubMed]
- Ramos-Romero, S.; Martínez-Maqueda, D.; Hereu, M.; Amézqueta, S.; Torres, J.L.; Pérez-Jiménez, J. Modifications of gut microbiota after grape pomace supplementation in subjects at cardiometabolic risk: A randomized cross-over controlled clinical trial. Foods 2020, 9, 1279. [Google Scholar] [CrossRef]
- Ramos-Romero, S.; Léniz, A.; Martínez-Maqueda, D.; Amézqueta, S.; Fernández-Quintela, A.; Hereu, M.; Torres, J.L.; Portillo, M.P.; Pérez-Jiménez, J. Inter-Individual Variability in Insulin Response after Grape Pomace Supplementation in Subjects at High Cardiometabolic Risk: Role of Microbiota and miRNA. Mol. Nutr. Food Res. 2021, 65, 2000113. [Google Scholar] [CrossRef]
- Annunziata, G.; Ciampaglia, R.; Maisto, M.; D’avino, M.; Caruso, D.; Tenore, G.C.; Novellino, E. Taurisolo®, a grape pomace polyphenol nutraceutical reducing the levels of serum biomarkers associated with atherosclerosis. Front. Cardiovasc. Med. 2021, 8, 732. [Google Scholar] [CrossRef]
- Zhu, Y.; Li, Q.; Jiang, H. Gut microbiota in atherosclerosis: Focus on trimethylamine N-oxide. APMIS 2020, 128, 353–366. [Google Scholar] [CrossRef] [PubMed]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
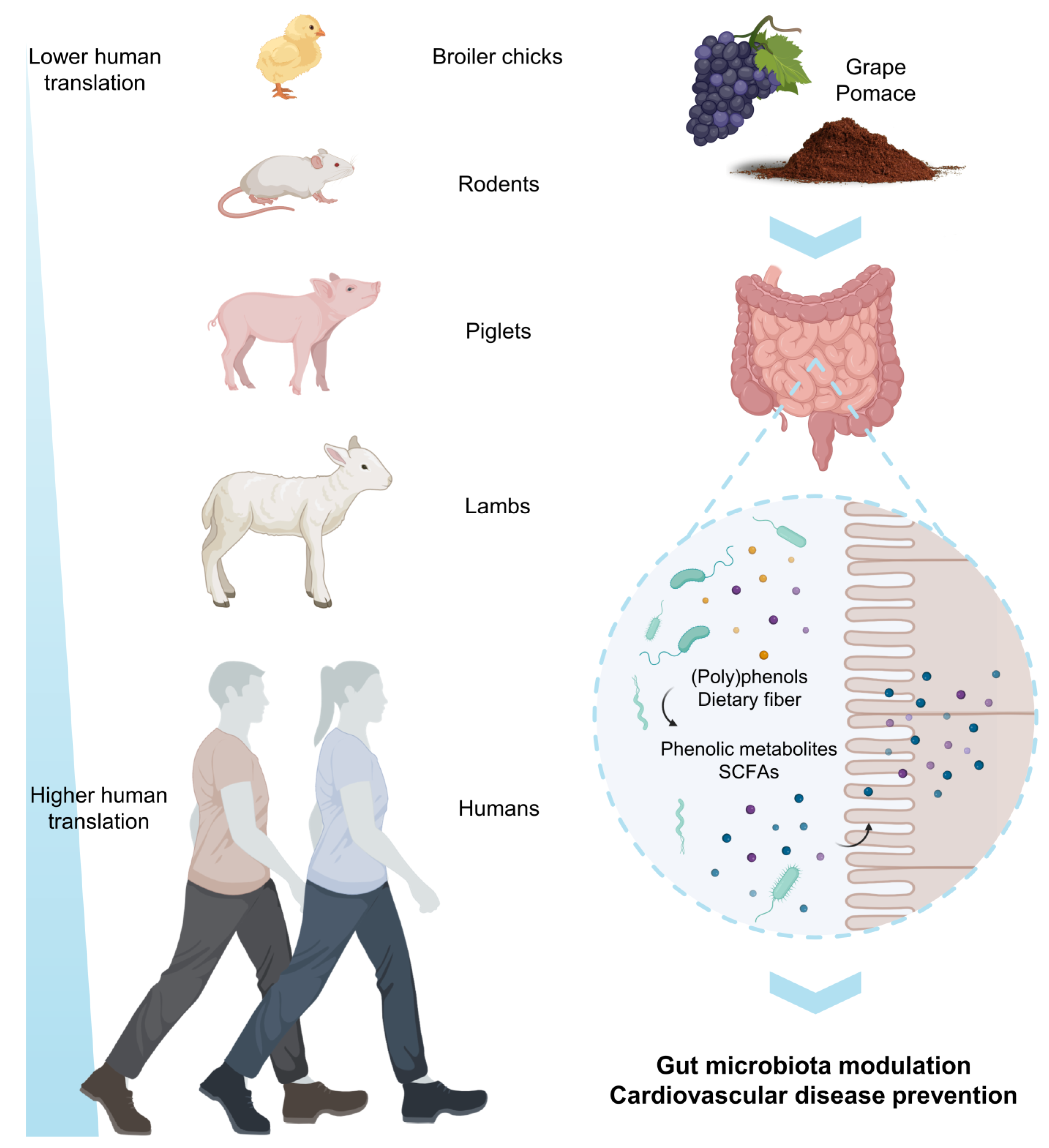{kind=link}
| RGPE | RGPE2 | ORGPE | WGPE | |
|---|---|---|---|---|
| Total (poly)phenols (mg/g) a | 48.0 ± 4.1 | 13.0 ± 2.3 | 5.7 ± 0.5 | 3.5 ± 0.5 |
| Phenolic acids (µg/g) a | 1843.0 ± 23.2 | 652.5 ± 12.6 | 853.7 ± 15.2 | 1211.8 ± 20.1 |
| Flavonols (µg/g) a | 72.7 ± 3.5 | nd | 87.0 ± 7.6 | nd |
| Flavan-3-ols (µg/g) a | 276.3 ± 7.6 | nd | 682.8 ± 8.7 | 295.2 ± 6.1 |
| Anthocyanins (µg/g) a | nd | nd | 1706.2 ± 21.7 | nd |
| AIR (mg/g) b | 930.5 ± 18.1 | 819.4 ± 18.9 | 969.8 ± 22.3 | 613.1 ± 21.8 |
| Glucose (mg/g AIR) b, c | 446.6 ± 2.1 | 270.4 ± 0.6 | 598.4 ± 4.5 | 839.3 ± 1.7 |
| Galactose (mg/g AIR) b, c | 28.8 ± 1.1 | 32 ± 0.7 | 36.9 ± 2.8 | 47.7 ± 1.1 |
| Mannose (mg/g AIR) b, c | 5.9 ± 0.3 | 17.4 ± 0.2 | 4 ± 0.2 | 2.7 ± 0.1 |
| Rhamnose (mg/g AIR) b, c | 3.2 ± 0.2 | 5.7 ± 0.2 | 2.1 ± 0.1 | 6.3 ± 0.1 |
| Uronic acids (mg/g AIR) b, c | 62.6 ± 4.2 | 44.1 ± 2.9 | 58.1 ± 4.8 | 52.2 ± 2.6 |
| Total sugars (mg/g AIR) | 555.9 | 374.3 | 702.3 | 969.1 |
| Klason lignin (mg/g AIR) | 161 ± 9.3 | 22.8 ± 3.6 | 118.4 ± 13.3 | 193.6 ± 26.8 |
| Nutrient | Effect of GP on Physical Impediment | Refs. | Effect of GP on Intestinal Enzymes | Refs. | Effect of GP on Nutrient Transport | Refs. |
|---|---|---|---|---|---|---|
| Carbohydrates | ↓ Glucose diffusion | [29] | α-amylase inhibition | [31,39,40,41,42,43,44,45] | SGLT1 downregulation | [52] |
| GLUT2 downregulation | [52,53,54] | |||||
| Starch digestibility | [27,28,30] | α-glucosidase inhibition | [28,31,40,42,43,44,45,46] | GLUT5 downregulation | [54] | |
| ↓ Intestinal glucose uptake | [24] | |||||
| Lipids | ↓ Triglyceride hydrolysis | [30] | Pancreatic lipase inhibition | [33,39,40,41,46] | CD36 downregulation | [57] |
| FABP1 downregulation | [57] | |||||
| Cholesterol | ↓ Micellar cholesterol | [33] | Cholesterol esterase inhibitor | [33] | NPC1L1 downregulation | [56] |
| ↓ Intestinal cholesterol uptake | [59] | |||||
| Bile salts | Binding | [33] | — | — | ASBT downregulation | [56,58] |
| Proteins | ↓ Protein hydrolysis | [30] | Trypsin inhibition Chymotrypsin inhibition | [40] | PEPT1 downregulation | [54] |
| Study Pl Design/Intervention | Microbiota-Related Measurements | Changes in Microbiota Composition | Relation to Other Biomarkers | Refs. |
|---|---|---|---|---|
| Broiler chicks | ||||
| Cobb broiler chicks; 1-day old n = 25 ♂ animals/group Duration: 21 days Diets:
| Plate counting 16S rRNA sequencing |
↓ Clostridium
↑ Lactobacillus ↑ Enterococcus ↑ Clostridium ↑ Bacterial diversity | =Weight gain
| [126] |
| Cobb-500 broiler chicks; 1-day old n = 192 animals/group Duration: 42 days Diets:
| 16S rRNA sequencing SCFAs by GC | ↑ Bacteroidetes ↓ Firmicutes ↑ Bacteroides ↑ Lactobacillus =SCFAs | ↑ Feed intake =Feed conversion ratio
↑ Villus height/crypt depth ratio
↑ Villus width
=Villus height/crypt depth ratio | [127] |
| Rodents (rats, mice) | ||||
| Wistar rats; n = 6 animals/group Duration: 14 months Diets:
| qPCR Urine phenolic metabolites by LC-QTOF | ↑ Bifidobacterium =Bacteroides =Clostridium leptum =Enterococcus ↓ Clostridium Cluster I | [128,129] | |
| C57BL/6J mice; 9-week old n = 14 animals/group Duration: 8 weeks Diets:
| 16S rRNA sequencing SCFAs by GC | ↓ Desulfovibrio ↓ Lactococcus ↑ Allobaculum ↑ Roseburia | ↓ Fat mass gain ↓ Adipose tissue inflammation =Food intake ↑ Glucose tolerance ↓ Insulin resistance index ↑ Antimicrobial peptides ↑ Tight junction proteins | [57] |
| C57BL/6J mice; 9-week old n = 10 ♂ animals/group Duration: 3-week antibiotics + 1 week diet Diets:
| 16S rRNA sequencing |
↑ Diversity ↑ Akkermansia | [130] | |
| Piglets | ||||
| Landrace × Large White–Duroc n = 12 animals/group Duration: 15 days Diets:
| Plate counting | ↑ Facultative probiotic bacteria ↑ Lactic acid bacteria ↓ Enterobacteriacae ↓ Campylobacter jejuni | ↑ Weight gain ↑ GSH ↑ H2O2 decomposition activity ↑ Total antioxidant capacity ↓ MDA (TBARS) ↓ Protein carbonyls | [105] |
| Songliao black pigs; 28-day old n = 6 animals/group Duration: 28 days Diets:
| 16S rRNA sequencing | ↑ Lactobacillus delbrueckii ↑ Olsenella umbonate ↑ Selenomonas bovis | =Growth
↑ Villus height/crypt depth ratio
=SCFAs receptors (GPR41/43) ↑ Serum IgG | [78] |
| Lambs | ||||
| Chios lambs; 15-day old n = 12 ♂ animals/group Duration: 55 days Diets:
| Plate counting | ↑ Facultative probiotic bacteria ↓ Enterobacteriacae ↓ Escherichia coli | ↑ CAT ↑ GSH ↓ MDA (TBARS) ↓ Protein carbonyls | [131] |
| Humans | ||||
| Healthy women (n = 10) Duration: 3 weeks Diet: 1.4 g/day of a red GP extract | qPCR SCFAs by GC-MS Phenolic metabolites by UPLC-ESI-MS/MS | =Faecal bacteria populations =Faecal/urine phenolic metabolites ↑ Fecal SCFAs | ↓ Blood fasting glucose Glucose metabolism-related miRNA modulation | [19] |
| ≥2 metabolic syndrome factors subjects (n = 49) Duration: 6 weeks Diet: 8 g/day of GP extract | qPCR SCFAs by GC | =Microbiota profile ↑ Lactobacilliales ↑ Bacteroides =SCFAs | ↑ insulin levels (responders) | [132] |
| Responders (n = 23) and non-responders (n = 26) to insulin reduction Duration: 6 weeks Diet: 8 g/day of GP extract | 16S rRNA sequencing Sequencing SCFAs by GC | ↓ Prevotella ↓ Firmicutes | ↑ miRNA-222 levels (responders) | [133] |
| High-risk cardiometabolic (n = 17) healthy (n = 12) subjects Duration: 6 weeks Diets:
| 16S rRNA sequencing SCFAs by GC-MS Phenolic metabolites by UPLC-ESI-MS/MS | =α and β diversity ↓ Peptoniphilus ↓ Clostridiaceae 1 ↓ Clostridium sensu stricto 1 ↓ Ezakiella ↓ Streptococcus ↓ Lachnospiraceae ND3007 ↓ Paraprevotella ↓ Senegalimassilia ↑ Streptococcaceae ↓ Eggerthellaceae ↓ Coribacteriales Incertae Sedis ↓ Propionic acid ↑ Protocatechuic acid | ↓ Blood pressure ↓ Fasting blood glucose Bacterial communities associated with cardiovascular and metabolic data | [16] |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Taladrid, D.; Rebollo-Hernanz, M.; Martin-Cabrejas, M.A.; Moreno-Arribas, M.V.; Bartolomé, B. Grape Pomace as a Cardiometabolic Health-Promoting Ingredient: Activity in the Intestinal Environment. Antioxidants 2023, 12, 979. https://doi.org/10.3390/antiox12040979
Taladrid D, Rebollo-Hernanz M, Martin-Cabrejas MA, Moreno-Arribas MV, Bartolomé B. Grape Pomace as a Cardiometabolic Health-Promoting Ingredient: Activity in the Intestinal Environment. Antioxidants. 2023; 12(4):979. https://doi.org/10.3390/antiox12040979
Chicago/Turabian StyleTaladrid, Diego, Miguel Rebollo-Hernanz, Maria A. Martin-Cabrejas, M. Victoria Moreno-Arribas, and Begoña Bartolomé. 2023. "Grape Pomace as a Cardiometabolic Health-Promoting Ingredient: Activity in the Intestinal Environment" Antioxidants 12, no. 4: 979. https://doi.org/10.3390/antiox12040979
APA StyleTaladrid, D., Rebollo-Hernanz, M., Martin-Cabrejas, M. A., Moreno-Arribas, M. V., & Bartolomé, B. (2023). Grape Pomace as a Cardiometabolic Health-Promoting Ingredient: Activity in the Intestinal Environment. Antioxidants, 12(4), 979. https://doi.org/10.3390/antiox12040979