
H2S-Enriched Flush out Does Not Increase Donor Organ Quality in a Porcine Kidney Perfusion Model

 , , ,
, , ,  and
and
Abstract
1. Introduction
2. Methods
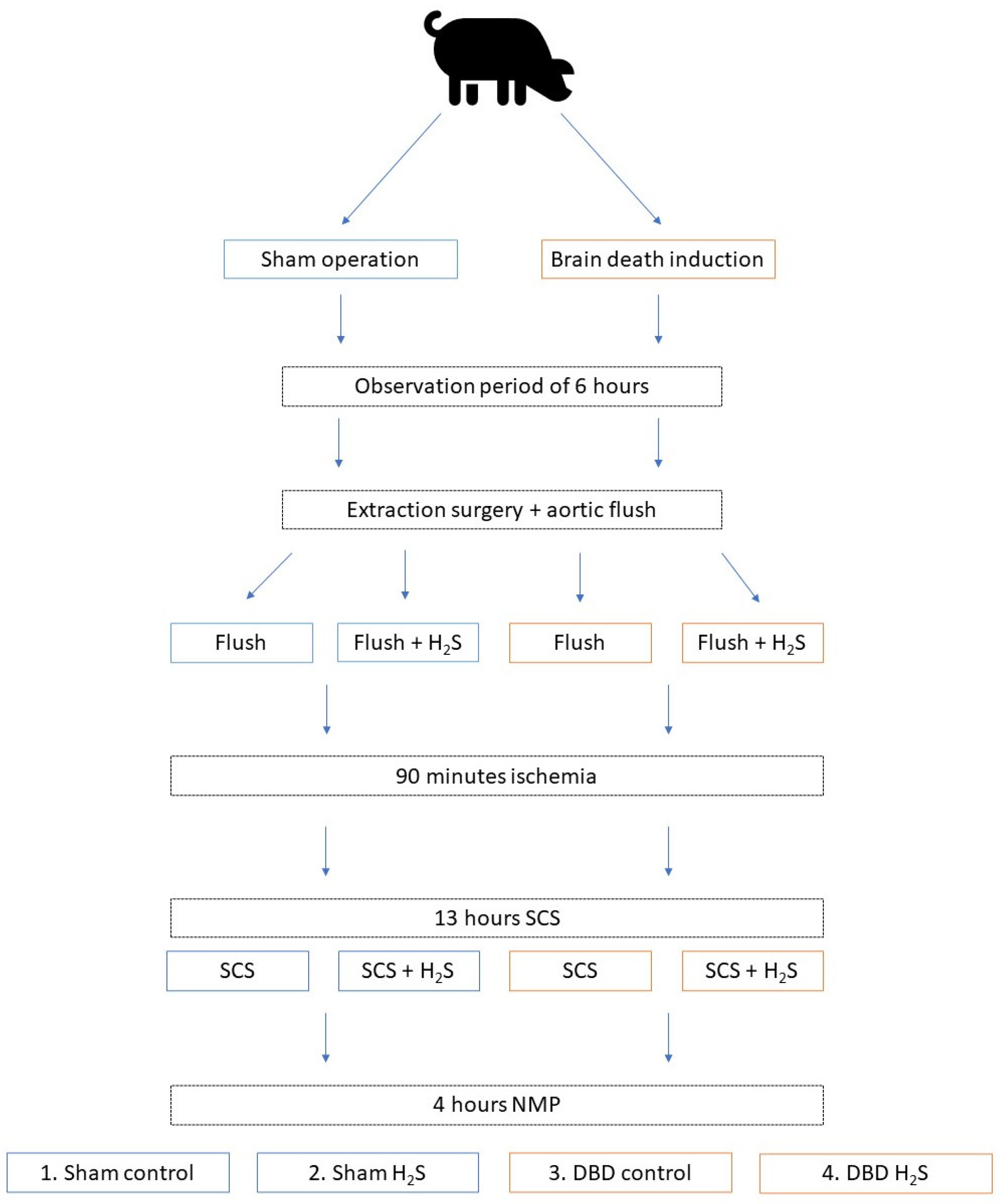
2.1. Study Design
2.2. Ethics and Animals
2.3. Anaesthesia and Induction of Brain Death
2.4. Organ Retrieval and Organ Flush
2.5. Normothermic Machine Perfusion
2.6. Flush Samples and Biopsies
2.7. Cytokines and Complement Activation Products
2.8. Renal Function and Injury Markers
2.9. Statistics
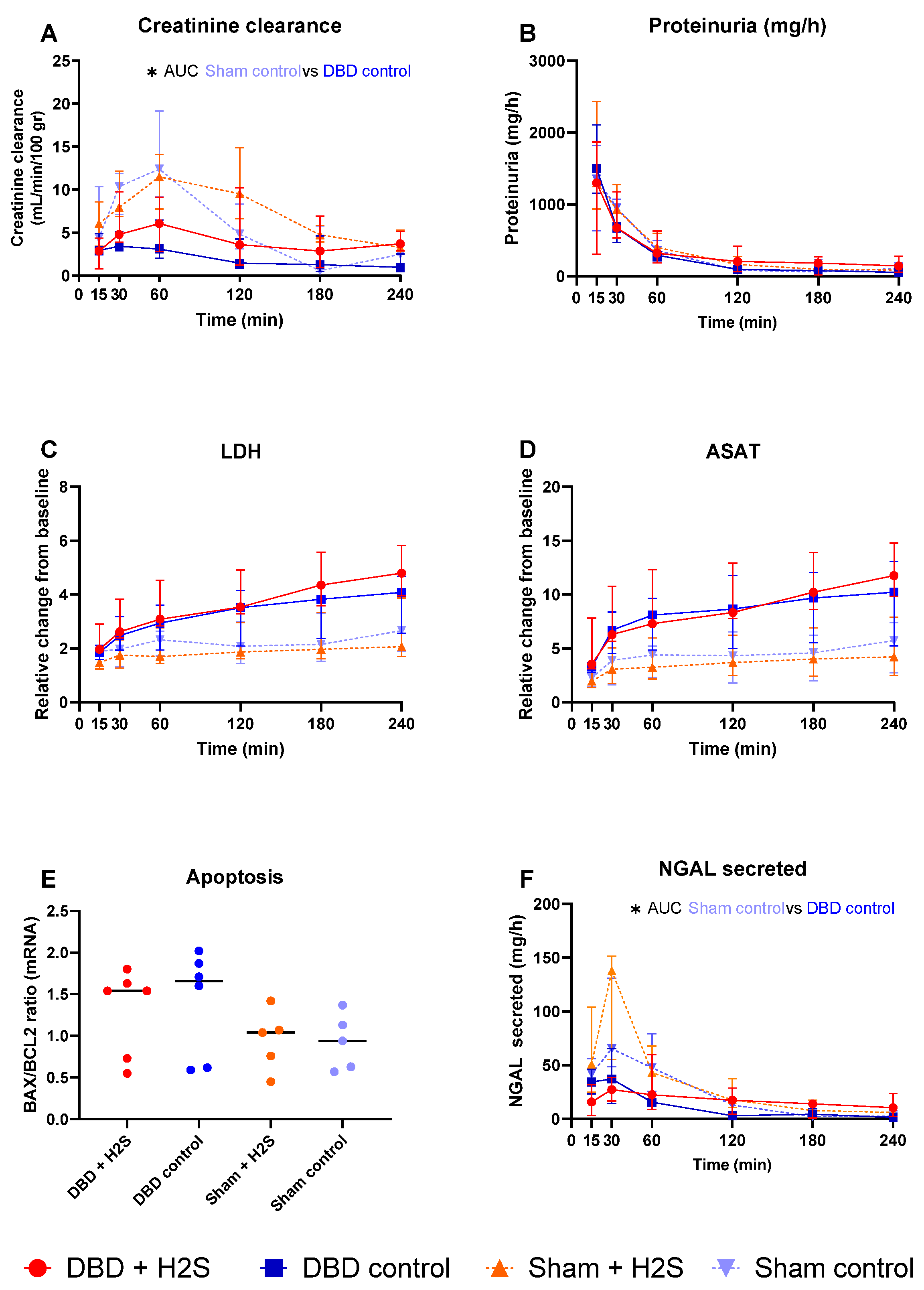
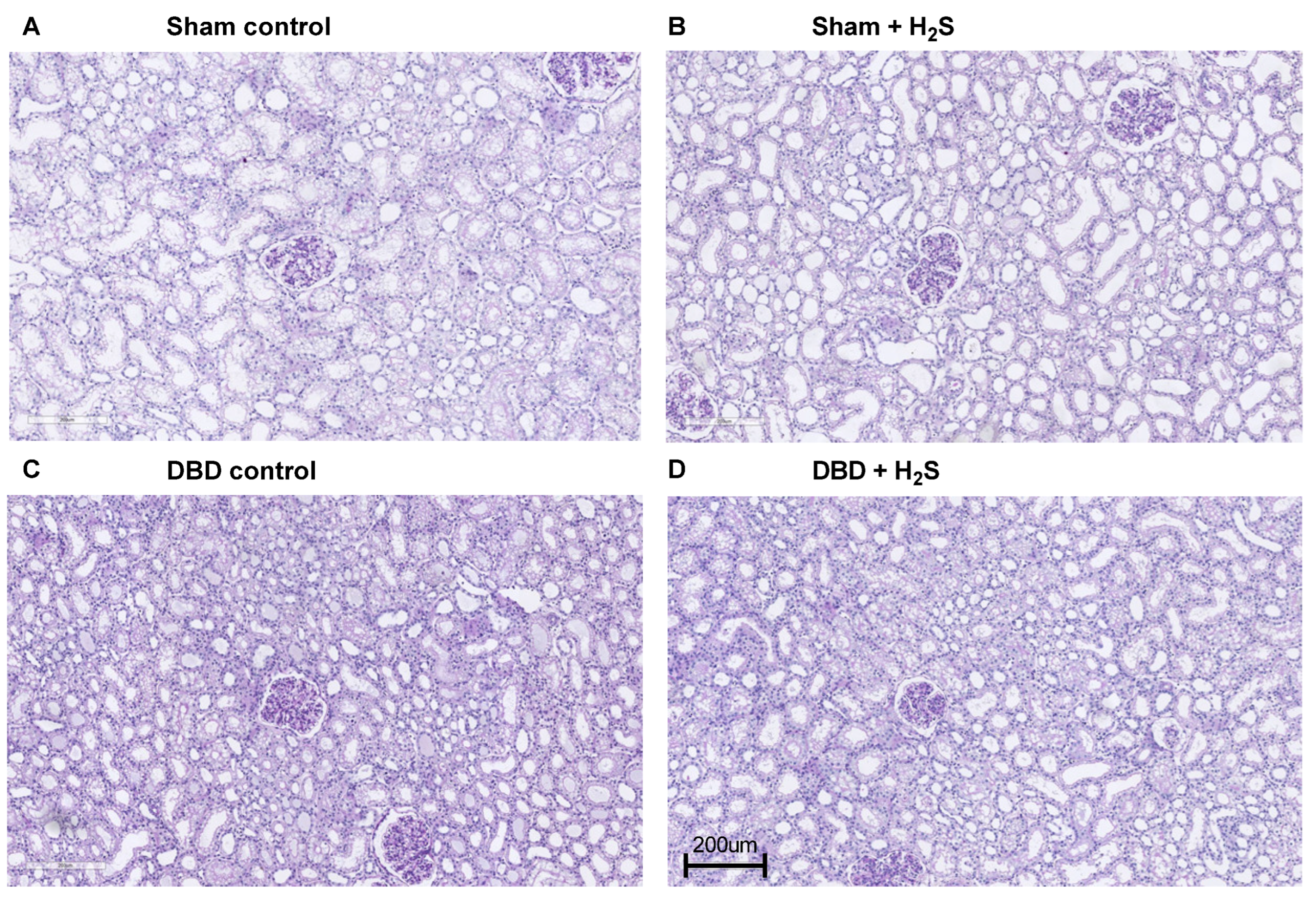
3. Results
3.1. Baseline Characteristics
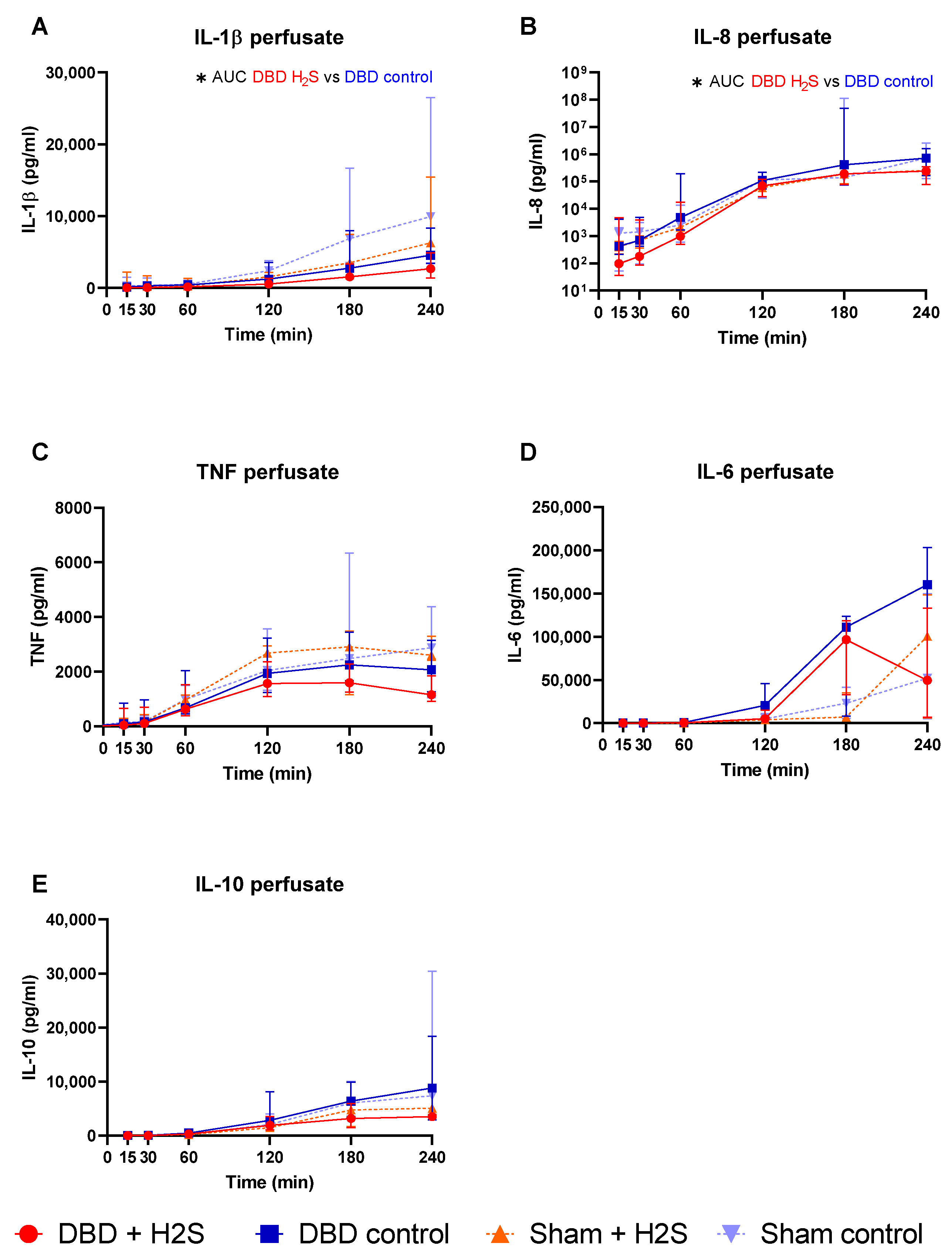
3.2. Cytokine Release during Preservation and NMP
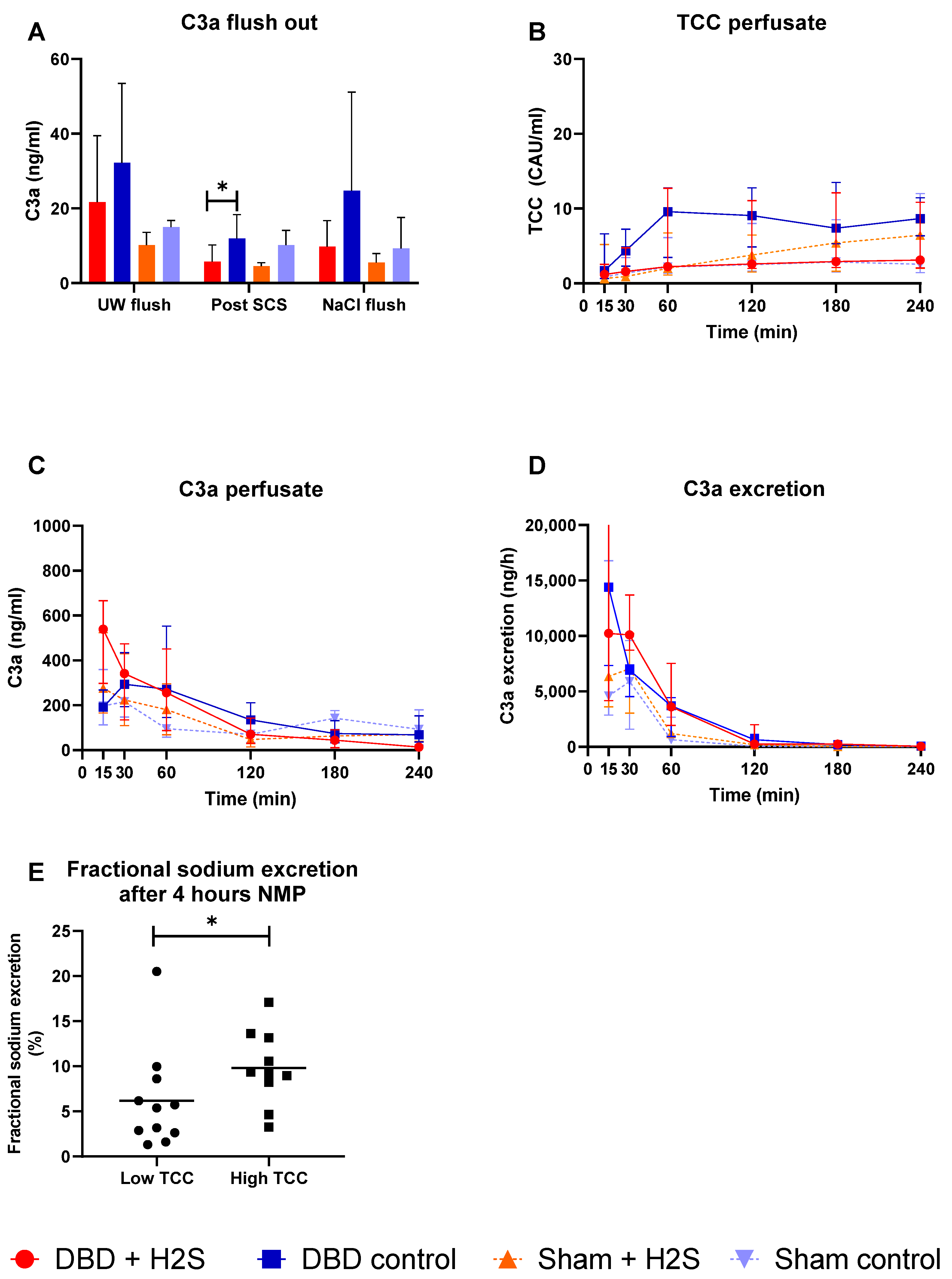
3.3. Complement Activation during Preservation and NMP
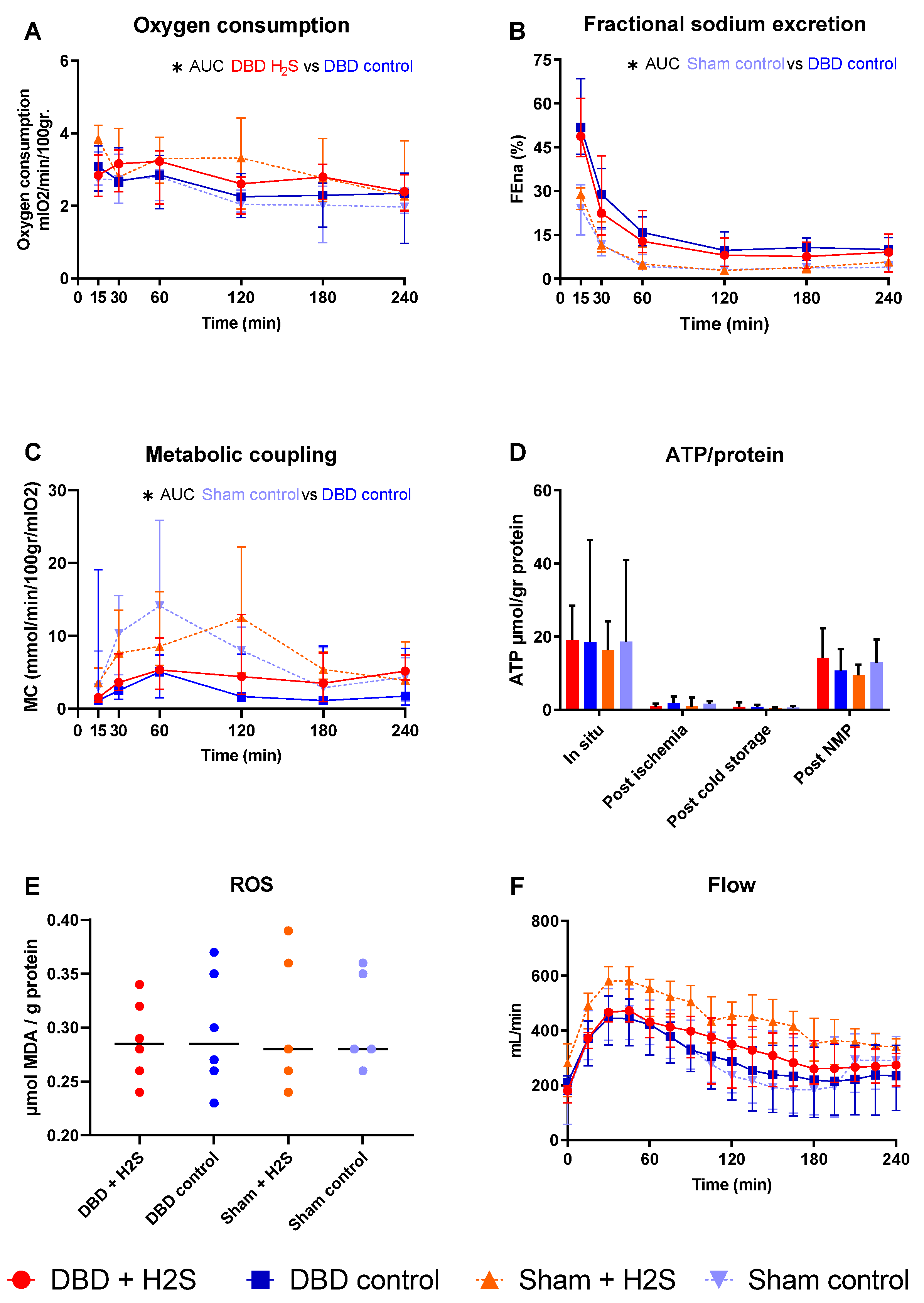
3.4. Metabolic Activity and ROS
3.5. Renal Function and Injury Markers
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Westendorp, W.H.; Leuvenink, H.G.; Ploeg, R.J. Brain death induced renal injury. Curr. Opin. Organ Transpl. 2011, 16, 151–156. [Google Scholar] [CrossRef] [PubMed]
- Bouma, H.R.; Ploeg, R.J.; Schuurs, T.A. Signal transduction pathways involved in brain death-induced renal injury. Am. J. Transpl. 2009, 9, 989–997. [Google Scholar] [CrossRef]
- Osband, A.J.; James, N.T.; Segev, D.L. Extraction Time of Kidneys From Deceased Donors and Impact on Outcomes. Am. J. Transpl. 2016, 16, 700–703. [Google Scholar] [CrossRef]
- Osband, A.J.; Zaki, R.F. Extraction time of kidneys during organ procurement impacts function. Clin. Transpl. 2011, 25, 235–238. [Google Scholar] [CrossRef]
- Heylen, L.; Pirenne, J.; Samuel, U.; Tieken, I.; Coemans, M.; Naesens, M.; Sprangers, B.; Jochmans, I. Effect of donor nephrectomy time during circulatory-dead donor kidney retrieval on transplant graft failure. Br. J. Surg. 2019, 107, 87–95. [Google Scholar] [CrossRef]
- Debout, A.; Foucher, Y.; Trébern-Launay, K.; Legendre, C.; Kreis, H.; Mourad, G.; Garrigue, V.; Morelon, E.; Buron, F.; Rostaing, L.; et al. Each additional hour of cold ischemia time significantly increases the risk of graft failure and mortality following renal transplantation. Kidney Int. 2015, 87, 343–349. [Google Scholar] [CrossRef]
- Tennankore, K.K.; Kim, S.J.; Alwayn, I.P.; Kiberd, B.A. Prolonged warm ischemia time is associated with graft failure and mortality after kidney transplantation. Kidney Int. 2016, 89, 648–658. [Google Scholar] [CrossRef]
- Pratschke, J.; Wilhelm, M.J.; Kusaka, M.; Beato, F.; Milford, E.L.; Hancock, W.W.; Tilney, N.L. Accelerated Rejection of Renal Allografts From Brain-Dead Donors. Ann. Surg. 2000, 232, 263–271. [Google Scholar] [CrossRef] [PubMed]
- Pratschke, J.; Wilhelm, M.J.; Laskowski, I.; Kusaka, M.; Beato, F.; Tullius, S.G.; Neuhaus, P.; Hancock, W.W.; Tilney, N.L. Influence of Donor Brain Death on Chronic Rejection of Renal Transplants in Rats. J. Am. Soc. Nephrol. 2001, 12, 2474–2481. [Google Scholar] [CrossRef]
- Terasaki, P.I.; Cecka, J.M.; Gjertson, D.W.; Takemoto, S. High survival rates of kidney transplants from spousal and living unrelated donors. N. Engl. J. Med. 1995, 333, 333–336. [Google Scholar] [CrossRef]
- van Erp, A.C.; van Dullemen, L.F.; Ploeg, R.J.; Leuvenink, H.G. Systematic review on the treatment of deceased organ donors. Transpl. Rev. 2018, 32, 194–206. [Google Scholar] [CrossRef]
- Hendriks, K.; Maassen, H.; van Dijk, P.R.; Henning, R.; van Goor, H.; Hillebrands, J.-L. Gasotransmitters in health and disease: A mitochondria-centered view. Curr. Opin. Pharmacol. 2019, 45, 87–93. [Google Scholar] [CrossRef]
- Koning, A.M.; Frenay, A.-R.S.; Leuvenink, H.G.D.; van Goor, H. Hydrogen sulfide in renal physiology, disease and transplantation—The smell of renal protection. Nitric Oxide 2015, 46, 37–49. [Google Scholar] [CrossRef]
- Maassen, H.; Hendriks, K.D.W.; Venema, L.H.; Henning, R.H.; Hofker, S.H.; Van Goor, H.; Leuvenink, H.G.D.; Coester, A.M. Hydrogen sulphide-induced hypometabolism in human-sized porcine kidneys. PLoS ONE 2019, 14, e0225152. [Google Scholar] [CrossRef]
- Lobb, I.; Jiang, J.; Lian, D.; Liu, W.; Haig, A.; Saha, M.N.; Torregrossa, R.; Wood, M.E.; Whiteman, M.; Sener, A. Hydrogen Sulfide Protects Renal Grafts Against Prolonged Cold Ischemia–Reperfusion Injury via Specific Mitochondrial Actions. Am. J. Transpl. 2017, 17, 341–352. [Google Scholar] [CrossRef]
- Dirkes, M.C.; Milstein, D.M.; Heger, M.; van Gulik, T.M. Absence of Hydrogen Sulfide-Induced Hypometabolism in Pigs: A Mechanistic Explanation in Relation to Small Nonhibernating Mammals. Eur. Surg. Res. 2015, 54, 178–191. [Google Scholar] [CrossRef] [PubMed]
- Haouzi, P.; Notet, V.; Chenuel, B.; Chalon, B.; Sponne, I.; Ogier, V.; Bihain, B. H2S induced hypometabolism in mice is missing in sedated sheep. Respir. Physiol. Neurobiol. 2008, 160, 109–115. [Google Scholar] [CrossRef]
- Chouchani, E.T.; Pell, V.R.; Gaude, E.; Aksentijević, D.; Sundier, S.Y.; Robb, E.L.; Logan, A.; Nadtochiy, S.M.; Ord, E.N.J.; Smith, A.C.; et al. Ischaemic accumulation of succinate controls reperfusion injury through mitochondrial ROS. Nature 2014, 515, 431–435. [Google Scholar] [CrossRef] [PubMed]
- Hamelink, T.L.; Ogurlu, B.; De Beule, J.; Lantinga, V.A.; Pool, M.B.F.; Venema, L.H.; Leuvenink, H.G.D.; Jochmans, I.; Moers, C. Renal Normothermic Machine Perfusion: The Road Toward Clinical Implementation of a Promising Pretransplant Organ Assessment Tool. Transplantation 2021. ahead of print. [Google Scholar] [CrossRef]
- Barklin, A.; Larsson, A.; Vestergaard, C.; Koefoed-Nielsen, J.; Bach, A.; Nyboe, R.; Wogensen, L.; Tønnesen, E. Does brain death induce a pro-inflammatory response at the organ level in a porcine model? Acta Anaesthesiol. Scand. 2008, 52, 621–627. [Google Scholar] [CrossRef] [PubMed]
- Lohmann, S.; Pool, M.B.; Rozenberg, K.M.; Keller, A.K.; Moers, C.; Møldrup, U.; Møller, B.K.; Lignell, S.J.; Krag, S.; Sierra-Parraga, J.M.; et al. Mesenchymal stromal cell treatment of donor kidneys during ex vivo normothermic machine perfusion: A porcine renal autotransplantation study. Am. J. Transpl. 2020, 21, 2348–2359. [Google Scholar] [CrossRef]
- Høgåsen, K.; Jansen, J.; Mollnes, T.; Harboe, M. Extensive complement activation in hereditary porcine membranoproliferative glomerulonephritis type II (porcine dense deposit disease). Mol. Immunol. 1993, 30, 17. [Google Scholar] [CrossRef]
- Nilsson, P.H.; Pettersen, K.; Oppermann, M.; Skjeflo, E.W.; Fure, H.; Christiansen, D.; Mollnes, T.E. Quantification of Porcine Complement Activation Fragment C3a by a Neoepitope-Based Enzyme-Linked Immunosorbent Assay. Methods Mol. Biol. 2021, 2227, 51–59. [Google Scholar] [CrossRef]
- Ueland, N.L.; Ludvigsen, J.K.; Hellerud, B.C.; Mollnes, T.E.; Skjeflo, E.W. Choice of immunoassay to evaluate porcine cytokine levels. Veter-Immunol. Immunopathol. 2020, 230, 110129. [Google Scholar] [CrossRef]
- Venema, L.H.; Brat, A.; Moers, C.; Hart, N.A.; Ploeg, R.J.; Hannaert, P.; Minor, T.; Leuvenink, H. Effects of Oxygen During Long-term Hypothermic Machine Perfusion in a Porcine Model of Kidney Donation After Circulatory Death. Transplantation 2019, 103, 2057–2064. [Google Scholar] [CrossRef]
- Mahboub, P.; Ottens, P.; Seelen, M.; t Hart, N.; Van Goor, H.; Ploeg, R.; Martins, P.N.; Leuvenink, H. Gradual rewarming with gradual increase in pressure during machine perfusion after cold static preservation reduces kidney ischemia reperfusion injury. PLoS ONE 2015, 10, e0143859. [Google Scholar] [CrossRef]
- van Erp, A.C.; Qi, H.; Jespersen, N.R.; Hjortbak, M.V.; Ottens, P.J.; Wiersema-Buist, J.; Nørregaard, R.; Pedersen, M.; Laustsen, C.; Leuvenink, H.G.D.; et al. Organ-specific metabolic profiles of the liver and kidney during brain death and afterwards during normothermic machine perfusion of the kidney. Am. J. Transpl. 2020, 20, 2425–2436. [Google Scholar] [CrossRef]
- Rebolledo, R.A.; Liu, B.; Akhtar, M.Z.; Ottens, P.J.; Zhang, J.-N.; Ploeg, R.J.; Leuvenink, H.G.D. Steroid Anti-Inflammatory Effects Did Not Improve Organ Quality in Brain-Dead Rats. BioMed Res. Int. 2015, 2015, 207534. [Google Scholar] [CrossRef]
- Sivarajah, A.; Collino, M.; Yasin, M.; Benetti, E.; Gallicchio, M.; Mazzon, E.; Cuzzocrea, S.; Fantozzi, R.; Thiemermann, C. Anti-apoptotic and anti-inflammatory effects of hydrogen sulfide in a rat model of regional myocardial I/R. Shock 2009, 31, 267–274. [Google Scholar] [CrossRef]
- Sun, X.; Wang, W.; Dai, J.; Huang, J.; Shi, M.; Chu, X.; Wang, F.; Guo, C.; Wang, C.; Pang, L.; et al. Donor heart preservation with a novel long-term and slow-releasing hydrogen sulfide system. Nitric Oxide 2018, 81, 1–10. [Google Scholar] [CrossRef]
- Chen, X.; Xu, W.; Wang, Y.; Luo, H.; Quan, S.; Zhou, J.; Yang, N.; Zhang, T.; Wu, L.; Liu, J.; et al. Hydrogen sulfide reduces kidney injury due to urinary-derived sepsis by inhibiting NF-κB expression, decreasing TNF-α levels and increasing IL-10 levels. Exp. Ther. Med. 2014, 8, 464–470. [Google Scholar] [CrossRef] [PubMed]
- Duque, G.A.; Descoteaux, A. Macrophage Cytokines: Involvement in Immunity and Infectious Diseases. Front. Immunol. 2014, 5, 491. [Google Scholar] [CrossRef] [PubMed]
- Kusaka, M.; Pratschke, J.; Wilhelm, M.J.; Ziai, F.; Zandi-Nejad, K.; Mackenzie, H.S.; Hancock, W.W.; Tilney, N.L. Activation of inflammatory mediators in rat renal isografts by donor brain death. Transplantation 2000, 69, 405–410. [Google Scholar] [CrossRef]
- Stone, J.P.; Ball, A.L.; Critchley, W.R.; Major, T.; Edge, R.J.; Amin, K.; Clancy, M.J.; Fildes, J.E. Ex Vivo Normothermic Perfusion Induces Donor-Derived Leukocyte Mobilization and Removal Prior to Renal Transplantation. Kidney Int. Rep. 2016, 1, 230–239. [Google Scholar] [CrossRef]
- Nieuwenhuijs-Moeke, G.J.; Pischke, S.E.; Berger, S.P.; Sanders, J.S.F.; Pol, R.A.; Struys, M.M.R.F.; Ploeg, R.J.; Leuvenink, H.G.D. Ischemia and Reperfusion Injury in Kidney Transplantation: Relevant Mechanisms in Injury and Repair. J. Clin. Med. 2020, 9, 253. [Google Scholar] [CrossRef]
- Nauser, C.L.; Farrar, C.A.; Sacks, S.H. Complement Recognition Pathways in Renal Transplantation. J. Am. Soc. Nephrol. 2017, 28, 2571–2578. [Google Scholar] [CrossRef]
- Damman, J.; Seelen, M.A.; Moers, C.; Daha, M.R.; Rahmel, A.; Leuvenink, H.G.; Paul, A.; Pirenne, J.; Ploeg, R.J. Systemic Complement Activation in Deceased Donors Is Associated With Acute Rejection After Renal Transplantation in the Recipient. Transplantation 2011, 92, 163–169. [Google Scholar] [CrossRef]
- Trouw, L.A.; Bengtsson, A.A.; Gelderman, K.A.; Dahlbäck, B.; Sturfelt, G.; Blom, A.M. C4b-binding Protein and Factor H Compensate for the Loss of Membrane-bound Complement Inhibitors to Protect Apoptotic Cells against Excessive Complement Attack. J. Biol. Chem. 2007, 282, 28540–28548. [Google Scholar] [CrossRef]
- Jager, N.M.; Poppelaars, F.; Daha, M.R.; Seelen, M.A. Complement in renal transplantation: The road to translation. Mol. Immunol. 2017, 89, 22–35. [Google Scholar] [CrossRef]
- Gerritsma, J.S.; van Kooten, C.; Gerritsen, A.F.; van Es, L.A.; Daha, M.R. Transforming growth factor-β1 regulates chemokine and complement production by human proximal tubular epithelial cells. Kidney Int. 1998, 53, 609–616. [Google Scholar] [CrossRef]
- Poppelaars, F.; Jager, N.M.; Kotimaa, J.; Leuvenink, H.G.; Daha, M.R.; van Kooten, C.; Seelen, M.A.; Damman, J. C1-Inhibitor Treatment Decreases Renal Injury in an Established Brain-Dead Rat Model. Transplantation 2018, 102, 79–87. [Google Scholar] [CrossRef]
- Blackstone, E.; Morrison, M.; Roth, M.B. H2S Induces a Suspended Animation-Like State in Mice. Science 2005, 308, 518. [Google Scholar] [CrossRef]
- Cooper, C.E.; Brown, G.C. The inhibition of mitochondrial cytochrome oxidase by the gases carbon monoxide, nitric oxide, hydrogen cyanide and hydrogen sulfide: Chemical mechanism and physiological significance. J. Bioenerg. Biomembr. 2008, 40, 533–539. [Google Scholar] [CrossRef]
- Snijder, P.M.; De Boer, R.A.; Bos, E.M.; van den Born, J.C.; Ruifrok, W.-P.T.; Vreeswijk-Baudoin, I.; Van Dijk, M.C.R.F.; Hillebrands, J.-L.; Leuvenink, H.G.; Van Goor, H. Gaseous Hydrogen Sulfide Protects against Myocardial Ischemia-Reperfusion Injury in Mice Partially Independent from Hypometabolism. PLoS ONE 2013, 8, e63291. [Google Scholar] [CrossRef]
- Hosgood, S.A.; Nicholson, M.L. Hydrogen sulphide ameliorates ischaemia-reperfusion injury in an experimental model of non-heart-beating donor kidney transplantation. Br. J. Surg. 2010, 97, 202–209. [Google Scholar] [CrossRef]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Gene | Primers | Amplicon Size (bp) | Gene Accession Number |
|---|---|---|---|
| b-act | TCTGCGCAAGTTAGGTTTTGTC CGTCCACCGCAAATGCTT | 78 | AY_550069.1 |
| Bax BCL-2 | CAGACGGTGACCATCTTCGTG TTCTTCCAGATGGTGAGCGAG AATTACCATCGGCGTAGTGCA TCTAGAGCCCTTGACCTCCATCT | 58 82 | XM_003127290.5 XM_021099593.1 |
| In Situ | DBD Kidneys n = 12 (6 Control, 6 H2S) | Sham Kidneys n = 10 (5 Control, 5 H2S) | p-Value |
|---|---|---|---|
| Weight pigs (kg) | 63.5 (3.4) | 62 (3.78) | 0.689 |
| Time under anaestesia (hours) | 11:15 (0:26) | 11:08 (0:22) | 0.180 |
| Surgery time (hours) | 3:06 (0:25) | 3:15 (0:17) | 0.233 |
| Urine production pigs (mL) | 4213 (890) | 874 (214.8) | <0.001 |
| Post Nephrectomy | |||
| Flush time (min) | 18 (7.1) | 16.5 (5.7) | 0.354 |
| Cold ischemic time (min) | 786 (7.3) | 785 (11.2) | 0.974 |
| Warm ischemic time (min) | 1.5 (1.5) | 1 (1.1) | 0.964 |
| Weight kidneys (gr) | 145 (9.5) | 143 (7.9) | 0.974 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Maassen, H.; Venema, L.H.; Weiss, M.G.; Huijink, T.M.; Hofker, H.S.; Keller, A.K.; Mollnes, T.E.; Eijken, M.; Pischke, S.E.; Jespersen, B.; et al. H2S-Enriched Flush out Does Not Increase Donor Organ Quality in a Porcine Kidney Perfusion Model. Antioxidants 2023, 12, 749. https://doi.org/10.3390/antiox12030749
Maassen H, Venema LH, Weiss MG, Huijink TM, Hofker HS, Keller AK, Mollnes TE, Eijken M, Pischke SE, Jespersen B, et al. H2S-Enriched Flush out Does Not Increase Donor Organ Quality in a Porcine Kidney Perfusion Model. Antioxidants. 2023; 12(3):749. https://doi.org/10.3390/antiox12030749
Chicago/Turabian StyleMaassen, Hanno, Leonie H. Venema, Marc G. Weiss, Tobias M. Huijink, H. Sijbrand Hofker, Anna K. Keller, Tom E. Mollnes, Marco Eijken, Søren E. Pischke, Bente Jespersen, and et al. 2023. "H2S-Enriched Flush out Does Not Increase Donor Organ Quality in a Porcine Kidney Perfusion Model" Antioxidants 12, no. 3: 749. https://doi.org/10.3390/antiox12030749
APA StyleMaassen, H., Venema, L. H., Weiss, M. G., Huijink, T. M., Hofker, H. S., Keller, A. K., Mollnes, T. E., Eijken, M., Pischke, S. E., Jespersen, B., van Goor, H., & Leuvenink, H. G. D. (2023). H2S-Enriched Flush out Does Not Increase Donor Organ Quality in a Porcine Kidney Perfusion Model. Antioxidants, 12(3), 749. https://doi.org/10.3390/antiox12030749