
Effect of Astaxanthin on Tissue Transglutaminase and Cytoskeletal Protein Expression in Amyloid-Beta Stressed Olfactory Ensheathing Cells: Molecular and Delayed Luminescence Studies
 , , , and
, , , and 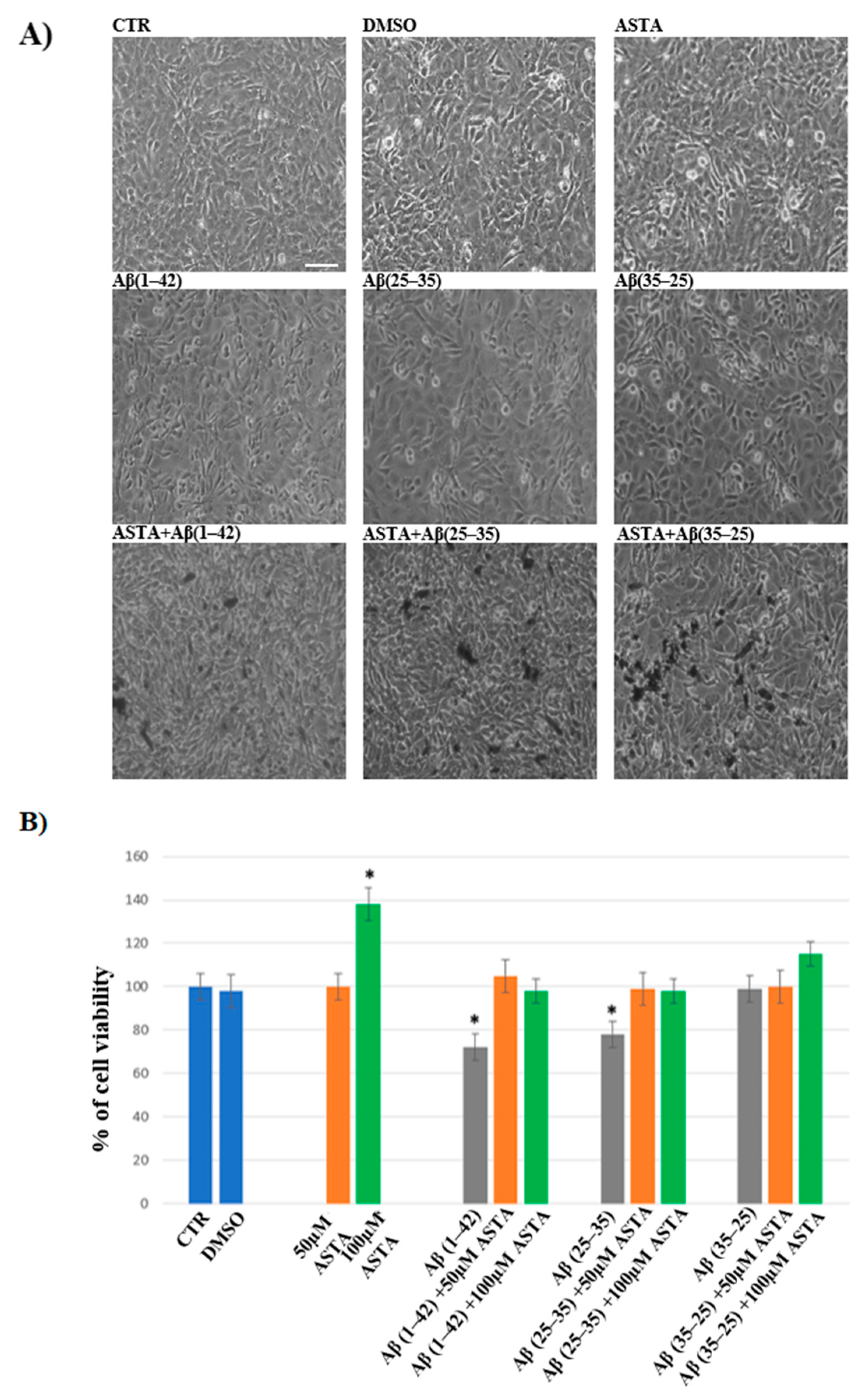{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
1. Introduction
2. Materials and Methods
2.1. Materials
2.2. Methods
2.2.1. OEC Cultures
2.2.2. Treatment of OECs
2.2.3. MTT Test
2.2.4. Total ROS
2.2.5. Immunocytochemistry and CLSM Analysis
2.2.6. Isolation of Total Protein and Western Blot Analysis
2.2.7. Delayed Luminescence Spectroscopy
2.2.8. Statistical Analysis
3. Results
3.1. Cell Viability
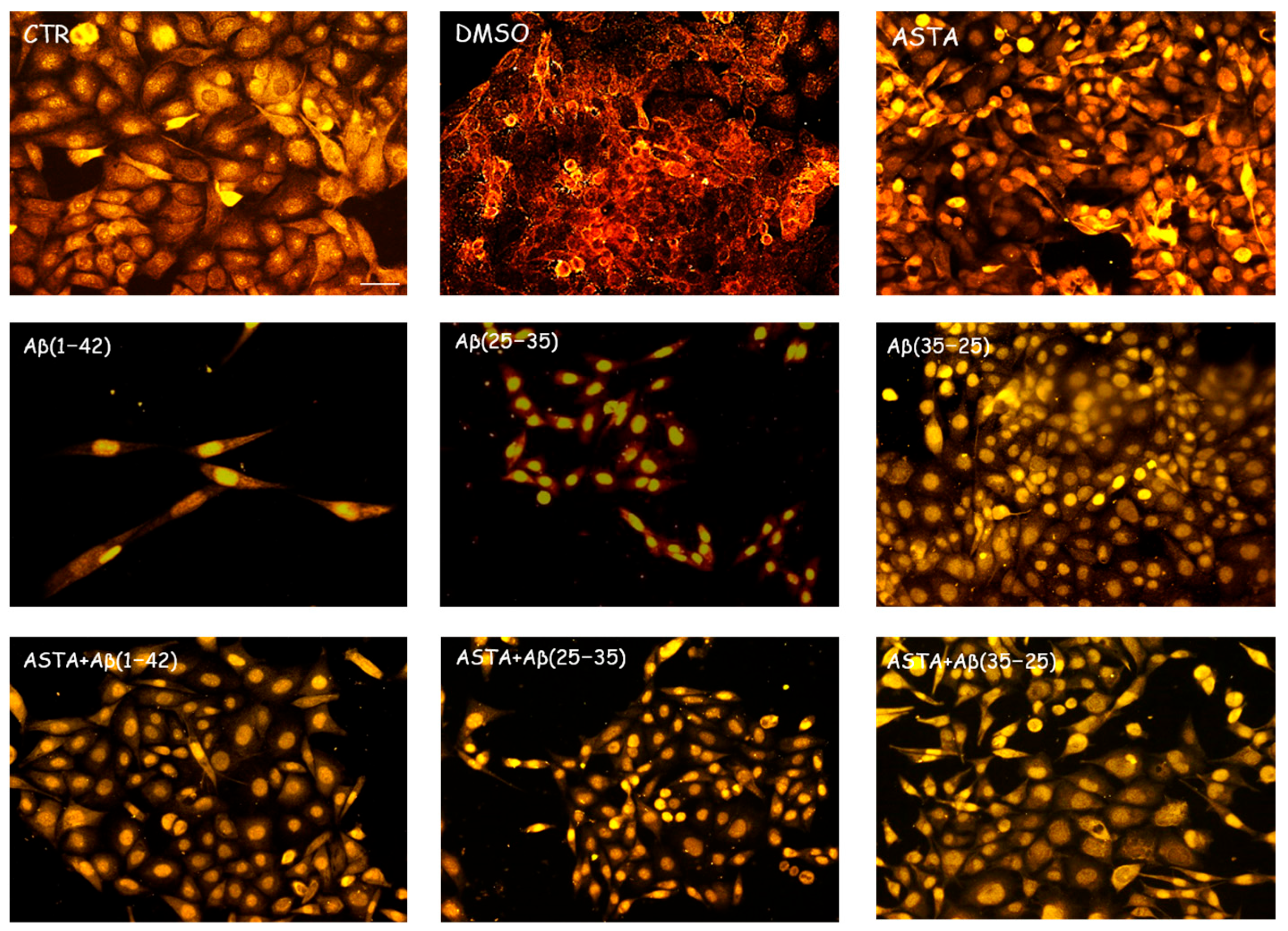
3.2. Expression of Markers by Immunofluorescence
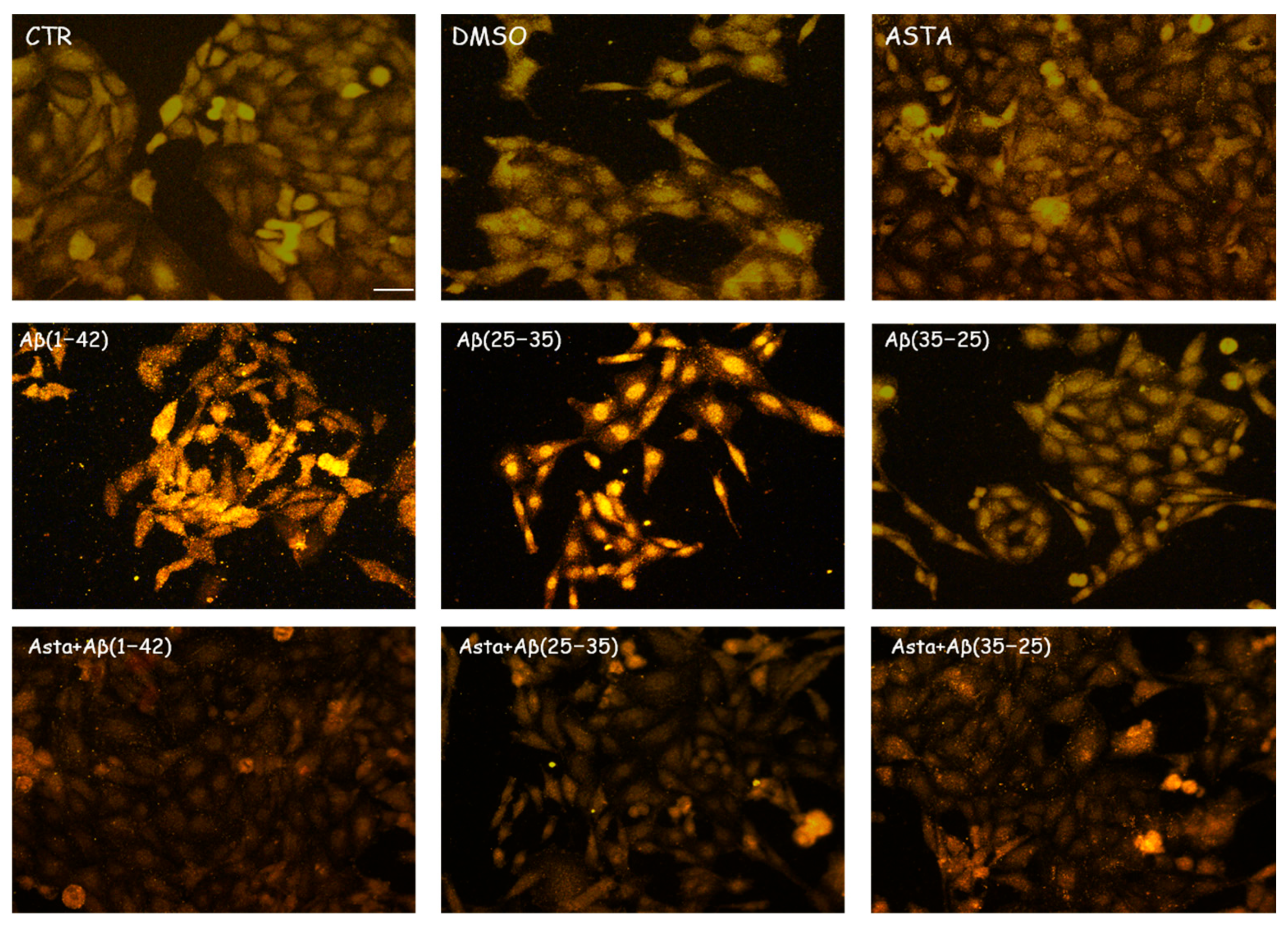
3.2.1. GFAP and Vimentin Expression
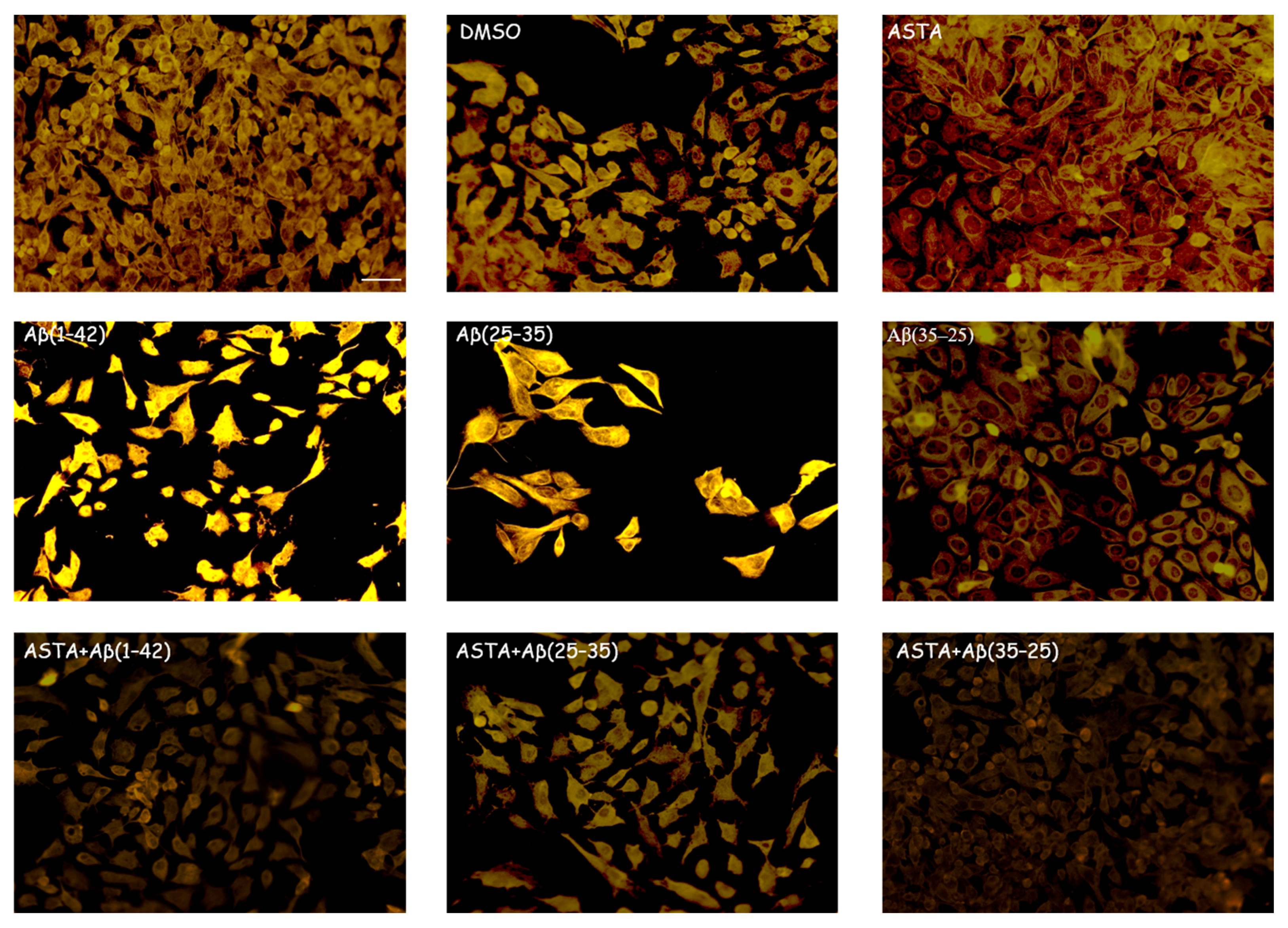
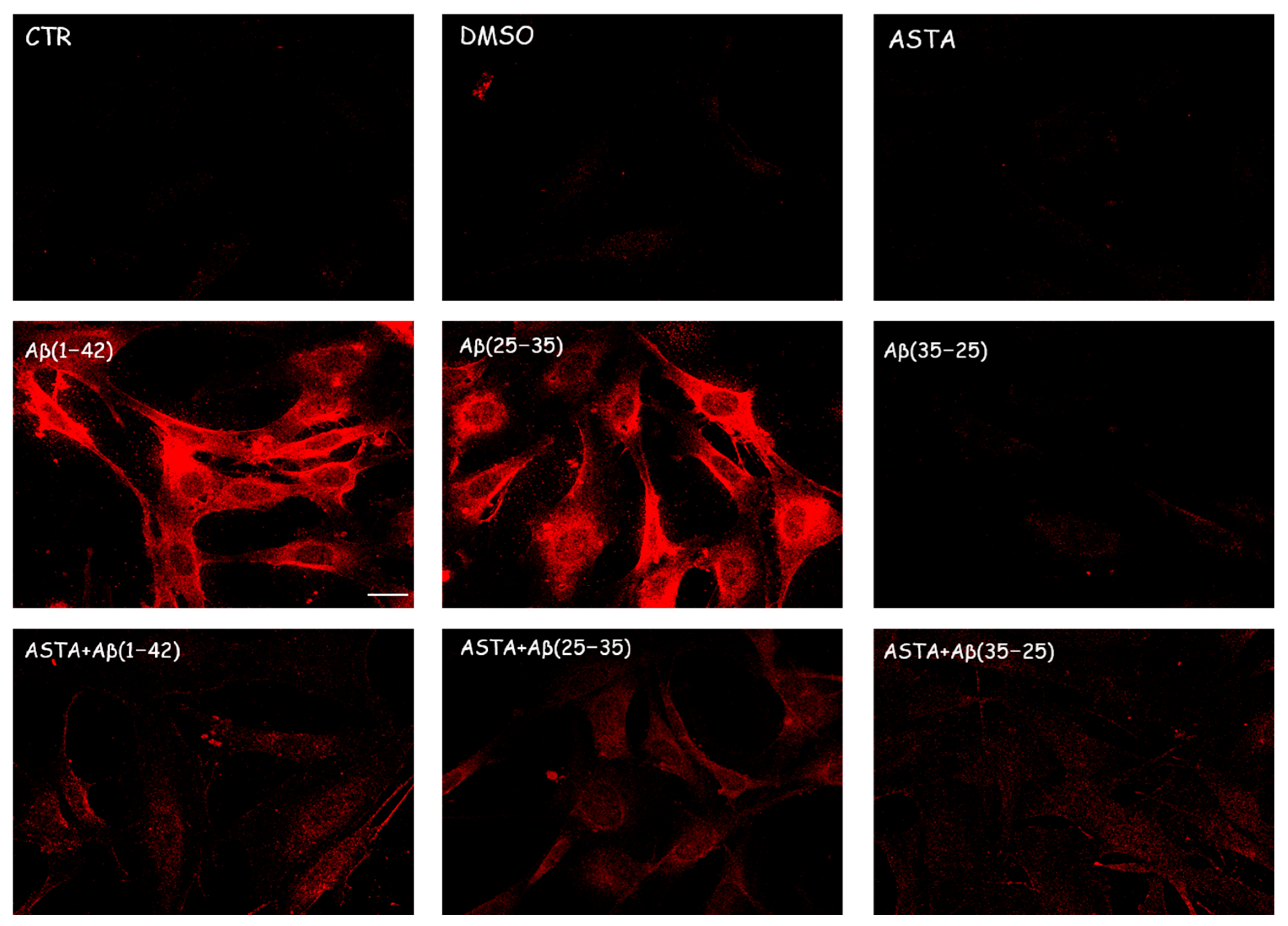
3.2.2. TG2 Expression
3.2.3. Nestin Expression
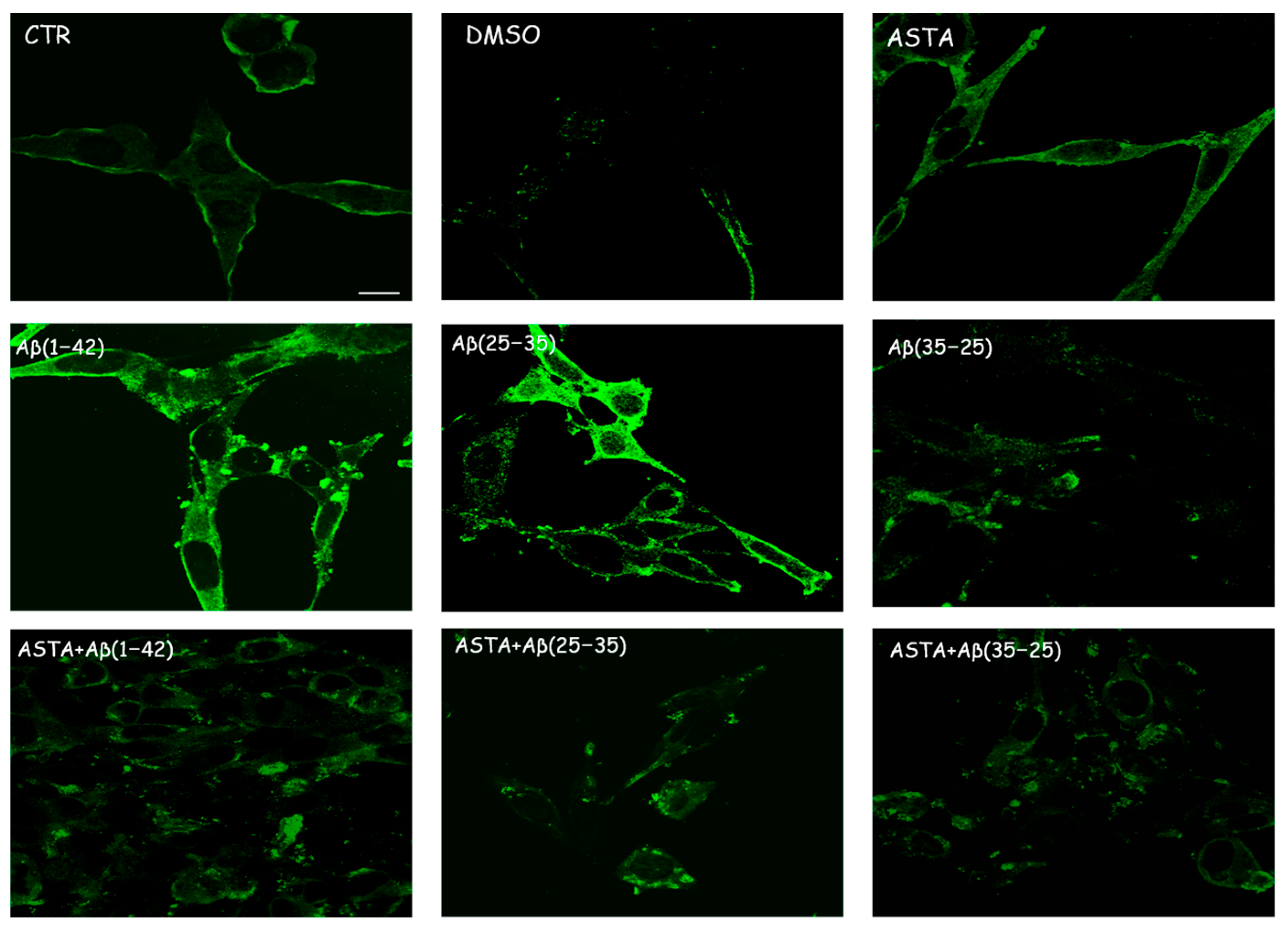
3.2.4. Caspase-3 Cleavage Immunolabeling
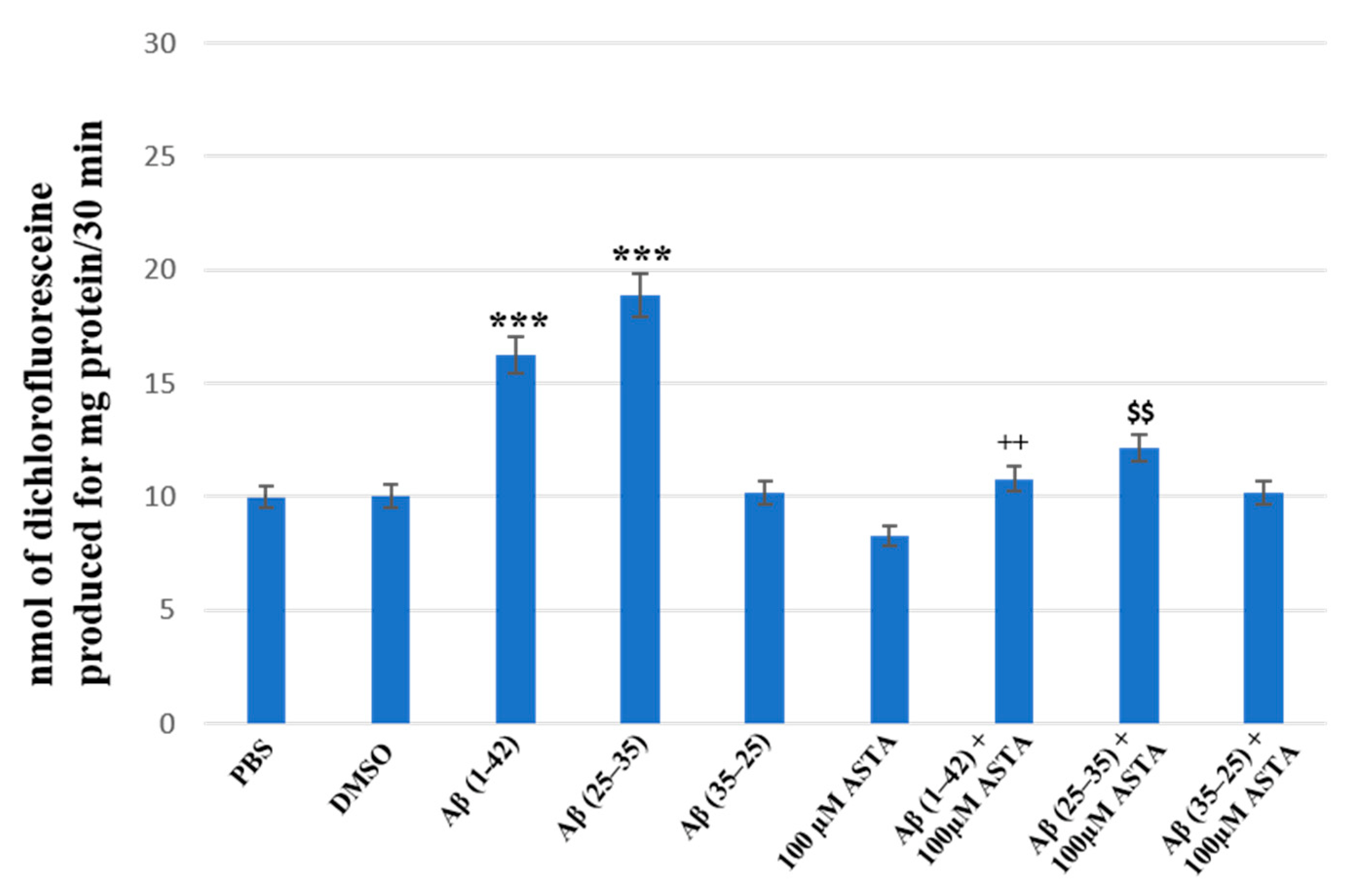
3.3. Total ROS Generation
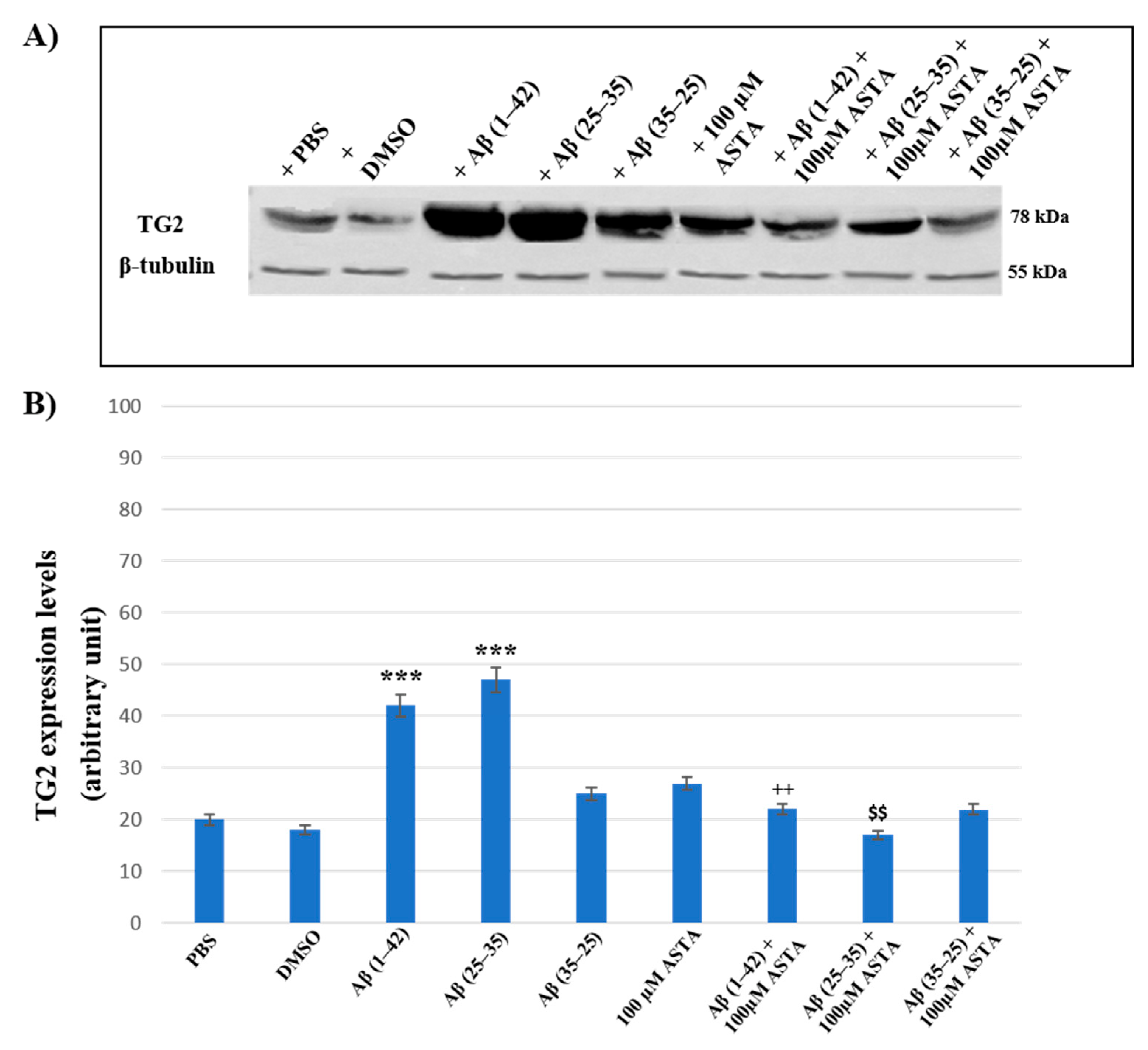
3.4. TG2 Expression Levels Immunolabeling
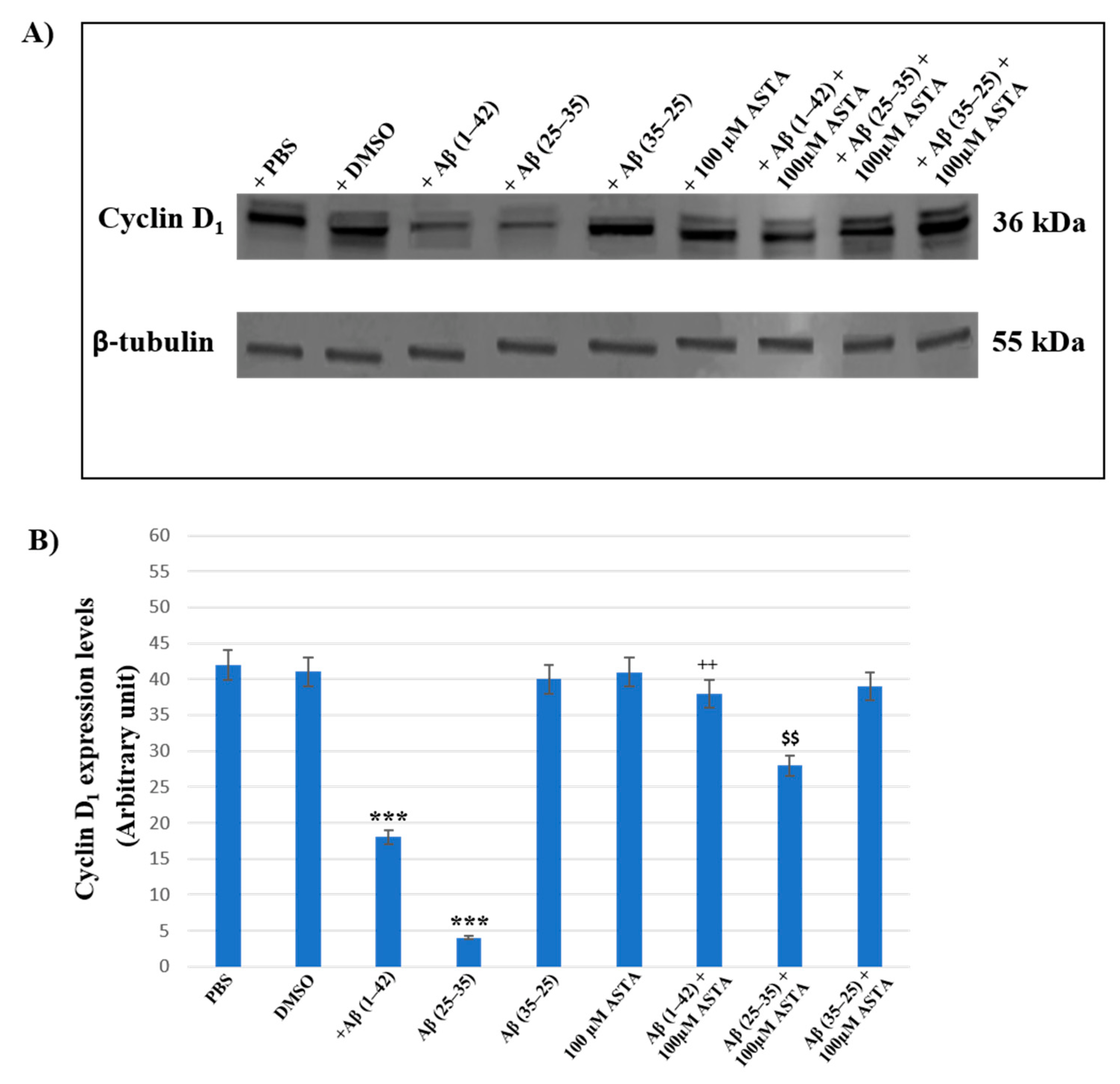
3.5. Cyclin D1 Expression Levels
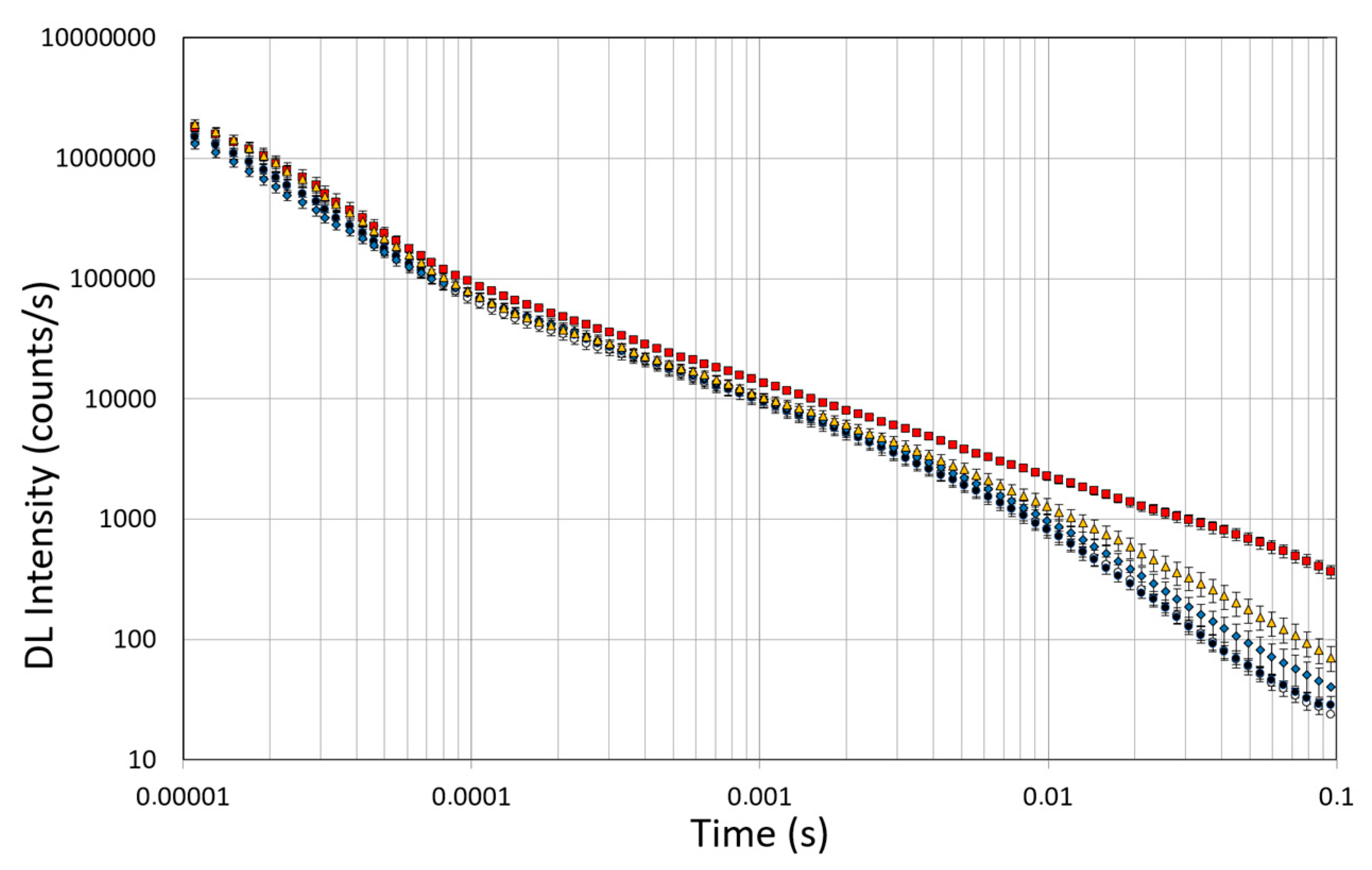
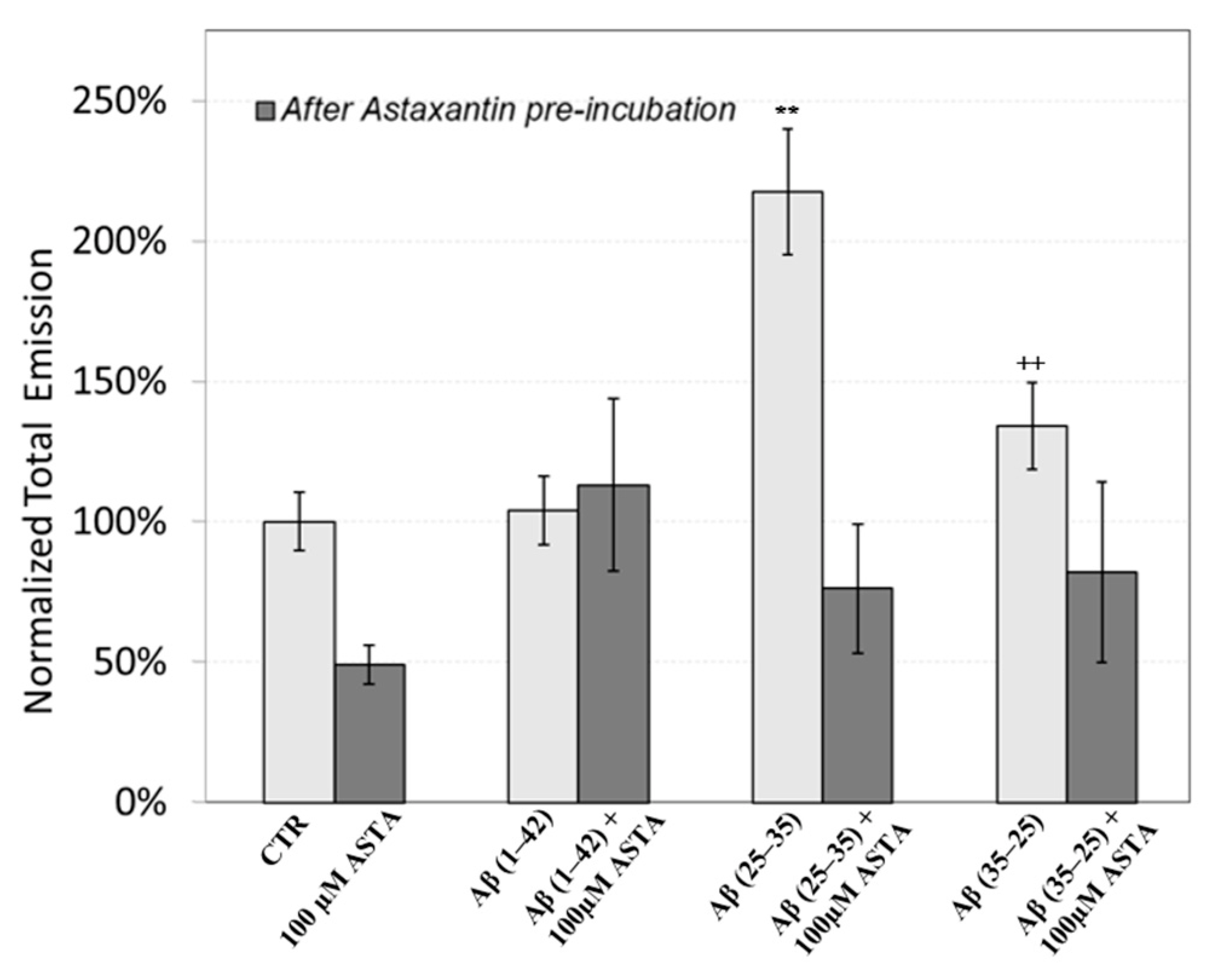
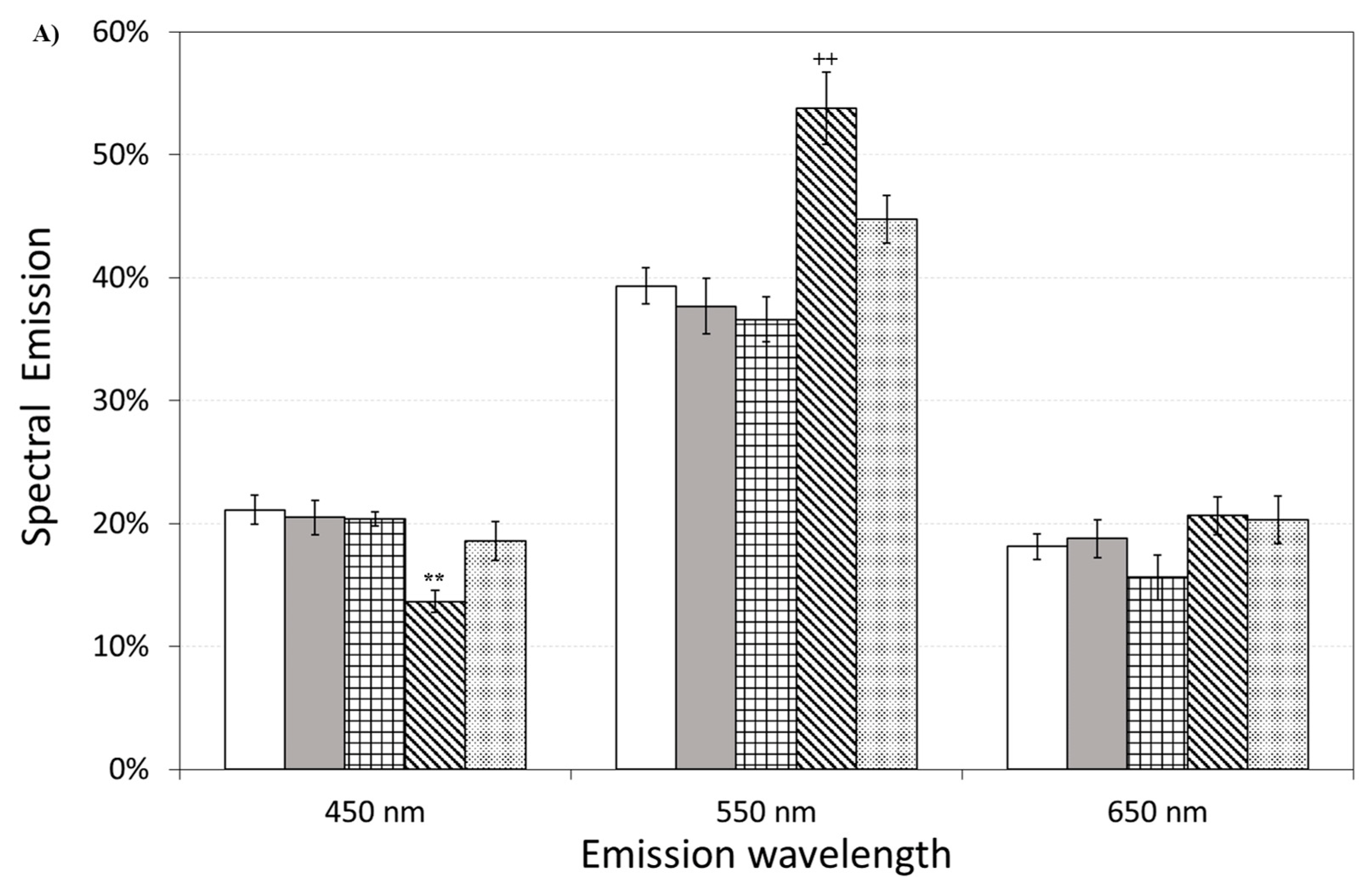
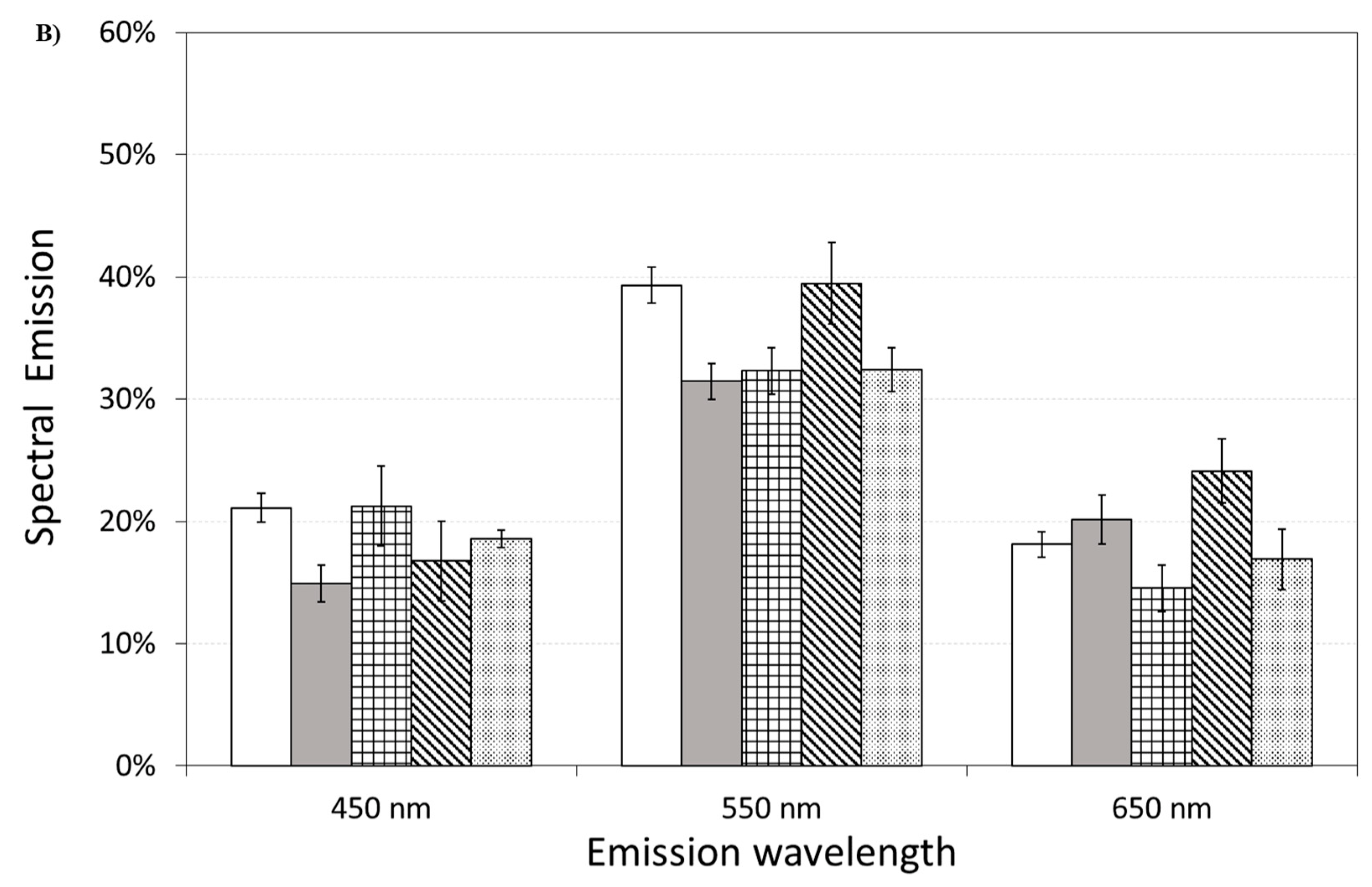
3.6. DL Spectroscopy
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Yuan, J.P.; Peng, J.; Yin, K.; Wang, J.H. Potential Health-Promoting Effects of Astaxanthin: A High-Value Carotenoid Mostly from Microalgae. Mol. Nutr. Food Res. 2011, 55, 150–165. [Google Scholar] [CrossRef]
- Galasso, C.; Corinaldesi, C.; Sansone, C. Carotenoids from Marine Organisms: Biological Functions and Industrial Applications. Antioxidants 2017, 6, 96. [Google Scholar] [CrossRef]
- Grimmig, B.; Kim, S.H.; Nash, K.; Bickford, P.C.; Douglas Shytle, R. Neuroprotective Mechanisms of Astaxanthin: A Potential Therapeutic Role in Preserving Cognitive Function in Age and Neurodegeneration. GeroScience 2017, 39, 19–32. [Google Scholar] [CrossRef]
- Pellitteri, R.; Bonfanti, R.; Spatuzza, M.; Cambria, M.T.; Ferrara, M.; Raciti, G.; Campisi, A. Effect of Some Growth Factors on Tissue Transglutaminase Overexpression Induced by β-Amyloid in Olfactory Ensheathing Cells. Mol. Neurobiol. 2017, 54, 6785–6794. [Google Scholar] [CrossRef]
- Campisi, A.; Raciti, G.; Sposito, G.; Grasso, R.; Chiacchio, M.A.; Spatuzza, M.; Attanzio, A.; Chiacchio, U.; Tesoriere, L.; Allegra, M.; et al. Amyloid-Beta Induces Different Expression Pattern of Tissue Transglutaminase and Its Isoforms on Olfactory Ensheathing Cells: Modulatory Effect of Indicaxanthin. Int. J. Mol. Sci. 2021, 22, 3388. [Google Scholar] [CrossRef]
- Campisi, A.; Sposito, G.; Pellitteri, R.; Santonocito, D.; Bisicchia, J.; Raciti, G.; Russo, C.; Nardiello, P.; Pignatello, R.; Casamenti, F.; et al. Effect of Unloaded and Curcumin-Loaded Solid Lipid Nanoparticles on Tissue Transglutaminase Isoforms Expression Levels in an Experimental Model of Alzheimer’s Disease. Antioxidants 2022, 11, 1863. [Google Scholar] [CrossRef]
- Lesort, M.; Tucholski, J.; Miller, M.L.; Johnson, G.V. Tissue Transglutaminase: A Possible Role in Neurodegenerative Diseases. Prog. Neurobiol. 2000, 61, 439–463. [Google Scholar] [CrossRef] [PubMed]
- Lee, M.Y.; Wu, M.F.; Cherng, S.H.; Chiu, L.Y.; Yang, T.Y.; Sheu, G.T. Tissue Transglutaminase 2 Expression Is Epigenetically Regulated in Human Lung Cancer Cells and Prevents Reactive Oxygen Species-Induced Apoptosis. Cancer Manag. Res. 2018, 10, 2835–2848. [Google Scholar] [CrossRef] [PubMed]
- Aeschlimann, D.; Wetterwald, A.; Fleisch, H.; Paulsson, M. Expression of Tissue Transglutaminase in Skeletal Tissues Correlates with Events of Terminal Differentiation of Chondrocytes. J. Cell Biol. 1993, 120, 1461–1470. [Google Scholar] [CrossRef] [PubMed]
- Gundemir, S.; Colak, G.; Tucholski, J.; Johnson, G.V.W. Transglutaminase 2: A Molecular Swiss Army Knife. Biochim. Biophys. Acta 2012, 1823, 406–419. [Google Scholar] [CrossRef]
- Mehta, K.; Lopez-Berestein, G. Expression of Tissue Transglutaminase in Cultured Monocytic Leukemia (THP-1) Cells during Differentiation. Cancer Res. 1986, 46, 1388–1394. [Google Scholar]
- Singh, G.; Zhang, J.; Ma, Y.; Cerione, R.A.; Antonyak, M.A. The Different Conformational States of Tissue Transglutaminase Have Opposing Affects on Cell Viability. J. Biol. Chem. 2016, 291, 9119–9132. [Google Scholar] [CrossRef]
- Lai, T.S.; Lin, C.J.; Wu, Y.T.; Wu, C.J. Tissue Transglutaminase (TG2) and Mitochondrial Function and Dysfunction. Front. Biosci. Landmark 2017, 22, 1114–1137. [Google Scholar] [CrossRef]
- Woodhall, E.; West, A.K.; Chuah, M.I. Cultured Olfactory Ensheathing Cells Express Nerve Growth Factor, Brain-Derived Neurotrophic Factor, Glia Cell Line-Derived Neurotrophic Factor and Their Receptors. Mol. Brain Res. 2001, 88, 203–213. [Google Scholar] [CrossRef]
- Mecocci, P.; Tinarelli, C.; Schulz, R.J.; Polidori, M.C. Nutraceuticals in Cognitive Impairment and Alzheimer’s Disease. Front. Pharmacol. 2014, 5, 147–158. [Google Scholar] [CrossRef] [PubMed]
- Allegra, M.; Tutone, M.; Tesoriere, L.; Almerico, A.M.; Culletta, G.; Livrea, M.A.; Attanzio, A. Indicaxanthin, a multi-target natural compound from Opuntia ficus-indica fruit: From its poly-pharmacological effects to biochemical mechanisms and molecular modelling studies. Eur. J. Med. Chem. 2019, 179, 753–764. [Google Scholar] [CrossRef] [PubMed]
- Brizhik, L.; Scordino, A.; Triglia, A.; Musumeci, F. Delayed Luminescence of Biological Systems Arising from Correlated Many-Soliton States. Phys. Rev. E Stat. Nonlin. Soft Matter. Phys. 2001, 64, 031902–031911. [Google Scholar] [CrossRef] [PubMed]
- Scordino, A.; Grasso, R.; Gulino, M.; Lanzanò, L.; Musumeci, F.; Privitera, G.; Tedesco, M.; Triglia, A.; Brizhik, L. Delayed Luminescence from Collagen as Arising from Soliton and Small Polaron States. Int. J. Quantum Chem. 2010, 110, 221–229. [Google Scholar] [CrossRef]
- Tudisco, S.; Musumeci, F.; Scordino, A.; Privitera, G. Advanced Research Equipment for Fast Ultraweak Luminescence Analysis. Rev. Sci. Instrum. 2003, 74, 4485–4490. [Google Scholar] [CrossRef]
- Lanzanò, L.; Scordino, A.; Privitera, S.; Tudisco, S.; Musumeci, F. Spectral Analysis of Delayed Luminescence from Human Skin as a Possible Non-Invasive Diagnostic Tool. Eur. Biophys. J. 2007, 39, 823–829. [Google Scholar] [CrossRef]
- Baran, I.; Ganea, C.; Scordino, A.; Musumeci, F.; Barresi, V.; Tudisco, S.; Privitera, S.; Grasso, R.; Condorelli, D.F.; Ursu, I.; et al. Effects of Menadione, Hydrogen Peroxide, and Quercetin on Apoptosis and Delayed Luminescence of Human Leukemia Jurkat T-Cells. Cell Biochem. Biophys. 2010, 58, 169–179. [Google Scholar] [CrossRef]
- Baran, I.; Ganea, C.; Privitera, S.; Scordino, A.; Barresi, V.; Musumeci, F.; Mocanu, M.M.; Condorelli, D.F.; Ursu, I.; Grasso, R.; et al. Detailed Analysis of Apoptosis and Delayed Luminescence of Human Leukemia Jurkat T Cells after Proton Irradiation and Treatments with Oxidant Agents and Flavonoids. Oxid. Med. Cell. Longev. 2012, 2012, 498914. [Google Scholar] [CrossRef]
- Scordino, A.; Campisi, A.; Grasso, R.; Bonfanti, R.; Gulino, M.; Iauk, L.; Parenti, R.; Musumeci, F. Delayed Luminescence to Monitor Programmed Cell Death Induced by Berberine on Thyroid Cancer Cells. J. Biomed. Opt. 2014, 19, 117005–117016. [Google Scholar] [CrossRef]
- Grasso, R.; Dell’Albani, P.; Carbone, C.; Spatuzza, M.; Bonfanti, R.; Sposito, G.; Puglisi, G.; Musumeci, F.; Scordino, A.; Campisi, A. Synergic Pro-Apoptotic Effects of Ferulic Acid and Nanostructured Lipid Carrier in Glioblastoma Cells Assessed through Molecular and Delayed Luminescence Studies. Sci. Rep. 2020, 10, 4680–4693. [Google Scholar] [CrossRef]
- Baran, I.; Ionescu, D.; Privitera, S.; Scordino, A.; Mocanu, M.M.; Musumeci, F.; Grasso, R.; Gulino, M.; Iftime, A.; Tofolean, I.T.; et al. Mitochondrial Respiratory Complex I Probed by Delayed Luminescence Spectroscopy. J. Biomed. Opt. 2013, 18, 127006–127028. [Google Scholar] [CrossRef]
- Ganne, A.; Balasubramaniam, M.; Griffin, W.S.T.; Shmookler Reis, R.J.; Ayyadevara, S. Glial Fibrillary Acidic Protein: A Biomarker and Drug Target for Alzheimer’s Disease. Pharmaceutics 2022, 14, 1354. [Google Scholar] [CrossRef] [PubMed]
- Chen, C.L.; Wang, L.J.; Yan, Y.T.; Hsu, H.W.; Su, H.L.; Chang, F.P.; Hsieh, P.C.H.; Hwang, S.M.; Shen, C.N. Cyclin D1 Acts as a Barrier to Pluripotent Reprogramming by Promoting Neural Progenitor Fate Commitment. FEBS Lett. 2014, 588, 4008–4017. [Google Scholar] [CrossRef]
- Pellitteri, R.; Spatuzza, M.; Russo, A.; Stanzani, S. Olfactory Ensheathing Cells Exert a Trophic Effect on the Hypothalamic Neurons in Vitro. Neurosci. Lett. 2007, 417, 24–29. [Google Scholar] [CrossRef] [PubMed]
- Chuah, M.I.; Au, C. Cultures of Ensheathing Cells from Neonatal Rat Olfactory Bulbs. Brain Res. 1993, 601, 213–220. [Google Scholar] [CrossRef] [PubMed]
- Campisi, A.; Spatuzza, M.; Russo, A.; Raciti, G.; Vanella, A.; Stanzani, S.; Pellitteri, R. Expression of Tissue Transglutaminase on Primary Olfactory Ensheathing Cells Cultures Exposed to Stress Conditions. Neurosci. Res. 2012, 72, 289–295. [Google Scholar] [CrossRef]
- Scordino, A.; Baran, I.; Gulino, M.; Ganea, C.; Grasso, R.; Niggli, J.H.; Musumeci, F. Ultra-Weak Delayed Luminescence in Cancer Research: A Review of the Results by the ARETUSA Equipment. J. Photochem. Photobiol. B Biol. 2014, 139, 76–84. [Google Scholar] [CrossRef] [PubMed]
- Nakamura, A.; Kaneko, N.; Villemagne, V.L.; Kato, T.; Doecke, J.; Doré, V.; Fowler, C.; Li, Q.X.; Martins, R.; Rowe, C.; et al. High Performance Plasma Amyloid-β Biomarkers for Alzheimer’s Disease. Nature 2018, 554, 249–254. [Google Scholar] [CrossRef] [PubMed]
- Butterfield, D.A.; Castegna, A.; Lauderback, C.M.; Drake, J. Evidence That Amyloid Beta-Peptide-Induced Lipid Peroxidation and Its Sequelae in Alzheimer’s Disease Brain Contribute to Neuronal Death. Neurobiol. Aging 2002, 23, 655–664. [Google Scholar] [CrossRef]
- Pradeepkiran, J.A.; Reddy, P.H. Defective Mitophagy in Alzheimer’s Disease. Ageing Res. Rev. 2020, 64, 101191–101229. [Google Scholar] [CrossRef]
- Naguib, Y.M. Antioxidant Activities of Astaxanthin and Related Carotenoids. J. Agric. Food Chem. 2000, 48, 1150–1154. [Google Scholar] [CrossRef] [PubMed]
- Nakajima, Y.; Inokuchi, Y.; Shimazawa, M.; Otsubo, K.; Ishibashi, T.; Hara, H. Astaxanthin, a Dietary Carotenoid, Protects Retinal Cells against Oxidative Stress in-Vitro and in Mice in-Vivo. J. Pharm. Pharmacol. 2008, 60, 1365–1374. [Google Scholar] [CrossRef]
- Zhang, X.S.; Zhang, X.; Wu, Q.; Li, W.; Zhang, Q.R.; Wang, C.X.; Zhou, X.M.; Li, H.; Shi, J.X.; Zhou, M.L. Astaxanthin Alleviates Early Brain Injury Following Subarachnoid Hemorrhage in Rats: Possible Involvement of Akt/Bad Signaling. Mar. Drugs 2014, 12, 4291–4310. [Google Scholar] [CrossRef]
- Campisi, A.; Caccamo, D.; Li Volti, G.; Currò, M.; Parisi, G.; Avola, R.; Vanella, A.; Ientile, R. Glutamate-Evoked Redox State Alterations Are Involved in Tissue Transglutaminase Upregulation in Primary Astrocyte Cultures. FEBS Lett. 2004, 578, 80–84. [Google Scholar] [CrossRef]
- Kim, J.H.; Nam, S.W.; Kim, B.W.; Choi, W.; Lee, J.H.; Kim, W.J.; Choi, Y.H. Astaxanthin Improves Stem Cell Potency via an Increase in the Proliferation of Neural Progenitor Cells. Int. J. Mol. Sci. 2010, 11, 5109–5119. [Google Scholar] [CrossRef]
- Cullen, D.K.; Simon, C.M.; LaPlaca, M.C. Strain Rate-Dependent Induction of Reactive Astrogliosis and Cell Death in Three-Dimensional Neuronal-Astrocytic Co-Cultures. Brain Res. 2007, 1158, 103–115. [Google Scholar] [CrossRef]
- Tian, M.; Li, Q.; Liu, Y.; Zheng, P.; Li, D.; Zhao, Y.; Wang, B.; Li, C.; Wang, J.; Gao, P.; et al. Relationship between delayed luminescence emission and mitochondrial status in Saccharomyces cerevisiae. Sci. Rep. 2022, 12, 394. [Google Scholar] [CrossRef]
- Swomley, A.M.; Förster, S.; Keeney, J.T.; Triplett, J.; Zhang, Z.; Sultana, R.; Butterfield, D.A. Abeta, Oxidative Stress in Alzheimer Disease: Evidence Based on Proteomics Studies. Biochim. Biophys. Acta (BBA)-Mol. Basis Dis. 2014, 1842, 1248–1257. [Google Scholar] [CrossRef] [PubMed]
- Varadarajan, S.; Kanski, J.; Aksenova, M.; Lauderback, C.; Butterfield, D.A. Different Mechanisms of Oxidative Stress and Neurotoxicity for Alzheimer’s A Beta(1–42) and A Beta(25–35). J. Am. Chem. Soc. 2001, 123, 5625–5631. [Google Scholar] [CrossRef]
- Croce, A.C.; Bottiroli, G. New Light in Flavin Autofluorescence. Eur. J. Histochem. 2015, 59, 2576–2578. [Google Scholar] [CrossRef] [PubMed]
- Clementi, M.E.; Marini, S.; Coletta, M.; Orsini, F.; Giardina, B.; Misiti, F. Aβ(31–35) and Aβ(25–35) Fragments of Amyloid Beta-Protein Induce Cellular Death through Apoptotic Signals: Role of the Redox State of Methionine-35. FEBS Lett. 2005, 579, 2913–2918. [Google Scholar] [CrossRef] [PubMed]













Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Campisi, A.; Sposito, G.; Grasso, R.; Bisicchia, J.; Spatuzza, M.; Raciti, G.; Scordino, A.; Pellitteri, R. Effect of Astaxanthin on Tissue Transglutaminase and Cytoskeletal Protein Expression in Amyloid-Beta Stressed Olfactory Ensheathing Cells: Molecular and Delayed Luminescence Studies. Antioxidants 2023, 12, 750. https://doi.org/10.3390/antiox12030750
Campisi A, Sposito G, Grasso R, Bisicchia J, Spatuzza M, Raciti G, Scordino A, Pellitteri R. Effect of Astaxanthin on Tissue Transglutaminase and Cytoskeletal Protein Expression in Amyloid-Beta Stressed Olfactory Ensheathing Cells: Molecular and Delayed Luminescence Studies. Antioxidants. 2023; 12(3):750. https://doi.org/10.3390/antiox12030750
Chicago/Turabian StyleCampisi, Agatina, Giovanni Sposito, Rosaria Grasso, Julia Bisicchia, Michela Spatuzza, Giuseppina Raciti, Agata Scordino, and Rosalia Pellitteri. 2023. "Effect of Astaxanthin on Tissue Transglutaminase and Cytoskeletal Protein Expression in Amyloid-Beta Stressed Olfactory Ensheathing Cells: Molecular and Delayed Luminescence Studies" Antioxidants 12, no. 3: 750. https://doi.org/10.3390/antiox12030750
APA StyleCampisi, A., Sposito, G., Grasso, R., Bisicchia, J., Spatuzza, M., Raciti, G., Scordino, A., & Pellitteri, R. (2023). Effect of Astaxanthin on Tissue Transglutaminase and Cytoskeletal Protein Expression in Amyloid-Beta Stressed Olfactory Ensheathing Cells: Molecular and Delayed Luminescence Studies. Antioxidants, 12(3), 750. https://doi.org/10.3390/antiox12030750