Abstract
The increase in environmental temperature due to global warming is a critical threat to plant growth and productivity. Heat stress can cause impairment in several biochemical and physiological processes. Plants sense and respond to this adverse environmental condition by activating a plethora of defense systems. Among them, the heat stress response (HSR) involves an intricate network of heat shock factors (HSFs) and heat shock proteins (HSPs). However, a growing amount of evidence suggests that reactive oxygen species (ROS), besides potentially being responsible for cellular oxidative damage, can act as signal molecules in HSR, leading to adaptative responses. The role of ROS as toxic or signal molecules depends on the fine balance between their production and scavenging. Enzymatic and non-enzymatic antioxidants represent the first line of defense against oxidative damage and their activity is critical to maintaining an optimal redox environment. However, the HS-dependent ROS burst temporarily oxidizes the cellular environment, triggering redox-dependent signaling cascades. This review provides an overview of the redox-activated mechanisms that participate in the HSR.
1. Introduction
In recent decades, along with natural causes, anthropic activities have brought about an alarming trend of global warming, which is driving drastic changes in our climate, resulting in more frequent and intense heat waves and causing substantial losses to agricultural production [1,2,3]. Plants can be exposed to different forms of heat stress (HS), such as heat shock, which is an increase in temperature of 10–15 °C above ambient for a short period (from several minutes to a few hours) or prolonged warming that consists in exposing plants to a modest increase in temperature (2–5 °C above ambient) for several days or weeks [4,5]. HS affects plant growth, development, and reproduction at morphological, physiological, and molecular levels [6], even though plants can use different strategies to combat these two types of HS [7].
One worrying consequence of temperature rise is how it severely limits crop productivity. HS occurring during gametogenesis causes morphological abnormalities in pollen and ovary ultrastructure and viability, thus affecting fertilization efficiency; moreover, HS impacts the duration of grain filling, the availability of assimilates, and the enzymes of starch metabolism while also altering the abscisic acid/ethylene ratio [8,9,10]. Like other alterations in environmental conditions, such as cold, drought, flooding or nutrient deficiency, heat directly affects plant growth and modifies plant relations with pathogens, thus strongly impacting crop yield [11]. As occurs in many other biotic and abiotic stresses [12,13,14], under HS, several biochemical and physiological processes are altered, including water potential, transpiration, nutrient uptake and transport, cell division and differentiation, respiration, and photosynthesis [15,16,17,18]. Photosynthesis is the main physiological process that is affected by heat. High temperatures modify the microscopic ultrastructure of chloroplasts, altering membrane fluidity and increasing thylakoid leakiness [19]. Heat damage in chloroplasts occurs principally on photosystem II (PSII) [20,21,22,23], the oxygen-evolving complex (OEC) [24], the ATP-generating system, and the cytochrome b6f complex [25,26]. In addition, HS also affects the activity of RUBISCO activase [27,28], the metabolism of carbon assimilation, and chloroplast biosynthesis, leading to leaf senescence, which is a clear symptom of heat injury [29,30,31,32]. At the cellular level, HS increases the production of reactive oxygen species (ROS), which directly or indirectly may cause many injuries such as protein denaturation, the inhibition of protein synthesis, the loss of membrane and cytoskeleton integrity, and the inactivation of enzymes [33,34].
As sessile organisms, plants have developed sophisticated signaling networks to sense HS and react to the harmful effects of high temperatures [34,35,36,37,38]. Since photosynthetic apparatus is the primary target of heat damage, chloroplasts can act as sensors of HS [26,38], activating retrograde signaling that induces changes in the expression of nuclear-encoded genes for the metabolic and molecular reprogramming required for stress adaptation [39,40,41]. One of the most well-studied mechanisms of the heat stress response (HSR) is the intricate network of heat shock factors (HSFs) and heat shock proteins (HSPs). HSPs are chaperone proteins that are able to repair aggregated, denatured, or misfolded proteins, allowing plants to maintain their delicate metabolic processes during heat and other stress [42].Through their association with the heat shock element (HSE), in the promoter of HSPs and other heat-responsive genes, HSFs alter gene expression to obtain adjustments in the molecular pathways involved in enabling plants to survive under high temperatures [43].
In recent years, many data have highlighted a significant role of ROS as signaling molecules in the HSR [44,45]; moreover, several genetic and biochemical studies have indicated that complex interactions take place between HSFs/HSPs, ROS, and the heat acclimatation response [46]. This review aims to provide and discuss data on the role of ROS and the redox-related signaling pathways that are involved in the acquisition of thermotolerance.
2. Production of Reactive Oxygen Species in Heat Stress Conditions
High temperature, like many environmental stresses, leads to the overproduction of ROS, the reactive forms of molecular oxygen (O2), including singlet oxygen (1O2), superoxide anion radical (O2•−), hydrogen peroxide (H2O2), and hydroxyl radical (HO•). Under environmental stress conditions, ROS accumulation results in oxidative damage to biomolecules, such as pigments, proteins, lipids, carbohydrates, and DNA, leading to cellular injuries and cell death. However, a fine balance between production and scavenging, controlling ROS levels, allows these molecules to act as molecular signals that induce changes in cellular metabolism, leading to adaptative responses or triggering programmed cell death (PCD) [47,48].
Under abiotic stress, ROS production occurs in different cellular compartments, such as chloroplasts, mitochondria, peroxisomes, cell wall, apoplast, and plasma membrane. During stress conditions, ROS production occurring in the photosynthetic and respiratory electron transport chains performs a regulatory function in alleviating over-reduction [49]. Moreover, the redox state of chloroplasts, peroxisomes, and mitochondria is connected by photorespiration, which is affected by several environmental stresses [50]. ROS accumulation in different organelles strictly depends on the plant tissues. In organs with low photosynthetic metabolism, such as fruits and flowers, ROS are principally formed in the mitochondria, peroxisomes, and apoplast [51], while in photosynthetic tissues, chloroplasts are the prime source of ROS [52]. Chloroplasts act as the principal sensors of environmental stresses, including heat [53]. HS in chloroplasts reduces photosynthetic efficiency, deactivating the reaction center of PSII and inhibiting electron flow to the plastoquinone pool and the induction of non-photochemical quenching [54]. Damage to the photosynthetic components can lead to excess excitation energy, resulting in the formation of a chlorophyll triplet state that reacts with molecular oxygen, forming 1O2 [55]. 1O2 formation at high temperatures is also associated with heat-induced lipid peroxidation in the thylakoid membrane and damage to PSII [56]. In the unicellular green alga, Chlamydomonas reinhardtii, which is exposed to 40 °C, it has been suggested that 1O2 formed by lipid peroxidation is initiated by an enzymatic reaction that is catalyzed by lipoxygenase [57]. Additionally, 1O2 can be produced by the direct decomposition of tetraoxide via the Russell reaction [57].
In chloroplasts, O2•− is formed by three processes: (i) in PSII, associated with incomplete water oxidation, (ii) in the electron transport chain, via plastosemiquinone interaction with O2, and (iii) in Photosystem I (PSI), due to the oxidation of ferredoxin that reduces O2 to O2•− and that takes place when the Calvin–Benson cycle does not operate properly and lowers NADP+ availability [50,53]. O2•− is disproportionate to H2O2 and O2 in a reaction catalyzed by superoxide dismutase (SOD) [58]. Moreover, H2O2 can also be produced in PSII when HS causes the cleavage and degradation of the D1 protein with the consequent release of PsbO, PsbP, and PsbQ [23,56]. These three proteins, which together with the inorganic cluster of Mn4O5Ca form the OEC, are located on the inner surface of the thylakoid membrane, in close association with PSII, and are involved in its thermal stability [59]. Indeed, the release of PsbO, PsbP, and PsbQ proteins leads to improper H2O accessibility to the water-splitting manganese complex and the consequent formation of H2O2. On the PSII electron donor site, H2O2, formed by incomplete water oxidation, can produce the more toxic HO•, through the Fenton reaction [60].
Under HS, the disturbances of respiratory metabolisms in the mitochondria trigger impairments of the electron transport chain (ETC), resulting in an increase in ROS levels. In tobacco cells, an impaired mitochondrial metabolism is responsible for the oxidative burst arising during heat-induced PCD [61,62]. O2•−, which is formed when a single electron leaks to O2 in ETC components, can act as a substrate for the generation of H2O2 by matrix-localized manganese SOD (MnSOD) [63]. HS promotes Ca2+ increase in mitochondria, which is accompanied by hyperpolarization of the inner mitochondrial membrane, leading to an increase in ROS generation [64]. HS increases membrane fluidity, impairing cytochrome c oxidase (COX), resulting in an over-reduction in the ETC that culminates in ROS production [65,66]. The maintenance of COX integrity is favored by the mitochondrial HSP70-1; consistently, mtHSC70-1 knockout lines show decreased COX activity and an increase in ROS production [67]. Additionally, during HS, an induction of COX1, COX2, and COX3 genes has been reported in Arabidopsis [7], highlighting the fact that plants try to counteract HS injury to mitochondria by preventing imbalances in the ETC. Moreover, in plants, the presence of alternative oxidase (AOX), which catalyzes the oxidation of ubiquinol and the reduction of O2 to H2O, helps to maintain electron flow, preventing the over-reduction in the ETC and O2•− formation [68]. The increase in the AOX protein after exposure to high temperatures confirms the participation of this enzyme in HSR [69].
During HS, ROS generation also comes from the activity of the plasma membrane NADPH-oxidase, which transfers an electron from intracellular NADPH to O2 in the apoplast, generating O2•−, which is then converted into H2O2 by SOD [70,71]. NADPH-oxidase is controlled by phosphorylation and by the direct binding of calcium. Thus, cytosolic calcium influx, occurring as a result of heat perception, activates the enzyme and leads to ROS accumulation [72]. In Arabidopsis, NADPH-oxidase is encoded by ten genes (respiratory burst oxidase homologues—RBOHs), which are involved in several biological processes, including biotic and abiotic stress responses. AtrbohB and AtrbohD play a key role in HSR, as suggested by the heat-sensitive phenotype observed in atrbohB and atrbohD Arabidopsis mutants [73]. In rice, six NADPH-oxidase genes increase their expression in response to HS [74]. NADPH-oxidase has a crucial function in thermotolerance acquisition, enhancing the expression of heat-induced genes and controlling the H2O2 level by regulating antioxidant enzymes [75].
3. Role of Antioxidants in Heat Stress Response
The role of ROS as toxic or signaling molecules depends on a delicate equilibrium between the production and scavenging of these reactive species in different cell compartments [46,47]. Antioxidants are the first line of defense against oxidative damage and their activity is critical to maintaining an optimal redox environment. The antioxidant machinery includes non-enzymatic components, such as ascorbate (ASC), glutathione (GSH), carotenoids and tocopherols, as well as enzymatic components such as ascorbate peroxidase (APX), catalase (CAT), SOD, peroxidases (PODs), glutathione peroxidase (GPX), glutathione S-transferase, and the enzymes involved in the reduction of the oxidized forms of ASC and GSH in the ASC–GSH cycle, namely, monodehydroascorbate reductase, dehydroascorbate reductase, and glutathione reductase (GR) [76].
A primary strategy used to Iimpart thermotolerance in plants involves enhancing the antioxidant machinery. In wheat, HS is better tolerated by plants that are subjected to heat priming. The improvement of thermotolerance is related to the enhanced activity of antioxidant enzymes, particularly to the upregulation of chloroplastic Cu/Zn-SOD and mitochondrial GR and POD [77,78]. Consistently, in Chinese Spring, an HS-sensitive wheat variety, the significant decrease in SOD, CAT, POD, and GPX activities, occurring after exposure to high temperature, causes ROS accumulation and increased levels of oxidized lipids [79]. In apple trees, HS (48 °C for 6 h)—despite the increases in antioxidant activity—causes a rise in H2O2, O2•− and lipid peroxidation, and a decrease in chlorophyll content. The overexpression of MdATG18a, encoding a key protein involved in autophagy, removes damaged chloroplasts and enhances antioxidant activity, thus improving the expression of heat-related genes. In transgenic plants, the reduction of ROS accumulation confers thermotolerance [80].
Many treatments that confer thermotolerance activate antioxidant machinery. In potato seedlings, a pre-treatment with sucrose, before exposure to HS, enhances SOD, CAT, APX, and POD activity, reducing ROS accumulation and resulting in increased thermotolerance [81]. Similar results have been obtained in potato plants treated with exogenous silicon, which mitigates heat-induced oxidative stress by increasing the expression of antioxidant enzymes [82]. In strawberries, the enhanced transcription of biosynthetic enzymes of ASC and GSH and antioxidant enzymes (APX, CAT, SOD, and GR), occurring in plants pre-treated with sodium hydrosulfide before HS, leads to a higher stress tolerance [83]. Furthermore, tomato thermotolerance due to the exogenous application of rosmarinic acid, a plant-derived phenolic compound, is linked to the increased transcript abundance and activity of ASC–GSH cycle enzymes, which modulate the ASC and GSH redox state [84].
The behavior of antioxidant machinery to counteract and/or mitigate heat injury depends on plant species, genotypes, phenological stage, and stress severity [85,86,87,88,89,90]. Moreover, different isoforms of antioxidant enzymes can show different performance in HSR; therefore, changes in the antioxidant systems in response to HS denote a complex scenario with a number of controversial points.
Among the antioxidant enzymes, CAT is one of the main scavengers of H2O2. Arabidopsis plants possess three CAT isoenzymes differing in their subcellular localization and exhibiting different tissue specificity. Analysis of cat mutants has suggested that CAT2 has a greater effect in removing H2O2 compared to CAT1 and CAT3, which are mainly “backup” or stress-specific enzymes [91]. Studies on Arabidopsis mutants lacking the three CATs have revealed that only cat2, but not cat1 and cat3, are hypersensitive to moderate long-term HS (35 °C for 5 days), but the same mutants do not differ from the wild type (wt) in response to short-term HS (42 °C for 40 min). However, the CAT2-overexpressing lines do not show improved thermotolerance, suggesting that CAT2 is necessary but insufficient to enhance long-term HS response [92]. In Brassica napus seedlings, the total CAT activity decreases after 24 and 48 h of HS at 38°; nevertheless, after selenium supply, which confers HS-tolerance, the increase in CAT activity, along with the subsequent reduction in oxidative stress, indicate a role for CAT in H2O2 detoxification, which is responsible for thermotolerance [93].
APX is a plant antioxidant enzyme that uses ascorbate as an electron donor and plays a key role in the scavenging of H2O2. In Arabidopsis, there are different isoenzymes classified based on their subcellular localization and that target cytosol (i.e., APX1 and APX2), mitochondria (mitAPX), chloroplastic stroma (sAPX), and thylakoid membranes (tAPX) [94]. Both cytosolic isoforms possess a heat shock element (HSE) in the promoters and are induced during HSR [95,96,97,98]. Arabidopsis apx2 mutants are more sensitive to HS at the seedling stage, showing a lower survival rate than the wt, but showing higher thermotolerance at the reproductive stage, producing more seeds under prolonged heat stress. This finding supports the idea that the lack of APX2 induces a signal suppressing ROS accumulation during stress, through the activation of different redundant ROS scavenging systems or the suppression of ROS production and that this signal may be stage- or tissue-specific [99]. The phosphorylation of APX2 by the calcium-dependent protein kinase CPK28 is needed to enhance thermotolerance; indeed, cpk28 mutants show increased HS-induced ROS accumulation and protein oxidation together with the reduced activity of APX [100]. The lack of APX1 in Arabidopsis causes stunted growth and enhanced sensitivity to oxidative stress [101]; moreover, APX1 seems to be necessary to increase tolerance to HS combined with drought [102,103]. Interestingly, APX1 subjected to heat treatment shows conformational changes, passing from dimeric to high-molecular-weight structures, losing peroxidase activity and acquiring chaperone activity, which can confer thermotolerance [104].
Under HS, the expression profile of APX1 and APX2 is different; in Arabidopsis, APX1 mRNA is already present in unstressed conditions and shows a modest increase under moderate HS, whereas APX2 mRNA is not detected in control conditions and has a fast, transient (1–2 h), and strong induction after HS [95,96]. Similarly, in independent studies, the maximal expression of APX1 and APX2 has been observed after 24 h and 15 min of HS, respectively [98,105]. These data suggest a similar but not superimposable role of these two isoforms.
The involvement of APX in HSR has also been widely studied in tobacco BY-2 cells. In these cells, short-term HS at 35 °C or 55 °C causes an increase or a decrease in APX activity, respectively [106]. The increase in APX activity under moderate HS is only imputable to the cytosolic isoenzymes, whereas exposure at 55 °C decreases all of the APX isoenzymes [107]. In this context, the inhibition of APX activity causes ROS accumulation and participates in the induction of PCD [61]. The reduction in cytosolic APX activity under PCD conditions is a precocious event, due to nitrosylation, ubiquitination, and the consequent proteasome-dependent degradation of the protein [108]. In tobacco BY-2 cells, cytosolic APX behaves differently depending on the duration of the moderate HS: activity and expression increase in the first 3 days of HS. However, when HS is protracted, a drop in enzyme levels is responsible for the rise in oxidative damage, leading to cell death [109].
Inconsistent behavior of isoenzyme regulation in the HSR has also been reported for SODs. In tomato plants, heat acclimation before HS favors an increase in the total SOD activity, but while Fe-SODs and Mn-SODs are induced, Cu/Zn-SODs decrease. The increase in Fe-SOD activity seems to be strictly related to conditions where the photosynthetic functions are unaffected by heat, indicating that Fe-SODs could participate either in reducing the spread of ROS and also protecting the thylakoid membrane and PSII reaction center from oxidative damage [110,111]. More recently, it has been reported that the decrease in Cu/Zn-SOD activity, with concomitant ROS accumulation, enhances thermotolerance in Arabidopsis [112]; similarly, a decrease in Cu/ZnSOD and Fe-SOD mRNA levels has been observed in Arabidopsis subject to 45 °C for 2 h [113]. This scenario suggests that the activity of these antioxidant enzymes in chloroplasts may be downregulated by HS, thus exacerbating ROS accumulation, in order to obtain cellular environment oxidation that functions as a signal to activate stress-responsive systems [113].
In tomato, an increase in APX and SOD activities, but not in gene expression, occurs after HS. The increase in the activity of these antioxidant enzymes is due to the induction of the HSP40, LeCDJ1, which acts as a chaperone that protects APX and SOD protein conformation, increasing their enzymatic activity, alleviating PSII damage, and conferring heat tolerance [114,115]. More recently, it has been reported that HSP40 interacts with SISNAT, an enzyme located in the chloroplast that participates in the biosynthesis of the ROS scavenger melatonin. SISNAT-overexpressing lines subjected to HS show decreased ROS levels, increased RUBISCO protection, and an upregulation of HSFs and HSPs [116]. Consistently, treatment with exogenous melatonin before HS increases thermotolerance, as a consequence of the reduction in oxidative stress [117,118].
HS tolerance in plants is also associated with non-enzymatic antioxidants. In mung bean plants, HS at 42 °C leads to a decrease in the ASC content and GSH/GSSG ratio, with a concomitant increase in oxidative markers; however, the supply of exogenous GSH induces tolerance to high temperatures by improving the antioxidant systems [119]. Similar results have been reported in tomato, where the decrease in GSH/GSSG ratio, which is attributed to the impairment of GSH recycling, supports the idea that the maintenance of GSSG under stress conditions is an indicator of high ROS accumulation [120,121].
4. Reactive Oxygen Species as Signaling Molecules in the Heat Stress Response
The heat-induced ROS accumulation is frequently considered in terms of oxidative stress, cell damage, and death. However, ROS can act as signaling molecules crucial for the acquisition of thermotolerance (Figure 1) [4,12,68,79,114,115,116,122,123]. Indeed, the amount of H2O2 increases after the exposure to high temperatures, causing HSFs activation and the accumulation of HSPs [124]. Consistently, in HS-exposed plants, inhibitors of H2O2 synthesis and ROS scavengers can prevent HSP expression [125]. Studies on mutants of ROS biosynthesis corroborate the idea of ROS requirements for achieving thermotolerance. Mutations in AtrbohB and AtrbohD, which participate in H2O2 production in the HSR, lead to an increased heat sensitivity [73]. In addition to not accumulating H2O2, atrbohB, atrbohD, and atrbohB/D double mutants fail to induce the expression of HSP17.7 and HSP21 [126]. H2O2 formed in the apoplast can be promptly sensed by membrane receptor kinases such as HPCA1, which causes a calcium influx that activates MAP kinases and additional signaling pathways [127].
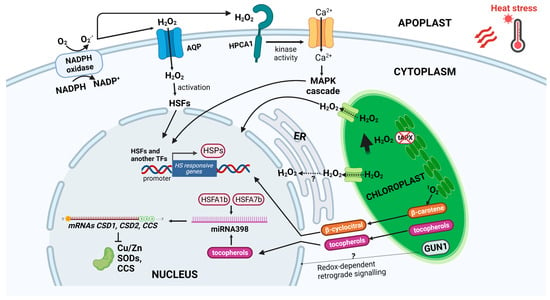
Figure 1.
Mechanisms of ROS accumulation and signaling in heat stress response. After heat stress, production, and accumulation of reactive oxygen species (ROS) occurs in both chloroplasts and apoplast. 1O2 accumulation in the chloroplast, oxidizing β-carotene, produces β-cyclocitral, which acts as a retrograde signal from chloroplast to nucleus, activating heat stress-responsive (HSR) genes. In chloroplasts, damage to photosynthetic apparatus, as well as inhibition of tAPX, lead to accumulation of H2O2, which moves to the nucleus through aquaporins, and/or ER, thus inducing HSR genes. The chloroplast-localized GUN1, by the redox-dependent plastid to nucleus communication, is involved in cell environment oxidation. ROS increase is also due to downregulation of CSD1, CSD2, and CCS by miR398, whose accumulation is dependent on HSFA1b and HSFA7b. Tocopherols produced in the chloroplast participate in miR398 induction. H2O2 produced in the apoplast by NADPH oxidase can diffuse in the cell via aquaporins, and/or sensed by membrane receptor HPCA1, which stimulates a calcium influx, triggering the MAP kinases cascade and transcription of HSPs. Abbreviations: 1O2, singlet oxygen; tAPX, thylakoidal ascorbate peroxidase; H2O2, hydrogen peroxide; ER; endoplasmic reticulum; GUN1, genomes uncoupled1; CSD, Cu/Zn superoxide dismutase; CCS, Cu chaperone for superoxide dismutase; HSF, heat shock factor; HPCA1, HYDROGEN-PEROXIDE-INDUCED CALCIUM INCREASES1; MAP; mitogen activated protein; and HSP, heat shock protein. Created with BioRender.com; accessed on 3 February 2023.
The downregulation of ROS scavenger enzymes also contributes to ROS accumulation and the activation of HSFs, giving a positive feedback loop regulating thermotolerance. During HS, HSFA1b and HSFA7b induce miR398, which downregulates CSD1, CSD2, and CCS genes, encoding cytosolic and chloroplast Cu/Zn SODs and a copper chaperone that delivers Cu for both SODs, respectively. Therefore, ROS accumulate and induce other HSF genes, promoting heat stress tolerance. Consistently csd1, csd2, and ccd mutants and plants over-expressing miRNA398 are more heat tolerant [112]. The lipid soluble antioxidant tocopherols, synthesized in chloroplast during HS, positively regulate the biogenesis of miRNAs, and this regulation involves the heat-induced accumulation of miRNA398. In the biosynthetic tocopherol mutant vte2-1, the induction of miR398 and suppression of CSD2 by heat are abolished; additionally, knocking down CSD2 in vte2-1 restores heat tolerance, supporting the evidence that SODs inhibition and ROS accumulation increase thermotolerance (Figure 1) [128].
Chloroplasts activate retrograde signaling to communicate environmental stress and induce changes in the expression of nuclear-encoded genes, which bring metabolic and molecular reprogramming for stress adaptation. Chloroplasts have been shown to play an important role in heat-induced ROS accumulation and the subsequent expression of nuclear heat-responsive genes [129]. ROS produced in chloroplasts can work as plastid signals to activate the expression of genes coding for antioxidant enzymes and to fine-tune the stress-responsive apparatus for a more effective adaptation to stresses [40].
Among ROS, 1O2 and H2O2 impact nuclear gene expression extensively. Singlet oxygen can function as a retrograde signal to activate nuclear gene expression. Given the very short half-life of 1O2 (200 ns) and its high reactivity, it can be located exclusively inside chloroplasts. Thus, more stable second messengers derived from 1O2 within the plastids are assumed to activate signaling pathways in controlling the expression of nuclear genes [40,130,131,132]. The 1O2-induced pathway involves the accumulation of β-cyclocitral, an oxidation product of β-carotene. β-cyclocitral could directly travel to the nucleus and activate 1O2-dependent retrograde signaling; it is positively correlated with heat tolerance because it is directly involved in the induction of responsive genes, such as genes responding to oxidative stress, HSE-binding proteins, and various defense genes (Figure 1) [125,126,127]. H2O2 derived from chloroplasts also acts as a specific signal for regulating plant responses to stress. In Arabidopsis, the t-APX silencing, increasing H2O2 levels in chloroplasts, affects the expression of a large set of genes associated with stress [133]. GUN1, a key player of plastid-to-nucleus retrograde signaling [134,135,136], has been proposed to be involved in ROS accumulation occurring in response to HS. Indeed, unlike wt plants, gun1 mutants do not accumulate O2− and H2O2, failing to induce the transient oxidative burst needed for HSR and resulting in more HS sensitivity [113] (Figure 1).
The direct transfer of H2O2 from chloroplasts to the nucleus has been proposed as a signaling way to induce nuclear heat-associated gene expression and the close association between the two compartments may provide the specificity needed for retrograde signal transduction. So that H2O2 passes from the chloroplast to the nucleus, it must cross the chloroplast double envelope, the perinuclear space, and the inner nuclear membrane; this movement probably occurs by diffusion via water channels [137]. Stromules may also promote chloroplasts-to-nucleus contacts, transferring H2O2 to the nucleus [138]. The close association between the endoplasmic reticulum (ER) with chloroplasts and nucleus has led to suppose that the ER may play a key role in mediating H2O2 retrograde signaling in a double manner: amplifying the signal, generating more H2O2 by a luminal ER oxidase, or attenuating it by activating antioxidant responses such as the ER glutathione peroxidase [139] (Figure 1).
5. Heat Stress-Dependent Crosstalk between Signaling Networks of Reactive Oxygen Species and Hormones
During HS, ROS signaling is also modulated by a wide interaction with hormone signaling [140,141]. Indeed, ROS and hormones that are apart to cooperate as single molecules are also connected by crucial signaling factors, such as calcium, cyclic nucleotides, MAPKs, G-proteins, and various transcription factors or regulator molecules. This delicate communication between ROS and hormones signaling organizes a suitable response that permits the plant to overcome the stress condition (Figure 2) [35,142,143,144,145].
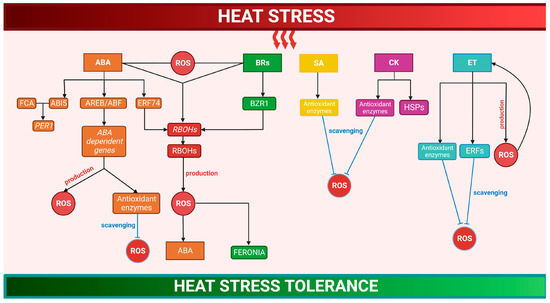
Figure 2.
Crosstalk between signaling networks of reactive oxygen species and hormones in heat stress response. Under HS, ABA activates AREB/ABF transcription factors controlling ABA-dependent gene expression, stimulating ROS production, or improving antioxidant machinery. ABA-induced ERF74 also increases ROS production by RBOHs induction. The interaction of FCA with ABI5 increases thermotolerance through the transcriptional regulation of PER1. BRs, by activating RBOHs, stimulate a transient H2O2 accumulation, which increases ABA levels resulting in additional rises in H2O2. In BR-mediated heat tolerance, the regulator BZR1 improves the production of ROS, which regulates the receptor-like kinase FERONIA. SA, CK, and ET enhance antioxidant systems, increasing ROS scavenging. In addition, CK stimulate HSPs. ROS can act in concert with ET in a self-intensifying feedback loop where ET stimulates H2O2 accumulation, which in turn improves ET production. ERFs contribute to lower ROS accumulation, balancing redox homeostasis. (See more details in the text). Abbreviations: ABA, abscisic acid; AREB/ABF, ABRE-binding/ABRE-binding factors; ERF74, ethylene response factor 74; RBOH, respiratory burst oxidase homolog; FCA, flowering control locus A; ABI5, ABA-INSENSITIVE 5; PER1, 1-Cys peroxiredoxin; BRs, brassinosteroids; BZR1, brassinosteroid-regulated transcription factor 1; SA, salicylic acid; CK, cytokine; ET, ethylene; and ERF, ethylene response factors. Created with BioRender.com.; accessed on 20 February 2023.
Hormones induced by HS can modify ROS amounts, activating RBOHs or modifying redox signaling, or ROS can act upstream the hormones. Moreover, ROS can play a role in mediating the interaction between different hormones, thus improving HS tolerance. These interactions lead to an amplification loop, controlling the gene expression and HSR [142,143,146,147,148].
After HS, the increase in abscisic acid (ABA) levels activates the transcription factors, such as AREB/ABFs, controlling the ABA-dependent gene expression that stimulates ROS production or improves antioxidant machinery [35,149,150,151,152]. For instance, the interaction of flowering control locus A, a plant-specific RNA-binding protein of the autonomous flowering pathway, with ABA-INSENSITIVE 5 increases thermotolerance through the transcriptional regulation of the PER1 gene encoding 1Cys-PRX [149]. The ABA-induced ERF74 induces RBOHD transcription, increasing ROS production, and enhancing HS tolerance [153] (Figure 2). The transcriptional co-activator MBF1c, as well as transcription factors of DREB2A and AP2-EREBPs families, induced by both ROS and ABA, play key roles in thermotolerance acquisition [103,154,155]. In the interaction between the regulative pathways of ROS, ABA, and auxin, the ROS-scavenger enzyme APX6 is involved in the HS protection of seeds during germination [156].
In the HSR of tomato plants, a crosstalk between ABA and brassinosteroids (BRs) mediated by ROS occurs. BRs, by activating RBOHs, stimulate a transient H2O2 accumulation, which increases the ABA levels and results in additional rises in H2O2 and enhanced HS tolerance [146]. In BR-mediated tolerance to HS, the regulator BZR1 improves the production of apoplastic H2O2 that regulates the receptor-like kinase FERONIA, resulting in HS tolerance [157] (Figure 2).
During heat stress, salicylic acid (SA) also increases and protects PSII from the high ROS levels [158,159]. The Arabidopsis mutants sid2, which are deficient in SA signaling, accumulate elevated ROS levels and reveal increased HS sensitivity, showing that SA has a crucial role in thermotolerance [154,155,160]. In tomatoes and barley, SA enhances antioxidant systems, increasing ROS scavenging and heat tolerance [161,162]. Moreover, treatment with SA improves the activity of antioxidant enzymes and by controlling ROS level, it relieves the heat-dependent decline in pollen viability and floret fertility [163,164] (Figure 2).
Ethylene (ET) production during HS has been related to the beginning of leaf senescence and reduction in pollen growth and germination, spike fertility, and grain weight, which are survival strategies to preserve the optimal progeny capacity under high temperatures [165]. Under HS, ROS can act in concert with ET in a self-intensifying feedback loop where ET stimulates H2O2 accumulation, which in turn improves ET production, initiating leaf senescence or chlorosis [142,166]. ET can additionally control ROS metabolism by altering antioxidant enzymes [167]. Rice seedlings treated with the ET precursor, 1-aminocyclopropane-1-caroboxylic, recover the activities of CAT, APX, and POD and show a decreased H2O2 accumulation and cell damage under HS [168]. Moreover, ET-induced ERFs are involved in redox regulation, while oxidative stress induces ERFs [169]. In tobacco, ERF3 contributes to lower ROS accumulation, enhancing abiotic stress tolerance [170], while in Arabidopsis, ERF6 controls ROS signaling during HS. These data suggest that ERFs control HS tolerance by modulating ROS homeostasis [171] (Figure 2).
Heat stress reduces the endogenous levels of cytokinins (CKs) [172]. However, the treatment with CKs usually improves HS tolerance by the increase in antioxidant enzymes and HSPs [173]; consistently, the inhibition of CKs degradation improves HS resistance, principally by the enhancement of antioxidant enzymes [173,174,175] (Figure 2).
6. Heat Stress-Dependent Oxidation of Cellular Environment
The increase in temperature provokes an enhanced ROS production and accumulation that temporarily oxidizes the cellular environment, triggering downstream signaling cascades. Because of their high reactivity, when ROS escape antioxidant-mediated scavenging, they react with biomolecules causing lipid peroxidation, protein oxidation, and other protein modification, which often might serve as signaling factors that trigger the effective defense response [176]. Thus, not only ROS accumulation, but also the HS-related redox changes are closely associated with the HSR that supports thermotolerance. Indeed, the inhibited expression of scavenger enzymes leads to an increase in the oxidative power required to activate the HSFs in the initial steps of HSR [46]. Consistently, miRNA398, which downregulates the CSD1, CSD2, and CCS genes, achieves its highest expression within 2 h from HS; moreover, heat improves tolerance to subsequent high temperature stress [77,177]. In Arabidopsis, a transient oxidative burst, occurring immediately after HS and which is due to a reduction in APX and CAT activity, causes a transient oxidation of the cellular environment, which helps trigger a signaling cascade that protects plants from HS. Accordingly, after 3 h of physiological recovery from HS, both non-enzymatic and enzymatic antioxidants significantly increase, lowering ROS accumulation and preventing oxidative damage. On the other hand, gun1 mutants that fail to increase ROS levels promptly exhibit impaired HSR and accumulate ROS and oxidative damage during physiological recovery at the growth temperature. Thus, GUN1 seems to be required to oxidize the cellular environment, participating in the acquisition of basal thermotolerance through the redox-dependent plastid-to-nucleus communication [113].
A recent study on HS response, performed using the redox-sensitive green fluorescent protein, roGFP2, clearly demonstrated that HS, by triggering large shifts in the glutathione redox potentials of cells, causes a huge rise in the extent of oxidation of both cytosol and nuclear compartments [105]. In this context, the analysis of transcript profiles indicates a very specific redox-processing response to HS, since APX2 was the only increased antioxidant transcript; moreover, the enhanced expression of ZAT12 in the seedlings after HS suggests altered oxidative stress signaling [105,178].
Cellular environment oxidation permits redox-dependent changes to proteins that include conformational changes of proteins involved in HSR, as well as modifications to transcription factors or associated proteins that can also trigger nuclear translocation [179]. Cysteine (Cys) residues of proteins may act as sensors of redox changes triggering different post-translation modifications, which in turn modulate redox-dependent pathways until a plant’s response to environmental insults is achieved [180,181]. Thiol residues could be deprotonated into a thiolate residue (R-S-), which leads to successive oxidations to sulfenic, sulfinic, and sulfonic acids [182]. Thiol groups can also form a disulfide bridge (S-S) or react with reactive nitrogen species (RNS), resulting in S-nitrosylation. Depending on their nature, most of these thiol modifications can be reversed by dedicated thiol reduction systems (thioredoxin-Trx, glutaredoxins-GRX, and S-nitrosoglutathione reductase GSNOR).
Exposure to high temperatures usually leads to an increase in RNS, which are essential for the acclimatization mechanism [183,184]. It has been demonstrated that the exogenous application of nitric oxide (NO) to plants subjected to HS promotes the activation of enzymatic and non-enzymatic defense systems to oxidative stress [122,185,186]. In contrast, treatment with NO scavengers eliminates the beneficial effects caused by NO, corroborating the idea that NO is involved in tolerance to high temperatures [185,187]. S-nitrosoglutathione reductase, GSNOR1, controls the turnover of GSNO under stress conditions [188,189]. In the Arabidopsis thaliana thermotolerance-defective mutant, hot5 (sensitive to hot temperature 5), with HOT5 encoding GSNOR1, the loss of enzymatic activity increases s-nitrosothiols and leads to a high heat sensitivity [190]. Thus, the absence of GSNOR1 activity undeniably affects NO/GSNO levels, which might correlate with heat sensitivity and thermotolerance [190,191].
7. Redox-Dependent Changes of Proteins Involved in the Heat Stress Response
ROS work efficiently in cell signaling through chemical reactions with specific target protein residues that lead to protein modifications. The primary targets of ROS are proteins containing amino acids with Cys residues. The redox changes that involve thiol-disulfide switches bring about various effects, such as the induction of protein conformational changes, the alteration of protein–protein interactions, and changes in protein subcellular localization [192].
In mild HS, any imbalance in the redox state of the chloroplast may lead to the inhibition of a specific redox-sensitive kinase (STN7), a transmembrane protein with catalytic domains containing two conserved Cys residues. STN7 phosphorylates the light-harvesting complex II (LHCII), which migrates to PSI and shifts the excitation energy to PSI. Thus, the redox-dependent STN7 inhibition, which alters LHCII phosphorylation, does not permit the formation of the PSI-LHCII complex with a consequent alteration in the thylakoid ultrastructure and a strong depletion in the photosynthetic rates [193]. Interestingly, one of the four phosphorylation sites in the C-terminal region of STN7 is phosphorylated by CK2, which is a chloroplast nuclear casein kinase regulated by the redox signal [194].
Under heat stress, many proteins undergo redox-dependent conformational changes that are associated with the acquisition of a new function (Table 1).

Table 1.
List of redox-dependent conformational changes and functions of proteins under heat stress.
An interesting example showing how changes in the protein oxidation state lead to a switch of function during HS is represented by the Arabidopsis h-type thioredoxin (Trx), AtTrx-h3. This protein forms several protein structures varying from low and oligomeric to high molecular weight (HMW) complexes, performing the function of disulfide reductase in the low molecular weight (LMW), but acting as a molecular chaperone in the HMW complexes. The structures of AtTrx-h3 are regulated both by HS and the redox state. Two active Cys residues in AtTrx-h3 are necessary for disulfide reductase activity, but not for the chaperone function. The reduction of AtTrx changes the HMW structures into LMW, while oxidation permits the formation of the HMW structure. It has been demonstrated that AtTrx-h3 confers HS tolerance in Arabidopsis, mainly in the oxidized state, through its chaperone function [195].
Another redox protein revealing an analogous regulation mode of AtTrx-h3 is the C-type NADPH-dependent Trx reductase (NTRC), a member of the plant-specific NADPH-dependent Trx reductase family, which comprises an N-terminal Trx reductase domain and a C-terminal Trx domain. This protein works as an efficient electron donor to 2-Cys peroxiredoxins [200,201]. The structure of NTRC shows different oligomeric conformations in various plant species [202,203,204], acting as a disulfide reductase and a foldase and holdase chaperone, depending on the protein structures. The LMW structures display greater disulfide reductase and foldase chaperone activities, while HMW ones have stronger activity as a holdase chaperone. Heat stress mediates the redox-dependent oligomeric changes of NTRC, converting LMW proteins into HMW complexes and progressively enhancing the holdase chaperone activity, which confers higher thermotolerance [196].
The 2-Cys peroxiredoxins (Prx) belong to the Prxs family, which are peroxidases that reduce H2O2, peroxynitrite, and alkyl hydroperoxides [197,205,206]. The overexpression of 2-Cys Prx in Festuca arundinacea, reducing oxidative damage under high temperature, confers thermotolerance [207]. The 2-Cys Prx is essential in protecting the cellular environment from oxidative stress, since under physiological conditions it forms LMW structures such as dimers through disulfide bonds and removes H2O2. However, an increase in ROS under HS leads to an overoxidation of the 2-Cys Prx, which, aggregating into HMW structures, loses its predominant function of antioxidant enzyme and gains the chaperone one, which is essential to mediate the proper folding of proteins in response to HS [197]. In this way, Prx can act as a sort of “dam” to H2O2 flow. In stress conditions, Prx inactivation may be a way to leave sufficient time for H2O2 to mediate stress signaling and to promote a variety of redox-dependent processes [208,209].
Redox-dependent conformational changes associated with different functions in response to HS also occur in the Arabidopsis glutaredoxin GRXS17: this member of the thiol reductase families in Arabidopsis is a nucleocytosolic monothiol GRX, containing an N-terminal thioredoxin domain and three conserved monocysteinic active sites that coordinate three iron–sulfur (Fe-S) clusters in a glutathione-dependent manner. GRXS17 is involved in the maturation of cytosolic and nuclear Fe–S proteins. However, GRXS17 also shows foldase and redox-dependent holdase activities. After HS, this protein loses its Fe–S clusters, forming disulfide bonds between Cys in the Trx domains, thus leading to the formation of HMW structures, which shows holdase activity in vitro. Therefore, upon HS, GRXS17 shows a redox-dependent chaperone activity, interacting with different proteins and protecting them from heat damage [198]. Consistently, the expression of Arabidopsis GRXS17 in maize confers thermotolerance due to an enhanced chaperone activity. Under heat stress, the GRXS17-overexpressing maize lines show a reduced protein damage and higher thermotolerance in the reproductive stages with an increase in grain production when compared to non-transgenic lines [210].
The Arabidopsis non-expressor of pathogenesis-related protein 1 (AtNPR1), which plays a significant role in plant systemic acquired immune responses [211], has also been shown to have a redox-dependent chaperone activity that is required to protect plants from HS. As a result of its structural switch into oligomeric form, the recombinant AtNPR1 works as a protein chaperone. The structural changes and the chaperone activity are dependent on its redox state since dithiothreitol treatment dissociates its structure to a monomer and reduces its chaperone activity [199].
8. Redox-Dependent Changes of Proteins Impacting Gene Expression during Heat Stress Response
As a result of HS, the glutathione redox potentials of Arabidopsis cell nuclei are greatly changed, indicating nuclear oxidation [105]. Since transcript profiles of the control and heat-treated seedlings showed significant differences in HSPs, mitochondrial proteins, transcription factors, and other nuclear-localized components, the authors suggested that nuclear oxidation plays a key role in both the genetic and epigenetic control of HSR [105]. Nuclear oxidation can be responsible for the epigenome changes occurring in heat-induced priming [212], because many of the enzymes involved in histone and DNA methylation are subject to redox regulation [213,214]. Furthermore, nuclear oxidation may be responsible for the regulation of redox-sensitive transcription factors (Figure 3) [215].
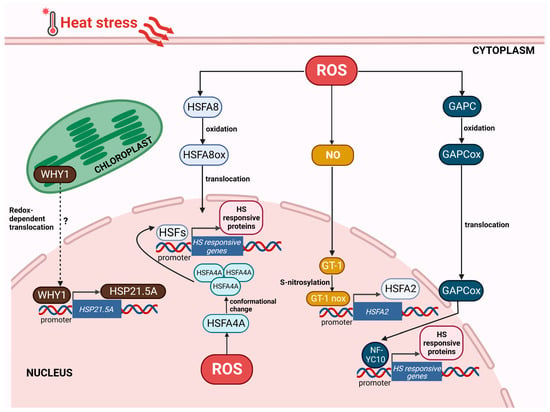
Figure 3.
Redox-dependent changes of proteins impacting gene expression in response to heat stress. Heat stress, triggering oxidation of cellular environment, causes redox changes in proteins involved in the induction of heat stress response. The oxidation of HSFA8 and GAPC permits these proteins to move into the nucleus: HSFA8 directly and GAPC by the interaction with NF-YC10 promote the expression of heat stress-responsive (HSR) genes. S-nitrosylation of GT-1, triggered by ROS-dependent NO accumulation, induces the transcription of HsfA2. Nuclear redox oxidation is sensed by HSFA4A, which is stabilized and activated, inducing HSR genes. In the chloroplast, over-reduction in the components of the thylakoid electron transport chain might provoke the redox-dependent translocation of WHY1 in the nucleus, where the protein activates the transcription of HSP21.5A. Abbreviations: HSF, heat shock factor; GAPC, glyceraldehyde-3-phosphate dehydrogenase, cytosolic; NF-YC10, nuclear factor Y, subunit C10; and WHY1, Whirly1. Created with BioRender.com; accessed on 3 February 2023.
It is known that HSFs sense the variation in temperature and in turn, modulate the expression of HSPs and other stress-associated genes [216]. In vitro stress treatments cause monomer-to-trimer transitions of HSFs, while the presence of the reducing agent dithiothreitol reverses this action. HSFs have been proposed to participate in peroxide sensing, as H2O2 can stabilize HSF trimers by reversible oxidation of Cys residues and by forming Cys–Cys bonds [217,218]. HSFA1a, the master regulator of HSR [219,220], contains one Cys residue located at the N-terminal portion of the trimerization domain [221]. N-terminal deletions of HSFA1a negatively affect H2O2 sensing, suggesting that trimerizations are induced by conformational changes [222]. However, the N-terminal deletion of HSFA1a does not inhibit heat sensing. This shows that, despite a common dependency on oxidative activity, the activation of this transcription factor is stress-specifically regulated [217]. The ROS signal has been shown to induce two signaling pathways associated with NO: ROS generated by RbohB and RbohD trigger NO accumulation [126] and the NO signal activates CaM3, which then leads to DNA binding of HSFs [223]. Full activation of HSFA1 seems to involve many processes rising from several signaling pathways [224]. However, recent data have shown that the binding of HSFA1a to its targets can be obtained independently of HS, by altering the plant redox state through light-dependent chloroplast signaling [39]. On the other hand, trimerization of HSFA4A is strictly dependent on the redox environment, since the intramolecular interactions of HSFA4A are weakened when the three conserved Cys residues are replaced with Ala [225,226]. In Arabidopsis, the MAPKs that phosphorylate this HSF, namely MAPK3 and MAPK6, have also been shown to be activated by H2O2 [225]. These results suggest that HSFA4A can function as redox sensors in stress conditions.
Recently, it has been reported that NO bursts under HS induce S-nitrosylation at Cys324 or Cys347 of the trihelix transcription factor GT-1, which promotes its binding to the HSFA2 promoter (Figure 3). GT-1s loss of function disturbs HSFA2 activation and heat tolerance, revealing that GT-1 is a requested mediator connecting signal perception to the triggering of cellular heat responses [227].
In the HSR, ROS affects nuclear gene expression due to the nuclear translocation of cytoplasmatic transcription factors or associated proteins, subjected to redox-dependent conformational changes (Figure 3). Indeed, cytosol oxidation can trigger the movement of redox-sensitive proteins into the nucleus [179].
HSFA8 shows redox-dependent translocation to nuclei, which is associated with the oxidation of key Cys residues and the formation of disulfide bonds [228]. Mutagenesis of the conserved residues, Cys24 and Cys269, blocks the translocation of HSFA8 to the nucleus, suggesting that this HSF functions as a redox-sensing transcription factor within the stress-responsive transcriptional network (Figure 3).
Cytosolic glyceraldehyde-3-phosphate dehydrogenase (GAPC) is a glycolytic enzyme that is also involved in plant responses to oxidative stress, which could result from abiotic and biotic challenges [229]. One mechanism for the action of GAPC in HSR is stress-induced nuclear translocation [230,231,232]. Under HS, cytosol oxidation triggers GAPC movement to the nucleus, where it interacts with the transcription factor NF-YC10 to promote heat tolerance through the expression of heat-inducible genes [233]. The overexpression of GAPC improves heat tolerance and the effect is removed when NF-YC10 is lacking, heat-induced nuclear accumulation of GAPC is repressed, or the GAPC-NF-YC10 interaction is disrupted. Moreover, the overexpression of GAPC also improves the binding ability of NF-YC10 to its target promoter (Figure 3) [233].
WHIRLY1 (WHY1) is a chloroplast and nucleus-localized plant-specific DNA- and RNA-binding protein that carries out several cellular tasks [234,235]. In the chloroplast, WHY1 mainly works in safeguarding the genome, but it also controls the photochemical activity of PSI [236,237,238]. In the nucleus, WHY1 functions as a transcriptional activator that can bind the elicitor response element of the pathogenesis-related gene PR-10 in Arabidopsis [239,240], or the HvS40 promoter in barley, regulating plant senescence [237]. Recently, a clear association between WHY1 and HSR has been reported. In tomato, HS induces WHY1 expression, and the protein binds the HSE in the promoter of the ER-localized small heat shock protein, HSP21.5A, to activate its transcription (Figure 3). WHY1 overexpression increased thermotolerance; conversely, the inhibition of WHY1 expression leads to an impaired HSR with reduced membrane stability and soluble sugar content and increased ROS accumulation [241]. WHY1 has been proposed as a sensor of photosynthetic apparatus redox changes and represents an ideal candidate for retrograde signaling. In physiological conditions, WHY1 is closely associated with thylakoid membranes. The over-reduction in one or more components of the thylakoid electron transport system, such as the PQ pool/cytochrome b6f complex, may destabilize the oligomeric WHY1 structure on the thylakoid membrane, leading to a release of monomeric proteins, which are then translocated to the nucleus [242].
9. Conclusions
Progressively higher temperatures, which are intensified by global warming, may have a strong impact on plant yield in the future. Thus, the study of molecular processes that allow plants to overcome climate change is essential to reduce its negative influence on growth and productivity. Plants use multiple intracellular signaling pathways to sense and respond to high temperature. The effects of heat stress are often correlated with an increase in ROS production occurring in organelles as well as in the apoplast. Even though ROS can potentially cause damage to plant cells, they may also act as signaling molecules and trigger the adaptative response [44,47]. The different and antagonistic role of these reactive molecules depends on a subtle balance between production and scavenging. Therefore, the enhancement of antioxidants is a key strategy to confer heat stress tolerance [53]. However, the study of antioxidant systems in HSR requires further investigation, since antioxidants display a complex and intricate network, which depends on the characteristics of the plant and heat stress, and on the different and sometime antagonistic behaviors of the various antioxidant isoenzymes.
ROS accumulation has been shown to be a requirement for the acquisition of plant thermotolerance. Thus, the research on the sites of ROS generation, the mechanisms that regulate ROS transport, as well as how ROS signaling is interconnected in different compartments is crucial for the comprehension of HS tolerance. The knowledge of the complex dynamics of ROS networks in the different cell compartments can be strongly enhanced using genetically encoded ROS and redox sensors.
Chloroplasts have been shown to play an important role in heat-induced ROS accumulation and the subsequent expression of nuclear heat-responsive genes [129]. Recently, it has been suggested that GUN1, a key player in plastid-to-nucleus retrograde signaling, contributes to the acquisition of basal thermotolerance, being involved in ROS accumulation and oxidation of the cellular environment [113], but, further research is needed to determine the mechanisms involved in this signaling pathway.
Not only HS-induced ROS production, but also HS-related redox changes are strongly linked with the HSR that confers thermotolerance [44,105]. In this context, Cys residues play a significant role as sensors of redox changes, triggering different post-translation modifications, which in turn regulate redox-dependent paths, comprising conformational changes of protein involved in HSR, as well as modifications to transcription factors or associated proteins that can also cause nuclear translocation [179,180,181]. The massive protein regulation capability may strongly affect adaptation to HS. Thus, the continuous advancement in proteomic approaches that permits a more accurate recognition of potential modifications will improve understanding of the significant signaling functions of redox changes occurring in HSR. Moreover, since HS also leads to nuclear oxidation [105], determining the redox regulation of nuclear proteins may help to identify the important mechanisms of genetic and epigenetic regulation of thermotolerance.
The use of model systems and the study of HSR in a controlled environment are crucial to elucidate the redox signaling network activated in response to high temperatures. However, improved knowledge about HSR has to come from experimental where environmental changes need to be simulated as realistically as possible. Indeed, it should be considered that under field conditions, HS is frequently accompanied by other types of stress, such as drought, air humidity, or the presence of pathogens [243,244]. Recent studies reveal that the response of plants to a combination of different abiotic and biotic stresses is unique and cannot be directly extrapolated from simply analyzing each of the different stresses applied individually [103,245,246,247]. Several literature studies have documented the effects of HS combined with other stress factors [160,248,249,250]; nevertheless, comprehensive analyses on the complex network between hormones, ROS, and redox signals in combined stress occurring under complex environmental conditions deserve more attention. Knowledge of how ROS and redox signals combine, and the identification of the numerous compartment-specific redox pathways that are activated might be crucial to modify HSR as a function of HS intensity and duration, and of plant characteristics, in order to help plants deal with climate change.
Author Contributions
Conceptualization, S.F., C.L. and M.C.d.P.; writing—review and editing S.F., C.L., F.V., N.D. and M.C.d.P., supervision, S.F. and M.C.d.P. All authors have read and agreed to the published version of the manuscript.
Funding
This research received no external funding.
Institutional Review Board Statement
Not applicable.
Informed Consent Statement
Not applicable.
Data Availability Statement
No new data were created or analyzed in this study. Data sharing is not applicable to this article.
Conflicts of Interest
The authors declare no conflict of interest.
References
- de Larminat, P. Earth Climate Identification vs. Anthropic Global Warming Attribution. Annu. Rev. Control 2016, 42, 114–125. [Google Scholar] [CrossRef]
- Bailey-Serres, J.; Parker, J.E.; Ainsworth, E.A.; Oldroyd, G.E.D.; Schroeder, J.I. Genetic Strategies for Improving Crop Yields. Nature 2019, 575, 109–118. [Google Scholar] [CrossRef] [PubMed]
- IPCC. 2021: Climate Change 2021: The Physical Science Basis. Contribution of Working Group I to the Sixth Assessment Report of the Intergovernmental Panel on Climate Change; Masson-Delmotte, V.P., Zhai, A., Pirani, S.L., Connors, C., Péan, S., Berger, N., Caud, Y., Chen, L., Goldfarb, M.I., Eds.; Cambridge University Press: Cambridge, UK; New York, NY, USA, In Press. [CrossRef]
- Wahid, A.; Gelani, S.; Ashraf, M.; Foolad, M.R. Heat Tolerance in Plants: An Overview. Environ. Exp. Bot. 2007, 61, 199–223. [Google Scholar] [CrossRef]
- Wolkovich, E.M.; Cook, B.I.; Allen, J.M.; Crimmins, T.M.; Betancourt, J.L.; Travers, S.E.; Pau, S.; Regetz, J.; Davies, T.J.; Kraft, N.J.B.; et al. Warming Experiments Underpredict Plant Phenological Responses to Climate Change. Nature 2012, 485, 494–497. [Google Scholar] [CrossRef]
- Zhao, J.; Lu, Z.; Wang, L.; Jin, B. Plant Responses to Heat Stress: Physiology, Transcription, Noncoding Rnas, and Epigenetics. Int. J. Mol. Sci. 2021, 22, 117. [Google Scholar] [CrossRef]
- Wang, L.; Ma, K.B.; Lu, Z.G.; Ren, S.X.; Jiang, H.R.; Cui, J.W.; Chen, G.; Teng, N.J.; Lam, H.M.; Jin, B. Differential Physiological, Transcriptomic and Metabolomic Responses of Arabidopsis Leaves under Prolonged Warming and Heat Shock. BMC Plant Biol. 2020, 20, 86. [Google Scholar] [CrossRef]
- Jagadish, S.V.K.; Murty, M.V.R.; Quick, W.P. Rice Responses to Rising Temperatures—Challenges, Perspectives and Future Directions. Plant Cell Environ. 2015, 38, 1686–1698. [Google Scholar] [CrossRef]
- Ullah, A.; Nadeem, F.; Nawaz, A.; Siddique, K.H.M.; Farooq, M. Heat Stress Effects on the Reproductive Physiology and Yield of Wheat. J. Agron. Crop Sci. 2022, 208, 1–17. [Google Scholar] [CrossRef]
- Ahammed, G.J.; Yu, J.Q. Plant Hormones under Challenging Environmental Factors; Springer Dordrecht: Berlin, Germany, 2016; pp. 1–269. [Google Scholar] [CrossRef]
- Peck, S.; Mittler, R. Plant Signaling in Biotic and Abiotic Stress. J. Exp. Bot. 2020, 71, 1649–1651. [Google Scholar] [CrossRef]
- Yadav, S.; Modi, P.; Dave, A.; Vijapura, A.; Dish, P.; Patel, M. Effect of Abiotic Stress on Crops. Sustain. Crop Prod. 2020, 3. [Google Scholar] [CrossRef]
- Qi, J.; Song, C.P.; Wang, B.; Zhou, J.; Kangasjärvi, J.; Zhu, J.K.; Gong, Z. Reactive Oxygen Species Signaling and Stomatal Movement in Plant Responses to Drought Stress and Pathogen Attack. J. Integr. Plant Biol. 2018, 60, 805–826. [Google Scholar] [CrossRef] [PubMed]
- Gull, A.; Lone, A.A.; Wani, N.U.I. Biotic and Abiotic Stresses in Plants. In Abiotic and Biotic Stress in Plants; IntechOpen: London, UK, 2019; pp. 1–19. [Google Scholar] [CrossRef]
- Hassan, M.U.; Chattha, M.U.; Khan, I.; Chattha, M.B.; Barbanti, L.; Aamer, M.; Iqbal, M.M.; Nawaz, M.; Mahmood, A.; Ali, A.; et al. Heat Stress in Cultivated Plants: Nature, Impact, Mechanisms, and Mitigation Strategies—A Review. Plant Biosyst. 2021, 155, 211–234. [Google Scholar] [CrossRef]
- Hasanuzzaman, M.; Nahar, K.; Alam, M.M.; Roychowdhury, R.; Fujita, M. Physiological, Biochemical, and Molecular Mechanisms of Heat Stress Tolerance in Plants. Int. J. Mol. Sci. 2013, 14, 9643–9684. [Google Scholar] [CrossRef] [PubMed]
- Giri, A.; Heckathorn, S.; Mishra, S.; Krause, C. Heat Stress Decreases Levels of Nutrient-Uptake and -Assimilation Proteins in Tomato Roots. Plants 2017, 6, 6. [Google Scholar] [CrossRef]
- Lippmann, R.; Babben, S.; Menger, A.; Delker, C.; Quint, M. Development of Wild and Cultivated Plants under Global Warming Conditions. Curr. Biol. 2019, 29, R1326–R1338. [Google Scholar] [CrossRef]
- Sharkey, T.D. Effects of Moderate Heat Stress on Photosynthesis: Importance of Thylakoid Reactions, Rubisco Deactivation, Reactive Oxygen Species, and Thermotolerance Provided by Isoprene. Plant Cell Environ. 2005, 28, 269–277. [Google Scholar] [CrossRef]
- Allakhverdiev, S.I.; Kreslavski, V.D.; Klimov, V.V.; Los, D.A.; Carpentier, R.; Mohanty, P. Heat Stress: An Overview of Molecular Responses in Photosynthesis. Photosynth. Res. 2008, 98, 541–550. [Google Scholar] [CrossRef]
- Murata, N.; Takahashi, S.; Nishiyama, Y.; Allakhverdiev, S.I. Photoinhibition of Photosystem II under Environmental Stress. Biochim. Biophys. Acta Bioenerg. 2007, 1767, 414–421. [Google Scholar] [CrossRef]
- Takahashi, S.; Murata, N. How Do Environmental Stresses Accelerate Photoinhibition? Trends Plant Sci. 2008, 13, 178–182. [Google Scholar] [CrossRef]
- Yamamoto, Y.; Ryota, A.E.; Ae, A.; Yoshioka, M.; Mahbuba, A.E.; Ae, K.; Komayama, K.; Daichi, A.E.; Ae, T.; Yamashita, A.; et al. Quality Control of Photosystem II: Impact of Light and Heat Stresses. Photosynth. Res. 2008, 98, 589–608. [Google Scholar] [CrossRef]
- Chen, K.; Zhang, M.; Zhu, H.; Huang, M.; Zhu, Q.; Tang, D.; Han, X.; Li, J.; Sun, J.; Fu, J. Ascorbic Acid Alleviates Damage from Heat Stress in the Photosystem II of Tall Fescue in Both the Photochemical and Thermal Phases. Front. Plant Sci. 2017, 8, 1373. [Google Scholar] [CrossRef] [PubMed]
- Yamori, W.; Takahashi, S.; Makino, A.; Price, G.D.; Badger, M.R.; von Caemmerer, S. The Roles of ATP Synthase and the Cytochrome b6/f Complexes in Limiting Chloroplast Electron Transport and Determining Photosynthetic Capacity. Plant Physiol. 2011, 155, 956–962. [Google Scholar] [CrossRef] [PubMed]
- Sharkey, T.D.; Zhang, R. High Temperature Effects on Electron and Proton Circuits of Photosynthesis. J. Integr. Plant Biol. 2010, 52, 712–722. [Google Scholar] [CrossRef] [PubMed]
- Mathur, S.; Jajoo, A.; Mehta, P.; Bharti, S. Analysis of Elevated Temperature-induced Inhibition of Photosystem II Using Chlorophyll a Fluorescence Induction Kinetics in Wheat Leaves (Triticum Aestivum). Plant Biol. 2011, 13, 1–6. [Google Scholar] [CrossRef]
- Perdomo, J.A.; Capó-Bauçà, S.; Carmo-Silva, E.; Galmés, J. Rubisco and Rubisco Activase Play an Important Role in the Biochemical Limitations of Photosynthesis in Rice, Wheat, and Maize under High Temperature and Water Deficit. Front. Plant Sci. 2017, 8, 490. [Google Scholar] [CrossRef]
- Khanna-Chopra, R. Leaf Senescence and Abiotic Stresses Share Reactive Oxygen Species-Mediated Chloroplast Degradation. Protoplasma 2012, 249, 469–481. [Google Scholar] [CrossRef]
- Rossi, S.; Burgess, P.; Jespersen, D.; Huang, B. Heat-Induced Leaf Senescence Associated with Chlorophyll Metabolism in Bentgrass Lines Differing in Heat Tolerance. Crop Sci. 2017, 57, S-169–S-178. [Google Scholar] [CrossRef]
- Liang, M.H.; Jiang, J.G.; Wang, L.; Zhu, J. Transcriptomic Insights into the Heat Stress Response of Dunaliella Bardawil. Enzyme Microb. Technol. 2020, 132, 109436. [Google Scholar] [CrossRef]
- Wang, Q.L.; Chen, J.H.; He, N.Y.; Guo, F.Q. Metabolic Reprogramming in Chloroplasts under Heat Stress in Plants. Int. J. Mol. Sci. 2018, 19, 849. [Google Scholar] [CrossRef]
- Prasch, C.M.; Sonnewald, U. Signaling Events in Plants: Stress Factors in Combination Change the Picture. Environ. Exp. Bot. 2015, 114, 4–14. [Google Scholar] [CrossRef]
- Haider, S.; Iqbal, J.; Naseer, S.; Yaseen, T.; Shaukat, M.; Bibi, H.; Ahmad, Y.; Daud, H.; Abbasi, N.L.; Mahmood, T. Molecular Mechanisms of Plant Tolerance to Heat Stress: Current Landscape and Future Perspectives. Plant Cell Rep. 2021, 40, 2247–2271. [Google Scholar] [CrossRef] [PubMed]
- Zhu, J.K. Abiotic Stress Signaling and Responses in Plants. Cell 2016, 167, 313–324. [Google Scholar] [CrossRef] [PubMed]
- Bigot, S.; Buges, J.; Gilly, L.; Jacques, C.; Le Boulch, P.; Berger, M.; Delcros, P.; Domergue, J.B.; Koehl, A.; Ley-Ngardigal, B.; et al. Pivotal Roles of Environmental Sensing and Signaling Mechanisms in Plant Responses to Climate Change. Glob. Chang. Biol. 2018, 24, 5573–5589. [Google Scholar] [CrossRef] [PubMed]
- Raza, A. Metabolomics: A Systems Biology Approach for Enhancing Heat Stress Tolerance in Plants. Plant Cell Rep. 2020, 41, 741–763. [Google Scholar] [CrossRef] [PubMed]
- Sharma, A.; Kumar, V.; Shahzad, B.; Ramakrishnan, M.; Singh Sidhu, G.P.; Bali, A.S.; Handa, N.; Kapoor, D.; Yadav, P.; Khanna, K.; et al. Photosynthetic Response of Plants Under Different Abiotic Stresses: A Review. J. Plant Growth Regul. 2020, 39, 509–531. [Google Scholar] [CrossRef]
- Dickinson, P.J.; Kumar, M.; Martinho, C.; Yoo, S.J.; Lan, H.; Artavanis, G.; Charoensawan, V.; Schöttler, M.A.; Bock, R.; Jaeger, K.E.; et al. Chloroplast Signaling Gates Thermotolerance in Arabidopsis. Cell Rep. 2018, 22, 1657–1665. [Google Scholar] [CrossRef] [PubMed]
- Fey, V.; Wagner, R.; Bräutigam, K.; Wirtz, M.; Hell, R.; Dietzmann, A.; Leister, D.; Oelmüller, R.; Pfannschmidt, T. Retrograde Plastid Redox Signals in the Expression of Nuclear Genes for Chloroplast Proteins of Arabidopsis Thaliana. J. Biol. Chem. 2005, 280, 5318–5328. [Google Scholar] [CrossRef]
- Sun, A.-Z.; Guo, F.-Q. Chloroplast Retrograde Regulation of Heat Stress Responses in Plants. Front. Plant Sci. 2016, 7, 398. [Google Scholar] [CrossRef]
- Haslbeck, M.; Vierling, E. A First Line of Stress Defense: Small Heat Shock Proteins and Their Function in Protein Homeostasis. J. Mol. Biol. 2015, 427, 1537–1548. [Google Scholar] [CrossRef]
- Scharf, K.D.; Berberich, T.; Ebersberger, I.; Nover, L. The Plant Heat Stress Transcription Factor (Hsf) Family: Structure, Function and Evolution. Biochim. Biophys. Acta Gene Regul. Mech. 2012, 1819, 104–119. [Google Scholar] [CrossRef]
- Mittler, R.; Zandalinas, S.I.; Fichman, Y.; Van Breusegem, F. Reactive Oxygen Species Signalling in Plant Stress Responses. Nat. Rev. Mol. Cell Biol. 2022, 23, 663–679. [Google Scholar] [CrossRef]
- Suzuki, N. Fine Tuning of ROS, Redox and Energy Regulatory Systems Associated with the Functions of Chloroplasts and Mitochondria in Plants under Heat Stress. Int. J. Mol. Sci. 2023, 24, 1356. [Google Scholar] [CrossRef]
- Driedonks, N.; Xu, J.; Peters, J.L.; Park, S.; Rieu, I. Multi-Level Interactions between Heat Shock Factors, Heat Shock Proteins, and the Redox System Regulate Acclimation to Heat. Front. Plant Sci. 2015, 6, 999. [Google Scholar] [CrossRef] [PubMed]
- Mittler, R. ROS Are Good. Trends Plant Sci. 2017, 22, 11–19. [Google Scholar] [CrossRef] [PubMed]
- Das, K.; Roychoudhury, A. Reactive Oxygen Species (ROS) and Response of Antioxidants as ROS-Scavengers during Environmental Stress in Plants. Front. Environ. Sci. 2014, 2, 53. [Google Scholar] [CrossRef]
- Noctor, G.; Mhamdi, A.; Foyer, C.H. The Roles of Reactive Oxygen Metabolism in Drought: Not so Cut and Dried. Plant Physiol. 2014, 164, 1636–1648. [Google Scholar] [CrossRef] [PubMed]
- Czarnocka, W.; Karpiński, S. Friend or Foe? Reactive Oxygen Species Production, Scavenging and Signaling in Plant Response to Environmental Stresses. Free Radic. Biol. Med. 2018, 122, 4–20. [Google Scholar] [CrossRef] [PubMed]
- Decros, G.; Baldet, P.; Beauvoit, B.; Stevens, R.; Flandin, A.; Colombié, S.; Gibon, Y.; Pétriacq, P. Get the Balance Right: ROS Homeostasis and Redox Signalling in Fruit. Front. Plant Sci. 2019, 10, 1091. [Google Scholar] [CrossRef] [PubMed]
- Rogers, H.; Munné-Bosch, S. Production and Scavenging of Reactive Oxygen Species and Redox Signaling during Leaf and Flower Senescence: Similar but Different. Plant Physiol. 2016, 171, 1560–1568. [Google Scholar] [CrossRef]
- Hasanuzzaman, M.; Bhuyan, M.H.M.B.; Zulfiqar, F.; Raza, A.; Mohsin, S.M.; Al Mahmud, J.; Fujita, M.; Fotopoulos, V. Reactive Oxygen Species and Antioxidant Defense in Plants under Abiotic Stress: Revisiting the Crucial Role of a Universal Defense Regulator. Antioxidants 2020, 9, 681. [Google Scholar] [CrossRef]
- Doğru, A. Effects of Heat Stress on Photosystem II Activity and Antioxidant Enzymes in Two Maize Cultivars. Planta 2021, 253, 85. [Google Scholar] [CrossRef] [PubMed]
- Krieger-Liszkay, A.; Fufezan, C.; Trebst, A. Singlet Oxygen Production in Photosystem II and Related Protection Mechanism. Photosynth. Res. 2008, 98, 551–564. [Google Scholar] [CrossRef] [PubMed]
- Yamashita, A.; Nijo, N.; Pospíšil, P.; Morita, N.; Takenaka, D.; Aminaka, R.; Yamamoto, Y.; Yamamoto, Y. Quality Control of Photosystem II reactive oxygen species are responsible for the damage to photosystem ii under moderate heat stress. J. Biol. Chem. 2008, 283, 28380–28391. [Google Scholar] [CrossRef] [PubMed]
- Prasad, A.; Ferretti, U.; Sedlaová, M.; Pospíšil, P. Singlet Oxygen Production in Chlamydomonas Reinhardtii under Heat Stress. Sci. Rep. 2016, 6, 20094. [Google Scholar] [CrossRef]
- Asada, K. Production and Scavenging of Reactive Oxygen Species in Chloroplasts and Their Functions. Plant Physiol. 2006, 141, 391–396. [Google Scholar] [CrossRef] [PubMed]
- Gupta, R. The Oxygen-Evolving Complex: A Super Catalyst for Life on Earth, in Response to Abiotic Stresses. Plant Signal. Behav. 2020, 15, 1824721. [Google Scholar] [CrossRef]
- Yadav, D.K.; Pospíšil, P. Role of Chloride Ion in Hydroxyl Radical Production in Photosystem II under Heat Stress: Electron Paramagnetic Resonance Spin-Trapping Study. J. Bioenerg. Biomembr. 2012, 44, 365–372. [Google Scholar] [CrossRef]
- Vacca, R.A.; de Pinto, M.C.; Valenti, D.; Passarella, S.; Marra, E.; De Gara, L. Production of Reactive Oxygen Species, Alteration of Cytosolic Ascorbate Peroxidase, and Impairment of Mitochondrial Metabolism Are Early Events in Heat Shock-Induced Programmed Cell Death in Tobacco Bright-Yellow 2 Cells. Plant Physiol. 2004, 134, 1100. [Google Scholar] [CrossRef]
- Valenti, D.; Vacca, R.A.; de Pinto, M.C.; De Gara, L.; Marra, E.; Passarella, S. In the Early Phase of Programmed Cell Death in Tobacco Bright Yellow 2 Cells the Mitochondrial Adenine Nucleotide Translocator, Adenylate Kinase and Nucleoside Diphosphate Kinase Are Impaired in a Reactive Oxygen Species-Dependent Manner. Biochim. Biophys. Acta Bioenerg. 2007, 1767, 66–78. [Google Scholar] [CrossRef]
- Cvetkovska, M.; Alber, N.A.; Vanlerberghe, G.C. The Signaling Role of a Mitochondrial Superoxide Burst during Stress. Plant Signal. Behav. 2013, 8, 1121–1136. [Google Scholar] [CrossRef]
- Rikhvanov, E.G.; Fedoseeva, I.V.; Pyatrikas, D.V.; Borovskii, G.B.; Voinikov, V.K. Role of Mitochondria in the Operation of Calcium Signaling System in Heat-Stressed Plants. Russ. J. Plant Physiol. 2014, 61, 141–153. [Google Scholar] [CrossRef]
- Scafaro, A.P.; Fan, Y.; Posch, B.C.; Garcia, A.; Coast, O.; Atkin, O.K. Responses of Leaf Respiration to Heatwaves. Plant Cell Environ. 2021, 44, 2090–2101. [Google Scholar] [CrossRef] [PubMed]
- Turrens, J.F. Mitochondrial Formation of Reactive Oxygen Species. J. Physiol. 2003, 552, 335–344. [Google Scholar] [CrossRef]
- Wei, S.S.; Niu, W.T.; Zhai, X.T.; Liang, W.Q.; Xu, M.; Fan, X.; Lv, T.T.; Xu, W.Y.; Bai, J.T.; Jia, N.; et al. Arabidopsis MtHSC70-1 Plays Important Roles in the Establishment of COX-Dependent Respiration and Redox Homeostasis. J. Exp. Bot. 2019, 70, 5575–5590. [Google Scholar] [CrossRef]
- Cvetkovska, M.; Vanlerberghe, G.C. Alternative Oxidase Modulates Leaf Mitochondrial Concentrations of Superoxide and Nitric Oxide. New Phytol. 2012, 195, 32–39. [Google Scholar] [CrossRef] [PubMed]
- Borovik, O.A.; Grabelnych, O.I. Mitochondrial Alternative Cyanide-Resistant Oxidase Is Involved in an Increase of Heat Stress Tolerance in Spring Wheat. J. Plant Physiol. 2018, 231, 310–317. [Google Scholar] [CrossRef] [PubMed]
- Sagi, M.; Fluhr, R. Production of Reactive Oxygen Species by Plant. Structure 2006, 141, 336–340. [Google Scholar] [CrossRef]
- Suzuki, N.; Miller, G.; Morales, J.; Shulaev, V.; Torres, M.A.; Mittler, R. Respiratory Burst Oxidases: The Engines of ROS Signaling. Curr. Opin. Plant Biol. 2011, 14, 691–699. [Google Scholar] [CrossRef]
- Miller, G.; Schlauch, K.; Tam, R.; Cortes, D.; Torres, M.A.; Shulaev, V.; Dangl, J.L.; Mittler, R. The Plant NADPH Oxidase RBOHD Mediates Rapid Systemic Signaling in Response to Diverse Stimuli. Sci. Signal. 2009, 2, ra45. [Google Scholar] [CrossRef]
- Larkindale, J.; Hall, J.D.; Knight, M.R.; Vierling, E. Heat Stress Phenotypes of Arabidopsis Mutants Implicate Multiple Signaling Pathways in the Acquisition of Thermotolerance. Plant Physiol. 2005, 138, 882–897. [Google Scholar] [CrossRef]
- Suriyasak, C.; Harano, K.; Tanamachi, K.; Matsuo, K.; Tamada, A.; Iwaya-inoue, M.; Ishibashi, Y. Reactive Oxygen Species Induced by Heat Stress during Grain Fi Lling of Rice (Oryza Sativa L.) Are Involved in Occurrence of Grain Chalkiness. J. Plant Physiol. 2017, 216, 52–57. [Google Scholar] [CrossRef]
- Sun, M.; Jiang, F.; Cen, B.; Wen, J.; Zhou, Y.; Wu, Z. Respiratory Burst Oxidase Homologue-Dependent H2O2 and Chloroplast H2O2 Are Essential for the Maintenance of Acquired Thermotolerance during Recovery after Acclimation. Plant Cell Environ. 2018, 41, 2373–2389. [Google Scholar] [CrossRef]
- Foyer, C.H.; Noctor, G. Redox Signaling in Plants. Antioxidants Redox Signal. 2013, 18, 2087–2090. [Google Scholar] [CrossRef] [PubMed]
- Fan, Y.; Ma, C.; Huang, Z.; Abid, M.; Jiang, S.; Dai, T.; Zhang, W.; Ma, S.; Jiang, D.; Han, X. Heat Priming during Early Reproductive Stages Enhances Thermo-Tolerance to Post-Anthesis Heat Stress via Improving Photosynthesis and Plant Productivity in Winter Wheat (Triticum Aestivum L.). Front. Plant Sci. 2018, 9, 805. [Google Scholar] [CrossRef] [PubMed]
- Wang, X.; Cai, J.; Liu, F.; Dai, T.; Cao, W.; Wollenweber, B.; Jiang, D. Multiple Heat Priming Enhances Thermo-Tolerance to a Later High Temperature Stress via Improving Subcellular Antioxidant Activities in Wheat Seedlings. Plant Physiol. Biochem. PPB 2014, 74, 185–192. [Google Scholar] [CrossRef] [PubMed]
- Djanaguiraman, M.; Boyle, D.L.; Welti, R.; Jagadish, S.V.K.; Prasad, P.V.V. Decreased Photosynthetic Rate under High Temperature in Wheat Is Due to Lipid Desaturation, Oxidation, Acylation, and Damage of Organelles. BMC Plant Biol. 2018, 18, 55. [Google Scholar] [CrossRef]
- Huo, L.; Sun, X.; Guo, Z.; Jia, X.; Che, R.; Sun, Y.; Zhu, Y.; Wang, P.; Gong, X.; Ma, F. MdATG18a Overexpression Improves Basal Thermotolerance in Transgenic Apple by Decreasing Damage to Chloroplasts. Hortic. Res. 2020, 7, 21. [Google Scholar] [CrossRef]
- Gong, H.L.; Chen, Q.Q. Exogenous Sucrose Protects Potato Seedlings Against Heat Stress by Enhancing the Antioxidant Defense System. J. Soil Sci. Plant Nutr. 2021, 21, 1511–1519. [Google Scholar] [CrossRef]
- Khan, A.; Khan, A.L.; Imran, M.; Asaf, S.; Kim, Y.H.; Bilal, S.; Numan, M.; Al-Harrasi, A.; Al-Rawahi, A.; Lee, I.J. Silicon-Induced Thermotolerance in Solanum Lycopersicum L. via Activation of Antioxidant System, Heat Shock Proteins, and Endogenous Phytohormones. BMC Plant Biol. 2020, 20, 248. [Google Scholar] [CrossRef]
- Christou, A.; Filippou, P.; Manganaris, G.A.; Fotopoulos, V. Sodium Hydrosulfide Induces Systemic Thermotolerance to Strawberry Plants through Transcriptional Regulation of Heat Shock Proteins and Aquaporin. BMC Plant Biol. 2014, 14, 42. [Google Scholar] [CrossRef]
- Zhou, Z.; Li, J.; Zhu, C.; Jing, B.; Shi, K.; Yu, J.; Hu, Z. Exogenous Rosmarinic Acid Application Enhances Thermotolerance in Tomatoes. Plants 2022, 11, 1172. [Google Scholar] [CrossRef]
- Kaur, N.; Kaur, J.; Grewal, S.K.; Singh, I. Effect of Heat Stress on Antioxidative Defense System and Its Amelioration by Heat Acclimation and Salicylic Acid Pre-Treatments in Three Pigeonpea Genotypes. Indian J. Agric. Biochem. 2019, 32, 106–110. [Google Scholar] [CrossRef]
- Hameed, A.; Goher, M.; Iqbal, N. Heat Stress-Induced Cell Death, Changes in Antioxidants, Lipid Peroxidation, and Protease Activity in Wheat Leaves. J. Plant Growth Regul. 2012, 31, 283–291. [Google Scholar] [CrossRef]
- de Pinto, M.C.; Locato, V.; Paradiso, A.; De Gara, L. Role of Redox Homeostasis in Thermo-Tolerance under a Climate Change Scenario. Ann. Bot. 2015, 116, 487–496. [Google Scholar] [CrossRef] [PubMed]
- Gupta, N.K.; Khan, A.; Maheshwari, A.; Narayan, S.; Chhapola, O.P.; Arora, A.; Singh, G. Effect of Post Anthesis High Temperature Stress on Growth, Physiology and Antioxidative Defense Mechanisms in Contrasting Wheat Genotypes. Indian J. Plant Physiol. 2015, 20, 103–110. [Google Scholar] [CrossRef]
- Khan, N.A.; Khan, S.; Naz, N.; Shah, M. Effect of Heat Stress on Growth, Physiological and Biochemical Activities of Wheat (Triticum Aestivum L.). Int. J. Biosci. 2017, 11, 173–183. [Google Scholar] [CrossRef]
- Rani, B.; Kumari, N.; Pooja, D.; Jain, V.; Dhawan, K.; Monika, R.A.; Kumar, A.; Sheoran, P. Antioxidative System as Influenced by High Temperature Stress in Brassica Juncea (L) Czern & Coss. Curr. Trends Biotechnol. Pharm. 2016, 10, 118–125. [Google Scholar]
- Yang, Z.; Mhamdi, A.; Noctor, G. Analysis of Catalase Mutants Underscores the Essential Role of CATALASE2 for Plant Growth and Day Length-Dependent Oxidative Signalling. Plant Cell Environ. 2019, 42, 688–700. [Google Scholar] [CrossRef]
- Ono, M.; Isono, K.; Sakata, Y.; Taji, T. Biochemical and Biophysical Research Communications CATALASE2 Plays a Crucial Role in Long-Term Heat Tolerance of Arabidopsis Thaliana. Biochem. Biophys. Res. Commun. 2021, 534, 747–751. [Google Scholar] [CrossRef]
- Hasanuzzaman, M.; Nahar, K.; Alam, M.M.; Fujita, M. Modulation of Antioxidant Machinery and the Methylglyoxal Detoxification System in Selenium-Supplemented Brassica Napus Seedlings Confers Tolerance to High Temperature Stress. Biol. Trace Elem. Res. 2014, 161, 297–307. [Google Scholar] [CrossRef]
- Maruta, T.; Sawa, Y.; Shigeoka, S.; Ishikawa, T. Diversity and Evolution of Ascorbate Peroxidase Functions in Chloroplasts: More than Just a Classical Antioxidant Enzyme? Plant Cell Physiol. 2016, 57, 1377–1386. [Google Scholar] [CrossRef] [PubMed]
- Volkov, R.A.; Panchuk, I.I.; Mullineaux, P.M.; Schöffl, F. Heat Stress-Induced H2O2 Is Required for Effective Expression of Heat Shock Genes in Arabidopsis. Plant Mol. Biol. 2006, 61, 733–746. [Google Scholar] [CrossRef]
- Panchuk, I.I.; Volkov, R.A.; Schöffl, F. Heat Stress- and Heat Shock Transcription Factor-Dependent Expression and Activity of Ascorbate Peroxidase in Arabidopsis. Plant Physiol. 2002, 129, 838–853. [Google Scholar] [CrossRef] [PubMed]
- Schramm, F.; Ganguli, A.; Kiehlmann, E.; Englich, G.; Walch, D.; Von Koskull-Döring, P. The Heat Stress Transcription Factor HsfA2 Serves as a Regulatory Amplifier of a Subset of Genes in the Heat Stress Response in Arabidopsis. Plant Mol. Biol. 2006, 60, 759–772. [Google Scholar] [CrossRef] [PubMed]
- Reddy, R.A.; Kumar, B.; Reddy, P.S.; Mishra, R.N.; Mahanty, S.; Kaul, T.; Nair, S.; Sopory, S.K.; Reddy, M.K. Molecular Cloning and Characterization of Genes Encoding Pennisetum Glaucum Ascorbate Peroxidase and Heat-Shock Factor: Interlinking Oxidative and Heat-Stress Responses. J. Plant Physiol. 2009, 166, 1646–1659. [Google Scholar] [CrossRef] [PubMed]
- Suzuki, N.; Miller, G.; Sejima, H.; Harper, J.; Mittler, R. Enhanced Seed Production under Prolonged Heat Stress Conditions in Arabidopsis Thaliana Plants Deficient in Cytosolic Ascorbate Peroxidase 2. J. Exp. Bot. 2013, 64, 253–263. [Google Scholar] [CrossRef] [PubMed]
- Hu, Z.; Li, J.; Ding, S.; Cheng, F.; Li, X.; Jiang, Y.; Yu, J.; Foyer, C.H.; Shi, K. The Protein Kinase CPK28 Phosphorylates Ascorbate Peroxidase and Enhances Thermotolerance in Tomato. Plant Physiol. 2021, 186, 1302–1317. [Google Scholar] [CrossRef]
- Miller, G.; Suzuki, N.; Rizhsky, L.; Hegie, A.; Koussevitzky, S.; Mittler, R. Double Mutants Deficient in Cytosolic and Thylakoid Ascorbate Peroxidase Reveal a Complex Mode of Interaction between Reactive Oxygen Species, Plant Development, and Response to Abiotic Stresses. Plant Physiol. 2007, 144, 1777–1785. [Google Scholar] [CrossRef]
- Koussevitzky, S.; Suzuki, N.; Huntington, S.; Armijo, L.; Sha, W.; Cortes, D.; Shulaev, V.; Mittler, R. Ascorbate Peroxidase 1 Plays a Key Role in the Response of Arabidopsis Thaliana to Stress Combination S. J. Biol. Chem. 2008, 283, 34197–34203. [Google Scholar] [CrossRef]
- Zandalinas, S.I.; Balfagón, D.; Arbona, V.; Gómez-Cadenas, A.; Inupakutika, M.A.; Mittler, R. ABA Is Required for the Accumulation of APX1 and MBF1c during a Combination of Water Deficit and Heat Stress. J. Exp. Bot. 2016, 67, 5381–5390. [Google Scholar] [CrossRef]
- Kaur, S.; Prakash, P.; Bak, D.-H.; Hong, S.H.; Cho, C.; Chung, M.-S.; Kim, J.-H.; Lee, S.; Bai, H.-W.; Lee, S.Y.; et al. Regulation of Dual Activity of Ascorbate Peroxidase 1 From Arabidopsis Thaliana by Conformational Changes and Posttranslational Modifications. Front. Plant Sci. 2021, 12, 1177. [Google Scholar] [CrossRef] [PubMed]
- Babbar, R.; Karpinska, B.; Grover, A.; Foyer, C.H. Heat-Induced Oxidation of the Nuclei and Cytosol. Front. Plant Sci. 2021, 11, 2184. [Google Scholar] [CrossRef] [PubMed]
- Locato, V.; Gadaleta, C.; De Gara, L.; de Pinto, M.C. Production of Reactive Species and Modulation of Antioxidant Network in Response to Heat Shock: A Critical Balance for Cell Fate. Plant Cell Environ. 2008, 31, 1606–1619. [Google Scholar] [CrossRef] [PubMed]
- Locato, V.; de Pinto, M.C.; De Gara, L. Different Involvement of the Mitochondrial, Plastidial and Cytosolic Ascorbate–Glutathione Redox Enzymes in Heat Shock Responses. Physiol. Plant. 2009, 135, 296–306. [Google Scholar] [CrossRef]
- de Pinto, M.C.; Locato, V.; Sgobba, A.; Romero-Puertas, M.; del, C.; Gadaleta, C.; Delledonne, M.; De Gara, L. S-Nitrosylation of Ascorbate Peroxidase Is Part of Programmed Cell Death Signaling in Tobacco Bright Yellow-2 Cells. Plant Physiol. 2013, 163, 1766–1775. [Google Scholar] [CrossRef] [PubMed]
- Sgobba, A.; Paradiso, A.; Dipierro, S.; de Gara, L.; de Pinto, M.C. Changes in Antioxidants Are Critical in Determining Cell Responses to Short- and Long-Term Heat Stress. Physiol. Plant. 2015, 153, 68–78. [Google Scholar] [CrossRef]
- Camejo, D.; Martí, D.C.; Nicolá, E.; Alarcó, N.B.J.J.; Jimé Nez, A.; Sevilla, F. Response of Superoxide Dismutase Isoenzymes in Tomato Plants (Lycopersicon Esculentum) during Thermo-Acclimation of the Photosynthetic Apparatus. Physiol. Plant. 2007, 131, 367. [Google Scholar] [CrossRef]
- Camejo, D.; Jiménez, A.; Alarcón, J.J.; Torres, W.; Gómez, J.M.; Sevilla, F. Changes in Photosynthetic Parameters and Antioxidant Activities Following Heat-Shock Treatment in Tomato Plants. Funct. Plant Biol. 2006, 33, 177–187. [Google Scholar] [CrossRef]
- Guan, Q.; Lu, X.; Zeng, H.; Zhang, Y.; Zhu, J. Heat Stress Induction of MiR398 Triggers a Regulatory Loop That Is Critical for Thermotolerance in Arabidopsis. Plant J. 2013, 74, 840–851. [Google Scholar] [CrossRef]
- Lasorella, C.; Fortunato, S.; Dipierro, N.; Jeran, N.; Tadini, L.; Vita, F.; Pesaresi, P.; de Pinto, M.C. Chloroplast-Localized GUN1 Contributes to the Acquisition of Basal Thermotolerance in Arabidopsis Thaliana. Front. Plant Sci. 2022, 13, 1058831. [Google Scholar] [CrossRef]
- Kong, F.; Deng, Y.; Wang, G.; Wang, J.; Liang, X.; Meng, Q. LeCDJ1, a Chloroplast DnaJ Protein, Facilitates Heat Tolerance in Transgenic Tomatoes. J. Integr. Plant Biol. 2014, 56, 63–74. [Google Scholar] [CrossRef] [PubMed]
- Wang, G.; Cai, G.; Xu, N.; Zhang, L.; Sun, X.; Guan, J.; Meng, Q. Novel DnaJ Protein Facilitates Thermotolerance of Transgenic Tomatoes. Int. J. Mol. Sci. 2019, 20, 367. [Google Scholar] [CrossRef] [PubMed]
- Wang, X.; Zhang, H.; Xie, Q.; Liu, Y.; Lv, H.; Bai, R.; Ma, R.; Li, X.; Zhang, X.; Guo, Y.D.; et al. SlSNAT Interacts with HSP40, a Molecular Chaperone, to Regulate Melatonin Biosynthesis and Promote Thermotolerance in Tomato. Plant Cell Physiol. 2020, 61, 909–921. [Google Scholar] [CrossRef] [PubMed]
- Jahan, M.S.; Shu, S.; Wang, Y.; Chen, Z.; He, M.; Tao, M.; Sun, J.; Guo, S. Melatonin Alleviates Heat-Induced Damage of Tomato Seedlings by Balancing Redox Homeostasis and Modulating Polyamine and Nitric Oxide Biosynthesis. BMC Plant Biol. 2019, 19, 414. [Google Scholar] [CrossRef] [PubMed]
- Wang, G.; Xing, M.; Hu, T.; Ji, M.; Li, X.; Amombo, E.; Shao, A.; Xu, X.; Fu, J. Photosystem II Photochemical Adjustment of Tall Fescue against Heat Stress after Melatonin Priming. J. Plant Physiol. 2022, 275, 153758. [Google Scholar] [CrossRef]
- Nahar, K.; Hasanuzzaman, M.; Alam, M.M.; Fujita, M. Exogenous Glutathione Confers High Temperature Stress Tolerance in Mung Bean (Vigna Radiata L.) by Modulating Antioxidant Defense and Methylglyoxal Detoxification System. Environ. Exp. Bot. 2014, 112, 44–54. [Google Scholar] [CrossRef]
- Raja, V.; Qadir, S.U.; Alyemeni, M.N.; Ahmad, P. Impact of Drought and Heat Stress Individually and in Combination on Physio-Biochemical Parameters, Antioxidant Responses, and Gene Expression in Solanum Lycopersicum. 3 Biotech 2020, 10, 208. [Google Scholar] [CrossRef]
- Foyer, C.H.; Noctor, G. Ascorbate and Glutathione: The Heart of the Redox Hub. Plant Physiol. 2011, 155, 2–18. [Google Scholar] [CrossRef]
- Hasanuzzaman, M.; Nahar, K.; Fujit, M. Extreme Temperature Responses, Oxidative Stress and Antioxidant Defense in Plants. In Abiotic Stress—Plant Responses and Applications in Agriculture; InTechOpen: London, UK, 2013; pp. 169–205. [Google Scholar] [CrossRef]
- Eyshi Rezaei, E.; Webber, H.; Gaiser, T.; Naab, J.; Ewert, F. Heat Stress in Cereals: Mechanisms and Modelling. Eur. J. Agron. 2015, 64, 98–113. [Google Scholar] [CrossRef]
- Banti, V.; Mafessoni, F.; Loreti, E.; Alpi, A.; Perata, P. The Heat-Inducible Transcription Factor HsfA2 Enhances Anoxia Tolerance in Arabidopsis. Plant Physiol. 2010, 152, 1471. [Google Scholar] [CrossRef]
- Königshofer, H.; Tromballa, H.-W.; Löppert, H.-G. Early Events in Signalling High-Temperature Stress in Tobacco BY2 Cells Involve Alterations in Membrane Fluidity and Enhanced Hydrogen Peroxide Production. Plant Cell Environ. 2008, 31, 1771–1780. [Google Scholar] [CrossRef] [PubMed]
- Wang, L.; Guo, Y.; Jia, L.; Chu, H.; Zhou, S.; Chen, K.; Wu, D.; Zhao, L. Hydrogen Peroxide Acts Upstream of Nitric Oxide in the Heat Shock Pathway in Arabidopsis Seedlings. Plant Physiol. 2014, 164, 2184. [Google Scholar] [CrossRef] [PubMed]
- Wu, F.; Chi, Y.; Jiang, Z.; Xu, Y.; Xie, L.; Huang, F.; Wan, D.; Ni, J.; Yuan, F.; Wu, X.; et al. Hydrogen Peroxide Sensor HPCA1 Is an LRR Receptor Kinase in Arabidopsis. Nature 2020, 578, 577–581. [Google Scholar] [CrossRef]
- Fang, X.; Zhao, G.; Zhang, S.; Li, Y.; Gu, H.; Li, Y.; Zhao, Q.; Qi, Y. Chloroplast-to-Nucleus Signaling Regulates MicroRNA Biogenesis in Arabidopsis. Dev. Cell 2019, 48, 371–382.e4. [Google Scholar] [CrossRef]
- Hu, S.; Ding, Y.; Zhu, C. Sensitivity and Responses of Chloroplasts to Heat Stress in Plants. Front. Plant Sci. 2020, 11, 375. [Google Scholar] [CrossRef] [PubMed]
- Khorobrykh, S.; Havurinne, V.; Mattila, H.; Tyystjärvi, E. Oxygen and ROS in Photosynthesis. Plants 2020, 9, 91. [Google Scholar] [CrossRef]
- Zeng, C.; Jia, T.; Gu, T.; Su, J.; Hu, X. Progress in Research on the Mechanisms Underlying. Genes 2021, 12, 1343. [Google Scholar] [CrossRef]
- Ramel, F.; Birtic, S.; Ginies, C.; Soubigou-Taconnat, L.; Triantaphylidès, C.; Havaux, M. Carotenoid Oxidation Products Are Stress Signals That Mediate Gene Responses to Singlet Oxygen in Plants. Proc. Natl. Acad. Sci. USA 2012, 109, 5535–5540. [Google Scholar] [CrossRef]
- Maruta, T.; Noshi, M.; Tanouchi, A.; Tamoi, M.; Yabuta, Y.; Yoshimura, K.; Ishikawa, T.; Shigeoka, S. H2O2-Triggered Retrograde Signaling from Chloroplasts to Nucleus Plays Specific Role in Response to Stress. J. Biol. Chem. 2012, 287, 11717–11729. [Google Scholar] [CrossRef]
- Koussevitzky, S.; Nott, A.; Mockler, T.C.; Hong, F.; Sachetto-Martins, G.; Surpin, M.; Lim, J.; Mittler, R.; Chory, J. Signals from Chloroplasts Converge to Regulate Nuclear Gene Expression. Science 2007, 316, 715–719. [Google Scholar] [CrossRef]
- Wu, G.Z.; Chalvin, C.; Hoelscher, M.; Meyer, E.H.; Wu, X.N.; Bock, R. Control of Retrograde Signaling by Rapid Turnover of GENOMES UNCOUPLED1. Plant Physiol. 2018, 176, 2472–2495. [Google Scholar] [CrossRef] [PubMed]
- Pesaresi, P.; Kim, C. Current Understanding of GUN1: A Key Mediator Involved in Biogenic Retrograde Signaling. Plant Cell Rep. 2019, 38, 819–823. [Google Scholar] [CrossRef] [PubMed]
- Borisova, M.M.; Kozuleva, M.A.; Rudenko, N.N.; Naydov, I.A.; Klenina, I.B.; Ivanov, B.N. Photosynthetic Electron Flow to Oxygen and Diffusion of Hydrogen Peroxide through the Chloroplast Envelope via Aquaporins. Proc. Biochim. et Biophys. Acta Bioenerg. 2012, 1817, 1314–1321. [Google Scholar] [CrossRef] [PubMed]
- Mullineaux, P.M.; Exposito-Rodriguez, M.; Laissue, P.P.; Smirnoff, N.; Park, E. Spatial Chloroplast-to-Nucleus Signalling Involving Plastid-Nuclear Complexes and Stromules. Philos. Trans. R Soc. Lond. B Biol. Sci. 2020, 375, 20190405. [Google Scholar] [CrossRef]
- Breeze, E.; Mullineaux, P.M. The Passage of H2O2 from Chloroplasts to Their Associated Nucleus during Retrograde Signalling: Reflections on the Role of the Nuclear Envelope. Plants 2022, 11, 552. [Google Scholar] [CrossRef] [PubMed]
- Devireddy, A.R.; Zandalinas, S.I.; Fichman, Y.; Mittler, R. Integration of Reactive Oxygen Species and Hormone Signaling during Abiotic Stress. Plant J. 2021, 105, 459–476. [Google Scholar] [CrossRef] [PubMed]
- Devireddy, A.R.; Tschaplinski, T.J.; Tuskan, G.A.; Muchero, W.; Chen, J.G. Role of Reactive Oxygen Species and Hormones in Plant Responses to Temperature Changes. Int. J. Mol. Sci. 2021, 22, 8843. [Google Scholar] [CrossRef]
- Xia, X.J.; Zhou, Y.H.; Shi, K.; Zhou, J.; Foyer, C.H.; Yu, J.Q. Interplay between Reactive Oxygen Species and Hormones in the Control of Plant Development and Stress Tolerance. J. Exp. Bot. 2015, 66, 2839–2856. [Google Scholar] [CrossRef]
- Kollist, H.; Zandalinas, S.I.; Sengupta, S.; Nuhkat, M.; Kangasjärvi, J.; Mittler, R. Rapid Responses to Abiotic Stress: Priming the Landscape for the Signal Transduction Network. Trends Plant Sci. 2019, 24, 25–37. [Google Scholar] [CrossRef]
- Zandalinas, S.I.; Sengupta, S.; Burks, D.; Azad, R.K.; Mittler, R. Identification and Characterization of a Core Set of ROS Wave-Associated Transcripts Involved in the Systemic Acquired Acclimation Response of Arabidopsis to Excess Light. Plant J. 2019, 98, 126–141. [Google Scholar] [CrossRef]
- Zandalinas, S.I.; Fichman, Y.; Devireddy, A.R.; Sengupta, S.; Azad, R.K.; Mittler, R. Systemic Signaling during Abiotic Stress Combination in Plants. Proc. Natl. Acad. Sci. USA 2020, 117, 13810–13820. [Google Scholar] [CrossRef] [PubMed]
- Zhou, J.; Wang, J.; Li, X.; Xia, X.J.; Zhou, Y.H.; Shi, K.; Chen, Z.; Yu, J.Q. H2O2 Mediates the Crosstalk of Brassinosteroid and Abscisic Acid in Tomato Responses to Heat and Oxidative Stresses. J. Exp. Bot. 2014, 65, 4371–4383. [Google Scholar] [CrossRef] [PubMed]
- Han, Y.; Chaouch, S.; Mhamdi, A.; Queval, G.; Zechmann, B.; Noctor, G. Functional Analysis of Arabidopsis Mutants Points to Novel Roles for Glutathione in Coupling H2O2 to Activation of Salicylic Acid Accumulation and Signaling. Antioxid. Redox Signal. 2013, 18, 2106–2121. [Google Scholar] [CrossRef]
- Prodhan, M.Y.; Munemasa, S.; Nahar, M.N.E.N.; Nakamura, Y.; Murata, Y. Guard Cell Salicylic Acid Signaling Is Integrated into Abscisic Acid Signaling via the Ca2+/CPK-Dependent Pathway. Plant Physiol. 2018, 178, 441–450. [Google Scholar] [CrossRef]
- Lee, S.; Lee, H.J.; Jung, J.H.; Park, C.M. The Arabidopsis Thaliana RNA-Binding Protein FCA Regulates Thermotolerance by Modulating the Detoxification of Reactive Oxygen Species. New Phytol. 2015, 205, 555–569. [Google Scholar] [CrossRef] [PubMed]
- Kuromori, T.; Seo, M.; Shinozaki, K. ABA Transport and Plant Water Stress Responses. Trends Plant Sci. 2018, 23, 513–522. [Google Scholar] [CrossRef] [PubMed]
- Yoshida, T.; Obata, T.; Feil, R.; Lunn, J.E.; Fujita, Y.; Yamaguchi-Shinozaki, K.; Fernie, A.R. The Role of Abscisic Acid Signaling in Maintaining the Metabolic Balance Required for Arabidopsis Growth under Nonstress Conditions. Plant Cell. 2019, 31, 84–105. [Google Scholar] [CrossRef]
- Mittler, R.; Blumwald, E. The Roles of ROS and ABA in Systemic Acquired Acclimation. Plant Cell 2015, 27, 64–70. [Google Scholar] [CrossRef]
- Yao, Y.; He, R.J.; Xie, Q.L.; Zhao, X.h.; Deng, X.m.; He, J.b.; Song, L.; He, J.; Marchant, A.; Chen, X.Y.; et al. ETHYLENE RESPONSE FACTOR 74 (ERF74) Plays an Essential Role in Controlling a Respiratory Burst Oxidase Homolog D (RbohD)-Dependent Mechanism in Response to Different Stresses in Arabidopsis. New Phytol. 2017, 213, 1667–1681. [Google Scholar] [CrossRef]
- Suzuki, N.; Bajad, S.; Shuman, J.; Shulaev, V.; Mittler, R. The Transcriptional Co-Activator MBF1c Is a Key Regulator of Thermotolerance in Arabidopsis Thaliana. J. Biol. Chem. 2008, 283, 9269–9275. [Google Scholar] [CrossRef]
- Zandalinas, S.I.; Fritschi, F.B.; Mittler, R. Signal Transduction Networks during Stress Combination. J. Exp. Bot. 2020, 71, 1734–1741. [Google Scholar] [CrossRef] [PubMed]
- Chen, C.; Letnik, I.; Hacham, Y.; Dobrev, P.; Ben-Daniel, B.H.; Vanková, R.; Amir, R.; Miller, G. ASCORBATE PEROXIDASE6 Protects Arabidopsis Desiccating and Germinating Seeds from Stress and Mediates Cross Talk between Reactive Oxygen Species, Abscisic Acid, And Auxin. Plant Physiol. 2014, 166, 370–383. [Google Scholar] [CrossRef] [PubMed]
- Yin, Y.; Qin, K.; Song, X.; Zhang, Q.; Zhou, Y.; Xia, X.; Yu, J. BZR1 Transcription Factor Regulates Heat Stress Tolerance through FERONIA Receptor-like Kinase-Mediated Reactive Oxygen Species Signaling in Tomato. Plant Cell Physiol. 2018, 59, 2239–2254. [Google Scholar] [CrossRef] [PubMed]
- Wang, L.J.; Fan, L.; Loescher, W.; Duan, W.; Liu, G.J.; Cheng, J.S.; Luo, H.B.; Li, S.H. Salicylic Acid Alleviates Decreases in Photosynthesis under Heat Stress and Accelerates Recovery in Grapevine Leaves. BMC Plant Biol. 2010, 10, 34. [Google Scholar] [CrossRef] [PubMed]
- Duan, J.; Lee, K.P.; Dogra, V.; Zhang, S.; Liu, K.; Caceres-Moreno, C.; Lv, S.; Xing, W.; Kato, Y.; Sakamoto, W.; et al. Impaired Psii Proteostasis Promotes Retrograde Signaling via Salicylic Acid1. Plant Physiol. 2019, 180, 2182–2197. [Google Scholar] [CrossRef]
- Suzuki, N.; Katano, K. Coordination between ROS Regulatory Systems and Other Pathways under Heat Stress and Pathogen Attack. Front. Plant Sci. 2018, 9, 490. [Google Scholar] [CrossRef]
- Shah Jahan, M.; Wang, Y.; Shu, S.; Zhong, M.; Chen, Z.; Wu, J.; Sun, J.; Guo, S. Exogenous Salicylic Acid Increases the Heat Tolerance in Tomato (Solanum Lycopersicum L) by Enhancing Photosynthesis Efficiency and Improving Antioxidant Defense System through Scavenging of Reactive Oxygen Species. Sci. Hortic. 2019, 247, 421–429. [Google Scholar] [CrossRef]
- Torun, H.; Novák, O.; Mikulík, J.; Pěnčík, A.; Strnad, M.; Ayaz, F.A. Timing-Dependent Effects of Salicylic Acid Treatment on Phytohormonal Changes, ROS Regulation, and Antioxidant Defense in Salinized Barley (Hordeum Vulgare L.). Sci. Rep. 2020, 10, 13886. [Google Scholar] [CrossRef]
- Zhao, Q.; Zhou, L.; Liu, J.; Cao, Z.; Du, X.; Huang, F.; Pan, G.; Cheng, F. Involvement of CAT in the Detoxification of HT-Induced ROS Burst in Rice Anther and Its Relation to Pollen Fertility. Plant Cell Rep. 2018, 37, 741–757. [Google Scholar] [CrossRef]
- Feng, B.; Zhang, C.; Chen, T.; Zhang, X.; Tao, L.; Fu, G. Salicylic Acid Reverses Pollen Abortion of Rice Caused by Heat Stress. BMC Plant Biol. 2018, 18, 245. [Google Scholar] [CrossRef]
- Valluru, R.; Reynolds, M.P.; Davies, W.J.; Sukumaran, S. Phenotypic and Genome-Wide Association Analysis of Spike Ethylene in Diverse Wheat Genotypes under Heat Stress. New Phytol. 2017, 214, 271–283. [Google Scholar] [CrossRef] [PubMed]
- Wi, S.J.; Jang, S.J.; Park, K.Y. Inhibition of Biphasic Ethylene Production Enhances Tolerance to Abiotic Stress by Reducing the Accumulation of Reactive Oxygen Species in Nicotiana Tabacum. Mol. Cells 2010, 30, 37–49. [Google Scholar] [CrossRef]
- Takács, Z.; Poór, P.; Borbély, P.; Czékus, Z.; Szalai, G.; Tari, I. H2O2 Homeostasis in Wild-Type and Ethylene-Insensitive Never Ripe Tomato in Response to Salicylic Acid Treatment in Normal Photoperiod and in Prolonged Darkness. Plant Physiol. Biochem. 2018, 126, 74–85. [Google Scholar] [CrossRef] [PubMed]
- Wu, Y.S.; Yang, C.Y. Ethylene-Mediated Signaling Confers Thermotolerance and Regulates Transcript Levels of Heat Shock Factors in Rice Seedlings under Heat Stress. Bot. Stud. 2019, 60, 23. [Google Scholar] [CrossRef] [PubMed]
- Müller, M.; Munné-Bosch, S. Ethylene Response Factors: A Key Regulatory Hub in Hormone and Stress Signaling. Plant Physiol. 2015, 169, 32–41. [Google Scholar] [CrossRef]
- Wu, L.; Zhang, Z.; Zhang, H.; Wang, X.C.; Huang, R. Transcriptional Modulation of Ethylene Response Factor Protein JERF3 in the Oxidative Stress Response Enhances Tolerance of Tobacco Seedlings to Salt, Drought, and Freezing. Plant Physiol. 2008, 148, 1953–1963. [Google Scholar] [CrossRef]
- Xie, Z.; Nolan, T.M.; Jiang, H.; Yin, Y. AP2/ERF Transcription Factor Regulatory Networks in Hormone and Abiotic Stress Responses in Arabidopsis. Front. Plant Sci. 2019, 10, 288. [Google Scholar] [CrossRef]
- Skalák, J.; Cerný, M.; Jedelský, P.; Dobrá, J.; Ge, E.; Novák, J.; Hronková, M.; Dobrev, P.; Vanková, R.; Brzobohatý, B. Stimulation of Ipt Overexpression as a Tool to Elucidate the Role of Cytokinins in High Temperature Responses of Arabidopsis Thaliana. J. Exp. Bot. 2016, 67, 2861–2873. [Google Scholar] [CrossRef]
- Cortleven, A.; Leuendorf, J.E.; Frank, M.; Pezzetta, D.; Bolt, S.; Schmülling, T. Cytokinin Action in Response to Abiotic and Biotic Stresses in Plants. Plant Cell Environ. 2019, 42, 998–1018. [Google Scholar] [CrossRef]
- Macková, H.; Hronková, M.; Dobrá, J.; Turečková, V.; Novák, O.; Lubovská, Z.; Motyka, V.; Haisel, D.; Hájek, T.; Prášil, I.T.; et al. Enhanced Drought and Heat Stress Tolerance of Tobacco Plants with Ectopically Enhanced Cytokinin Oxidase/Dehydrogenase Gene Expression. J. Exp. Bot. 2013, 64, 2805–2815. [Google Scholar] [CrossRef]
- Prerostova, S.; Dobrev, P.I.; Kramna, B.; Gaudinova, A.; Knirsch, V.; Spichal, L.; Zatloukal, M.; Vankova, R. Heat Acclimation and Inhibition of Cytokinin Degradation Positively Affect Heat Stress Tolerance of Arabidopsis. Front. Plant Sci. 2020, 11, 87. [Google Scholar] [CrossRef]
- Foyer, C.H.; Noctor, G. Redox Regulation in Photosynthetic Organisms. Regulation 2009, 11, 861–905. [Google Scholar] [CrossRef]
- Bäurle, I. Plant Heat Adaptation: Priming in Response to Heat Stress. F1000Research 2016, 18, 694. [Google Scholar] [CrossRef] [PubMed]
- Nguyen, X.C.; Kim, S.H.; Lee, K.; Kim, K.E.; Liu, X.M.; Han, H.J.; Hoang, M.H.T.; Lee, S.W.; Hong, J.C.; Moon, Y.H.; et al. Identification of a C2H2-Type Zinc Finger Transcription Factor (ZAT10) from Arabidopsis as a Substrate of MAP Kinase. Plant Cell Rep. 2012, 31, 737–745. [Google Scholar] [CrossRef] [PubMed]
- Foyer, C.H.; Baker, A.; Wright, M.; Sparkes, I.A.; Mhamdi, A.; Schippers, J.H.M.; Van Breusegem, F. On the Move: Redox-Dependent Protein Relocation in Plants. J. Exp. Bot. 2020, 71, 620–631. [Google Scholar] [CrossRef] [PubMed]
- Couturier, J.; Chibani, K.; Jacquot, J.P.; Rouhier, N. Cysteine-Based Redox Regulation and Signaling in Plants. Front. Plant Sci. 2013, 4, 105. [Google Scholar] [CrossRef] [PubMed]
- Spadaro, D.; Yun, B.W.; Spoel, S.H.; Chu, C.; Wang, Y.Q.; Loake, G.J. The Redox Switch: Dynamic Regulation of Protein Function by Cysteine Modifications. Physiol. Plant. 2010, 138, 360–371. [Google Scholar] [CrossRef]
- Davies, M.J. The Oxidative Environment and Protein Damage. Biochim. Biophys. Acta Proteins Proteom. 2005, 1703, 93–109. [Google Scholar] [CrossRef]
- Uchida, A.; Jagendorf, A.T.; Hibino, T.; Takabe, T.; Takabe, T. Effects of Hydrogen Peroxide and Nitric Oxide on Both Salt and Heat Stress Tolerance in Rice. Plant Sci. 2002, 163, 515–523. [Google Scholar] [CrossRef]
- Zhang, J.; Liao, W. Protein S-Nitrosylation in Plant Abiotic Stresses. Funct. Plant Biol. 2019, 47, 1–10. [Google Scholar] [CrossRef]
- Song, L.; Ding, W.; Zhao, M.; Sun, B.; Zhang, L. Nitric Oxide Protects against Oxidative Stress under Heat Stress in the Calluses from Two Ecotypes of Reed. Plant Sci. 2006, 171, 449–458. [Google Scholar] [CrossRef]
- Zhao, M.G.; Chen, L.; Zhang, L.L.; Zhang, W.H. Nitric Reductase-Dependent Nitric Oxide Production Is Involved in Cold Acclimation and Freezing Tolerance in Arabidopsis. Plant Physiol. 2009, 151, 755–767. [Google Scholar] [CrossRef]
- Parankusam, S.; Adimulam, S.S.; Bhatnagar-Mathur, P.; Sharma, K.K. Nitric Oxide (NO) in Plant Heat Stress Tolerance: Current Knowledge and Perspectives. Front. Plant Sci. 2017, 8, 1582. [Google Scholar] [CrossRef] [PubMed]
- Cheng, T.; Chen, J.; EF, A.A.; Wang, P.; Wang, G.; Hu, X.; Shi, J. Quantitative Proteomics Analysis Reveals That S-Nitrosoglutathione Reductase (GSNOR) and Nitric Oxide Signaling Enhance Poplar Defense against Chilling Stress. Planta 2015, 242, 1361–1390. [Google Scholar] [CrossRef] [PubMed]
- Li, B.; Sun, L.; Huang, J.; Göschl, C.; Shi, W.; Chory, J.; Busch, W. GSNOR Provides Plant Tolerance to Iron Toxicity via Preventing Iron-Dependent Nitrosative and Oxidative Cytotoxicity. Nat. Commun. 2019, 10, 3896. [Google Scholar] [CrossRef] [PubMed]
- Lee, U.; Wie, C.; Fernandez, B.O.; Feelisch, M.; Vierling, E. Modulation of Nitrosative Stress by S-Nitrosoglutathione Reductase Is Critical for Thermotolerance and Plant Growth in Arabidopsis. Plant Cell 2008, 20, 786–802. [Google Scholar] [CrossRef] [PubMed]
- Xu, S.; Guerra, D.; Lee, U.; Vierling, E. S-Nitrosoglutathione Reductases Are Low-Copy Number, Cysteine-Rich Proteins in Plants That Control Multiple Developmental and Defense Responses in Arabidopsis. Front. Plant Sci. 2013, 4, 430. [Google Scholar] [CrossRef]
- Jacques, S.; Ghesquière, B.; Van Breusegem, F.; Gevaert, K. Plant Proteins under Oxidative Attack. Proteomics 2013, 13, 932–940. [Google Scholar] [CrossRef]
- Ancín, M.; Millan, A.F.S.; Larraya, L.; Morales, F.; Veramendi, J.; Aranjuelo, I.; Farran, I. Overexpression of Thioredoxin m in Tobacco Chloroplasts Inhibits the Protein Kinase STN7 and Alters Photosynthetic Performance. J. Exp. Bot. 2019, 70, 1005–1016. [Google Scholar] [CrossRef]
- Reiland, S.; Messerli, G.; Baerenfaller, K.; Gerrits, B.; Endler, A.; Grossmann, J.; Gruissem, W.; Baginsky, S. Large-Scale Arabidopsis Phosphoproteome Profiling Reveals Novel Chloroplast Kinase Substrates and Phosphorylation Networks1[W]. Plant Physiol. 2009, 150, 889–903. [Google Scholar] [CrossRef]
- Park, S.K.; Jung, Y.J.; Lee, J.R.; Lee, Y.M.; Jang, H.H.; Lee, S.S.; Park, J.H.; Kim, S.Y.; Moon, J.C.; Lee, S.Y.; et al. Heat-Shock and Redox-Dependent Functional Switching of an h-Type Arabidopsis Thioredoxin from a Disulfide Reductase to a Molecular Chaperone. Plant Physiol. 2009, 150, 552–561. [Google Scholar] [CrossRef] [PubMed]
- Chae, H.B.; Moon, J.C.; Shin, M.R.; Chi, Y.H.; Jung, Y.J.; Lee, S.Y.; Nawkar, G.M.; Jung, H.S.; Hyun, J.K.; Kim, W.Y.; et al. Thioredoxin Reductase Type C (NTRC) Orchestrates Enhanced Thermotolerance to Arabidopsis by Its Redox-Dependent Holdase Chaperone Function. Mol. Plant 2013, 6, 323–336. [Google Scholar] [CrossRef] [PubMed]
- Conway, M.E.; Lee, C. The Redox Switch That Regulates Molecular Chaperones. Biomol. Concepts 2015, 6, 269–284. [Google Scholar] [CrossRef] [PubMed]
- Martins, L.; Knuesting, J.; Bariat, L.; Dard, A.; Freibert, S.A.; Marchand, C.H.; Young, D.; Dung, N.H.T.; Voth, W.; Debures, A.; et al. Redox Modification of the Iron-Sulfur Glutaredoxin GRXS17 Activates Holdase Activity and Protects Plants from Heat Stress. Plant Physiol. 2020, 184, 676–692. [Google Scholar] [CrossRef]
- Paeng, S.K.; Chi, Y.H.; Kang, C.H.; Chae, H.B.; Lee, E.S.; Park, J.H.; Wi, S.D.; Bae, S.B.; Phan, K.A.T.; Lee, S.Y. Chaperone Function of Arabidopsis NPR1. Plant Biotechnol. Rep. 2020, 14, 227–233. [Google Scholar] [CrossRef]
- Moon, J.C.; Jang, H.H.; Chae, H.B.; Lee, J.R.; Lee, S.Y.; Jung, Y.J.; Shin, M.R.; Lim, H.S.; Chung, W.S.; Yun, D.J.; et al. The C-Type Arabidopsis Thioredoxin Reductase ANTR-C Acts as an Electron Donor to 2-Cys Peroxiredoxins in Chloroplasts. Biochem. Biophys. Res. Commun. 2006, 348, 478–484. [Google Scholar] [CrossRef]
- Pérez-Ruiz, J.M.; Spínola, M.C.; Kirchsteiger, K.; Moreno, J.; Sahrawy, M.; Cejudo, F.J. Rice NTRC Is a High-Efficiency Redox System for Chloroplast Protection against Oxidative Damage. Plant Cell 2006, 18, 2356. [Google Scholar] [CrossRef]
- Alkhalfioui, F.; Renard, M.; Vensel, W.H.; Wong, J.; Tanaka, C.K.; Hurkman, W.J.; Buchanan, B.B.; Montrichard, F. Thioredoxin-Linked Proteins Are Reduced during Germination of Medicago Truncatula Seeds. Plant Physiol. 2007, 144, 1559–1579. [Google Scholar] [CrossRef]
- Pérez-Ruiz, J.M.; Cejudo, F.J. A Proposed Reaction Mechanism for Rice NADPH Thioredoxin Reductase C, an Enzyme with Protein Disulfide Reductase Activity. FEBS Lett. 2009, 583, 1399–1402. [Google Scholar] [CrossRef]
- Wulff, R.P.; Lundqvist, J.; Rutsdottir, G.; Hansson, A.; Stenbaek, A.; Elmlund, D.; Elmlund, H.; Jensen, P.E.; Hansson, M. The Activity of Barley NADPH-Dependent Thioredoxin Reductase C Is Independent of the Oligomeric State of the Protein: Tetrameric Structure Determined by Cryo-Electron Microscopy. Biochemistry 2011, 50, 3713–3723. [Google Scholar] [CrossRef]
- Chae, H.Z.; Oubrahim, H.; Park, J.W.; Rhee, S.G.; Chock, P.B. Protein Glutathionylation in the Regulation of Peroxiredoxins: A Family of Thiol-Specific Peroxidases That Function As Antioxidants, Molecular Chaperones, and Signal Modulators. Antioxid. Redox Signal. 2012, 16, 506. [Google Scholar] [CrossRef] [PubMed]
- Rhee, S.G.; Woo, H.A.; Kil, I.S.; Bae, S.H. Peroxiredoxin Functions as a Peroxidase and a Regulator and Sensor of Local Peroxides. J. Biol. Chem. 2012, 287, 4403–4410. [Google Scholar] [CrossRef] [PubMed]
- Kim, K.H.; Alam, I.; Lee, K.W.; Sharmin, S.A.; Kwak, S.S.; Lee, S.Y.; Lee, B.H. Enhanced Tolerance of Transgenic Tall Fescue Plants Overexpressing 2-Cys Peroxiredoxin against Methyl Viologen and Heat Stresses. Biotechnol. Lett. 2010, 32, 571–576. [Google Scholar] [CrossRef] [PubMed]
- Poole, L.B.; Nelson, K.J. Discovering Mechanisms of Signaling-Mediated Cysteine Oxidation. Curr. Opin. Chem. Biol. 2008, 12, 18–24. [Google Scholar] [CrossRef]
- Phalen, T.J.; Weirather, K.; Deming, P.B.; Anathy, V.; Howe, A.K.; Van Der Vliet, A.; Jönsson, T.J.; Poole, L.B.; Heintz, N.H. Oxidation State Governs Structural Transitions in Peroxiredoxin II That Correlate with Cell Cycle Arrest and Recovery. J. Cell Biol. 2006, 175, 779–789. [Google Scholar] [CrossRef]
- Sprague, S.A.; Tamang, T.M.; Steiner, T.; Wu, Q.; Hu, Y.; Kakeshpour, T.; Park, J.; Yang, J.; Peng, Z.; Bergkamp, B.; et al. Redox-Engineering Enhances Maize Thermotolerance and Grain Yield in the Field. Plant Biotechnol. J. 2022, 20, 1819–1832. [Google Scholar] [CrossRef]
- Birkenbihl, R.P.; Liu, S.; Somssich, I.E. Transcriptional Events Defining Plant Immune Responses. Curr. Opin. Plant Biol. 2017, 38, 1–9. [Google Scholar] [CrossRef]
- Liu, J.; Feng, L.; Li, J.; He, Z. Genetic and Epigenetic Control of Plant Heat Responses. Front. Plant Sci. 2015, 6, 267. [Google Scholar] [CrossRef]
- Zhu, J.-K. Active DNA Methylation Mediated by DNA Glycosylases The Interplay between Intragenic Heterochromatin and RNA Processing View Project Stomatal Development View Project. Artic. Annu. Rev. Genet. 2009, 43, 143–166. [Google Scholar] [CrossRef]
- Niu, Y.; Desmarais, T.L.; Tong, Z.; Yao, Y.; Costa, M. Oxidative Stress Alters Global Histone Modification and DNA Methylation. Free Radic. Biol. Med. 2015, 82, 22–28. [Google Scholar] [CrossRef]
- He, H.; Van Breusegem, F.; Mhamdi, A. Redox-Dependent Control of Nuclear Transcription in Plants. J. Exp. Bot. 2018, 69, 3359–3372. [Google Scholar] [CrossRef] [PubMed]
- Kumar, J.; Gupta, D.S.; Kesari, R.; Verma, R.; Murugesan, S.; Basu, P.S.; Soren, K.R.; Gupta, S.; Singh, N.P. Comprehensive RNAseq Analysis for Identification of Genes Expressed under Heat Stress in Lentil. Physiol. Plant. 2021, 173, 1785–1807. [Google Scholar] [CrossRef] [PubMed]
- Mittler, R.; Vanderauwera, S.; Suzuki, N.; Miller, G.; Tognetti, V.B.; Vandepoele, K.; Gollery, M.; Shulaev, V.; Van Breusegem, F. ROS Signaling: The New Wave? Trends Plant Sci. 2011, 16, 300–309. [Google Scholar] [CrossRef] [PubMed]
- Miller, G.; Mittler, R. Could Heat Shock Transcription Factors Function as Hydrogen Peroxide Sensors in Plants? Ann. Bot. 2006, 98, 279–288. [Google Scholar] [CrossRef]
- Mishra, S.K.; Tripp, J.; Winkelhaus, S.; Tschiersch, B.; Theres, K.; Nover, L.; Scharf, K.D. In the Complex Family of Heat Stress Transcription Factors, HsfA1 Has a Unique Role as Master Regulator of Thermotolerance in Tomato. Genes Dev. 2002, 16, 1555–1567. [Google Scholar] [CrossRef]
- Yoshida, T.; Ohama, N.; Nakajima, J.; Kidokoro, S.; Mizoi, J.; Nakashima, K.; Maruyama, K.; Kim, J.M.; Seki, M.; Todaka, D.; et al. Arabidopsis HsfA1 Transcription Factors Function as the Main Positive Regulators in Heat Shock-Responsive Gene Expression. Mol. Genet. Genomics 2011, 286, 321–332. [Google Scholar] [CrossRef]
- Hübel, A.; Schöffl, F. Arabidopsis Heat Shock Factor: Isolation and Characterization of the Gene and the Recombinant Protein. Plant Mol. Biol. 1994, 26, 353–362. [Google Scholar] [CrossRef]
- Liu, Y.; Zhang, C.; Chen, J.; Guo, L.; Li, X.; Li, W.; Yu, Z.; Deng, J.; Zhang, P.; Zhang, K.; et al. Arabidopsis Heat Shock Factor HsfA1a Directly Senses Heat Stress, PH Changes, and Hydrogen Peroxide via the Engagement of Redox State. Plant Physiol. Biochem. 2013, 64, 92–98. [Google Scholar] [CrossRef]
- Xuan, Y.; Zhou, S.; Wang, L.; Cheng, Y.; Zhao, L. Nitric Oxide Functions as a Signal and Acts Upstream of AtCaM3 in Thermotolerance in Arabidopsis Seedlings. Plant Physiol. 2010, 153, 1895–1906. [Google Scholar] [CrossRef]
- Andrási, N.; Pettkó-Szandtner, A.; Szabados, L. Diversity of Plant Heat Shock Factors: Regulation, Interactions, and Functions. J. Exp. Bot. 2021, 72, 1558–1575. [Google Scholar] [CrossRef]
- Pérez-Salamó, I.; Papdi, C.; Rigó, G.; Zsigmond, L.; Vilela, B.; Lumbreras, V.; Nagy, I.; Horváth, B.; Domoki, M.; Darula, Z.; et al. The Heat Shock Factor A4A Confers Salt Tolerance and Is Regulated by Oxidative Stress and the Mitogen-Activated Protein Kinases MPK3 and MPK6. Plant Physiol. 2014, 165, 319–334. [Google Scholar] [CrossRef]
- Andrási, N.; Rigó, G.; Zsigmond, L.; Pérez-Salamó, I.; Papdi, C.; Klement, E.; Pettkó-Szandtner, A.; Baba, A.I.; Ayaydin, F.; Dasari, R.; et al. The Mitogen-Activated Protein Kinase 4-Phosphorylated Heat Shock Factor A4A Regulates Responses to Combined Salt and Heat Stresses. J. Exp. Bot. 2019, 70, 4903–4918. [Google Scholar] [CrossRef] [PubMed]
- He, N.Y.; Chen, L.S.; Sun, A.Z.; Zhao, Y.; Yin, S.N.; Guo, F.Q. A Nitric Oxide Burst at the Shoot Apex Triggers a Heat-Responsive Pathway in Arabidopsis. Nat. Plants 2022, 8, 434–450. [Google Scholar] [CrossRef] [PubMed]
- Giesguth, M.; Sahm, A.; Simon, S.; Dietz, K.J. Redox-Dependent Translocation of the Heat Shock Transcription Factor AtHSFA8 from the Cytosol to the Nucleus in Arabidopsis Thaliana. FEBS Lett. 2015, 589, 718–725. [Google Scholar] [CrossRef]
- Schneider, M.; Knuesting, J.; Birkholz, O.; Heinisch, J.J.; Scheibe, R. Cytosolic GAPDH as a Redox-Dependent Regulator of Energy Metabolism. BMC Plant Biol. 2018, 18, 184. [Google Scholar] [CrossRef]
- Vescovi, M.; Zaffagnini, M.; Festa, M.; Trost, P.; Lo Schiavo, F.; Costa, A. Nuclear Accumulation of Cytosolic Glyceraldehyde-3-Phosphate Dehydrogenase in Cadmium-Stressed Arabidopsis Roots. Plant Physiol. 2013, 162, 333. [Google Scholar] [CrossRef] [PubMed]
- Henry, E.; Fung, N.; Liu, J.; Drakakaki, G.; Coaker, G. Beyond Glycolysis: GAPDHs Are Multi-Functional Enzymes Involved in Regulation of ROS, Autophagy, and Plant Immune Responses. PLoS Genet. 2015, 11, e1005199. [Google Scholar] [CrossRef] [PubMed]
- Han, S.; Wang, Y.; Zheng, X.; Jia, Q.; Zhao, J.; Bai, F.; Hong, Y.; Liu, Y. Cytoplastic Glyceraldehyde-3-Phosphate Dehydrogenases Interact with ATG3 to Negatively Regulate Autophagy and Immunity in Nicotiana Benthamiana. Plant Cell 2015, 27, 1316–1331. [Google Scholar] [CrossRef] [PubMed]
- Kim, S.C.; Guo, L.; Wang, X. Nuclear Moonlighting of Cytosolic Glyceraldehyde-3-Phosphate Dehydrogenase Regulates Arabidopsis Response to Heat Stress. Nat. Commun. 2020, 11, 3439. [Google Scholar] [CrossRef]
- Krause, K.; Kilbienski, I.; Mulisch, M.; Rödiger, A.; Schäfer, A.; Krupinska, K. DNA-Binding Proteins of the Whirly Family in Arabidopsis Thaliana Are Targeted to the Organelles. FEBS Lett. 2005, 579, 3707–3712. [Google Scholar] [CrossRef]
- Grabowski, E.; Miao, Y.; Mulisch, M.; Krupinska, K. Single-Stranded DNA-Binding Protein Whirly1 in Barley Leaves Is Located in Plastids and the Nucleus of the Same Cell. Plant Physiol. 2008, 147, 1800. [Google Scholar] [CrossRef] [PubMed]
- Cappadocia, L.; Parent, J.S.; Zampini, É.; Lepage, É.; Sygusch, J.; Brisson, N. A Conserved Lysine Residue of Plant Whirly Proteins Is Necessary for Higher Order Protein Assembly and Protection against DNA Damage. Nucleic Acids Res. 2012, 40, 258–269. [Google Scholar] [CrossRef] [PubMed]
- Krupinska, K.; Oetke, S.; Desel, C.; Mulisch, M.; Schäfer, A.; Hollmann, J.; Kumlehn, J.; Hensel, G. WHIRLY1 Is a Major Organizer of Chloroplast Nucleoids. Front. Plant Sci. 2014, 5, 432. [Google Scholar] [CrossRef] [PubMed]
- Huang, D.; Lin, W.; Deng, B.; Ren, Y.; Miao, Y. Dual-Located WHIRLY1 Interacting with LHCA1 Alters Photochemical Activities of Photosystem i and Is Involved in Light Adaptation in Arabidopsis. Int. J. Mol. Sci. 2017, 18, 2352. [Google Scholar] [CrossRef]
- Desveaux, D.; Subramaniam, R.; Després, C.; Mess, J.N.; Lévesque, C.; Fobert, P.R.; Dangl, J.L.; Brisson, N. A “Whirly” Transcription Factor Is Required for Salicylic Acid-Dependent Disease Resistance in Arabidopsis. Dev. Cell 2004, 6, 229–240. [Google Scholar] [CrossRef]
- Desveaux, D.; Despres, C.; Joyeux, A.; Subramaniam, R.; Brisson, N. PBF-2 Is a Novel Single-Stranded DNA Binding Factor Implicated in PR-10a Gene Activation in Potato. Plant Cell 2000, 12, 1477. [Google Scholar] [CrossRef]
- Zhuang, K.; Gao, Y.; Liu, Z.; Diao, P.; Sui, N.; Meng, Q.; Meng, C.; Kong, F. WHIRLY1 Regulates HSP21.5A Expression to Promote Thermotolerance in Tomato. Plant Cell Physiol. 2020, 61, 169–177. [Google Scholar] [CrossRef]
- Foyer, C.H.; Karpinska, B.; Krupinska, K. The Functions of WHIRLY1 and REDOXRESPONSIVE TRANSCRIPTION FACTOR 1 in Cross Tolerance Responses in Plants: A Hypothesis. Philos. Trans. R. Soc. B Biol. Sci. 2014, 369, 15–17. [Google Scholar] [CrossRef]
- Georgii, E.; Jin, M.; Zhao, J.; Kanawati, B.; Schmitt-Kopplin, P.; Albert, A.; Winkler, J.B.; Schäffner, A.R. Relationships between Drought, Heat and Air Humidity Responses Revealed by Transcriptome-Metabolome Co-Analysis. BMC Plant Biol. 2017, 17, 120. [Google Scholar] [CrossRef]
- Pandey, P.; Ramegowda, V.; Senthil-Kumar, M. Shared and Unique Responses of Plants to Multiple Individual Stresses and Stress Combinations: Physiological and Molecular Mechanisms. Front. Plant Sci. 2015, 6, 723. [Google Scholar] [CrossRef]
- Pandey, P.; Irulappan, V.; Bagavathiannan, M.V.; Senthil-Kumar, M. Impact of Combined Abiotic and Biotic Stresses on Plant Growth and Avenues for Crop Improvement by Exploiting Physio-Morphological Traits. Front. Plant Sci. 2017, 8, 537. [Google Scholar] [CrossRef] [PubMed]
- Choudhury, F.K.; Rivero, R.M.; Blumwald, E.; Mittler, R. Reactive Oxygen Species, Abiotic Stress and Stress Combination. Plant J. 2017, 90, 856–867. [Google Scholar] [CrossRef] [PubMed]
- Suzuki, N.; Rivero, R.M.; Shulaev, V.; Blumwald, E.; Mittler, R. Abiotic and Biotic Stress Combinations. New Phytol. 2014, 203, 32–43. [Google Scholar] [CrossRef] [PubMed]
- Francesca, S.; Vitale, L.; Arena, C.; Raimondi, G.; Olivieri, F.; Cirillo, V.; Paradiso, A.; de Pinto, M.C.; Maggio, A.; Barone, A.; et al. The Efficient Physiological Strategy of a Novel Tomato Genotype to Adapt to Chronic Combined Water and Heat Stress. Plant Biol. 2022, 24, 62–74. [Google Scholar] [CrossRef] [PubMed]
- Sehgal, A.; Sita, K.; Siddique, K.H.M.; Kumar, R.; Bhogireddy, S.; Varshney, R.K.; HanumanthaRao, B.; Nair, R.M.; Prasad, P.V.V.; Nayyar, H. Drought or/and Heat-Stress Effects on Seed Filling in Food Crops: Impacts on Functional Biochemistry, Seed Yields, and Nutritional Quality. Front. Plant Sci. 2018, 871, 1705. [Google Scholar] [CrossRef] [PubMed]
- Fahad, S.; Bajwa, A.A.; Nazir, U.; Anjum, S.A.; Farooq, A.; Zohaib, A.; Sadia, S.; Nasim, W.; Adkins, S.; Saud, S.; et al. Crop Production under Drought and Heat Stress: Plant Responses and Management Options. Front. Plant Sci. 2017, 8, 1147. [Google Scholar] [CrossRef]



Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).