1. Introduction
Anthropogenic activities and modern lifestyles frequently result in the intentional or unintentional release of thousands of chemicals, including pesticides, into the environment. The aquatic ecosystem is one of the most polluted places, since it is the final resting place for most contaminants [
1]. Fish are primarily affected by pesticide hazards due to their absorption through their skin and through gill uptake [
2]. Methomyl (MET) is a carbamate pesticide widely used worldwide due to its high efficacy, high solubility, and wide biological effects [
3]. Its use has contributed to pest management (controlling insects and nematodes), increasing agricultural productivity. Detectable levels of MET residue have been identified in various water bodies and foods due to its high water solubility, extensive use, and agricultural and industrial release into the environment [
4]. MET residue levels in the aquatic environment have been documented to range from 0 to 55.3 g L
−1 [
5].
The World Health Organization (WHO) categorized MET as a very dangerous product (class 1B) [
6]. Its exposure poses a serious health risk and causes toxicity and death as it has been found in humans and animals’ blood, liver, kidneys, and brain [
7]. Several studies revealed that MET exposure causes serious harm to various biological and metabolic processes in fish organs. For instance, Islamy et al. [
8] reported its genotoxic effects on Nile tilapia (
Oreochromis niloticus). Additionally, Wang Yuqin [
9] confirmed the increased oxidative stress in the liver and a decreased growth rate of
O. niloticus due to MET exposure. Meng et al. [
3] recorded the inhibition of the
O. niloticus antioxidant system when exposed to 0.2–200 μg MET/L
−1. Moreover, it reduced proinflammatory cytokine expression and abrogated defense against bacterial infections in tilapia [
10].
The aquaculture industry must develop nutritional strategies capable of mitigating the risk of waterborne pollution while improving the immunological response and growth of different cultured fish species [
11,
12,
13]. Parsley (
Petroselinum crispum) is a culinary herb native to the Mediterranean region. Parsley is a member of the Umbelliferae family and is used in the pharmaceutical, cosmetic, and food industries [
14]. It contains alpha-linolenic acid, a fatty acid important for growth and reproduction [
15]. Flavonoids, terpenoids, carotenoids, myristicin, coumarins, ascorbic acid, and apiole have been identified as major components of parsley [
16]. It boosts and promotes organ activity, enhancing their ability to absorb and utilize nutrients [
17]. Moreover, it has been shown to have various potential medicinal properties, including antioxidant, antimicrobial, antihyperlipidemic, and hepatoprotective effects [
18,
19,
20]. In modern medicine, parsley has been shown to have a wide range of pharmacological activity, including hepatoprotective, neuroprotective, antidiabetic, analgesic, immunostimulant, antioxidant, anti-platelet, cytoprotective, antibacterial, and antifungal properties [
21,
22]. However, there is little information about the effects of using PSM as a fish diet supplement.
Nile tilapia (
Oreochromis niloticus) is a commonly produced fish for human consumption globally [
23]. Toxic chemicals in the water, on the other hand, constitute a threat to
O. niloticus health and growth [
24]. To the best of our knowledge, no scientific published study has been conducted on the potential benefits of PSM dietary supplementation in mitigating the MET-induced adverse effects on the health of
O. niloticus. Consequently, biochemical and histopathological endpoints were assessed, including hematological indices, hepatic enzymes, renal damage products, oxidative stress indices, acetylcholinesterase (AchE) activity, protein profile, and innate immune biomarkers. A comprehensive morphometric histological examination of hepatic and renal tissues in fish exposed to MET and fed a PSM-enriched diet was also carried out. In addition, the fish body’s chemical composition and growth performance were assessed. Moreover, susceptibility to
Pseudomonas aeruginosa infection, one of the most prevalent bacteria that infect fish and causes significant damage of fish tissues and mortality [
25,
26], was also investigated.
2. Materials and Methods
2.1. Tested Compounds and Chemicals
Methomyl (97%) (methyl N-(methylcarbamoyloxy) ethanimidothioate was obtained from Sigma Aldrich (St. Louis, MO, USA). Parsley seeds were purchased in dry packages from the local market for herbs and medicinal plants in Zagazig, Egypt. The dried parsley seeds were washed first, dried, and then crushed and kept at 4 °C till use. All other analytical-grade chemicals were purchased from Sigma-Aldrich, St. Louis, MO, USA.
2.2. Experimental Fish
Apparently healthy fish, Oreochromis niloticus, were gathered from a commercial fish farm in Kafr El-Sheik governorate, Egypt (average body weight: 25.33 ± 0.26 g). Before the experiments, fish were stocked and acclimated for two weeks in 75 L glass aquaria (80 × 40 × 30 cm) with dechlorinated tap water and continual aeration from a central air compressor via an air stone. During the acclimatization period, fish were fed a basal diet. To eliminate excreta, approximately 30% of the water was emptied thrice weekly. The fish were subjected to a photoperiod of 12 h of light and 12 h of darkness. The physicochemical properties of the water used in the aquaria were investigated; the temperature was maintained at 26 ± 0.5 °C, and the pH and dissolved oxygen were kept at 7.5 ± 0.5 and 6.8 ± 0.23 mg L−1, respectively. The averages of ammonia, nitrite, and nitrate were recorded at 0.12 ± 0.02, 0.14 ± 0.04, and 2.30 ± 0.05 mg L−1, respectively.
The trial was conducted at the Fish Diseases and Management Department, Faculty of Veterinary Medicine, Zagazig University, Egypt. This institution’s Animal Use in Research Committee (IACUC) approved the experiment. The experiments were carried out in compliance with the National Institutes of Health’s (NIH) Ethical Guidelines for the Use and Care of Laboratory Animals in Scientific Investigations.
2.3. Diet Formulation and Experimental Plan
Two hundred and twenty-five healthy
O. niloticus fish were randomly dispersed among five different groups. Each fish group was divided into three aquaria, with 15 fish each. In the control group (G1), fish were fed a basal diet without PSM supplementation and reared in aquaria containing clean water (
Table 1), while G2 was exposed to 1/20 LC
50 (20.39 µg L
−1) MET [
10] and fed a basal diet without PSM supplementation, and G3, G4, and G5 received basal diets enriched with 0.5%, 1.0%, and 2.0% PSM, respectively, with concomitant MET exposure at the same concentration. The experimental diet ingredients were well mixed before being mechanically pelleted, air-dried for 24 h at room temperature, and then stored at 4 °C until use. The basal diet was formulated to meet the optimal dietary needs of fish as specified by the Nutrient Requirements of Fish [
27]. Each experimental diet was assessed for crude protein using the macro-Kjeldahl method, crude fat via the ether extraction technique, moisture by a forced-air oven, total ash using a muffle furnace, and crude fiber following AOAC [
28] guidelines. Fish were fed until satiation thrice daily (8:00 a.m., 12:00 p.m., and 16:00 p.m.). The trial lasted 60 days. The total fish weight per aquarium was measured every two weeks to monitor fish growth. Every 48 h, the water was fully replaced by moving the fish to aquaria containing freshly prepared MET solutions.
2.4. Growth Performance Assessment
To assess growth performance, each replicate’s fish was weighed at the start of the trial and every two weeks. The final body weight (FBW), weight gain (WG), specific growth rate (SGR), weight gain percentage, condition factor (K), feed intake (FI), and feed conversion ratio (FCR) were calculated as follows:
FBW = total weight of fish, divided by the fish number in each replicate
WG = total body weight minus initial body weight
Weight gain percentage = [(final average BW − initial average BW)/initial average BW] × 100
SGR = [(Ln final BW − Ln initial BW)/experiment duration (days)] × 100
FI = feed consumed/number of survival fish
FCR = dry feed fed (g)/wet weight gain (g)
Condition factor (K) = (W/L3) × 100
where W = total fish weight (g) and L = fish length (cm), measured from the tip of the snout to the end of the middle caudal fin.
The mortality rate was calculated using the following formula: mortality rate = (number of dead fish/total initial fish number) × 100.
2.5. Whole-Body Chemical Analysis
At the end of the 60-day feeding trial, five fish from each replication were used to analyze the chemical composition of the total fish body. Following AOAC [
28] protocols, crude protein levels were determined by a Kjeldahl distillation unit (UDK 129, Velp Scientifica, Usmate Velate, Italy). Moisture was determined by a natural convection oven (JSON-100, Gongju-City, Korea). The ash content was determined using muffle furnaces (Barnstead/Thermolyne Benchtop 47900, Thermo Scientific, Waltham, MA, USA). Soxhlet extractor glassware was also used to estimate crude lipids.
2.6. Blood and Tissue Sampling
After the 60-day feeding trial, blood was drawn from caudal vessels and split into two halves. The first half was obtained with EDTA tubes for a complete blood cell count. The second was collected without an anticoagulant until clotting happened; the tubes were kept at 4 °C overnight and centrifuged at 150× g for 5 min, followed by 350× g for 15 min. The serum was utilized to test immunological function, serum protein electrophoretic pattern, oxidant/antioxidant status, and liver and kidney functions. After collecting blood samples, the fish were dissected to obtain the liver and kidney, then fixed for 48 h in 10% neutral buffered formalin for histopathological investigations.
2.7. Estimation of Hematological Values
Hematological indices such as red blood cell counts (RBCs), packed cell volume (PCV), mean corpuscular volume (MCV), hemoglobin content (Hb), mean corpuscular hemoglobin (MCH), and mean corpuscular hemoglobin concentration (MCHC) were determined using a Hema Screen 18 automatic hematology analyzer (Hospitex Diagnostics, Sesto Fiorentino, Italy). Dacie and Lewis’s manual technique was used for counting total leukocytes (WBCs), differential leukocyte counts (lymphocytes, neutrophils, and monocytes), and platelets [
29].
2.8. Serum Biochemical Analysis
2.8.1. Hepatorenal Damage Products
Alanine aminotransferase (ALT), aspartate aminotransferase (AST), lactate dehydrogenase (LDH), alkaline phosphatase (ALP), creatinine, urea, ammonia, total bilirubin, and cholesterol were assessed in serum by Spinreact kits (Esteve De Bas, Girona, Spain) in accordance to the protocols described by Burtis and Ashwood [
30], Wenger et al. [
31], Murray and Kaplan [
32], Pesce [
33], Neely and Phillipson [
34], Martinen [
35], Naito [
36], Fossati et al. [
37], and Kaplan and Glucose [
38], respectively.
2.8.2. Immune System Response
Serum complement3 (C3) levels were measured by fish-specific ELISA kits and the manufacturer’s instructions. Serum lysozyme activity was determined using spectrophotometry [
39]. The concentration of nitric oxide (NO) was determined following the protocol of Dumock [
40].
2.8.3. Stress, Inflammatory Status Assays, and Acetyl Choline Esterase Enzyme (AchE) Activity
Stress indicators were measured by commercial colorimetric kits of Biodiagnostic Co. (Cairo, Egypt). The catalase enzyme (CAT) level was determined using the Aebi [
41] method. The superoxide dismutase (SOD) activity was measured by the Nishikimi et al. [
42] protocol. Malondialdehyde (MDA) was measured according to the Uchiyama and Mihara [
43] method. The EnzyChromTM Glutathione Peroxidase Assay Kit (EGPX-100) of Bio-Assay Systems (Hayward, CA, USA) was used to determine quantitative colorimetric glutathione peroxidase (GPx) [
44].
Myeloperoxidase (MPO) activity in fish serum was determined based on Kumari and Sahoo [
45]. Fish ELISA kits of My Biosource Co. for tumor necrosis factor-alpha (TNF-α) were used according to the instructions in the enclosed pamphlets of the kits. The acetylcholinesterase (AchE) activity was evaluated according to Ellman et al. [
46]. According to Badawi [
47], the electrophoretic pattern of serum proteins, comprising total proteins, albumin, and globulins, was analyzed.
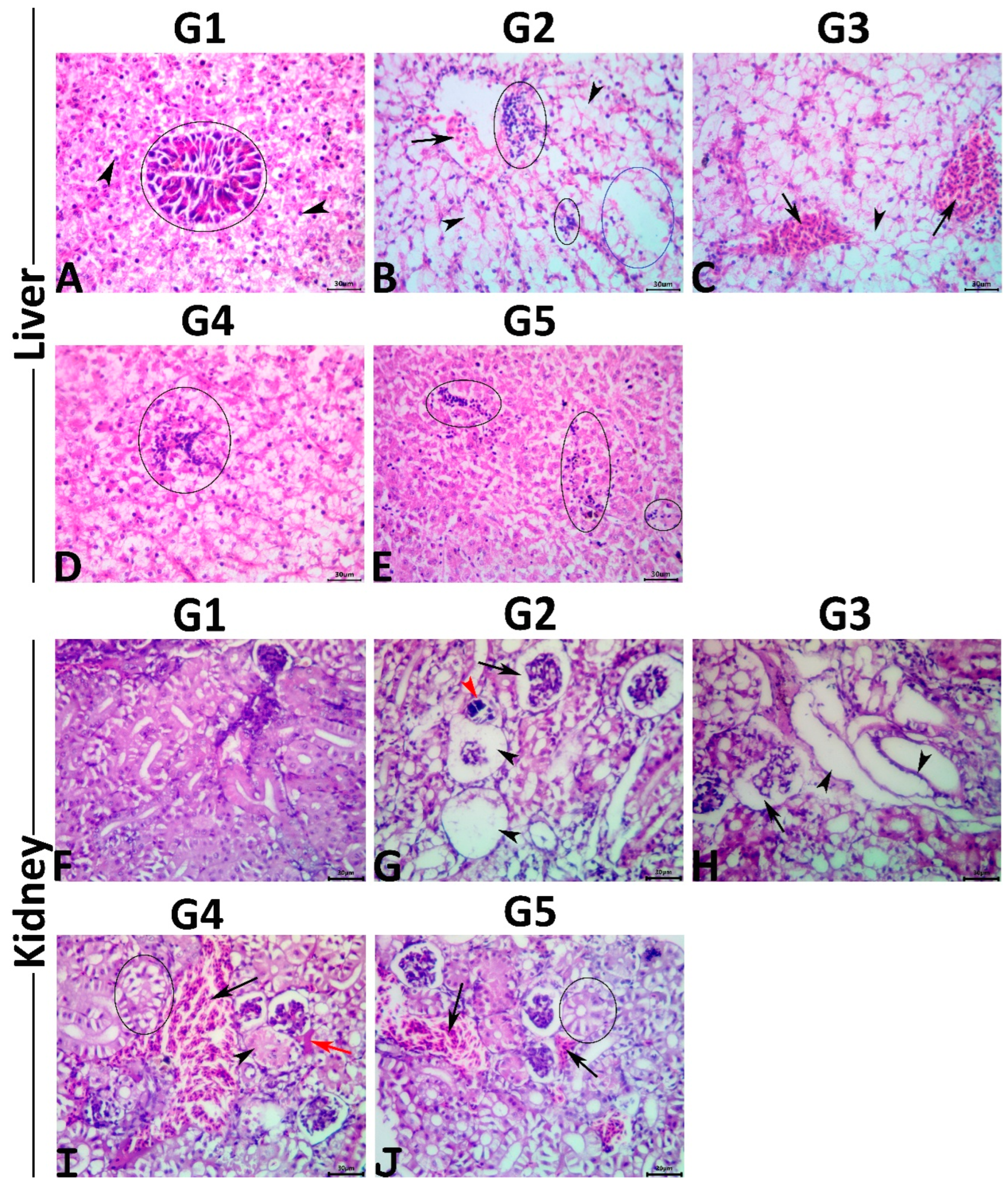
2.9. Histopathological Study
At the end of the trial, the fish were necropsied using standard finfish necropsy procedures [
48]. Liver and kidney tissue samples were obtained from nine randomly selected fish per group, rinsed in distilled water, and immediately fixed in a 10% neutral buffered formalin solution for 48 h. Post fixation, the specimens were processed using the paraffin technique, sectioned at four μm thick, and stained with hematoxylin and eosin following the protocol described by Suvarna et al. [
49]. The stained slides were examined microscopically for morphological alterations. A multiparametric numerical histopathological assessment of the hepatic and renal tissues was performed with minor modifications to the protocol proposed by Bernet et al. [
50]. In brief, five non-overlapped randomly chosen microscopic fields (10 objectives) per organ per fish were snapshotted, and the images were then analyzed to calculate lesion frequencies, liver and kidney indices (the higher the indices, the worse the histopathological alterations), and the total indices for a fish using the following formulas:
The following formula determined the frequency of lesions (FQ):
where (N
lesion) represents fish that exhibited a lesion, and (N
total) represents the total number of fish in the group.
The liver and kidney indices were calculated by the following formula:
where (rp) is the reaction pattern, (alt) the histopathological alteration, (
a) the score value (indicates the size of tissue affected by the alteration, and its value ranged between six (diffuse lesion) and zero (lack of the alteration)), and (
w) the importance factor (indicates how much worse the alteration is, with its value ranging between three and one).
The total index for each individual fish was calculated by adding the indices for the liver and kidney according to the formula:
2.10. Challenge Test
Pseudomonas aeruginosa (previously isolated from naturally infected O. niloticus and previously identified and validated as pathogenic) was dispersed on tryptic soy agar (TSA) (Hi-media, Thane West, Maharashtra, India) and incubated for 48 h at 30 °C. The P. aeruginosa lethal dose (LD50) was evaluated. Fish were intraperitoneally (IP) injected with diverse doses of 24-h live bacteria; then, the infected fish mortality was reported for 3-days post-injection. The LD50, which resulted in 50% fish mortality, was 1 × 108 CFU/mL. Five fish per replicate (n = 15 fish/group) were intraperitoneally injected with 0.5 mL of the produced bacterial solution containing 1 × 108 CFU/mL after the feeding trial (60 days). The injected fish were monitored for two weeks to determine the percentage of mortality and post-mortem lesions.
2.11. Statistical Analysis
Using SPSS, the data were statistically investigated using one-way Analysis of Variance (ANOVA) (version 16.0, SPSS Inc., Chicago, IL, USA). A Tukey’s multiple comparisons post hoc test was used to compare means between groups, with statistical significance set at p ˂ 0.05. The results were displayed as means ± standard error (SE).
4. Discussion
In the present study, MET exposure significantly reduced the
O. niloticus growth performance in FBW, DWG, SGR, and feed utilization values. These findings followed the study of Mohamed et al. [
10], which verified that chronic exposure to MET for 60 days resulted in low growth performance in
O. niloticus. Additionally, Trachantong et al. [
51] reported a decrease in the amphibian growth rate after MET exposure. The MET-induced growth retardation could be related to toxic stress-induced metabolic alterations in protein and carbohydrate metabolism, resulting in energy storage to repair tissue damage, leaving limited energy for growth. Moreover, MET could induce endocrine-disruption, resulting in smaller sizes, as previously reported in exposed tadpoles [
51]. Remarkably, concurrent PSM administration in the
O. niloticus diet improved growth even with MET exposure. It has been demonstrated that spices, herbs, and plant extracts can increase appetite and digestion in fish while also having antibacterial properties [
12,
52,
53]. The PSM growth-augmenting effect could be attributed to the richness of parsley with active compounds (phenols, flavonoids, turbines, and glycosides) that serve as digestive system catalysts by enhancing the elasticity of the intestine’s vasculature [
54], thus enhancing the secretion of enzymes (trypsin, chymotrypsin, amylase, lipase) that help in digestive function and excretion [
55]. Moreover, the antimicrobial action of parsley is dominated by modulation of the gastrointestinal microbiota [
21]. In line with our findings, improved feed conversion rate and growth were demonstrated in
Cyprinus carpio, which were fed parsley-supplemented diets [
56]. Previous studies have shown that parsley has a comparable protective activity against aflatoxins [
57] or bifenthrin [
58] in
O. niloticus. The mortality of
O. niloticus was increased by 13.33% after exposure to MET in the current investigation, indicating its significant toxicity. This could be due to MET-induced oxidative stress, immunotoxicity, genotoxicity, and neurobehavioral toxicity, which have previously been documented [
10]. Moreover, MET is a potent neurotoxin as it inhibits the AChE enzyme activity, as recorded in the current study. Likely, Jablonski et al. [
59] reported that MET affected embryo hatching and larva shape and behavior of zebrafish, resulting in smaller bodies and eyes, failure to inflate the swimming bladder, and hypolocomotor and highly neurotoxic activity. The PSM-enriched feed given concurrently with MET exposure, on the other hand, diminished mortality percent, perhaps via boosting growth performance and immunological response, as well as alleviation of oxidative damage [
58]. Moreover, the mitigating effect of parsley was detected on brain neurons, neurotransmitter levels, and neurobehavioral performance in Cd-treated mice [
60].
Tissue protein and lipid content are influenced by the dynamic equilibrium between their production and breakdown rates. In this study, the decrease in lipid content and crude protein was evident in the chemical composition analysis of fish exposed to MET. The former observation could be linked to cellular fraction disruption and the resulting impairment in protein synthesis machinery. The intense proteolysis and catabolic effects caused by MET toxicity release free amino acids to meet the increased energy needs caused by toxins [
61]. In times of stress, active glycogenolysis and glycolytic pathways offer surplus energy, resulting in decreased crude lipid content [
62]. Many studies reported a similar decrease in protein levels and carbohydrate levels [
63]. In contrast, PSM dietary supplementation restored crude protein and lipid deposition in the
O. niloticus muscles at the end of the feeding experiment. This could be due to parsley’s modulatory effect on the activity of fish digestive enzymes and intestinal microbiota, which improves the utilization of digestive products [
58]. Increased expression of fatty acid synthesis-related markers and up-regulation of urea cycle components suggest a recovery in oxidative stress and glycolysis [
64].
Blood biochemical indicators are crucial for assessing fish health, toxicity, and biomonitoring [
65,
66,
67,
68]. Herein, fish exposed to MET developed apparent anemia and a significant decrease in total leukocyte count with a parallel decline in heterophils, lymphocytes, monocytes, and eosinophils. Increased RBCs destruction, osmoregulatory impairment, or decreased erythropoiesis can cause a decline in RBC count [
69]. As a compensatory reaction to maintaining gas exchange in damaged gills while decreasing oxygen-carrying capacity, decreases in Hb and PCV values were observed. The decreased leukocyte production could be attributed to depression of leukopoiesis, alteration of the cell membrane or disintegration of white blood cells, and a disruption in the innate immune response [
70]. Additionally, lymphocytopenia may be connected to cortisol, a stress hormone that induces lymphocyte death or redistribution from the blood to the tissues [
71]. In line with our findings, Mohamed et al. [
10] found a decrease in erythrocytic, leukocytic, and Hb content in tilapia fish exposed to MET. MET also affected the erythrogram and total and differential leukocytic counts in rats’ blood [
72]. Notably, PSM dietary supplementation significantly reversed the MET-induced hematological alterations. Similarly, parsley extract completely reversed the Hb decrease in
O. niloticus with aflatoxin contamination [
57]. Additionally, the hematological indices of
O. niloticus fish exposed to MET and treated with taurine showed a significant improvement [
10]. This could be due to PSM’s ability to protect the Hb molecule and the erythrocyte membrane from oxidative damage and the resumption of erythropoiesis. The increase in leukocyte and lymphocytic counts may be linked to preserving the leukocyte redox state and the immunostimulant impact of parsley, which boosts leukocyte proliferation.
When a fish is exposed to various water toxicants, its innate immune system, the first line of pathogen defense, is compromised [
73]. The innate immune response system relies heavily on various cellular and humoral components [
74]. Lysozymes hydrolyze the peptidoglycan layer of the bacterial cell wall by acting as opsonins, causing the complement system and phagocytes to activate [
39]. In fish, complement C3 performs various immunological tasks, including removing invading pathogens, initiating inflammatory responses, and clearing other homeostatic cells [
75]. NO, which boosts macrophages’ ability to eliminate infections while also playing a role in the inflammatory response, is another key sign of innate immunity [
76]. Herein, immunological responses (lysozyme activity, NO, and C3) were significantly reduced in fish exposed to MET, indicating the innate immune response suppression. The decrease in lysozyme after MET exposure in fish could be attributed to decreased leukocyte count and function, including neutrophils and macrophages [
77]. Moreover, the noted reduction in NO amount may reveal declined phagocytosis. The immunosuppressive impact of MET could be due to a toxic insult that lowers total leukocytic and lymphocytic counts. It could also result from uncontrolled inflammatory cytokine production and increased lymphoid organ and immune cell apoptosis. Furthermore, MET has been implicated as an endocrine disruptor [
51]. In agreement with our findings, the activity of serum lysozyme, C3, and NO was reduced in
O. niloticus after 60 days of exposure to MET [
10].
Serum protein, albumin, and globulin levels are also linked to innate immunity [
78]. Following 60 days of exposure to MET in
O. niloticus, a significant fall in total serum protein, albumin, and globulin was detected, indicating immunoglobulin underproduction [
79]. Hypoproteinemia was triggered due to catabolism of protein and/or malfunction of the liver [
80]. As a result, the depleted immune components and the hypoproteinemic condition were restored when PSM was supplemented. This immunostimulant effect corresponds to an improvement in total leukocytic and lymphocytic counts. Previous research has shown that parsley boosts humoral and cellular immunity [
81,
82].
During normal metabolism, reactive oxygen species (ROS) such as superoxide anions and hydroxyl radicals are generated in aerobic cells, mostly through mitochondrial oxidative metabolism, and some of these intermediates are thought to be harmful to cells, causing oxidative stress and oxidative damage [
83]. Nonetheless, aerobic organisms have evolved mechanisms to activate antioxidant defense systems in different organs and tissues, which include antioxidant enzymes like CAT, SOD, GR, GPx, and GST, as well as antioxidant scavengers to protect against damage caused by high levels of ROS [
84]. In the current study, noticeable exhaustion of the GPx, CAT, and SOD activities but increased inflammatory markers (MPO and TNF-
α) and MDA concentrations were recorded in fish exposed to MET. It is possible to hypothesize that under conditions of chronic exposure to MET, it could not be metabolized and was not entirely eliminated from the tilapia body by the cells. As a result, the MET accumulated in the body and destroyed the protein structures of the antioxidant enzymes, resulting in decreased antioxidant enzyme activity, according to Jagetia and Aggarwal [
85]. The conjugation of MET or its degradation products to polyunsaturated fatty acids may be blamed for the MET-induced elevation in MDA levels [
86]. In this regard, Mohamed et al. [
10] proved that the main toxic effects of MET in
O. niloticus take place via oxidative stress, resulting in oxidative damage through reducing antioxidant enzyme activity and inducing DNA damage and lipid peroxidation. As proven by the fact that MET can cause oxidative damage, LPO levels are raised, and several antioxidant enzymes are disrupted [
87]. In contrast, PSM significantly suppresses the oxidative and lipid peroxidative damage induced by MET by stimulating the production of antioxidant enzymes and thereby boosting cellular antioxidant defenses. Parsley is abundant in polyphenolic flavonoid anti-oxidants such as apiin, apigenin, and luteolin [
88]. Along with luteolin and vitamin C, parsley acts as an anti-inflammatory agent, enhancing catalase and glutathione activity and lowering lipid peroxidation [
89]. Previous reports have evidenced parsley efficiency in heavy metal excretion from the body [
60,
90].
The liver is the first organ to be exposed to any foreign molecule delivered by portal circulation, and it sustains the most damage. The release of intracellular enzymes such as AST, ALT, LDH, and ALP into the circulation is one of the most sensitive and dramatic indications of hepatocyte injury in response to toxicity [
12,
91]. The current findings revealed a significant increase in the activities of AST, ALT, LDH, and ALP in the serum of fish together with metabolic products such as cholesterol and bilirubin, implying that MET could cause liver damage. This changed biochemical profile was linked to a variety of hepatic lesions, including lymphocytic infiltration, coagulative necrotic foci, congestion, sinusoidal dilatations, and melanomacrophage aggregate hyperplasia. Changes in membrane permeability and loss of functional integrity of cell membranes in the liver also cause cellular leakage, resulting in the pervasive release of these enzymes from the cell [
92]. Furthermore, MET-induced hypercholesterolemia may be due to organisms’ requirements for additional energy reserves to cope with stressors’ effects [
93]. Our observed increase in these parameters due to MET intoxication is in accordance with the findings of Hashish and Elgaml [
94]. Restoration of the disturbed enzyme activities and hepatic architecture with the correction of metabolic product concentrations after PSM supplementation is the primary explanation for their hepatoprotective effects. Indeed, the hepatoprotective effect of parsley arises from its capacity to scavenge ROS, as reported in numerous assays [
95,
96]. The decreased LDH and hypocholesterolemia due to the concurrent treatment with 20% parsley seed methanol extract was documented in rats [
97]. The stabilization of serum bilirubin levels by PSM is a clear indication of the improvement in the functional status of the liver cells. This activity could be attributed to the phytochemical analysis’s bioactive components, including flavonoids, lignans, alkaloids, bisbenzyl, coumarins, and terpenes [
98].
Fish kidneys gather the most post-branchial blood, so renal diseases could be effective indicators of environmental contamination [
99]. MET caused a considerable increase in serum ammonia, urea, and creatinine activity in the present study. The elevated levels of ammonia, urea, and creatinine may be linked to the effects of MET on muscle metabolism and its limited impact on purine metabolism and renal tubule injury. These findings matched those of Hashish and Elgaml [
94]. Conversely, the present study found that PSM supplementation might reverse MET-induced nephrotoxicity in
O. niloticus fish, as indicated by restoration of the disturbed metabolites and renal architecture. Apigenin and its glucosidal flavonoids, which are abundant in parsley, have been discovered to have anti-inflammatory, antioxidant, and anticancer properties, particularly in the case of renal inflammation [
100]. A similar nephroprotective effect of PSM has been previously recognized [
101].
In the current study,
O. niloticus was challenged with
P. aeruginosa after MET exposure for 60 days. The findings demonstrated that the RPS was dropped noticeably in MET-exposed and fed basal diet following the challenge. Moreover, severe abnormal behavior and clinical signs were observed. The loss of cellular and humoral immune components caused by MET could be a major contributor to lowering the RPS and, as a result, compromising the fish response to
P. aeruginosa in the challenge. Importantly, when PSM was added to the fish diet with MET exposure, it improved the outcomes of suppressed innate immunity against
P. aeruginosa, which could be strongly linked to the immune-stimulatory activity observed in this trial. Correspondingly, Farag et al. [
58] demonstrated that dietary parsley essential oil supplementation in the
O. niloticus diet alleviated the adverse effects of bifenthrin and enhanced disease resistance against
Aeromonas hydrophila.
Importantly, the effects of new feed ingredients or additives on fish health indicators and safety should be carefully considered for an appropriate assessment of any nutritional strategy [
13]. In this regard, the liver and kidney function indicators are important determinants in assessing feed ingredient safety in fish diets because of their roles in detoxification, biotransformation, and excretion of xenobiotics [
102,
103]. Of note, in the current study, supplementing fish with diets fortified with up to 2% of PSM, even with MET exposure, did not negatively impacted the liver and kidney function indicators, indicating a wide safety margin of PSM as a feed supplement in fish.


 ,
, 



{kind=link}