
Phytochemicals, Nutrition, Metabolism, Bioavailability, and Health Benefits in Lettuce—A Comprehensive Review

 , ,
, ,  , , ,
, , ,  ,
,  , ,
, ,  and
and 
Abstract
:1. Introduction
2. Methodology
3. Lettuce Types and Production
4. Taxonomical Classification
4.1. Crisphead
4.2. Butterhead
4.3. Loose-Leaf
4.4. Romaine
5. Phytochemicals and Nutritional Composition of Lettuce
5.1. Phenolic Compounds
5.2. Carotenoids
5.3. Chlorophyll
5.4. Vitamins
5.5. Minerals
5.6. Anti-Nutrients Compounds
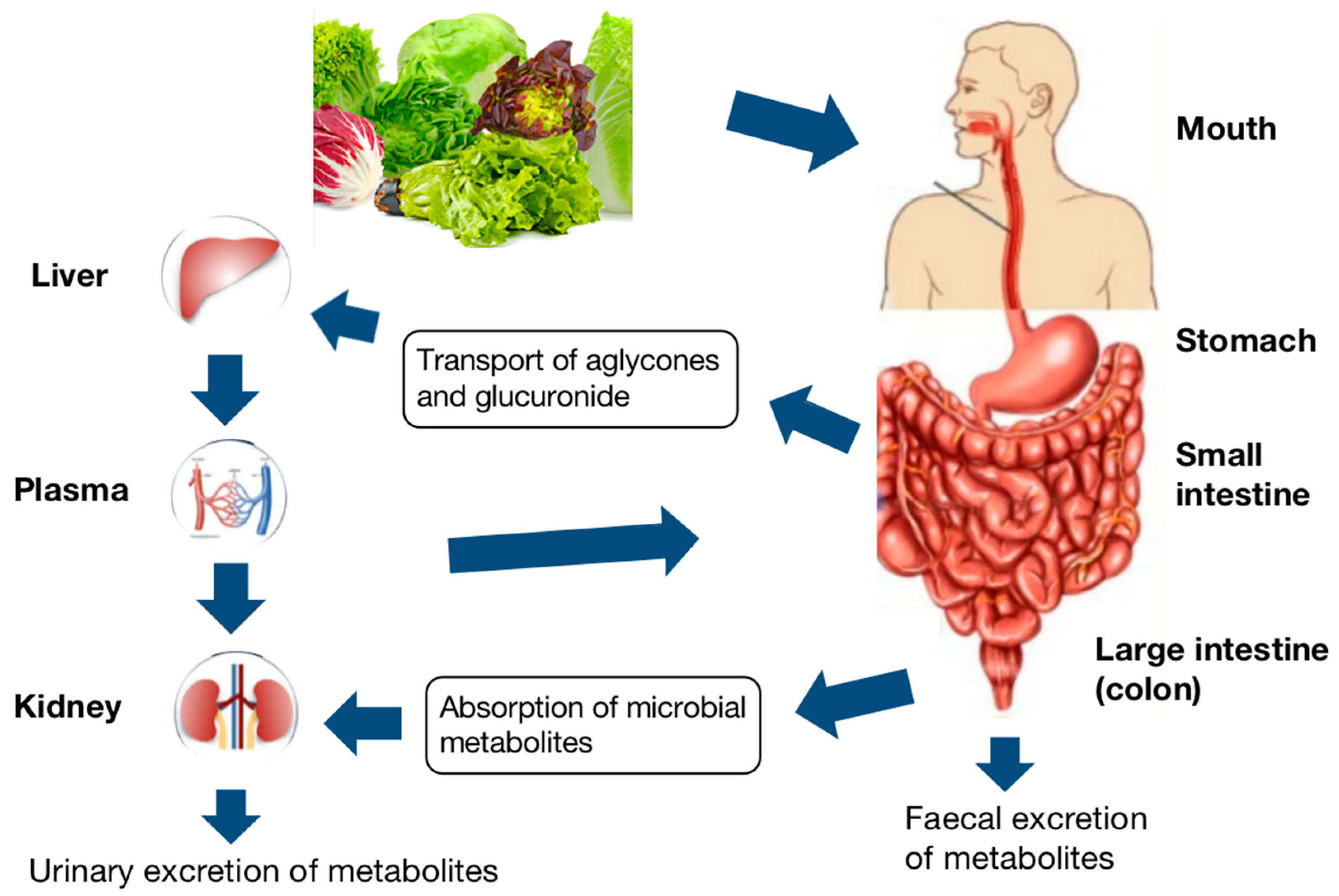
6. Bioavailability of Bioactive Compounds from Lettuce
7. Digestion, Absorption, and Metabolism
7.1. Phenolic Compounds
7.2. Carotenoids
7.3. Vitamin C
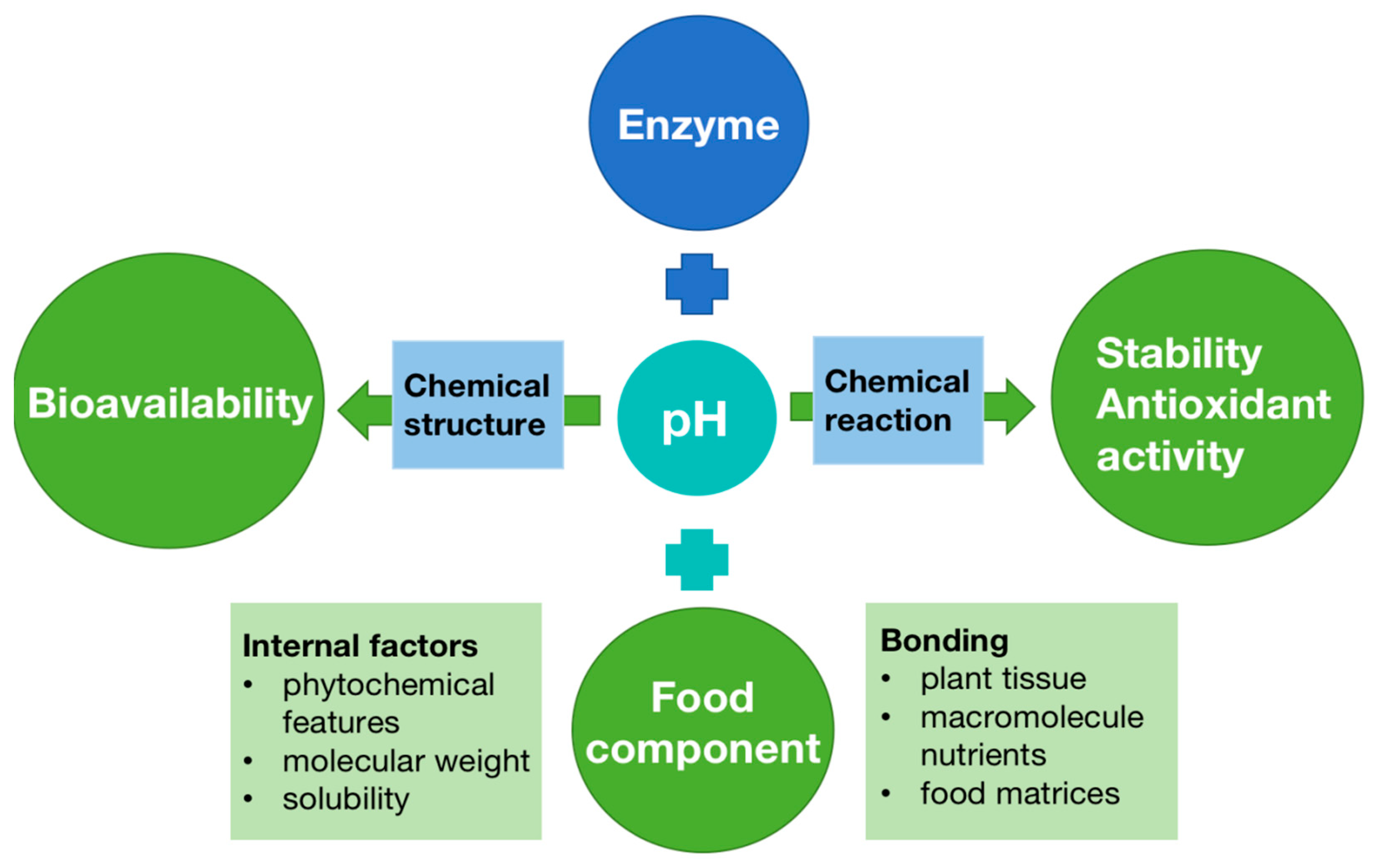
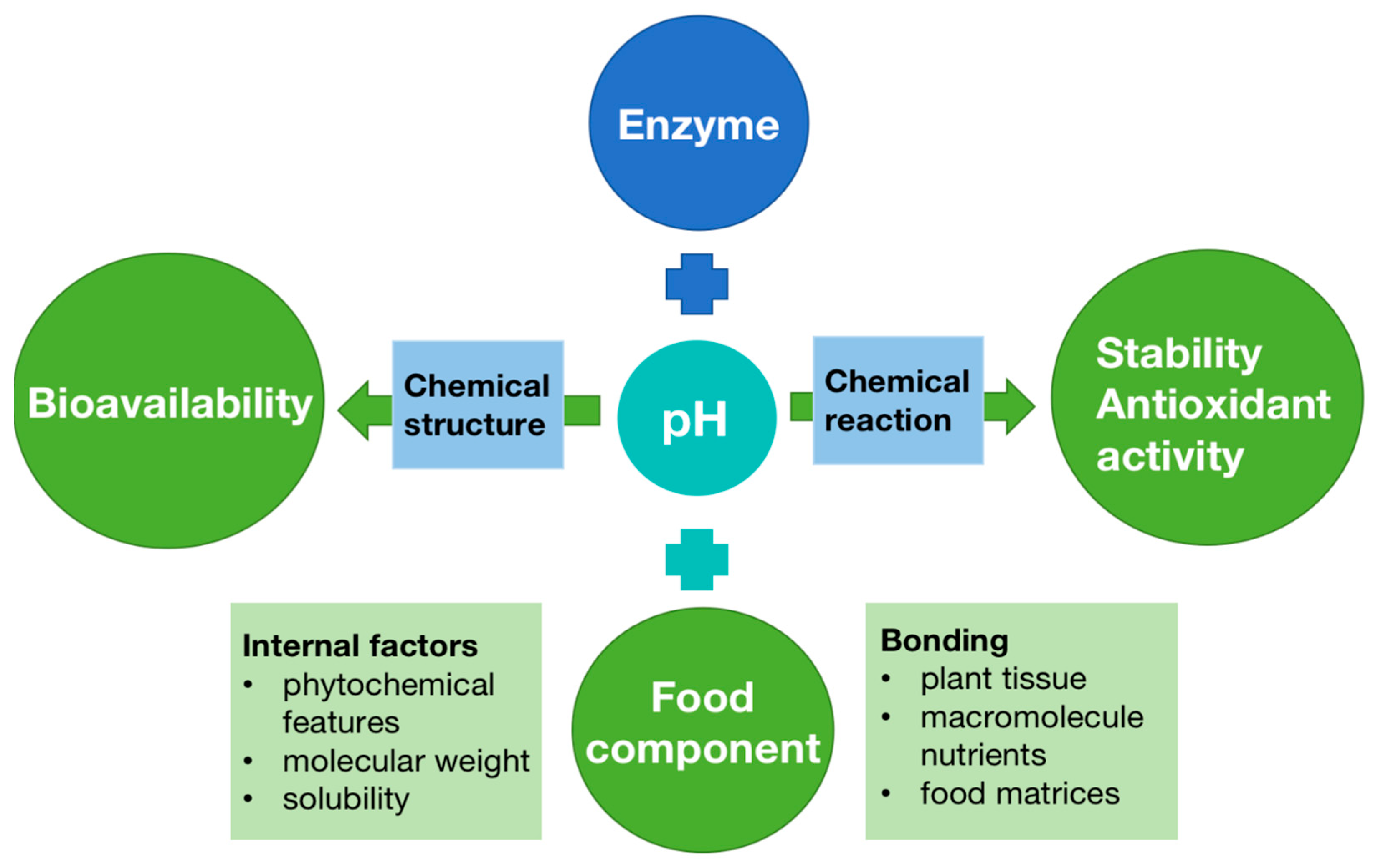
7.4. Factors Affecting the Bioavailability
8. Health Benefits and Risks of Lettuce
8.1. Healthy Benefits
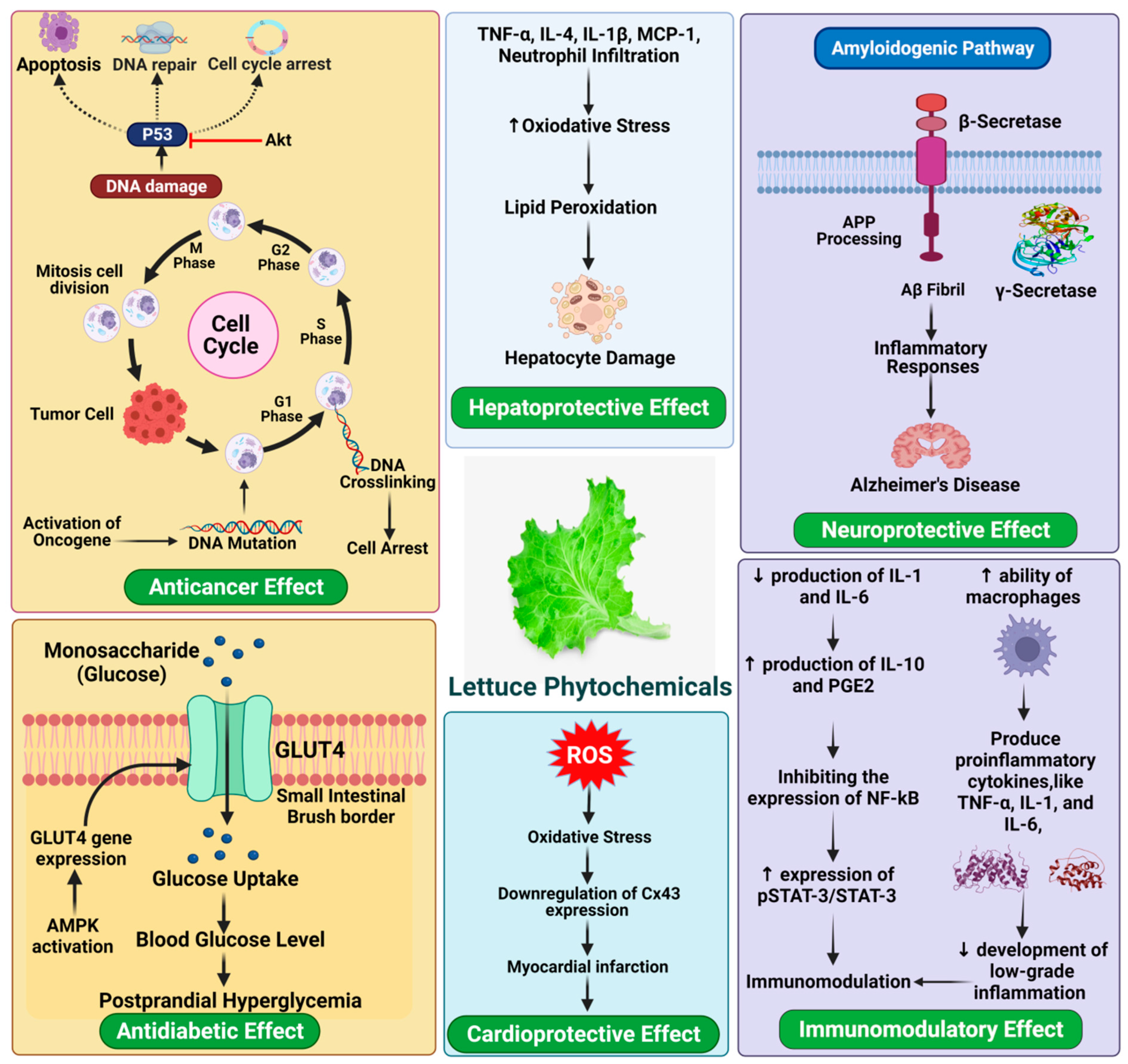
8.1.1. Cardio-Protective Effect
8.1.2. Anti-Cancer Effect
8.1.3. Anti-Diabetic Effect
8.1.4. Immunomodulatory Effect
8.1.5. Hepatoprotective Effect
8.1.6. Neuroprotective Activity
8.1.7. Antioxidant properties
8.1.8. Other Health Benefits
8.2. Health Risks
9. Future Perspectives
10. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Funk, V.A.; Bayer, R.J.; Keeley, S.; Chan, R.; Watson, L.; Gemeinholzer, B.; Schilling, E.; Panero, J.L.; Baldwin, B.G.; Garcia-Jacas, N. Everywhere but Antarctica: Using a supertree to understand the diversity and distribution of the Compositae. Biol. Skr. 2005, 55, 343–374. [Google Scholar]
- Mampholo, B.M.; Maboko, M.M.; Soundy, P.; Sivakumar, D. Phytochemicals and overall quality of leafy lettuce (Lactuca sativa L.) varieties grown in closed hydroponic system. J. Food Qual. 2016, 39, 805–815. [Google Scholar] [CrossRef]
- Yang, X.; Gil, M.I.; Yang, Q.; Tomás-Barberán, F.A. Bioactive compounds in lettuce: Highlighting the benefits to human health and impacts of preharvest and postharvest practices. Compr. Rev. Food Sci. Food Saf. 2022, 21, 4–45. [Google Scholar] [CrossRef] [PubMed]
- Wilson, P.; Morrison, S.; Hedges, L.; Kerkhofs, N.; Lister, C. Phenolics contribute significantly to higher antioxidant activity of red lettuce compared to green lettuce. In Proceedings of the XXII International Conference on Polyphenols, Helsinki, Finland, 25–28 August 2004; pp. 273–274. [Google Scholar]
- Medina-Lozano, I.; Bertolín, J.R.; Díaz, A. Nutritional value of commercial and traditional lettuce (Lactuca sativa L.) and wild relatives: Vitamin C and anthocyanin content. Food Chem. 2021, 359, 129864. [Google Scholar] [CrossRef] [PubMed]
- Coria-Cayupán, Y.S.; Sánchez de Pinto, M.a.I.; Nazareno, M.n.A. Variations in bioactive substance contents and crop yields of lettuce (Lactuca sativa L.) cultivated in soils with different fertilization treatments. J. Agric. Food Chem. 2009, 57, 10122–10129. [Google Scholar] [CrossRef]
- Hung, H.-C.; Joshipura, K.J.; Jiang, R.; Hu, F.B.; Hunter, D.; Smith-Warner, S.A.; Colditz, G.A.; Rosner, B.; Spiegelman, D.; Willett, W.C. Fruit and vegetable intake and risk of major chronic disease. J. Natl. Cancer Inst. 2004, 96, 1577–1584. [Google Scholar] [CrossRef] [Green Version]
- Sepehri, N.Z.; Parvizi, M.M.; Habibzadeh, S.; Handjani, F. Lettuce as an Effective Remedy in Uremic Pruritus: Review of the Literature Supplemented by an In Silico Study. Evid. Based Complementary Altern. Med. 2022, 2022, 4231854. [Google Scholar] [CrossRef]
- Al Mahmud, Z.; Emran, T.B.; Qais, N.; Bachar, S.C.; Sarker, M.; Uddin, M.M.N. Evaluation of analgesic, anti-inflammatory, thrombolytic and hepatoprotective activities of roots of Premna esculenta (Roxb). J. Basic Clin. Physiol. Pharmacol. 2016, 27, 63–70. [Google Scholar] [CrossRef]
- Rein, M.J.; Renouf, M.; Cruz-Hernandez, C.; Actis-Goretta, L.; Thakkar, S.K.; da Silva Pinto, M. Bioavailability of bioactive food compounds: A challenging journey to bioefficacy. Br. J. Clin. Pharmacol. 2013, 75, 588–602. [Google Scholar] [CrossRef] [Green Version]
- Lafarga, T.; Villaró, S.; Rivera, A.; Bobo, G.; Aguiló-Aguayo, I. Bioaccessibility of polyphenols and antioxidant capacity of fresh or minimally processed modern or traditional lettuce (Lactuca sativa L.) varieties. J. Food Sci. Technol. 2020, 57, 754–763. [Google Scholar] [CrossRef]
- Uddin, M.Z.; Rana, M.S.; Hossain, S.; Ferdous, S.; Dutta, E.; Dutta, M.; Emran, T.B. In vivo neuroprotective, antinociceptive, anti-inflammatory potential in Swiss albino mice and in vitro antioxidant and clot lysis activities of fractionated Holigarna longifolia Roxb. bark extract. J. Complement. Integr. Med. 2020, 17, 1–12. [Google Scholar] [CrossRef] [PubMed]
- Mitra, S.; Lami, M.S.; Uddin, T.M.; Das, R.; Islam, F.; Anjum, J.; Hossain, M.J.; Emran, T.B. Prospective multifunctional roles and pharmacological potential of dietary flavonoid narirutin. Biomed. Pharmacother. 2022, 150, 112932. [Google Scholar] [CrossRef] [PubMed]
- Mitra, S.; Tareq, A.M.; Das, R.; Emran, T.B.; Nainu, F.; Chakraborty, A.J.; Ahmad, I.; Tallei, T.E.; Idris, A.M.; Simal-Gandara, J. Polyphenols: A first evidence in the synergism and bioactivities. Food Rev. Int. 2022, 2022, 1–23. [Google Scholar] [CrossRef]
- Mitra, S.; Rauf, A.; Tareq, A.M.; Jahan, S.; Emran, T.B.; Shahriar, T.G.; Dhama, K.; Alhumaydhi, F.A.; Aljohani, A.S.; Rebezov, M. Potential health benefits of carotenoid lutein: An updated review. Food Chem. Toxicol. 2021, 154, 112328. [Google Scholar] [CrossRef] [PubMed]
- Serafini, M.; Bugianesi, R.; Salucci, M.; Azzini, E.; Raguzzini, A.; Maiani, G. Effect of acute ingestion of fresh and stored lettuce (Lactuca sativa) on plasma total antioxidant capacity and antioxidant levels in human subjects. Br. J. Nutr. 2002, 88, 615–623. [Google Scholar] [CrossRef] [Green Version]
- Naseem, S.; Ismail, H. In vitro and in vivo evaluations of antioxidative, anti-Alzheimer, antidiabetic and anticancer potentials of hydroponically and soil grown Lactuca sativa. BMC Complementary Med. Ther. 2022, 22, 30. [Google Scholar] [CrossRef]
- Kabir, M.; Uddin, M.; Jeandet, P.; Emran, T.B.; Mitra, S.; Albadrani, G.M.; Sayed, A.A.; Abdel-Daim, M.M.; Simal-Gandara, J. Anti-Alzheimer’s molecules derived from marine life: Understanding molecular mechanisms and therapeutic potential. Mar. Drugs 2021, 19, 251. [Google Scholar] [CrossRef]
- Bahbah, E.I.; Ghozy, S.; Attia, M.S.; Negida, A.; Emran, T.B.; Mitra, S.; Albadrani, G.M.; Abdel-Daim, M.M.; Uddin, M.; Simal-Gandara, J. Molecular mechanisms of astaxanthin as a potential neurotherapeutic agent. Mar. Drugs 2021, 19, 201. [Google Scholar] [CrossRef]
- Vázquez, G.; Santos, J.; Freire, M.S.; Antorrena, G.; González-Álvarez, J. Extraction of antioxidants from eucalyptus (Eucalyptus globulus) bark. Wood Sci. Technol. 2012, 46, 443–457. [Google Scholar] [CrossRef]
- Page, M.J.; McKenzie, J.E.; Bossuyt, P.M.; Boutron, I.; Hoffmann, T.C.; Mulrow, C.D.; Shamseer, L.; Tetzlaff, J.M.; Akl, E.A.; Brennan, S.E. The PRISMA 2020 statement: An updated guideline for reporting systematic reviews. Int. J. Surg. 2021, 88, 105906. [Google Scholar] [CrossRef]
- Jaiswal, A.K. Nutritional Composition and Antioxidant Properties of Fruits and Vegetables; Academic Press: Cambridge, MA, USA, 2020. [Google Scholar]
- Shatilov, M.V.; Razin, A.F.; Ivanova, M.I. Analysis of the world lettuce market. IOP Conf. Ser. Earth Environ. Sci. 2019, 395, 012053. [Google Scholar] [CrossRef] [Green Version]
- Tony, N. Field lettuce production. NSW Agric. 2004, 2004, 1–16. [Google Scholar]
- Mitchell, J.; Horwath, W.; Southard, R.; Baker, J.; Munk, D.; Hembree, K.; Klonsky, K.; DeMoura, R.; Miyao, G. Expansion of conservation tillage initiatives in California. In Proceedings of the California Plant and Soil Conference, University of California, Berkeley, CA, USA, 14 May 2002. [Google Scholar]
- Mohamed, S.J.; Rihan, H.Z.; Aljafer, N.; Fuller, M.P. The Impact of Light Spectrum and Intensity on the Growth, Physiology, and Antioxidant Activity of Lettuce (Lactuca sativa L.). Plants 2021, 10, 2162. [Google Scholar] [CrossRef] [PubMed]
- Rader, H.B.; Karlsson, M.G. Northern field production of leaf and romaine lettuce using a high tunnel. HortTechnology 2006, 16, 649–654. [Google Scholar] [CrossRef] [Green Version]
- Simko, I.; Hayes, R.J.; Mou, B.; McCreight, J.D. Lettuce and spinach. In Yield Gains in Major US Field Crops; American Society of Agronomy: Madison WI, USA, 2014; Volume 33, pp. 53–85. [Google Scholar]
- Dai, J.; Mumper, R.J. Plant phenolics: Extraction, analysis and their antioxidant and anticancer properties. Molecules 2010, 15, 7313–7352. [Google Scholar] [CrossRef]
- Hodges, D.M.; Forney, C.F. Postharvest ascorbate metabolism in two cultivars of spinach differing in their senescence rates. J. Am. Soc. Hortic. Sci. 2003, 128, 930–935. [Google Scholar] [CrossRef] [Green Version]
- López, A.; Javier, G.-A.; Fenoll, J.; Hellín, P.; Flores, P. Chemical composition and antioxidant capacity of lettuce: Comparative study of regular-sized (Romaine) and baby-sized (Little Gem and Mini Romaine) types. J. Food Compos. Anal. 2014, 33, 39–48. [Google Scholar] [CrossRef]
- Zhao, X.; Carey, E.E.; Young, J.E.; Wang, W.; Iwamoto, T. Influences of organic fertilization, high tunnel environment, and postharvest storage on phenolic compounds in lettuce. HortScience 2007, 42, 71–76. [Google Scholar] [CrossRef]
- Llorach, R.; Martínez-Sánchez, A.; Tomás-Barberán, F.A.; Gil, M.I.; Ferreres, F. Characterisation of polyphenols and antioxidant properties of five lettuce varieties and escarole. Food Chem. 2008, 108, 1028–1038. [Google Scholar] [CrossRef]
- Gazula, A.; Kleinhenz, M.D.; Scheerens, J.C.; Ling, P.P. Anthocyanin levels in nine lettuce (Lactuca sativa) cultivars: Influence of planting date and relations among analytic, instrumented, and visual assessments of color. HortScience 2007, 42, 232–238. [Google Scholar] [CrossRef]
- Heimler, D.; Isolani, L.; Vignolini, P.; Tombelli, S.; Romani, A. Polyphenol content and antioxidative activity in some species of freshly consumed salads. J. Agric. Food Chem. 2007, 55, 1724–1729. [Google Scholar] [CrossRef] [PubMed]
- Rice-Evans, C.; Miller, N.; Paganga, G. Antioxidant properties of phenolic compounds. Trends Plant Sci. 1997, 2, 152–159. [Google Scholar] [CrossRef]
- Kumar, N.; Goel, N. Phenolic acids: Natural versatile molecules with promising therapeutic applications. Biotechnol. Rep. 2019, 24, e00370. [Google Scholar] [CrossRef]
- Kumar, N.; Pruthi, V.; Goel, N. Structural, thermal and quantum chemical studies of p-coumaric and caffeic acids. J. Mol. Struct. 2015, 1085, 242–248. [Google Scholar] [CrossRef]
- Tapas, A.R.; Sakarkar, D.M.; Kakde, R.B. Flavonoids as Nutraceuticals: A Review. Trop. J. Pharm. Res. 2008, 7, 1089–1099. [Google Scholar] [CrossRef]
- Cushnie, T.P.T.; Lamb, A.J. Antimicrobial activity of flavonoids. Int. J. Antimicrob. Agents 2005, 26, 343–356. [Google Scholar] [CrossRef] [PubMed]
- Rokkaku, T.; Kimura, R.; Ishikawa, C.; Yasumoto, T.; Senba, M.; Kanaya, F.; Mori, N. Anticancer Effects of Marine Carotenoids, Fucoxanthin and its Deacetylated Product, Fucoxanthinol, on Osteosarcoma. Int. J. Oncol. 2013, 43, 1176–1186. [Google Scholar] [CrossRef] [Green Version]
- Dwyer, J.H.; Navab, M.; Dwyer, K.M.; Hassan, K.; Sun, P.; Shircore, A.; Hama-Levy, S.; Hough, G.; Wang, X.; Drake, T.; et al. Oxygenated carotenoid lutein and progression of early atherosclerosis: The Los Angeles atherosclerosis study. Circulation 2001, 103, 2922–2927. [Google Scholar] [CrossRef] [Green Version]
- Ferruzzi, M.G.; Blakeslee, J. Digestion, absorption, and cancer preventative activity of dietary chlorophyll derivatives. Nutr. Res. 2007, 27, 1–12. [Google Scholar] [CrossRef]
- Pareek, S.; Sagar, N.A.; Sharma, S.; Kumar, V.; Agarwal, T.; González-Aguilar, G.A.; Yahia, E.M. Chlorophylls: Chemistry and biological functions. In Fruit and Vegetable Phytochemicals; Wiley-Blackwell: Hoboken, NJ, USA, 2017; pp. 269–284. [Google Scholar]
- Maiani, G.; Periago Castón, M.J.; Catasta, G.; Toti, E.; Cambrodón, I.G.; Bysted, A.; Granado-Lorencio, F.; Olmedilla-Alonso, B.; Knuthsen, P.; Valoti, M. Carotenoids: Actual knowledge on food sources, intakes, stability and bioavailability and their protective role in humans. Mol. Nutr. Food Res. 2009, 53, S194–S218. [Google Scholar] [CrossRef]
- Carnat, A.; Fraisse, D.; Lamaison, J.L.; Rock, E.; Michel, H.; Amouroux, P.; Remesy, C. Characterisation and variation of antioxidant micronutrients in lettuce (Lactuca sativa folium). J. Sci. Food Agric. 2004, 84, 2061–2069. [Google Scholar]
- Khachik, F. Distribution and metabolism of dietary carotenoids in humans as a criterion for development of nutritional supplements. Pure Appl. Chem. 2006, 78, 1551–1557. [Google Scholar] [CrossRef]
- Mou, B. Genetic variation of beta-carotene and lutein contents in lettuce. J. Am. Soc. Hortic. Sci. 2005, 130, 870–876. [Google Scholar] [CrossRef] [Green Version]
- Baslam, M.; Morales, F.; Garmendia, I.; Goicoechea, N. Nutritional quality of outer and inner leaves of green and red pigmented lettuces (Lactuca sativa L.) consumed as salads. Sci. Hortic. 2013, 151, 103–111. [Google Scholar] [CrossRef]
- Turkmen, N.; Poyrazoglu, E.S.; Sari, F.; Sedat Velioglu, Y. Effects of cooking methods on chlorophylls, pheophytins and colour of selected green vegetables. Int. J. Food Sci. Technol. 2006, 41, 281–288. [Google Scholar] [CrossRef]
- Ozgen, S.; Sekerci, S. Effect of leaf position on the distribution of phytochemicals and antioxidant capacity among green and red lettuce cultivars. Span. J. Agric. Res. 2011, 801–809. [Google Scholar] [CrossRef] [Green Version]
- Mou, B. Nutrient content of lettuce and its improvement. Curr. Nutr. Food Sci. 2009, 5, 242–248. [Google Scholar] [CrossRef]
- Survase, S.A.; Bajaj, I.B.; Singhal, R.S. Biotechnological production of vitamins. Food Technol. Biotechnol. 2006, 44, 381–396. [Google Scholar]
- Wang, C.; Riedl, K.M.; Schwartz, S.J. Fate of folates during vegetable juice processing—Deglutamylation and interconversion. Food Res. Int. 2013, 53, 440–448. [Google Scholar] [CrossRef]
- Dewhirst, R.A.; Fry, S.C. The oxidation of dehydroascorbic acid and 2,3-diketogulonate by distinct reactive oxygen species. Biochem. J. 2018, 475, 3451–3470. [Google Scholar] [CrossRef] [Green Version]
- Szeto, Y.T.; Tomlinson, B.; Benzie, I.F. Total antioxidant and ascorbic acid content of fresh fruits and vegetables: Implications for dietary planning and food preservation. Br. J. Nutr. 2002, 87, 55–59. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bahorun, T.; Luximon-Ramma, A.; Crozier, A.; Aruoma, O.I. Total phenol, flavonoid, proanthocyanidin and vitamin C levels and antioxidant activities of Mauritian vegetables. J. Sci. Food Agric. 2004, 84, 1553–1561. [Google Scholar] [CrossRef]
- Chun, J.; Lee, J.; Ye, L.; Exler, J.; Eitenmiller, R.R. Tocopherol and tocotrienol contents of raw and processed fruits and vegetables in the United States diet. J. Food Compos. Anal. 2006, 19, 196–204. [Google Scholar] [CrossRef]
- Szymańska, R.; Kruk, J. Tocopherol content and isomers’ composition in selected plant species. Plant Physiol. Biochem. 2008, 46, 29–33. [Google Scholar] [CrossRef] [PubMed]
- Byrdwell, W.C.; Kubzdela, N.; Goldschmidt, R. Changes in compositions of galactolipids, triacylglycerols, and tocopherols of lettuce varieties (Lactuca sativa L.) with type, age, and light source. J. Food Compos. Anal. 2021, 100, 103631. [Google Scholar] [CrossRef]
- Saini, R.K.; Shang, X.M.; Ko, E.Y.; Choi, J.H.; Keum, Y.-S. Stability of carotenoids and tocopherols in ready-to-eat baby-leaf lettuce and salad rocket during low-temperature storage. Int. J. Food Sci. Nutr. 2016, 67, 489–495. [Google Scholar] [CrossRef] [PubMed]
- Saini, R.; Manoj, P.; Shetty, N.; Srinivasan, K.; Giridhar, P. Dietary iron supplements and Moringa oleifera leaves influence the liver hepcidin messenger RNA expression and biochemical indices of iron status in rats. Nutr. Res. 2014, 34, 630–638. [Google Scholar] [CrossRef] [PubMed]
- Soetan, K.; Olaiya, C.; Oyewole, O. The importance of mineral elements for humans, domestic animals and plants—A review. Afr. J. Food Sci. 2010, 4, 200–222. [Google Scholar]
- Food, I.o.M.; Board, N. Dietary Reference Intakes for Water, Potassium, Sodium, Chloride, and Sulfate; National Academy Press: Washington, DC, USA, 2005. [Google Scholar]
- Santos, J.; Oliva-Teles, M.; Delerue-Matos, C.; Oliveira, M. Multi-elemental analysis of ready-to-eat “baby leaf” vegetables using microwave digestion and high-resolution continuum source atomic absorption spectrometry. Food Chem. 2014, 151, 311–316. [Google Scholar] [CrossRef]
- Sinha, K.; Khare, V. Review on: Antinutritional factors in vegetable crops. Pharma Innov. J. 2017, 6, 353–358. [Google Scholar]
- Greenwood, M.J.; Hunt, G.L.; McDowell, J.M. Migration and employment change: Empirical evidence on the spatial and temporal dimensions of the linkage. J. Reg. Sci. 1986, 26, 223–234. [Google Scholar] [CrossRef] [PubMed]
- Aletor, V. Allelochemicals in plant foods and feedingstuffs: 1. Nutritional, biochemical and physiopathological aspects in animal production. Vet. Hum. Toxicol. 1993, 35, 57–67. [Google Scholar] [PubMed]
- Fernando, R.; Pinto, M.; Pathmeswaran, A. Goitrogenic food and prevalence of goitre in Sri Lanka. Int. J. Intern. Med. 2012, 1, 17–20. [Google Scholar]
- Natesh, H.; Abbey, L.; Asiedu, S. An overview of nutritional and antinutritional factors in green leafy vegetables. Hortic. Int. J. 2017, 1, 00011. [Google Scholar] [CrossRef] [Green Version]
- Shahidi, F.; Peng, H. Bioaccessibility and bioavailability of phenolic compounds. J. Food Bioact. 2018, 4, 11–68. [Google Scholar] [CrossRef] [Green Version]
- Saura-Calixto, F.; Serrano, J.; Goñi, I. Intake and bioaccessibility of total polyphenols in a whole diet. Food Chem. 2007, 101, 492–501. [Google Scholar] [CrossRef] [Green Version]
- Gonthier, M.-P.; Verny, M.-A.; Besson, C.; Rémésy, C.; Scalbert, A. Chlorogenic acid bioavailability largely depends on its metabolism by the gut microflora in rats. J. Nutr. 2003, 133, 1853–1859. [Google Scholar] [CrossRef] [Green Version]
- Wang, S.-J.; Zeng, J.; Yang, B.-K.; Zhong, Y.-M. Bioavailability of caffeic acid in rats and its absorption properties in the Caco-2 cell model. Pharm. Biol. 2014, 52, 1150–1157. [Google Scholar] [CrossRef]
- Kishida, K.; Matsumoto, H. Urinary excretion rate and bioavailability of chlorogenic acid, caffeic acid, p-coumaric acid, and ferulic acid in non-fasted rats maintained under physiological conditions. Heliyon 2019, 5, e02708. [Google Scholar] [CrossRef] [Green Version]
- Bourne, L.C.; Rice-Evans, C. Bioavailability of ferulic acid. Biochem. Biophys. Res. Commun. 1998, 253, 222–227. [Google Scholar] [CrossRef]
- Dabeek, W.M.; Marra, M.V. Dietary quercetin and kaempferol: Bioavailability and potential cardiovascular-related bioactivity in humans. Nutrients 2019, 11, 2288. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Meyer, H.; Bolarinwa, A.; Wolfram, G.; Linseisen, J. Bioavailability of apigenin from apiin-rich parsley in humans. Ann. Nutr. Metab. 2006, 50, 167–172. [Google Scholar] [CrossRef] [PubMed]
- Lewandowska, U.; Szewczyk, K.; Hrabec, E.b.; Janecka, A.; Gorlach, S. Overview of metabolism and bioavailability enhancement of polyphenols. J. Agric. Food Chem. 2013, 61, 12183–12199. [Google Scholar] [CrossRef] [PubMed]
- Bohn, T. Dietary factors affecting polyphenol bioavailability. Nutr. Rev. 2014, 72, 429–452. [Google Scholar] [CrossRef] [PubMed]
- Ketnawa, S.; Reginio, F.C., Jr.; Thuengtung, S.; Ogawa, Y. Changes in bioactive compounds and antioxidant activity of plant-based foods by gastrointestinal digestion: A review. Crit. Rev. Food Sci. Nutr. 2021, 2021, 1–22. [Google Scholar] [CrossRef] [PubMed]
- Tarko, T.; Duda-Chodak, A.; Zajac, N. Digestion and absorption of phenolic compounds assessed by in vitro simulation methods. A review. Rocz. Państwowego Zakładu Hig. 2013, 64, 79–84. [Google Scholar]
- Karaś, M.; Jakubczyk, A.; Szymanowska, U.; Złotek, U.; Zielińska, E. Digestion and bioavailability of bioactive phytochemicals. Int. J. Food Sci. Technol. 2017, 52, 291–305. [Google Scholar] [CrossRef]
- Fernandez-Garcia, E.; Carvajal-Lerida, I.; Jaren-Galan, M.; Garrido-Fernandez, J.; Perez-Galvez, A.; Hornero-Mendez, D. Carotenoids bioavailability from foods: From plant pigments to efficient biological activities. Food Res. Int. 2012, 46, 438–450. [Google Scholar] [CrossRef]
- Hof, K.H.v.h.; West, C.E.; Westrate, J.A.; Hautvast, J.G.A.J. Dietary factors that affect the bioavailability of carotenoids. J. Nutr. 2000, 130, 503–506. [Google Scholar]
- Malo, C.; Wilson, J.X. Glucose Modulates Vitamin C Transport in Adult Human Small Intestinal Brush Border Membrane Vesicles. J. Nutr. 2000, 130, 63–69. [Google Scholar] [CrossRef] [Green Version]
- Fujita, I.; Akagi, Y.; Hirano, J.; Nakanishi, T.; Itoh, N.; Muto, N.; Tanaka, K. Distinct mechanisms of transport of ascorbic acid and dehydroascorbic acid in intestinal epithelial cells (IEC-6). Res. Commun. Mol. Pathol. 2000, 107, 219–231. [Google Scholar]
- Ti, H.; Zhang, R.; Li, Q.; Wei, Z.; Zhang, M. Effects of cooking and in vitro digestion of rice on phenolic profiles and antioxidant activity. Food Res. Int. 2015, 76, 813–820. [Google Scholar] [CrossRef] [PubMed]
- Burtch, H.; Ferruzzi, M.; Kay, C.; Lila, M.; Iorrizo, M.; Mengist, M. Adaptation of an in vitro digestion model for high throughput phenolic bioaccessibility phenotyping within cultivated (highbush) blueberry varieties. Curr. Dev. Nutr. 2019, 3, nzz031. [Google Scholar] [CrossRef] [Green Version]
- Dutta, T.; Paul, A.; Majumder, M.; Sultan, R.A.; Emran, T.B. Pharmacological evidence for the use of Cissus assamica as a medicinal plant in the management of pain and pyrexia. Biochem. Biophys. Rep. 2020, 21, 100715. [Google Scholar] [CrossRef] [PubMed]
- Hedges, L.; Lister, C. Nutritional attributes of salad vegetables. Crop Food Res. Confid. Rep. 2005, 9, 1–29. [Google Scholar]
- Altunkaya, A.; Becker, E.M.; Gökmen, V.; Skibsted, L.H. Antioxidant activity of lettuce extract (Lactuca sativa) and synergism with added phenolic antioxidants. Food Chem. 2009, 115, 163–168. [Google Scholar] [CrossRef]
- Gan, Y.; Azrina, A. Antioxidant properties of selected varieties of lettuce (Lactuca sativa L.) commercially available in Malaysia. Int. Food Res. J. 2016, 23, 2357–2362. [Google Scholar]
- Reis, J.F.; Monteiro, V.V.S.; de Souza Gomes, R.; do Carmo, M.M.; da Costa, G.V.; Ribera, P.C.; Monteiro, M.C. Action mechanism and cardiovascular effect of anthocyanins: A systematic review of animal and human studies. J. Transl. Med. 2016, 14, 315. [Google Scholar] [CrossRef] [Green Version]
- Praveen, P.A.; Roy, A.; Prabhakaran, D. Cardiovascular disease risk factors: A childhood perspective. Indian J. Pediatrics 2013, 80, 3–12. [Google Scholar] [CrossRef]
- Nicolle, C.; Cardinault, N.; Gueux, E.; Jaffrelo, L.; Rock, E.; Mazur, A.; Amouroux, P.; Rémésy, C. Health effect of vegetable-based diet: Lettuce consumption improves cholesterol metabolism and antioxidant status in the rat. Clin. Nutr. 2004, 23, 605–614. [Google Scholar] [CrossRef]
- Nishimura, N.; Taniguchi, Y.; Kiriyama, S. Plasma cholesterol-lowering effect on rats of dietary fiber extracted from immature plants. Biosci. Biotechnol. Biochem. 2000, 64, 2543–2551. [Google Scholar] [CrossRef] [PubMed]
- Stedronsky, E.R. Interaction of bile acids and cholesterol with non-systemic agents having hypocholesterolemic properties. Biochim. Et Biophys. Acta (BBA)-Lipids Lipid Metab. 1994, 1210, 255–287. [Google Scholar] [CrossRef]
- Proteggente, A.R.; Pannala, A.S.; Paganga, G.; Buren, L.v.; Wagner, E.; Wiseman, S.; Put, F.v.d.; Dacombe, C.; Rice-Evans, C.A. The antioxidant activity of regularly consumed fruit and vegetables reflects their phenolic and vitamin C composition. Free Radic. Res. 2002, 36, 217–233. [Google Scholar] [CrossRef] [PubMed]
- Aviram, M.; Dornfeld, L.; Rosenblat, M.; Volkova, N.; Kaplan, M.; Coleman, R.; Hayek, T.; Presser, D.; Fuhrman, B. Pomegranate juice consumption reduces oxidative stress, atherogenic modifications to LDL, and platelet aggregation: Studies in humans and in atherosclerotic apolipoprotein E–deficient mice. Am. J. Clin. Nutr. 2000, 71, 1062–1076. [Google Scholar] [CrossRef] [PubMed]
- García-Lafuente, A.; Guillamón, E.; Villares, A.; Rostagno, M.A.; Martínez, J.A. Flavonoids as anti-inflammatory agents: Implications in cancer and cardiovascular disease. Inflamm. Res. 2009, 58, 537–552. [Google Scholar] [CrossRef]
- Sularz, O.; Koronowicz, A.; Smoleń, S.; Kowalska, I.; Skoczylas, Ł.; Liszka-Skoczylas, M.; Tabaszewska, M.; Pitala, J. Anti-and pro-oxidant potential of lettuce (Lactuca sativa L.) biofortified with iodine by KIO 3, 5-iodo-and 3, 5-diiodosalicylic acid in human gastrointestinal cancer cell lines. RSC Adv. 2021, 11, 27547–27560. [Google Scholar] [CrossRef]
- Fernandez, E.; La Vecchia, C.; D’Avanzo, B.; Negri, E.; Franceschi, S. Risk factors for colorectal cancer in subjects with family history of the disease. Br. J. Cancer 1997, 75, 1381–1384. [Google Scholar] [CrossRef] [Green Version]
- Brennan, P.; Fortes, C.; Butler, J.; Agudo, A.; Benhamou, S.; Darby, S.; Gerken, M.; Jöckel, K.-H.; Kreuzer, M.; Mallone, S. A multicenter case–control study of diet and lung cancer among non-smokers. Cancer Causes Control 2000, 11, 49–58. [Google Scholar] [CrossRef]
- Donaldson, M.S. Nutrition and cancer: A review of the evidence for an anti-cancer diet. Nutr. J. 2004, 3, 19. [Google Scholar] [CrossRef] [Green Version]
- Tareq, A.M.; Farhad, S.; Uddin, A.N.; Hoque, M.; Nasrin, M.S.; Uddin, M.M.R.; Hasan, M.; Sultana, A.; Munira, M.S.; Lyzu, C.; et al. Chemical profiles, pharmacological properties, and in silico studies provide new insights on Cycas pectinata. Heliyon 2020, 6, e04061. [Google Scholar] [CrossRef]
- Martins, S.; Mussatto, S.I.; Martínez-Avila, G.; Montañez-Saenz, J.; Aguilar, C.N.; Teixeira, J.A. Bioactive phenolic compounds: Production and extraction by solid-state fermentation. A review. Biotechnol. Adv. 2011, 29, 365–373. [Google Scholar] [CrossRef] [Green Version]
- Rauf, A.; Shariati, M.A.; Imran, M.; Bashir, K.; Khan, S.A.; Mitra, S.; Emran, T.B.; Badalova, K.; Uddin, M.; Mubarak, M.S. Comprehensive review on naringenin and naringin polyphenols as a potent anticancer agent. Environ. Sci. Pollut. Res. 2022, 29, 31025–31041. [Google Scholar] [CrossRef] [PubMed]
- Rauf, A.; Abu-Izneid, T.; Khalil, A.A.; Imran, M.; Shah, Z.A.; Emran, T.B.; Mitra, S.; Khan, Z.; Alhumaydhi, F.A.; Aljohani, A.S. Berberine as a potential anticancer agent: A comprehensive review. Molecules 2021, 26, 7368. [Google Scholar] [CrossRef] [PubMed]
- Terry, L. Health-Promoting Properties of Fruits and Vegetables; CABI: Oxfordshore, UK, 2011. [Google Scholar]
- Thomford, N.E.; Mkhize, B.; Dzobo, K.; Mpye, K.; Rowe, A.; Parker, M.I.; Wonkam, A.; Skelton, M.; September, A.V.; Dandara, C. African lettuce (Launaea taraxacifolia) displays possible anticancer effects and herb–drug interaction potential by CYP1A2, CYP2C9, and CYP2C19 inhibition. Omics J. Integr. Biol. 2016, 20, 528–537. [Google Scholar] [CrossRef]
- Zhou, W.; Liang, X.; Dai, P.; Chen, Y.; Zhang, Y.; Zhang, M.; Lu, L.; Jin, C.; Lin, X. Alteration of phenolic composition in lettuce (Lactuca sativa L.) by reducing nitrogen supply enhances its anti-proliferative effects on colorectal cancer cells. Int. J. Mol. Sci. 2019, 20, 4205. [Google Scholar] [CrossRef] [Green Version]
- Imran, M.; Aslam Gondal, T.; Atif, M.; Shahbaz, M.; Batool Qaisarani, T.; Hanif Mughal, M.; Salehi, B.; Martorell, M.; Sharifi-Rad, J. Apigenin as an anticancer agent. Phytother. Res. 2020, 34, 1812–1828. [Google Scholar] [CrossRef] [PubMed]
- Kabir, M.S.H.; Hossain, M.M.; Kabir, M.I.; Rahman, M.M.; Hasanat, A.; Emran, T.B.; Rahman, M.A. Phytochemical screening, Antioxidant, Thrombolytic, alpha-amylase inhibition and cytotoxic activities of ethanol extract of Steudnera colocasiifolia K. Koch leaves. J. Young Pharm. 2016, 8, 391. [Google Scholar] [CrossRef] [Green Version]
- Rahman, M.M.; Islam, M.R.; Shohag, S.; Hossain, M.E.; Rahaman, M.S.; Islam, F.; Ahmed, M.; Mitra, S.; Khandaker, M.U.; Idris, A.M. The multifunctional role of herbal products in the management of diabetes and obesity: A comprehensive review. Molecules 2022, 27, 1713. [Google Scholar] [CrossRef]
- Gopal, S.S.; Lakshmi, M.J.; Sharavana, G.; Sathaiah, G.; Sreerama, Y.N.; Baskaran, V. Lactucaxanthin—A potential anti-diabetic carotenoid from lettuce (Lactuca sativa) inhibits α-amylase and α-glucosidase activity in vitro and in diabetic rats. Food Funct. 2017, 8, 1124–1131. [Google Scholar] [CrossRef]
- Dembinska-Kiec, A.; Mykkänen, O.; Kiec-Wilk, B.; Mykkänen, H. Antioxidant phytochemicals against type 2 diabetes. Br. J. Nutr. 2008, 99, ES109–ES117. [Google Scholar] [CrossRef] [Green Version]
- Cheng; Pogrebnyak, N.; Kuhn, P.; Poulev, A.; Waterman, C.; Rojas-Silva, P.; Johnson, W.D.; Raskin, I. Polyphenol-rich Rutgers Scarlet Lettuce improves glucose metabolism and liver lipid accumulation in diet-induced obese C57BL/6 mice. Nutrition 2014, 30, S52–S58. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Shah, A.; Mudassar, S. Nutraceutical-Based Pharmacological Intervention in the Management of Liver Diseases. In Nutraceuticals and Natural Product Derivatives: Disease Prevention and Drug Discovery; John Wiley and Sons, Inc.: Hoboken, NJ, USA, 2019; pp. 375–394. [Google Scholar]
- Guo, H.; Xia, M.; Zou, T.; Ling, W.; Zhong, R.; Zhang, W. Cyanidin 3-glucoside attenuates obesity-associated insulin resistance and hepatic steatosis in high-fat diet-fed and db/db mice via the transcription factor FoxO1. J. Nutr. Biochem. 2012, 23, 349–360. [Google Scholar] [CrossRef] [PubMed]
- Nie, C.; Zhu, P.; Ma, S.; Wang, M.; Hu, Y. Purification, characterization and immunomodulatory activity of polysaccharides from stem lettuce. Carbohydr. Polym. 2018, 188, 236–242. [Google Scholar] [CrossRef]
- Seo, H.-J.; Jeong, J.B. Immune-Enhancing Effects of Green Lettuce (Lactuca sativa L.) Extracts through the TLR4-MAPK/NF-κB Signaling Pathways in RAW264. 7 Macrophage Cells. Korean J. Plant Resour. 2020, 33, 183–193. [Google Scholar]
- Mitra, S.; Paul, S.; Roy, S.; Sutradhar, H.; Bin Emran, T.; Nainu, F.; Khandaker, M.U.; Almalki, M.; Wilairatana, P.; Mubarak, M.S. Exploring the Immune-Boosting Functions of Vitamins and Minerals as Nutritional Food Bioactive Compounds: A Comprehensive Review. Molecules 2022, 27, 555. [Google Scholar] [CrossRef] [PubMed]
- Awan, A.F.; Akhtar, M.S.; Anjum, I.; Mushtaq, M.N.; Fatima, A.; Mannan, A.; Ali, I. Anti-oxidant and hepatoprotective effects of Lactuca serriola and its phytochemical screening by HPLC and FTIR analysis. Pak. J. Pharm. Sci. 2020, 33, 2823–2830. [Google Scholar] [PubMed]
- James, A.; Olasore, H.; Babalola, O.; Ikujuni, A.; Magbagbeola, O. Hepatoprotective Potential of Ethanol Leaf Extract of Launaea taraxacifolia Willd.(Asteraceae) on Carbon Tetrachloride-induced Liver Toxicity in Sprague-Dawley Rats. Univ. Lagos J. Basic Med Sci. 2021, 7, 1–2. [Google Scholar]
- Hefnawy, H.T.M.; Ramadan, M.F. Protective effects of Lactuca sativa ethanolic extract on carbon tetrachloride induced oxidative damage in rats. Asian Pac. J. Trop. Dis. 2013, 3, 277–285. [Google Scholar] [CrossRef]
- Ghorbani, A.; Sadeghnia, H.R.; Asadpour, E. Mechanism of protective effect of lettuce against glucose/serum deprivation-induced neurotoxicity. Nutr. Neurosci. 2015, 18, 103–109. [Google Scholar] [CrossRef]
- Im, S.-E.; Yoon, H.; Nam, T.-G.; Heo, H.J.; Lee, C.Y.; Kim, D.-O. Antineurodegenerative effect of phenolic extracts and caffeic acid derivatives in romaine lettuce on neuron-like PC-12 cells. J. Med. Food 2010, 13, 779–784. [Google Scholar] [CrossRef]
- Zhan, H.; Li, S.; Sun, J.; Liu, R.; Yan, F.; Niu, B.; Zhang, H.; Wang, X. Lettuce glycoside B ameliorates cerebral ischemia reperfusion injury by increasing nerve growth factor and neurotrophin-3 expression of cerebral cortex in rats. Indian J. Pharmacol. 2014, 46, 63. [Google Scholar] [PubMed] [Green Version]
- Altunkaya, A.; Gökmen, V. Effect of various inhibitors on enzymatic browning, antioxidant activity and total phenol content of fresh lettuce (Lactuca sativa). Food Chem. 2008, 107, 1173–1179. [Google Scholar] [CrossRef]
- Altunkaya, A.; Gökmen, V.; Skibsted, L.H. pH dependent antioxidant activity of lettuce (L. sativa) and synergism with added phenolic antioxidants. Food Chem. 2016, 190, 25–32. [Google Scholar] [CrossRef] [PubMed]
- Palit, S.; Bhuiyan, R.H.; Aklima, J.; Emran, T.B.; Dash, R. A study of the prevalence of thalassemia and its correlation with liver function test in different age and sex group in the Chittagong district of Bangladesh. J. Basic Clin. Pharm. 2012, 3, 352. [Google Scholar]
- Rizvi, S.I.; Maurya, P.K. Alterations in antioxidant enzymes during aging in humans. Mol. Biotechnol. 2007, 37, 58–61. [Google Scholar] [CrossRef] [PubMed]
- Chaudhary, S. Vitamin K—The ignorant nutrient. Int. J. Clin. Biomed. Res. 2015, 2, 24–25. [Google Scholar]
- Santamaria, P. Nitrate in vegetables: Toxicity, content, intake and EC regulation. J. Sci. Food Agric. 2006, 86, 10–17. [Google Scholar] [CrossRef]
- Ysart, G.; Clifford, R.; Harrison, N. Monitoring for nitrate in UK-grown lettuce and spinach. Food Addit. Contam. 1999, 16, 301–306. [Google Scholar] [CrossRef]
- Regulation. 2006/EC (2006) Off. J. Eur. Union 1881, 364, 5–24. [Google Scholar]
- Iammarino, M.; Taranto, A.; Cristino, M. Monitoring of nitrites and nitrates levels in leafy vegetables (Spinach And Lettuce): A Contribution To Risk Assessment. J. Sci. Food Agric. 2014, 94, 773–778. [Google Scholar] [CrossRef]
- Negri, S.; Commisso, M.; Avesani, L.; Guzzo, F. The case of tryptamine and serotonin in plants: A mysterious precursor for an illustrious metabolite. J. Exp. Bot. 2021, 72, 5336–5355. [Google Scholar] [CrossRef] [PubMed]
- Besharat, S.; Besharat, M.; Jabbari, A. Wild lettuce (Lactuca virosa) toxicity. BMJ Case Rep. 2009, 2009, bcr0620080134. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Llorach, R.; Tomás-Barberán, F.A.; Ferreres, F. Lettuce and chicory byproducts as a source of antioxidant phenolic extracts. J. Agric. Food Chem. 2004, 52, 5109–5116. [Google Scholar] [CrossRef] [PubMed]
- Chemat, F.; Rombaut, N.; Meullemiestre, A.; Turk, M.; Perino, S.; Fabiano-Tixier, A.-S.; Abert-Vian, M. Review of green food processing techniques. Preservation, transformation, and extraction. Innov. Food Sci. Emerg. Technol. 2017, 41, 357–377. [Google Scholar] [CrossRef]
- Dziki, D.; Różyło, R.; Gawlik-Dziki, U.; Świeca, M. Current trends in the enhancement of antioxidant activity of wheat bread by the addition of plant materials rich in phenolic compounds. Trends Food Sci. Technol. 2014, 40, 48–61. [Google Scholar] [CrossRef]
- Plazzotta, S.; Sillani, S.; Manzocco, L. Exploitation of lettuce waste flour to increase bread functionality: Effect on physical, nutritional, sensory properties and on consumer response. Int. J. Food Sci. Technol. 2018, 53, 2290–2297. [Google Scholar] [CrossRef] [Green Version]




{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Phytochemicals | Subgroups | Specific Compounds | Biological Activities | Reference |
|---|---|---|---|---|
| Phenolic compounds | Phenolic acids | Caffeic acid, Chlorogenic acid, and their derivatives | Antidiabetic, antimicrobial, anti-inflammatory, skin care products, food preservatives | [37,38] |
| Flavonoids | Quercetin, Isorhamnetin, Kaempferol, Anthocyanins | Hepatoprotective, antibacterial, anti-inflammatory, anticancer, antiviral | [39,40] | |
| Carotenoids | Carotenes | α-carotene, β-carotene, Lycopene | Cardiovascular protective, anti-cancer, anti-obesity, pigment, antiproliferative | [41,42] |
| Xanthophylls | Lutein, Neoxanthin, Lactucaxanthin, Violaxanthin, Zeaxanthin | |||
| Chlorophyll | Chlorophyll A, Chlorophyll B | Antioxidant, anti-cancer, stimulating immune system, pigment, normalize blood pressure | [43,44] |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Shi, M.; Gu, J.; Wu, H.; Rauf, A.; Emran, T.B.; Khan, Z.; Mitra, S.; Aljohani, A.S.M.; Alhumaydhi, F.A.; Al-Awthan, Y.S.; et al. Phytochemicals, Nutrition, Metabolism, Bioavailability, and Health Benefits in Lettuce—A Comprehensive Review. Antioxidants 2022, 11, 1158. https://doi.org/10.3390/antiox11061158
Shi M, Gu J, Wu H, Rauf A, Emran TB, Khan Z, Mitra S, Aljohani ASM, Alhumaydhi FA, Al-Awthan YS, et al. Phytochemicals, Nutrition, Metabolism, Bioavailability, and Health Benefits in Lettuce—A Comprehensive Review. Antioxidants. 2022; 11(6):1158. https://doi.org/10.3390/antiox11061158
Chicago/Turabian StyleShi, Min, Jingyu Gu, Hanjing Wu, Abdur Rauf, Talha Bin Emran, Zidan Khan, Saikat Mitra, Abdullah S. M. Aljohani, Fahad A. Alhumaydhi, Yahya S. Al-Awthan, and et al. 2022. "Phytochemicals, Nutrition, Metabolism, Bioavailability, and Health Benefits in Lettuce—A Comprehensive Review" Antioxidants 11, no. 6: 1158. https://doi.org/10.3390/antiox11061158
APA StyleShi, M., Gu, J., Wu, H., Rauf, A., Emran, T. B., Khan, Z., Mitra, S., Aljohani, A. S. M., Alhumaydhi, F. A., Al-Awthan, Y. S., Bahattab, O., Thiruvengadam, M., & Suleria, H. A. R. (2022). Phytochemicals, Nutrition, Metabolism, Bioavailability, and Health Benefits in Lettuce—A Comprehensive Review. Antioxidants, 11(6), 1158. https://doi.org/10.3390/antiox11061158