
Tomato Allergy: The Characterization of the Selected Allergens and Antioxidants of Tomato (Solanum lycopersicum)—A Review




Abstract
:1. Introduction
2. Tomato Antioxidants
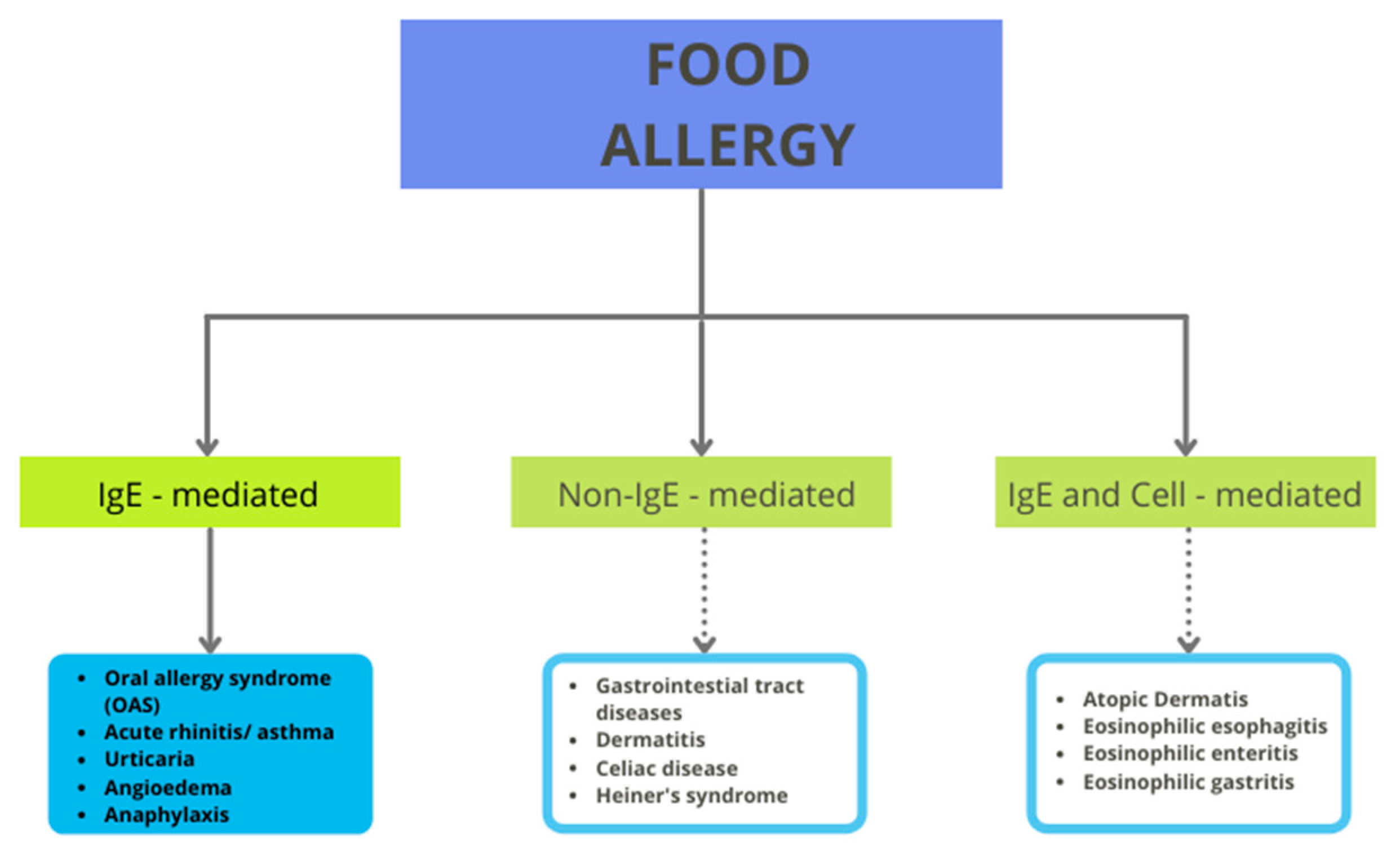
3. Tomato Allergy
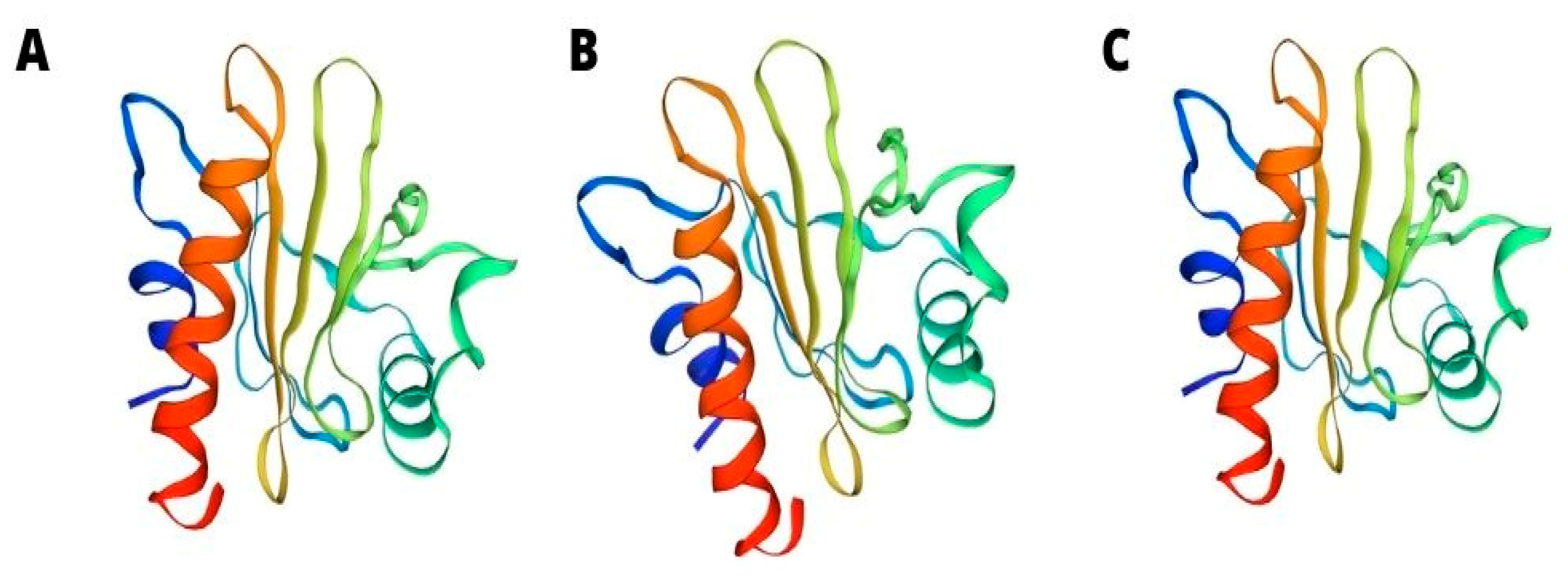
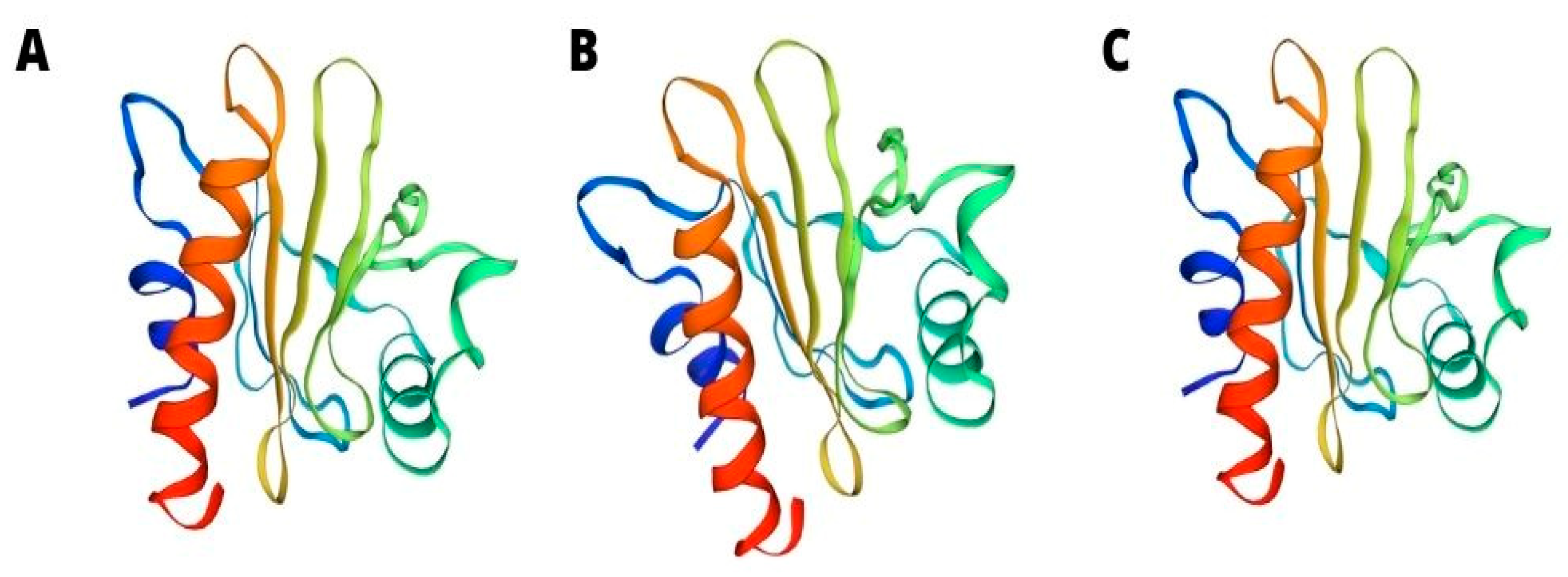
4. Profilins—Sola l 1
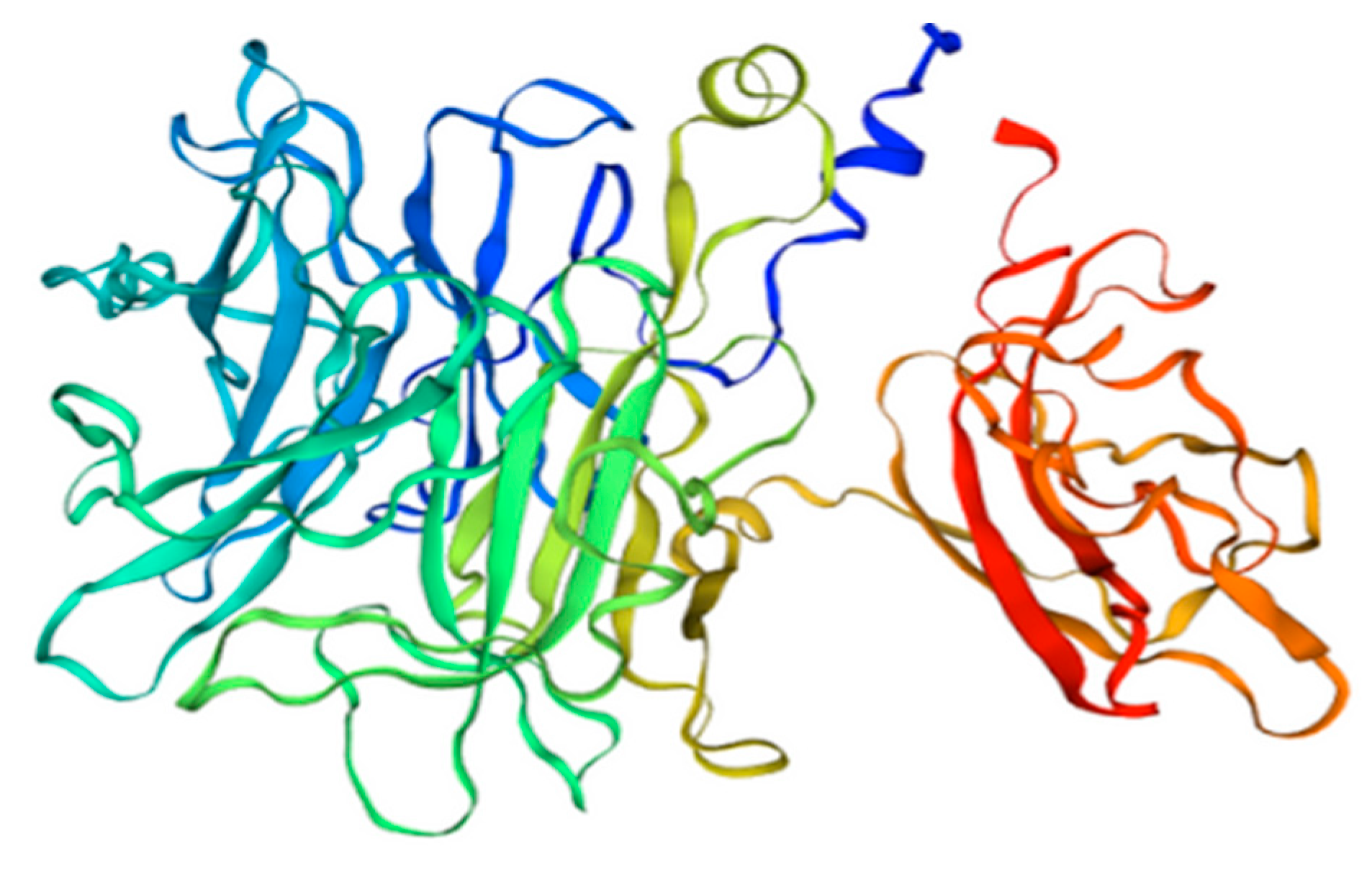
5. β-Fructofuranosidase—Sola l 2
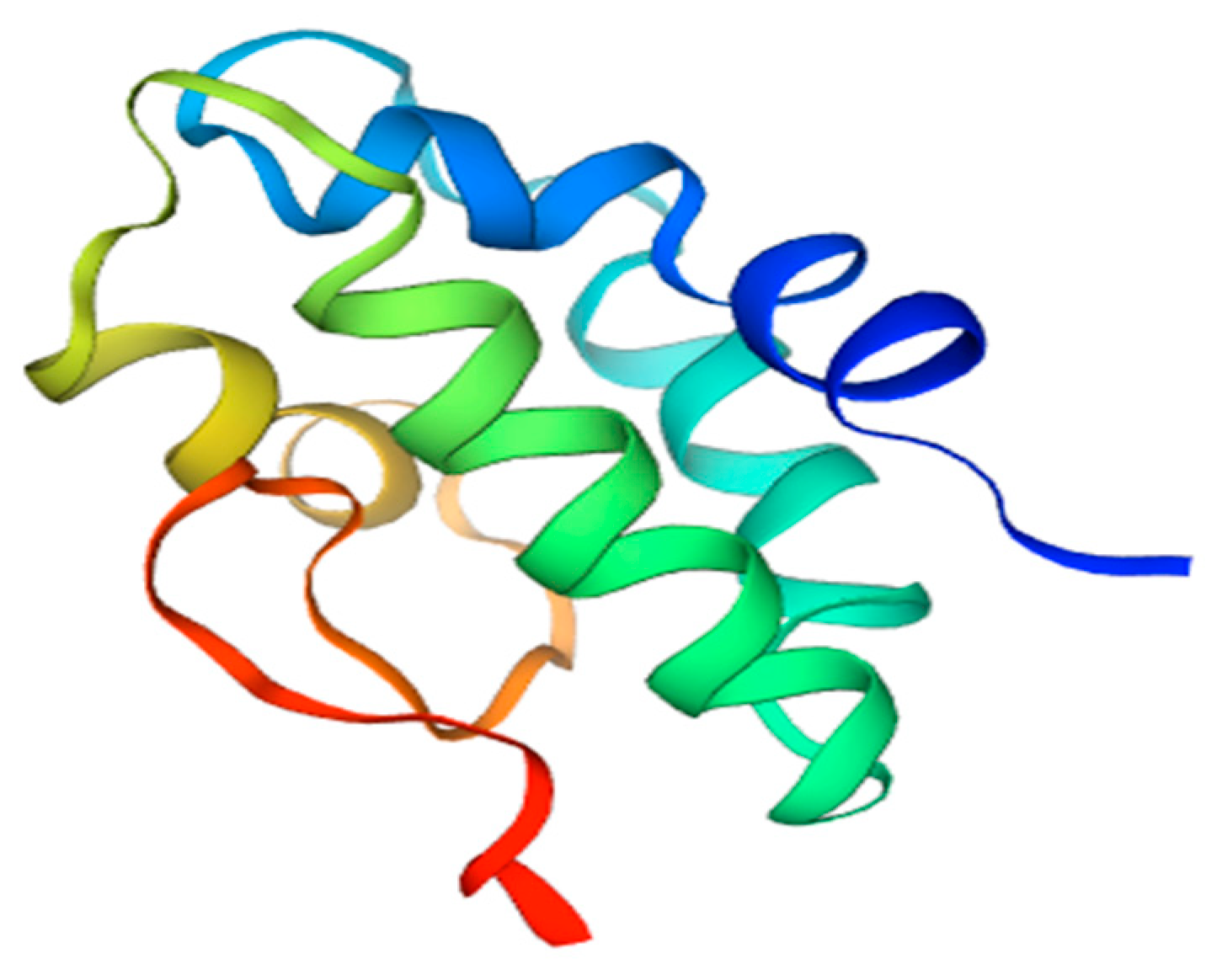
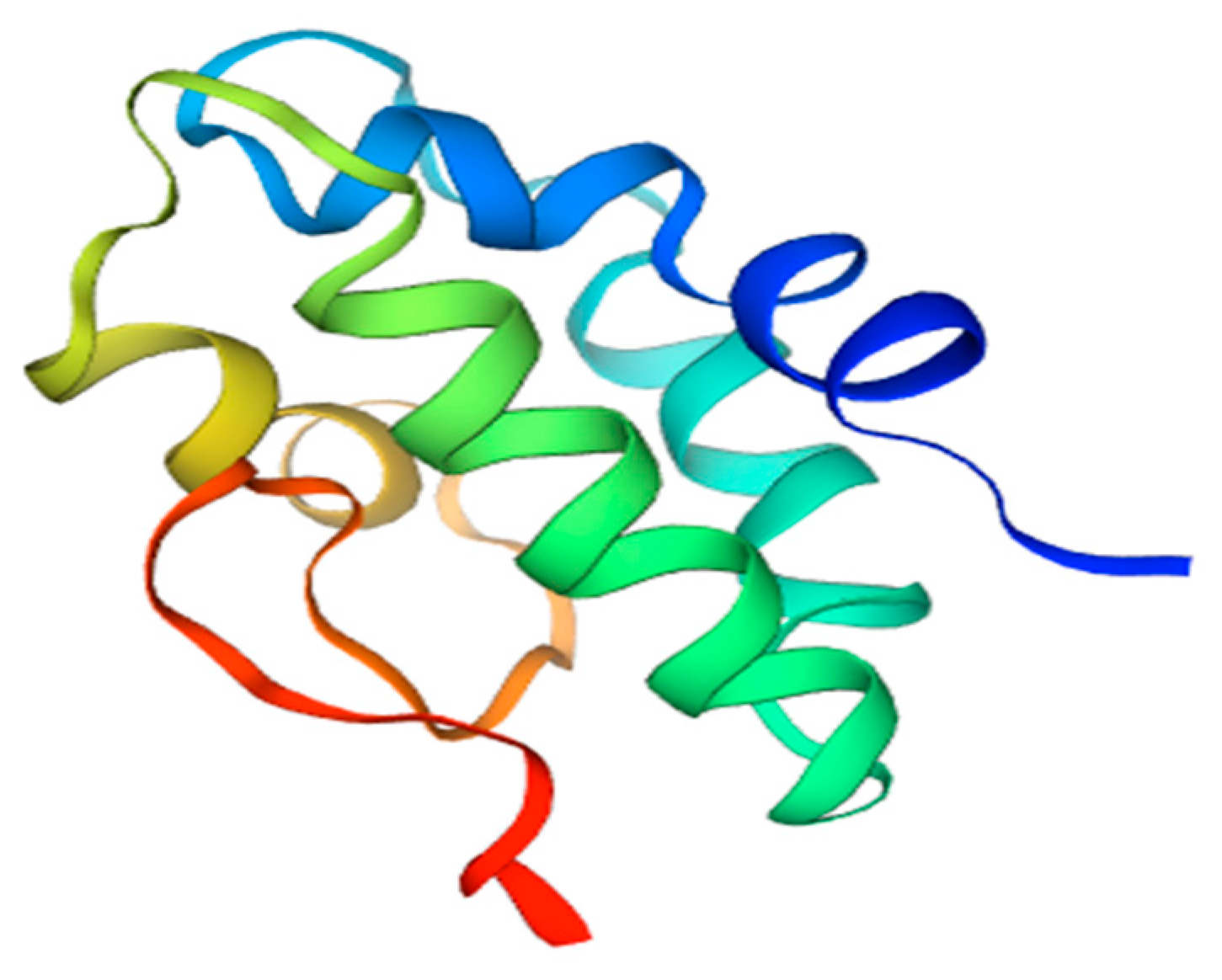
6. Lipid Transfer Protein—Sola l 3
7. The Pathogenesis-Related Proteins—Sola l 4
8. Cross-Reactivity
9. Transgenic Tomatoes
10. The Impact of Environmental Conditions and Processing Techniques on Tomato Allergens
10.1. Environmental Conditions
10.2. Food Processing
11. Current State of the Art
12. Concluding Remarks
Author Contributions
Funding
Conflicts of Interest
References
- Peralta, I.E.; Knapp, S.; Spooner, D.M. New Species of Wild Tomatoes (Solanum Section Lycopersicon: Solanaceae) from Northern Peru. Syst. Bot. 2009, 30, 424–434. [Google Scholar] [CrossRef]
- Sahlin, E.; Savage, G.; Lister, C. Investigation of the Antioxidant Properties of Tomatoes after Processing. J. Food Compos. Anal. 2004, 17, 635–647. [Google Scholar] [CrossRef]
- Tilahun, S.; Park, D.; Seo, M.; Jeong, C. Review on Factors Affecting the Quality and Antioxidant Properties of Tomatoes. AFRICAN J. Biotechnol. 2017, 16, 1678–1687. [Google Scholar] [CrossRef] [Green Version]
- Sainju, U.; Dris, R.; Singh, B. Mineral Nutrition of Tomato. Food Agric. Environ. 2003, 1, 176–183. [Google Scholar]
- Vashisth, A.; Nagarajan, S. Effect on Germination and Early Growth Characteristics in Sunflower (Helianthus annus) Seeds Exposed to Static Magnetic Field. J. Plant Physiol. 2009, 167, 149–156. [Google Scholar] [CrossRef]
- Costa, M.; Heuvelink, E. The Global Tomato Industry; CABI: Boston, MA, USA, 2018; pp. 1–26. [Google Scholar] [CrossRef]
- Onopiuk, A.; Półtorak, A.; Wojtasik-Kalinowska, I.; Szpicer, A.; Marcinkowska-Lesiak, M.; Pogorzelski, G.; Wierzbicka, A. Impact of the Storage Atmosphere Enriched with Ozone on the Quality of Lycopersicon esculentum Tomatoes. J. Food Process. Preserv. 2019, 43, e14252. [Google Scholar] [CrossRef]
- Kucuk, O.; Sarkar, F.; Sakr, W.; Djuric, Z.; Pollak, M.; Khachik, F.; Li, Y.W.; Banerjee, M.; Grignon, D.; Bertram, J.S.; et al. Phase II Randomized Clinical Trial of Lycopene Supplementation before Radical Prostatectomy. Cancer Epidemiol. Biomark. Prev. 2001, 10, 861–868. [Google Scholar]
- Nishino, H.; Murakoshi, M.; Ii, T.; Takemura, M.; Kuchide, M.; Kanazawa, M.; Yang Mou, X.; Wada, S.; Masuda, M.; Ohsaka, Y.; et al. Carotenoids in Cancer Chemoprevention. Cancer Metastasis Rev. 2002, 21, 257–264. [Google Scholar] [CrossRef]
- Riccioni, G. Carotenoids and Cardiovascular Disease. Curr. Atheroscler. Rep. 2009, 11, 434–439. [Google Scholar] [CrossRef] [PubMed]
- Hossin, I.; Talukdar, G.; Roy, N.; Shaha, R. Anti-Allergic Compounds from Red Tomato Peel. J. Adv. Lab. Res. Biol. 2012, 3, 181–189. [Google Scholar]
- Capanoglu, E.; Beekwilder, J.; Boyacioglu, D.; De Vos, R.C.H.; Hall, R.D. The Effect of Industrial Food Processing on Potentially Health-Beneficial Tomato Antioxidants. Crit. Rev. Food Sci. Nutr. 2010, 50, 919–930. [Google Scholar] [CrossRef]
- Chen, R.Y.; Wu, J.J.; Tsai, M.J.; Liu, M.S. Effects of Storage and Thermal Treatment on the Antioxidant Activity of Tomato Fruits. Taiwan J. Agric. Chem. Food Sci. 2000, 38, 353–360. [Google Scholar]
- Graziani, G.; Pernice, R.; Lanzuise, S.; Vitaglione, P.; Anese, M.; Fogliano, V. Effect of Peeling and Heating on Carotenoid Content and Antioxidant Activity of Tomato and Tomato-Virgin Olive Oil Systems. Eur. Food Res. Technol. 2003, 216, 116–121. [Google Scholar] [CrossRef]
- Chang, C.-H.; Lin, H.-Y.; Chang, C.-Y.; Liu, Y.-C. Comparisons on the Antioxidant Properties of Fresh, Freeze-Dried and Hot-Air-Dried Tomatoes. J. Food Eng. 2006, 77, 478–485. [Google Scholar] [CrossRef]
- Alda, L.; Gogoaşă, I.; Bordean, D.-M.; Gergen, I.; Alda, S.; Moldovan, C.; Niţă, L. Lycopene Content of Tomatoes and Tomato Products. J. Agroaliment. Process. Technol. 2009, 15, 540–542. [Google Scholar]
- Yamamoto, T.; Yoshimura, M.; Yamaguchi, F.; Kouchi, T.; Tsuji, R.; Saito, M.; Obata, A.; Kikuchi, M. Anti-Allergic Activity of Naringenin Chalcone from a Tomato Skin Extract. Biosci. Biotechnol. Biochem. 2004, 68, 1706–1711. [Google Scholar] [CrossRef] [Green Version]
- Gates, K.W. Food Allergen Testing: Molecular, Immunochemical, and Chromatographic Techniques, by George Siragakis and Dimosthenis Kizis. J. Aquat. Food Prod. Technol. 2015, 24, 632–635. [Google Scholar] [CrossRef]
- Petersen, A.; Vieths, S.; Aulepp, H.; Schlaak, M.; Becker, W.-M. Ubiquitous Structures Responsible for IgE Cross-Reactivity between Tomato Fruit and Grass Pollen Allergens. J. Allergy Clin. Immunol. 1996, 98, 805–815. [Google Scholar] [CrossRef]
- Ortolani, C.; Ispano, M.; Pastorello, E.A.; Ansaloni, R.; Magri, G.C. Comparison of Results of Skin Prick Tests (with Fresh Foods and Commercial Food Extracts) and RAST in 100 Patients with Oral Allergy Syndrome. J. Allergy Clin. Immunol. 1989, 83, 683–690. [Google Scholar] [CrossRef]
- Burney, P.G.J.; Potts, J.; Kummeling, I.; Mills, E.N.C.; Clausen, M.; Dubakiene, R.; Barreales, L.; Fernandez-Perez, C.; Fernandez-Rivas, M.; Le, T.-M.; et al. The Prevalence and Distribution of Food Sensitization in European Adults. Allergy 2014, 69, 365–371. [Google Scholar] [CrossRef] [Green Version]
- Ballmer-Weber, B.; Hoffmann-Sommergruber, K. Molecular Diagnosis of Fruit and Vegetable Allergy. Curr. Opin. Allergy Clin. Immunol. 2011, 11, 229–235. [Google Scholar] [CrossRef] [PubMed]
- Geroldinger-Simic, M.; Zelniker, T.; Aberer, W.; Ebner, C.; Egger, C.; Greiderer, A.; Prem, N.; Lidholm, J.; Ballmer-Weber, B.; Vieths, S.; et al. Birch Pollen-Related Food Allergy: Clinical Aspects and the Role of Allergen-Specific IgE and IgG(4) Antibodies. J. Allergy Clin. Immunol. 2011, 127, 616–622.e1. [Google Scholar] [CrossRef] [PubMed]
- Bencivenni, M.; Faccini, A.; Bottesini, C.; Rao, R.; Detoraki, A.; Ridolo, E.; Marone, G.; Dall’Aglio, P.; Dossena, A.; Marchelli, R.; et al. Assessing Allergenicity of Different Tomato Ecotypes by Using Pooled Sera of Allergic Subjects: Identification of the Main Allergens. Eur. Food Res. Technol. 2012, 234, 405–414. [Google Scholar] [CrossRef]
- López-Matas, M.A.; Larramendi, C.H.; Huertas, A.J.; Ferrer, A.; Moya, R.; Pagán, J.A.; Navarro, L.A.; García-Abujeta, J.L.; Carnés, J. Tomato NsLTP as an “In Vivo” Diagnostic Tool: Sensitization in a Mediterranean Population. J. Investig. Allergol. Clin. Immunol. 2015, 25, 196–204. [Google Scholar] [PubMed]
- Asero, R.; Antonicelli, L.; Arena, A.; Bommarito, L.; Colombo, G.; Crivellaro, M.; De Carli, M.; Della-Torre, E.; Della Torre, F.; Heffler, E.; et al. Causes of Food-Induced Anaphylaxis in Italian Adults: A Multi-Centre Study. Int. Arch. Allergy Immunol. 2009, 150, 271–277. [Google Scholar] [CrossRef]
- Zacharisen, M.C.; Elms, N.P.; Kurup, V.P. Severe Tomato Allergy (Lycopersicon esculentum). Allergy Asthma Proc. 2002, 23, 149–152. [Google Scholar]
- Worm, M.; Edenharter, G.; Ruëff, F.; Scherer, K.; Pföhler, C.; Mahler, V.; Treudler, R.; Lang, R.; Nemat, K.; Koehli, A.; et al. Symptom Profile and Risk Factors of Anaphylaxis in Central Europe. Allergy 2012, 67, 691–698. [Google Scholar] [CrossRef]
- Coombs, R.; Gell, P. The Classification of Allergic Reactions Underlying Disease. In Clinical Aspects of Immunology; Gell, P.G.H., Coombs, R.R.A., Eds.; SCIRP: Oxford, UK, 1963; pp. 317–337. [Google Scholar]
- Żukiewicz-Sobczak, W.A.; Wróblewska, P.; Adamczuk, P.; Kopczyński, P. Review Paper: Causes, symptoms and prevention of food allergy. Adv. Dermatol. Allergol. 2013, 30, 113–116. [Google Scholar] [CrossRef] [Green Version]
- Pravettoni, V.; Primavesi, L.; Farioli, L.; Brenna, O.; Pompei, C.; Conti, A.; Scibilia, J.; Piantanida, M.; Mascheri, A.; Pastorello, E. Tomato Allergy: Detection of IgE-Binding Lipid Transfer Proteins in Tomato Derivatives and in Fresh Tomato Peel, Pulp, and Seeds. J. Agric. Food Chem. 2009, 57, 10749–10754. [Google Scholar] [CrossRef] [PubMed]
- Bässler, O.Y.; Weiss, J.; Wienkoop, S.; Lehmann, K.; Scheler, C.; Dölle, S.; Schwarz, D.; Franken, P.; George, E.; Worm, M.; et al. Evidence for Novel Tomato Seed Allergens: IgE-Reactive Legumin and Vicilin Proteins Identified by Multidimensional Protein Fractionation−Mass Spectrometry and in Silico Epitope Modeling. J. Proteome Res. 2009, 8, 1111–1122. [Google Scholar] [CrossRef]
- Bleumink, E.; Berrens, L.; Young, E. Studies on the Atopic Allergen in Ripe Tomato Fruits. I. Isolation and Identification of the Allergen. Int. Arch. Allergy Appl. Immunol. 1966, 302, 132–145. [Google Scholar] [CrossRef]
- Bleumink, E. Food Allergy The Chemical Nature of the Substances Eliciting Symptoms. World Rev. Nutr. Diet. 1970, 12, 505–570. [Google Scholar] [CrossRef]
- Foetisch, K.; Son, D.; Altmann, F.; Aulepp, H.; Conti, A.; Haustein, D.; Vieths, S. Tomato (Lycopersicon esculentum) Allergens in Pollen-Allergic Patients. Eur. Food Res. Technol. 2001, 213, 259–266. [Google Scholar] [CrossRef]
- Kondo, Y.; Urisu, A.; Tokuda, R. Identification and Characterization of the Allergens in the Tomato Fruit by Immunoblotting. Int. Arch. Allergy Immunol. 2001, 126, 294–299. [Google Scholar] [CrossRef]
- Westphal, S.; Kolarich, D.; Foetisch, K.; Lauer, I.; Altmann, F.; Conti, A.; Crespo, J.; Rodríguez, J.; Enrique, E.; Vieths, S.; et al. Molecular Characterization and Allergenic Activity of Lyc e 2 (β-Fructofuranosidase), a Glycosylated Allergen of Tomato. Eur. J. Biochem. 2003, 270, 1327–1337. [Google Scholar] [CrossRef] [Green Version]
- Asero, R.; Mistrello, G.; Roncarolo, D.; de Vries, S.C.; Gautier, M.-F.; Ciurana, C.L.F.; Verbeek, E.; Mohammadi, T.; Knul-Brettlova, V.; Akkerdaas, J.H.; et al. Lipid Transfer Protein: A Pan-Allergen in Plant-Derived Foods That Is Highly Resistant to Pepsin Digestion. Int. Arch. Allergy Immunol. 2000, 122, 20–32. [Google Scholar] [CrossRef]
- Westphal, S.; Kempf, W.; Foetisch, K.; Retzek, M.; Vieths, S.; Scheurer, S. Tomato Profilin Lyc e 1: IgE Cross-Reactivity and Allergenic Potency. Allergy 2004, 59, 526–532. [Google Scholar] [CrossRef] [PubMed]
- Asero, R.; Mistrello, G.; Roncarolo, D.; Amato, S.; Arcidiacono, R.; Fortunato, D. Detection of a Novel Allergen in Raw Tomato. J. Investig. Allergol. Clin. Immunol. 2008, 18, 397–400. [Google Scholar] [PubMed]
- Palomares, O.; Villalba, M.; Quiralte, J.; Polo, F.; Rodriguez, R. 1,3-β-Glucanases as Candidates in Latex-Pollen-Vegetable Food Cross-Reactivity. Clin. Exp. Allergy 2005, 35, 345–351. [Google Scholar] [CrossRef] [PubMed]
- López, M.A.; Hernando de Larramendi, C.; Ferrer, A.; Huertas, A.; Pagán, J.; Garcia Abujeta, J.; Bartra, J.; Andreu, C.; Lavín, J.; Carnés, J. Identification and Quantification of Tomato Allergens: In Vitro Characterization of Six Different Varieties. Ann. Allergy. Asthma Immunol. 2011, 106, 230–238. [Google Scholar] [CrossRef]
- Diaz-Perales, A.; Collada, C.; Blanco, C.; Sanchez-Monge, R.; Carrillo, T.; Aragoncillo, C.; Salcedo, G. Cross-Reactions in the Latex-Fruit Syndrome: A Relevant Role of Chitinases but Not of Complex Asparagine-Linked Glycans. J. Allergy Clin. Immunol. 1999, 104, 681–687. [Google Scholar] [CrossRef]
- Krishnan, K.; Moens, P.D.J. Structure and Functions of Profilins. Biophys. Rev. 2009, 1, 71–81. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Carlsson, L.; Nyström, L.-E.; Sundkvist, I.; Markey, F.; Lindberg, U. Actin Polymerizability Is Influenced by Profilin, a Low Molecular Weight Protein in Non-Muscle Cells. J. Mol. Biol. 1977, 115, 465–483. [Google Scholar] [CrossRef]
- Rodríguez Del Río, P.; Díaz-Perales, A.; Sánchez-García, S.; Escudero, C.; Ibáñez, M.D.; Méndez-Brea, P.; Barber, D. Profilin, a Change in the Paradigm. J. Investig. Allergol. Clin. Immunol. 2018, 28, 1–12. [Google Scholar] [CrossRef] [Green Version]
- Darnowski, D.W.; Valenta, R.; Parthasarathy, M.V. Identification and Distribution of Profilin in Tomato (Lycopersicon esculentum Mill.). Planta 1996, 198, 158–161. [Google Scholar] [CrossRef]
- Zeleny, R.; Altmann, F.; Praznik, W. Structural Characterization of the N-Linked Oligosaccharides from Tomato Fruit. Phytochemistry 1999, 51, 199–210. [Google Scholar] [CrossRef]
- Priem, B.; Gitti, R.; Bush, C.A.; Gross, K.C. Structure of Ten Free N-Glycans in Ripening Tomato Fruit. Arabinose Is a Constituent of a Plant N-Glycan. Plant Physiol. 1993, 102, 445–458. [Google Scholar] [CrossRef] [Green Version]
- Valenta, R.; Duchene, M.; Ebner, C.; Valent, P.; Sillaber, C.; Deviller, P.; Ferreira, F.; Focke-Tejkl, M.; Edelmann, H.; Kraft, D. Profilins Constitute a Novel Family of Functional Plant Pan-Allergens. J. Exp. Med. 1992, 175, 377–385. [Google Scholar] [CrossRef] [Green Version]
- Vieths, S.; Scheurer, S.; Ballmer-Weber, B. Current Understanding of Cross-Reactivity of Food Allergens and Pollen. Ann. N. Y. Acad. Sci. 2002, 964, 47–68. [Google Scholar] [CrossRef] [PubMed]
- Valenta, R.; Duchêne, M.; Pettenburger, K.; Sillaber, C.; Valent, P.; Bettelheim, P.; Breitenbach, M.; Rumpold, H.; Kraft, D.; Scheiner, O.; et al. Identification of Profilin as a Novel Pollen Allergen; IgE Autoreactivity in Sensitized Individuals. Science 1991, 253, 557–560. [Google Scholar] [CrossRef] [PubMed]
- Willerroider, M.; Fuchs, H.C.; Ballmer-Weber, B.; Focke-Tejkl, M.; Susani, M.; Thalhamer, J.; Ferreira, F.; Wüthrich, B.; Scheiner, O.; Breiteneder, H.; et al. Cloning and Molecular and Immunological Characterisation of Two New Food Allergens, Cap a 2 and Lyc e 1, Profilins from Bell Pepper (Capsicum annuum) and Tomato (Lycopersicon esculentum). Int. Arch. Allergy Immunol. 2003, 131, 245–255. [Google Scholar] [CrossRef]
- de Martino, M.; Novembre, E.; Cozza, G.; de Marco, A.; Bonazza, P.; Vierucci, A. Sensitivity to Tomato and Peanut Allergens in Children Monosensitized to Grass Pollen. Allergy 1988, 43, 206–213. [Google Scholar] [CrossRef]
- Beezhold, D.H.; Sussman, G.L.; Liss, G.M.; Chang, N.S. Latex Allergy Can Induce Clinical Reactions to Specific Foods. Clin. Exp. Allergy 1996, 26, 416–422. [Google Scholar] [CrossRef]
- van Ree, R.; Voitenko, V.; van Leeuwen, W.A.; Aalberse, R.C. Profilin Is a Cross-Reactive Allergen in Pollen and Vegetable Foods. Int. Arch. Allergy Immunol. 1992, 98, 97–104. [Google Scholar] [CrossRef] [PubMed]
- Yu, L.X.; Parthasarathy, M.V. Molecular and Cellular Characterization of the Tomato Pollen Profilin, LePro1. PLoS ONE 2014, 9, e0086505. [Google Scholar] [CrossRef]
- Ebner, C.; Jensen-Jarolim, E.; Leitner, A.; Breiteneder, H. Characterization of Allergens in Plant-Derived Spices: Apiaceae Spices, Pepper (Piperaceae), and Paprika (Bell Peppers, Solanaceae). Allergy 1998, 53, 52–54. [Google Scholar] [CrossRef] [PubMed]
- Sturm, A.; Chrispeels, M. cDNA Cloning of Carrot Extracellular B-Fructosidase and Its Expression in Response to Wounding and Bacterial Infection. Plant Cell 1990, 2, 1107–1119. [Google Scholar] [CrossRef] [Green Version]
- Yelle, S.; Chetelat, R.T.; Dorais, M.; DeVerna, J.W.; Bennett, A.B. Sink Metabolism in Tomato Fruit 1: IV. Genetic and Biochemical Analysis of Sucrose Accumulation. Plant Physiol. 1991, 95, 1026–1035. [Google Scholar] [CrossRef] [Green Version]
- Sturm, A. Heterogeneity of the Complex N-Linked Oligosaccharides at Specific Glycosylation Sites of Two Secreted Carrot Glycoproteins. Eur. J. Biochem. 1991, 199, 169–179. [Google Scholar] [CrossRef]
- Asero, R.; Mistrello, G.; Roncarolo, D.; Amato, S.; Caldironi, G.; Barocci, F.; Ree, R. Immunological Cross-Reactivity between Lipid Transfer Proteins from Botanically Unrelated Plant-Derived Foods: A Clinical Study. Allergy 2002, 57, 900–906. [Google Scholar] [CrossRef] [PubMed]
- Sánchez-Monge, R.; Lombardero, M.; García-Sellés, F.J.; Barber, D.; Salcedo, G. Lipid-Transfer Proteins Are Relevant Allergens in Fruit Allergy. J. Allergy Clin. Immunol. 1999, 103, 514–519. [Google Scholar] [CrossRef]
- Salcedo, G.; Díaz-Perales, A.; Sánchez-Monge, R. Fruit Allergy: Plant Defence Proteins as Novel Potential Panallergens. Clin. Exp. Allergy J. Br. Soc. Allergy Clin. Immunol. 1999, 29, 1158–1160. [Google Scholar] [CrossRef]
- Boutrot, F.; Guirao, A.-L.; Alary, R.; Joudrier, P.; Gautier, M.-F. Wheat Non-Specific Lipid Transfer Protein Genes Display a Complex Pattern of Expression in Developing Seeds. Biochim. Biophys. Acta 2005, 1730, 114–125. [Google Scholar] [CrossRef] [PubMed]
- Douliez, J.-P.; Michon, T.; Khalil, E.; Marion, D. Structure, Biological and Technological Functions of Lipid Transfer Proteins and Indolines, the Major Lipid Binding Proteins from Cereal Kernels. J. Cereal Sci. 2000, 32, 1–20. [Google Scholar] [CrossRef]
- José-Estanyol, M.; Gomis-Rüth, F.X.; Puigdomènech, P. The Eight-Cysteine Motif, a Versatile Structure in Plant Proteins. Plant Physiol. Biochem. 2004, 42, 355–365. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yeats, T.H.; Rose, J.K.C. The Biochemistry and Biology of Extracellular Plant Lipid-Transfer Proteins (LTPs). Protein Sci. 2008, 17, 191–198. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Liu, F.; Zhang, X.; Lu, C.; Zeng, X.; Li, Y.; Fu, D.; Wu, G. Non-Specific Lipid Transfer Proteins in Plants: Presenting New Advances and an Integrated Functional Analysis. J. Exp. Bot. 2015, 66, 5663–5681. [Google Scholar] [CrossRef] [Green Version]
- Liu, W.; Huang, D.; Liu, K.; Hu, S.; Yu, J.; Gao, G.; Song, S. Discovery, Identification and Comparative Analysis of Non-Specific Lipid Transfer Protein (NsLtp) Family in Solanaceae. Genom. Proteom. Bioinform. 2010, 8, 229–237. [Google Scholar] [CrossRef] [Green Version]
- Li, F.; Fan, K.; Ma, F.; Yue, E.; Bibi, N.; Wang, M.; Shen, H.; Hasan, M.M.-U.; Wang, X. Genomic Identification and Comparative Expansion Analysis of the Non-Specific Lipid Transfer Protein Gene Family in Gossypium. Sci. Rep. 2016, 6, 38948. [Google Scholar] [CrossRef]
- D’Agostino, N.; Buonanno, M.; Ayoub, J.; Barone, A.; Monti, S.; Rigano, M. Identification of Non-Specific Lipid Transfer Protein Gene Family Members in Solanum lycopersicum and Insights into the Features of Sola l 3 Protein. Sci. Rep. 2019, 9, 1607. [Google Scholar] [CrossRef]
- Edstam, M.M.; Viitanen, L.; Salminen, T.A.; Edqvist, J. Evolutionary History of the Non-Specific Lipid Transfer Proteins. Mol. Plant 2011, 4, 947–964. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Van Loon, L.C.; Van Strien, E.A. The Families of Pathogenesis-Related Proteins, Their Activities, and Comparative Analysis of PR-1 Type Proteins. Physiol. Mol. Plant Pathol. 1999, 55, 85–97. [Google Scholar] [CrossRef]
- Asero, R.; Piantanida, M.; Pinter, E.; Pravettoni, V. The Clinical Relevance of Lipid Transfer Protein. Clin. Exp. Allergy 2017, 48, 6–12. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Matricardi, P.M.; Kleine-Tebbe, J.; Hoffmann, H.J.; Valenta, R.; Hilger, C.; Hofmaier, S.; Aalberse, R.C.; Agache, I.; Asero, R.; Ballmer-Weber, B.; et al. EAACI Molecular Allergology User’s Guide. Pediatr. Allergy Immunol. Off. Publ. Eur. Soc. Pediatr. Allergy Immunol. 2016, 27 (Suppl. 2), 1–250. [Google Scholar] [CrossRef] [PubMed]
- Salcedo, G.; Sánchez-Monge, R.; Barber, D.; Diaz-Perales, A. Plant Non-Specific Lipid Transfer Proteins: An Interface between Plant Defence and Human Allergy. Biochim. Biophys. Acta 2007, 1771, 781–791. [Google Scholar] [CrossRef] [PubMed]
- Volpicella, M.; Leoni, C.; Fanizza, I.; Rinalducci, S.; Placido, A.; Ceci, L. Expression and Characterization of a New Isoform of the 9 KDa Allergenic Lipid Transfer Protein from Tomato (Variety San Marzano). Plant Physiol. Biochem. 2015, 96, 64–71. [Google Scholar] [CrossRef]
- Martín-Pedraza, L.; Gonzalez Visiedo, M.; Gomez, F.; Blanca-Gómez, N.; Garrido-Arandia, M.; Rodriguez, R.; Torres, M.; Blanca, M.; Villalba, M.; Mayorga, C. Two Non-Specific Lipid Transfer Proteins (NsLTP) from Tomato Seeds Are Associated to Severe Symptoms of Tomato-Allergic Patients. Mol. Nutr. Food Res. 2016, 60, 1172–1182. [Google Scholar] [CrossRef]
- Giangrieco, I.; Alessandri, C.; Rafaiani, C.; Santoro, M.; Zuzzi, S.; Tuppo, L.; Tamburrini, M.; D’Avino, R.; Ciardiello, M.; Mari, A. Structural Features, IgE Binding and Preliminary Clinical Findings of the 7 KDa Lipid Transfer Protein from Tomato Seeds. Mol. Immunol. 2015, 66, 154–163. [Google Scholar] [CrossRef]
- Sinha, M.; Singh, R.P.; Kushwaha, G.S.; Iqbal, N.; Singh, A.; Kaushik, S.; Kaur, P.; Sharma, S.; Singh, T.P. Current Overview of Allergens of Plant Pathogenesis Related Protein Families. Sci. World J. 2014, 2014, 543195. [Google Scholar] [CrossRef] [Green Version]
- Wangorsch, A.; Jamin, A.; Foetisch, K.; Malczyk, A.; Reuter, A.; Vierecke, S.; Schülke, S.; Bartel, D.; Mahler, V.; Lidholm, J.; et al. Identification of Sola l 4 as Bet v 1 Homologous Pathogenesis Related-10 Allergen in Tomato Fruits. Mol. Nutr. Food Res. 2015, 59, 582–592. [Google Scholar] [CrossRef]
- Kurze, E.; Lo Scalzo, R.; Campanelli, G.; Schwab, W. Effect of Tomato Variety, Cultivation, Climate and Processing on Sola l 4, an Allergen from Solanum lycopersicum. PLoS ONE 2018, 13, e0197971. [Google Scholar] [CrossRef] [PubMed]
- Yokoi, H.; Yoshitake, H.; Mstsumoto, Y.; Kawada, M.; Takato, Y.; Shinagawa, K.; Sakurai, H.; Saito, K. Involvement of Cross-Reactive Carbohydrate Determinants-Specific IgE in Pollen Allergy Testing. Asia Pac. Allergy 2017, 7, 29. [Google Scholar] [CrossRef] [PubMed]
- Altmann, F. The Role of Protein Glycosylation in Allergy. Int. Arch. Allergy Immunol. 2007, 142, 99–115. [Google Scholar] [CrossRef]
- Le, L.Q.; Mahler, V.; Lorenz, Y.; Scheurer, S.; Biemelt, S.; Vieths, S.; Sonnewald, U. Reduced Allergenicity of Tomato Fruits Harvested from Lyc e 1-Silenced Transgenic Tomato Plants. J. Allergy Clin. Immunol. 2006, 118, 1176–1183. [Google Scholar] [CrossRef]
- Le, L.Q.; Mahler, V.; Scheurer, S.; Foetisch, K.; Braun, Y.; Weigand, D.; Enrique, E.; Lidholm, J.; Paulus, K.E.; Sonnewald, S.; et al. Yeast Profilin Complements Profilin Deficiency in Transgenic Tomato Fruits and Allows Development of Hypoallergenic Tomato Fruits. FASEB J. Off. Publ. Fed. Am. Soc. Exp. Biol. 2010, 24, 4939–4947. [Google Scholar] [CrossRef]
- Lien, L.; Lorenz, Y.; Scheurer, S.; Fötisch, K.; Enrique, E.; Bartra, J.; Biemelt, S.; Vieths, S.; Sonnewald, U. Design of Tomato Fruits with Reduced Allergenicity by DsRNAi-Mediated Inhibition of Ns-LTP (Lyc e 3) Expression. Plant Biotechnol. J. 2006, 4, 231–242. [Google Scholar] [CrossRef]
- Lorenz, Y.; Enrique, E.; Lequynh, L.; Fötisch, K.; Retzek, M.; Biemelt, S.; Sonnewald, U.; Vieths, S.; Scheurer, S. Skin Prick Tests Reveal Stable and Heritable Reduction of Allergenic Potency of Gene-Silenced Tomato Fruits. J. Allergy Clin. Immunol. 2006, 118, 711–718. [Google Scholar] [CrossRef]
- Dölle-Bierke, S.; Schwarz, D.; Lehmann, K.; Weckwerth, W.; George, E.; Worm, M.; Franken, P. Tomato Allergy: Impact of Genotype and Environmental Factors on the Biological Response. J. Sci. Food Agric. 2011, 91, 2234–2240. [Google Scholar] [CrossRef]
- Słowianek, M.; Skorupa, M.; Hallmann, E.; Rembialkowska, E.; Leszczynska, J. Allergenic Potential of Tomatoes Cultivated in Organic and Conventional Systems. Plant Foods Hum. Nutr. 2016, 71, 35–41. [Google Scholar] [CrossRef]
- Kiyota, K.; Yoshimitsu, M.; Satsuki-Murakami, T.; Akutsu, K.; Kajimura, K.; Yamano, T. Detection of the Tomato Allergen Sola l 1 and Evaluation of Its Reactivity after Heat and Papain Treatment. Food Agric. Immunol. 2017, 28, 1450–1459. [Google Scholar] [CrossRef] [Green Version]
- Pravettoni, V.; Primavesi, L.; Piantanida, M.; Brenna, O.V.; Farioli, L.; Scibilia, J.; Mascheri, A.; Pastorello, E.A. Tmoato Industrial Derivatives: Mallardo Reaction and Residual Allergenicity. Clin. Transl. Allergy 2011, 1, P19. [Google Scholar] [CrossRef] [Green Version]
- Primavesi, L.; Pravettoni, V.; Brenna, O.V.; Farioli, L.; Pastorello, E.A.; Pompei, C. Influence of Technological Processing on the Allergenicity of Tomato Products. Eur. Food Res. Technol. 2011, 232, 631–636. [Google Scholar] [CrossRef]
- Mócsai, R.; Maczó, A.; Grünwald-Gruber, C.; Majer-Baranyi, K.; Adányi, N.; Milotay, P.; Czelecz, J.; Tömösközi-Farkas, R. Investigation of Allergen Proteins in Five Tomato Cultivars. Acta Aliment. 2017, 46, 369–377. [Google Scholar] [CrossRef] [Green Version]


{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Description | Units | Concentration |
|---|---|---|
| Calcium (Ca) | mg/100 g | 105.21 ± 22,76 |
| Phosphorus (P) | mg/100 g | 300.99 ± 32.12 |
| Iron (Fe) | mg/100 g | 4.55 ± 2.18 |
| Sodium (Na) | mg/100 g | 70.38 ± 12.20 |
| Potassium (K) | mg/100 g | 403.02 ± 254.41 |
| Magnesium (Mg) | mg/100 g | 172.58 ± 58.92 |
| Vitamin A | IU/100 g | 614.44 ± 248.18 |
| Vitamin E | μg/100 g | 15.08 ± 1.06 |
| Thiamine | mg/100 g | 0.66 ± 0.44 |
| Riboflavin (mg) | mg/100 g | 0.48 ± 0.34 |
| Niacin (mg) | mg/100 g | 9.68 ± 0.00 |
| Ascorbic acid (mg) | mg/100 g | 36.16 ± 29.64 |
| Country | Total Production (×106 t) | Tomatoes for Processing | |
|---|---|---|---|
| (×106 t) | (% Total) | ||
| China | 55.72 | 5.60 | 11 |
| India | 18.74 | 0.13 | <1 |
| USA | 14.51 | 13.40 | 95 |
| Turkey | 11.85 | 2.70–3.9 | 25–30 |
| Egypt | 8.29 | 0.25 | 3 |
| Iran | 5.97 | 1.35 | 23 |
| Italy | 5.62 | 5.40 | 96 |
| Brazil | 4.30 | 1.30 | 33 |
| EU | 17.90 | 60 | 40 |
| World total | 171 | 41 | 24 |
| Plant Species | Allergen | Biochemical Name | MW (SDS-PAGE) |
|---|---|---|---|
| Solanum lycopersicum (Lycopersicon esculentum)—Tomato | Sola l 1 | Profilin | 14 kDa |
| Sola l 2 | Beta-fructofuranosidase | 50 kDa | |
| Sola l 3 | Non-specific lipid transfer protein type 1 (nsLTP1) | 9 kDa | |
| Sola l 4 | Pathogenesis-related protein, PR-10, Bet v 1 family member, TSI-1 | 20 kDa | |
| Sola l 5 | Cyclophilin | 19 kDa | |
| Sola l 6 | Non-specific lipid transfer protein type 2 (nsLTP2) | 7 kDa | |
| Sola l 7 | nsLTP type 1 | 12.5 kDa (SDS PAGE reducing) |
| Plant Species | Allergen Name | UNIPROT | Id (%) | |
|---|---|---|---|---|
| Tomato | Solanum lycopersicum | Sola l 1 | Q93YG7 | 100 |
| Bell Pepper | Capsicum annuum | Cap a 2 | Q93YI9 | 91.6 |
| Para rubber tree (Latex) | Hevea brasiliensis | Hev b 8 | O65812 | 84.7 |
| Pineapple | Ananas comosus | Ana c 1 | Q94JN2 | 80.6 |
| Apple | Malus domestica | Mal d 4 | Q9XF42 | 78.6 |
| Carrot | Daucus carota | Dau c 4 | Q8SAE6 | 77.6 |
| Celery | Apium graveolens | Api g 4 | Q9XF37 | 76.9 |
| Betula pendula | Betula erucosa | Bet v 2 | P25816 | 74.4 |
| Sugar Moiety | Mol-% in Sol l 2 |
|---|---|
| MUXF3 (Mana1–6(Xylb1–2)Man b1–4GlcNAcb1–4(Fuca1–3)GlcNAc) | 5.3 |
| MMX (Mana1–6(Mana1–3)(Xylb1–2)Manb1–4GlcNAcb1–4GlcNAc) | 8.2 |
| MMXF3 (Mana1–6(Mana1–3)(Xylb1–2)Man b1– 4GlcNAcb1–4(Fuca1–3)GlcNAc) | 83.6 |
| GnMXF3 (GlcNAcb1–2Mana1–6(GlcNAcb1–2Mana1–3)(Xylb1–2)Manb1–4GlcNAcb1–4(Fuca1–3)GlcNAc) | 2.3 |
| GnGnMXF3 (GlcNAcb1–2Mana1–6(Mana1–3)(Xylb1–2)Manb1–4GlcNAcb1–4(Fuca1–3)GlcNAc) | 0.6 |
| Plant Species | Bionomial Name | nsLTP | UNIPROT |
|---|---|---|---|
| Tomato | Solanum lycopersicum | Sol l 3 | P93224 |
| Peach | Prunus persica | Pru p 3 | P81402/Q9LED1 |
| Apple | Malus domestica | Mal d 3 | Q5J026 |
| Apricot | Prunus armeniaca | Pru ar 3 | P81651 |
| Plum | Prunus domestica | Pru d 3 | P82534 |
| Cherry | Prunus avium | Pru av 3 | Q9M5X8 |
| Strawberry | Fragaria ananassa | Fru a 3 | Q8VX12/Q4PLT9/Q4PLU0/Q4PLT6 |
| Lemon | Citrus limon | Cit l 3 | P84160 |
| Asparagus | Asparagus officinalis | Aspa o 1 | - |
| Lettuce | Lactuca sativa | Lec s 1 | - |
| Cabbage | Brassica oleracea | Bra o 3 | - |
| Latex | Hevea brasiliensis | Hev b 12 | Q8RYA8 |
| Parietaria | Parietaria judaica | Par j 1 | P43217/O04404/Q1JTN5/Q40905 |
| Ragweed | Ambrosia artemisiifolia | Amb a 6 | O04004 |
| Olive | Olea europoea | Ole e 7 | P81430 |
| Celery | Apium graveolens | Api g 2 | E6Y8S8 |
| Peanut | Arachis hypogaea | Ara h 9 | B6CEX8/B6CG41 |
| Asparagus | Asparagus officinalis | Aspa o 1 | - |
| Chestnut | Castanea sativa | Cas s 8 | - |
| Mugwort | Artimisia vulgaris | Art v 3 | P0C088/C4MGG9/C4MGH0/C4MGH1 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Włodarczyk, K.; Smolińska, B.; Majak, I. Tomato Allergy: The Characterization of the Selected Allergens and Antioxidants of Tomato (Solanum lycopersicum)—A Review. Antioxidants 2022, 11, 644. https://doi.org/10.3390/antiox11040644
Włodarczyk K, Smolińska B, Majak I. Tomato Allergy: The Characterization of the Selected Allergens and Antioxidants of Tomato (Solanum lycopersicum)—A Review. Antioxidants. 2022; 11(4):644. https://doi.org/10.3390/antiox11040644
Chicago/Turabian StyleWłodarczyk, Katarzyna, Beata Smolińska, and Iwona Majak. 2022. "Tomato Allergy: The Characterization of the Selected Allergens and Antioxidants of Tomato (Solanum lycopersicum)—A Review" Antioxidants 11, no. 4: 644. https://doi.org/10.3390/antiox11040644
APA StyleWłodarczyk, K., Smolińska, B., & Majak, I. (2022). Tomato Allergy: The Characterization of the Selected Allergens and Antioxidants of Tomato (Solanum lycopersicum)—A Review. Antioxidants, 11(4), 644. https://doi.org/10.3390/antiox11040644