1. Introduction
Certain aquaculture practices, such as overcrowding, periodic handling, sudden changes in temperature and poor nutritional quality, generate stress in reared fish [
1]. These stress situations could become ameliorable through the action of exogenous antioxidants, as their intake may mitigate oxidative damage by inhibiting the initiation or propagation of oxidative chain reactions [
2,
3]. Moreover, aquafeeds are usually rich in long chain polyunsaturated fatty acids (LC-PUFA), making them highly susceptible to lipid oxidation [
4]. In fish fillet, rancidity and fatty acid depletion are also promoted by the presence of highly unsaturated fatty acids, leading to a deterioration of fillet quality over time, albeit at different levels depending on species, age and diet composition [
5]. Thus, maintaining taste, colour, texture and freshness during storage is also a prime concern of the aquaculture industry [
6].
Given the increasing importance of aquaculture production, which represents 53% of total fish supply for human consumption [
7], the aforementioned issues associated with oxidative stress generate a rising necessity for the development of novel functional diets with physiological benefits beyond those provided by traditional feeds, particularly antioxidant benefits [
8,
9]. Due to their radical-scavenging properties and/or capacity of antioxidant system modulation, synthetic antioxidants are often used as additives for improving fish resistance to oxidative stress, as well as avoiding oxidative rancidity of fats and oils in feed mixtures [
10,
11]. Synthetic antioxidant compounds such as ethoxyquin, butylated hydroxyanisole (BHA) and butylated hydroxytoluene (BHT) are amongst the most widely used antioxidants in food and feed formulations [
4]. In 2017, the common use of ethoxyquin was banned in raw materials, feed premixes, additives and food produced and/or commercialized in the European Union (EU 2017/962). This occurred due to possible genotoxicity effects of one of its metabolites (ethoxyquin quinone imine), as well as the fact that p-phenetidine, an impurity of ethoxyquin, is recognised as a possible mutagen [
12]. Meanwhile, BHT (E321) and BHA (E320) are both widely used as antioxidants in feed for all animal species, with a maximum limit of 150 mg kg
−1 each, or in combination, authorised in the EU [
13]. Nevertheless, BHT has been found to inhibit humoral immune response in animals and to possess tumour promotion effects, and despite not being genotoxic, it modifies the genotoxicity of other agents [
14]. This, together with the proven effectiveness of natural antioxidants for inhibiting rancidity in food [
15] and in providing health advantages for consumers [
16,
17], prompted the scientific community and the feed industry to look for natural alternatives with strong anti-oxidant activity for inclusion in animal feeds [
9,
18,
19,
20,
21].
Vitamins, carotenoids and phenolic compounds are antioxidant molecules commonly present in fruits, herbs and vegetables such as tomato, carrot and coriander [
22,
23,
24]. Specifically, vitamin E is a generic term commonly used for describing eight different lipid-soluble antioxidant forms (tocopherols and tocotrienols), the form of α-tocopherol having the greatest impact on the fish antioxidant system [
25]. These compounds are easily oxidized due to their inherent hydroxyl moiety on carbon 6, which has an important role in the maintenance of normal metabolic processes and physiological functions in the body, mainly due to its radical-scavenging properties [
26,
27,
28], thus playing an important role in the protection of cell membranes from lipid peroxidation [
29,
30]. Vitamin E also shows a stimulatory effect on fish’s immune system, while improving fish health and disease resistance [
31,
32]. This antioxidant compound has been proven to have an essential protective role against the adverse effects of reactive oxygen species and other free radicals [
4], which is important for quality preservation of fish fillet, either raw or cooked. To date, the positive influence of vitamin E on seafood quality has been studied in several commercially reared fish species, such as rainbow trout (
Oncorhynchus mykiss) [
33], turbot (
Scophthalmus maximus) [
34] and European sea bass (
Dicentrarchus labrax) [
35]. The minimum recommended doses of vitamin E in fish dietary mixes range from 25 to 200 mg kg
−1, depending on the species and maturation state [
4]. Although vitamin E supplementation has been demonstrated as essential for the development of European sea bass larvae [
36], very few data are available regarding the antiperoxidative effects of vitamin E in commercial-sized European sea bass. Gatta et al. [
35] suggested that the α-tocopherol content of diets for European sea bass (208 g) should be above 254 mg kg
−1, and up to 942 mg kg
−1, in order to reduce lipid oxidation.
Phenolic compounds and carotenoids are also associated with a wide range of biological activities, including antioxidant properties that contribute both directly and indirectly to the inhibition or suppression of oxidation processes [
17,
37]. Carotenoid-rich diets are effective in terms of preventing oxidative stress and enhancing innate immune system defences. Ehsani et al. [
21] found that dietary lycopene, naturally abundant in carrots and tomatoes, is effective in preventing lipid oxidation in rainbow trout, subsequently delaying fillet rancidity. β-carotene, found in high doses in carrot and coriander acted as an antioxidant and immunostimulant, mitigating the negative effects of peroxide radicals in Nile tilapia (
Oreochromis niloticus) [
18]. Both synthetic and natural carotenoid dietary inclusion has been proven to stimulate antioxidant potential in European sea bass [
6,
38,
39], while immunomodulatory effects were reported with the dietary inclusion of carotenoid rich sources such as seaweed [
38].
Annual production of plants and aromatic herbs has passed 600 million tonnes per year [
39], generating a high amount of coproducts that are mostly wasted. Despite viability for consumption, about a third of total farm production in the EU is discarded, mostly due to cosmetic and retailers’ standards [
40]. These otherwise discarded coproducts can be valuable sources of nutrients, antioxidants and bioactive compounds for aquafeeds that at a reduced price may have a positive effect on weight gain, appetite stimulation and overall fish health [
41,
42,
43,
44], as well as in fillet quality traits [
17,
21]. Adding these vegetable coproducts to aquafeeds would also address current consumer trends in terms of preferring natural food sources over synthetic ones [
45], and promote a circular economy approach towards sustainability.
The novel approach of this study consisted of evaluating the potential of vegetable antioxidants-rich coproducts as additional antioxidant sources in diets for European sea bass, a carnivorous fish of high economical value, commonly reared in the Mediterranean region. The effects of such natural antioxidants on European sea bass growth, immune system and fillet quality traits will be compared to those of synthetic vitamin E included in diets at either a regular or reinforced dose (100 and 500 mg kg−1, respectively).
4. Discussion
Both synthetic and natural antioxidants are commonly included in aquafeeds in order to ameliorate the negative effects of oxidative stress, potentially increasing fish health and delaying flesh oxidation during storage [
4], the physical and chemical conditions that occur during feed formulation, namely light exposure and high temperatures, may negatively affect antioxidant stability [
76,
77].
The high vulnerability of carotenoids to high temperatures is thoroughly documented [
78]. Despite the fact that dried carrot and coriander used in this study revealed the presence of α-carotene and β-carotene, results showed a complete absence of these carotenoids in their respective experimental diets. In the case of lutein, although dried coriander was a very rich source, its 2% inclusion in CO diet was not reflected in the final dietary level. The measured amounts of lutein in the CO diet were 1.7 g 100 g
−1 of DM, representing 68% of the expected value, i.e., 2.5 g 100 g
−1 of DM. This result points towards a possible carotenoid degradation during the extrusion (110 °C) and/or drying (60 °C) stages of diet manufacture. Of the three ingredients used in this study, only coriander contained β-cryptoxanthin, but all diets just evidenced trace amounts of this pigments, again suggestion degradation during processing. The CA diet contained the highest amount of total polyphenols, thus significantly differing from the CTRL, possibly due to the dietary inclusion of dried carrot.
Despite synthetic vitamin E being a commonly used antioxidant in aquaculture, there is little knowledge regarding supplementation rates for commercial-sized European sea bass (
Dicentrarchus labrax). Overall, quantification of vitamin E in experimental diets used in this study showed tocopherol amounts that reflected the different supplementation levels, since the VITE diet had five times more α-tocopherol than the remaining diets. Moreover, the dried vegetables added at a 2% inclusion rate did not provide any further significant contribution of vitamin E to the dietary formulations. However, supplementation doses did not reach targeted levels (500 mg kg
−1 in VITE and 100 mg kg
−1 in all other diets) as values measured after feed manufacturing procedures of extrusion and drying (126 mg kg
−1 of DM and 25–30 mg kg
−1 of DM, respectively) indicated a 24–30% α-tocopherol retention after feed manufacturing. During the feed formulation process, namely during the extrusion and drying stages, tocopherol suffers considerable temperature-induced degradation, an effect which is thoroughly documented [
79,
80,
81]. Vitamin stability during extrusion depends on several factors, namely raw material, mixing, conditioning, temperature, pressure, moisture, energy input and extruder mechanical features [
82]. According to Morin et al. [
83], the chemical nature of the extruded matrix, moisture and temperature levels account for a variation of 63 ± 28% of tocopherol retention. Riaz and Ali [
82] showed that the pelleting and extrusion processes alone can account for a 25% loss of vitamin E at around 80–90 °C, whilst higher extrusion temperatures, above 100 °C, increases the sensitivity of this vitamin significantly [
83]. According to Anderson and Sunderland [
76], most vitamin E losses occur over the course of aquafeeds extrusion process, before drying procedures, and might negatively affect the antioxidant potential of the diets. Effective technologies (e.g., colder extrusion and softer drying procedures, microencapsulation) able to protect both pigments and natural antioxidants sources prior extrusion should be envisaged.
The VITE diet exhibited less DPPH
• scavenging capacity than the CO diet, despite showing no differences when compared to control. Moreover, several synergistic mechanisms between different natural antioxidants, such as those found in coriander, are known to heighten the antioxidant potential of biological samples, generating superior antioxidant characteristics as opposed the sums of each individual one [
84,
85,
86]. When compared to the control, the antioxidant capacity measured in diet VITE does not seem dose responsive. Previous studies suggested that the DPPH and ABTS method can be employed to examine lipophilic antioxidants such as tocopherols (vitamin E) [
85,
87]. However, the DPPH
• reaction rates are highly influenced by solvent composition [
88], suggesting that results have to be interpreted with caution. In the present study, the DPPH and ABTS assays were carried out using the same extract employed for polyphenol quantification (via the Folin–Ciocalteau method), that is, methanol and distilled water (4:1, vv
−1). This mixture is optimal for polyphenols due to the solvent’s polarity [
89], but methanol is not the most appropriate solvent for tocopherol extraction, as it is a hydrophilic substance [
90,
91]. Thus, observed values from the DPPH and ABTS assays might not reflect the totality of radical scavenging capacity resulting from the addition of synthetic vitamin E, which may explain the lack of differences between VITE and the CTRL diet.
The chemical structure of carotenoids and polyphenols allows these compounds to regulate antioxidant activity due to their radical-scavenging properties [
92]. Specifically, the antioxidant activity of lutein [
93,
94], β-carotene [
93,
95], lycopene [
93,
96], α-carotene and β-cryptoxanthin [
93] has been thoroughly evaluated in in vitro studies. Supplementation of fish feeds with carotenoids has shown health associated benefits, acting as antioxidants and immunostimulants, enhancing fish resistance to bacterial and fungal diseases [
97]. Additionally, polyphenols show an ability of scavenging free radicals, assisting in hindering the negative effects of ROS such as singlet oxygen, peroxynitrite and hydrogen peroxide, which must be continually removed from cells to maintain healthy metabolic function [
98,
99]. Natural phenolic compounds can also serve as potential additives for preventing quality deterioration or to retain the quality of fish and fish products [
100], and seem to be rather resistant to deactivation via high-temperature extrusion-cooking process [
101]. This antioxidant potential of polyphenols in terms of direct and indirect inhibition or suppression of oxidation processes is thoroughly established, mostly through trolox equivalent antioxidant capacity assays, as well as their scavenging capacity regarding the stable free radical 2,2-diphenyl-1-picryl-hydrazyl, commonly known as DPPH, due to its affinity with fat-soluble hydrophobic compounds, as is the case with polyphenols [
102,
103]. In this study, the CA diet revealed higher amounts of phenolic compounds (1179.1 ± 103.4 mg GAE 100 g
−1 DM) than all remaining diets. This could be advantageous for the conservation of feed properties, as the proven antioxidant potency of polyphenols [
17,
100] might provide additional resistance to feed oxidation during storage.
Data from this study show that the addition of natural antioxidants to the experimental diets did not affect any of the evaluated immune parameters in fish. Moreover, considering the results of liver antioxidant activity observed in this study, GR was significantly higher in fish fed CTRL and CO diets when compared to fish fed with the TO diet. GR is an essential enzyme for catalysing the reaction that reduces oxidized glutathione (GSSR) into reduced glutathione (GSH) [
104], the sum of which comprises total glutathione content (TG). GSH is an essential cofactor for antiperoxidative enzymes such as glutathione peroxidase (GPx) [
104]. Therefore, a lower GR associated with the TO diet, in combination with liver lipid peroxidation data that showed no significant differences between TO and CTRL, might mean that fish fed with TO require a lower endogenous antioxidant activity in order to maintain cell homeostasis. However, TAC, which is specific for non-enzymatic antioxidants, displayed no differences between experimental treatments, meaning that we cannot directly attribute this lower production of GR to an increase in antioxidant potential stemming from exogenous antioxidants. As expected, due to the absence of differences between treatments regarding TG, glutathione-dependent antiperoxidative enzymes GPx and GST also did not show any differences when experimental diets were compared to the CTRL, as these enzymes require glutathione as a cofactor in order to perform their biological functions [
105]. The antioxidant function of GPx and GST is largely dependent on its interaction with TG [
106], neutralizing hydroperoxides as GSH is oxidized to GSSG by GR. Considering the results obtained in this study, we can observe that a lesser production of GR did not lead to differences in glutathione dependent enzymes. Moreover, the heightened dose of polyphenols in the CA diet did not translate into additional antioxidant protection in fish liver. This raises further questions concerning the bioavailability of these polyphenols in sea bass organism. The bioaccessibility and bioavailability of natural antioxidants in the organism not only relies on the concentration of bioactive compounds in the ingredient, but also on dosage and form of administration, composition of the feed matrix, while also being heavily influenced by other factors such as pH variations, enzyme action and digestion time [
107,
108]. Hence, further research is still warranted to clarify the full potential of natural antioxidant sources for inclusion in aquafeeds.
Overall, neither a heightened dose of vitamin E compared to standard values nor a 2% inclusion of natural antioxidant induced an upregulation of sea bass liver antioxidant system. Evaluation of the antioxidant potential present in these diets after the manufacturing process (extrusion and drying) only provides a limited perspective on their potential biological effect, since this is also greatly conditioned by their digestive bioaccessibility and bioavailability as discussed above.
Dietary vitamin E and natural antioxidants at the levels used in this study had no effects on fish proximate composition and feed conversion rate, confirming data obtained by Gatta et al. [
35].
The beneficial effects of vitamin E as an antioxidant have been thoroughly evaluated in teleost fish, namely gilthead seabream (
Sparus aurata) [
109], red sea bream (
Pragus major) [
110] and rainbow trout (
Oncorhynchus mykiss) [
111,
112]. However, in Atlantic salmon (
Salmo salar) (IBW = 64 g), high supplementation levels of up to 1100 mg kg
−1 dietary vitamin E did not affect fish antioxidant defence, lipid peroxidation and overall fish muscle resistance to oxidative stress [
113]. Indeed, the available literature shows that the effectiveness of vitamin E as an antioxidant is largely dependent on fish life stage and species [
4].
Overall, studies concerning the antioxidant effects of vitamin E of commercial-sized European sea bass are rare. Silva et al. [
114] recommended a dosage of 500 mg kg
−1 of vitamin E for adult sea bass. However, data obtained in this study suggests that a 500 mg kg
−1 inclusion of vitamin E (VITE diet) in European sea bass feeds have no beneficial effect on muscle antioxidant potential compared to the traditional dosage of 100 mg kg
−1 (CTRL diet). Gatta et al. [
35] also reported decreased lipid peroxidation rates in sea bass fillets when α-tocopherol supplementation was increased from 139 mg kg
−1 feed to 493 mg kg
−1, but this could not be confirmed in the present study. It should be noted that all these results were obtained in optimised non-stressful rearing conditions for sea bass. Different conclusions regarding the antioxidant potential of these diets might have been reached if fish were submitted to a stress challenge, as enzymatic responses to oxidative stress are particularly promoted when fish face a pro-oxidant challenge [
115]. Further studies should hence be envisaged to explore fish response to stressful conditions after being fed natural antioxidant sources.
Although vegetable coproduct inclusion and different vitamin E inclusion levels failed to show any antioxidant benefits in European sea bass muscle, significant differences were identified between Day 0 and Day 8. Namely, the antioxidant potential measured through the scavenging potential for DPPH
• was higher in fish stored in ice for 8 days, irrespective of the dietary treatments. This might have contributed towards the absence of differences in muscle LPO between Day 0 and Day 8, showing that lipid oxidation levels were similar between samples from both days. Likewise, Gatta et al. [
35] showed that vitamin E supplementations between 139 mg kg
−1 and 942 mg kg
−1 fed to European sea bass (IBW = 200 g) showed no differences in terms of muscle lipid peroxidation between the first and last days of a 12-day storage time. The ability to scavenge free radicals is essential for ameliorating the negative effects of oxidative stress. Due to its relative stability, the free radical DPPH
• is a prime candidate as a first approach for evaluation of radical scavenging activity [
116]. The eventual breakdown of cohesiveness between tissues, liquefaction of most organs and subsequent decomposition of proteins by hydrolysis leads to an increase in amino acid content, which might consequentially increase muscle free radical scavenging activity [
117]. Moreover, naturally-occurring ROS via interactions with coproducts generated through the natural functioning processes of electron transport chains [
118], cease functioning after death. Thus, a post-mortem increase in amino acid content accompanied by a lesser formation of ROS might explain this increase in DPPH
• radical scavenging activity. Plant by-products have also been proven effective in delaying chemical changes and microbial growth, as well as maintaining sensory characteristics and extending the shelf-life of seafood during refrigerated storage [
102]. Natural antioxidants, namely phenolic compounds can have positive effects in terms of upregulation of muscle antibacterial properties and retardation of bacterial growth [
102], which would translate into a larger product shelf-life. This was, however, not accessed in the present study and merits further evaluation.
Besides their antioxidant properties, carotenoids are also sources of pigments, and their deposition in tissues may affect skin/muscle colour and appearance in fish [
119]. In this study, instrumental colour data of fish muscle was consistent with data found in the literature [
120]. Moreover, all diets with natural antioxidant inclusion showed a significant decrease in muscle yellowness (
b*) when compared to CTRL, whilst fish fed with CA presented a significantly higher
h* than CTRL. Dietary coriander (CO) was associated with decreased muscle chroma (
C*). Despite the evidence for dietary carotenoid degradation, the different diets still seem to modulate fish muscle colour. However, these differences in raw muscle colorimetric analyses between TO, CA and CO, when compared to CTRL, could not be perceived in the cooked muscle slices that were equally well accepted by the sensory panel. Similarly, in large-sized European sea bass, significant alterations in fillet colour of fish fed with
Isochrysis sp., could not be detected by a sensory panel [
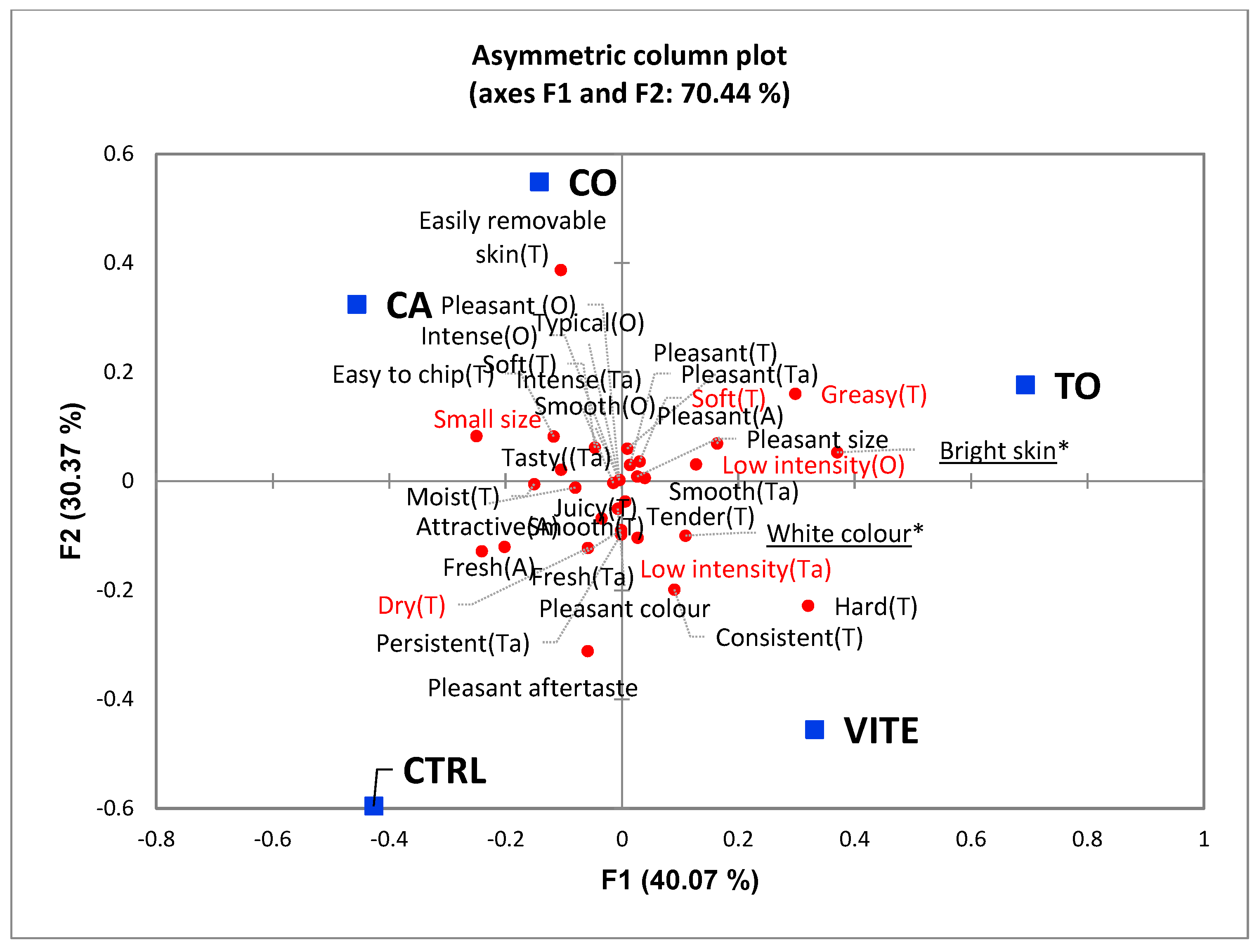
120]. It is important to note that consumers were only able to significantly differentiate two attributes. TO group presented brighter skin than others, while VITE samples presented a white colour significantly different from the other groups. Both differences were mentioned as positive aspects.
This is particularly important in commercial fish species, since colour and visual appearance are known to influence market value, flavour perception and acceptability of fish food products [
121], thus providing a measurable parameter for flesh freshness and ultimately affecting consumer perception quality [
122,
123]. Although differences in food colour can exert considerable influence on taste and consumers’ perception [
121], overall liking scores in terms of consumer acceptability, showed no significant differences between samples. Indeed, all samples had a high average score value, in line with a high mention of positive comments, revealing that consumers were unable to detect differences between dietary treatments.
One of the most important freshness quality attributes of fish muscle is texture, which is heavily dependent on several parameters such as hardness, cohesiveness, springiness, chewiness, resilience and adhesiveness, as well as fibre detachment and internal cross-linking of connective tissue [
124]. In terms of muscle texture, there were no significant differences among dietary treatments. Although hardness, adhesiveness, cohesiveness, gumminess and chewiness decreased after storage time, accompanied by an increase in springiness and resilience, none of these parameters were affected by the experimental diets.


 ,
, 







{kind=link}
{kind=link}