
How Does LC/MS Compare to UV in Coffee Authentication and Determination of Antioxidant Effects? Brazilian and Middle Eastern Coffee as Case Studies


 ,
, 
Abstract
1. Introduction
2. Materials and Methods
2.1. Samples Collection and Preparation
2.2. Samples Extraction and UHPLC-ESI–HRMS Analysis
2.3. UV Measurements and Multivariate Data Analysis
2.4. Tenative Identification of Metabolites Analyzed by UHPLC-ESI–HRMS
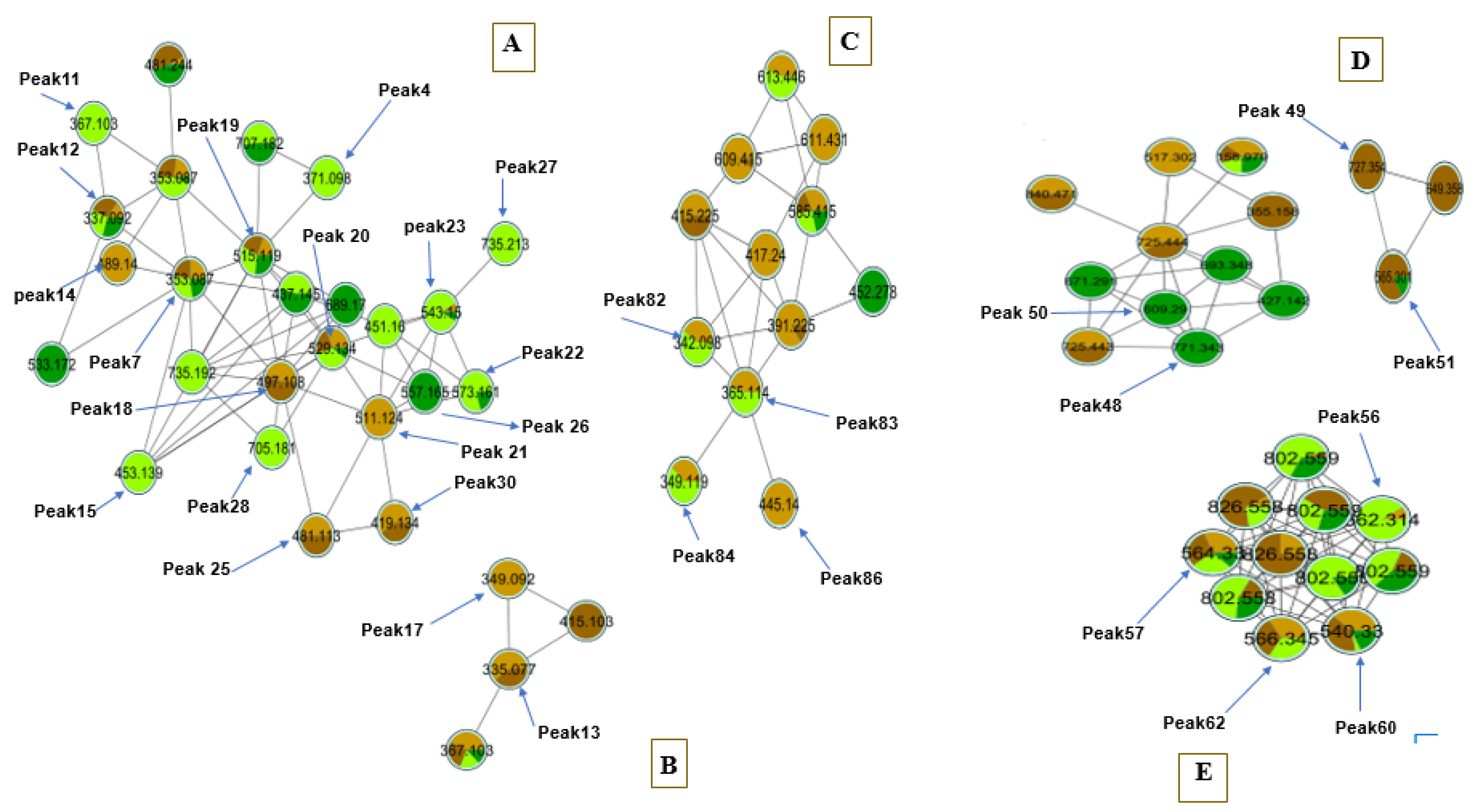
2.5. Molecular Based Networking of Coffee Specimens
2.6. Determination of Total Phenolic Content
2.7. Antioxidant Assays
2.7.1. DPPH Radical Scavenging Assay
2.7.2. Ferric Reducing Antioxidant Power (FRAP) Assay
3. Results and Discussion
3.1. Metabolite Profiling via UHPLC-ESI–HRMS
3.1.1. Alkaloids
3.1.2. Hydroxycinnamate Derivatives
3.1.3. Diterpenes
3.1.4. Fatty Acids and Sphingolipids
3.1.5. Serotonin Amides (Hydroxytryptamine Derivatives)
3.1.6. Miscellaneous
3.2. UHPLC–HRMS Based Multivariate Data Analyses and Fingerprinting of Coffee Samples
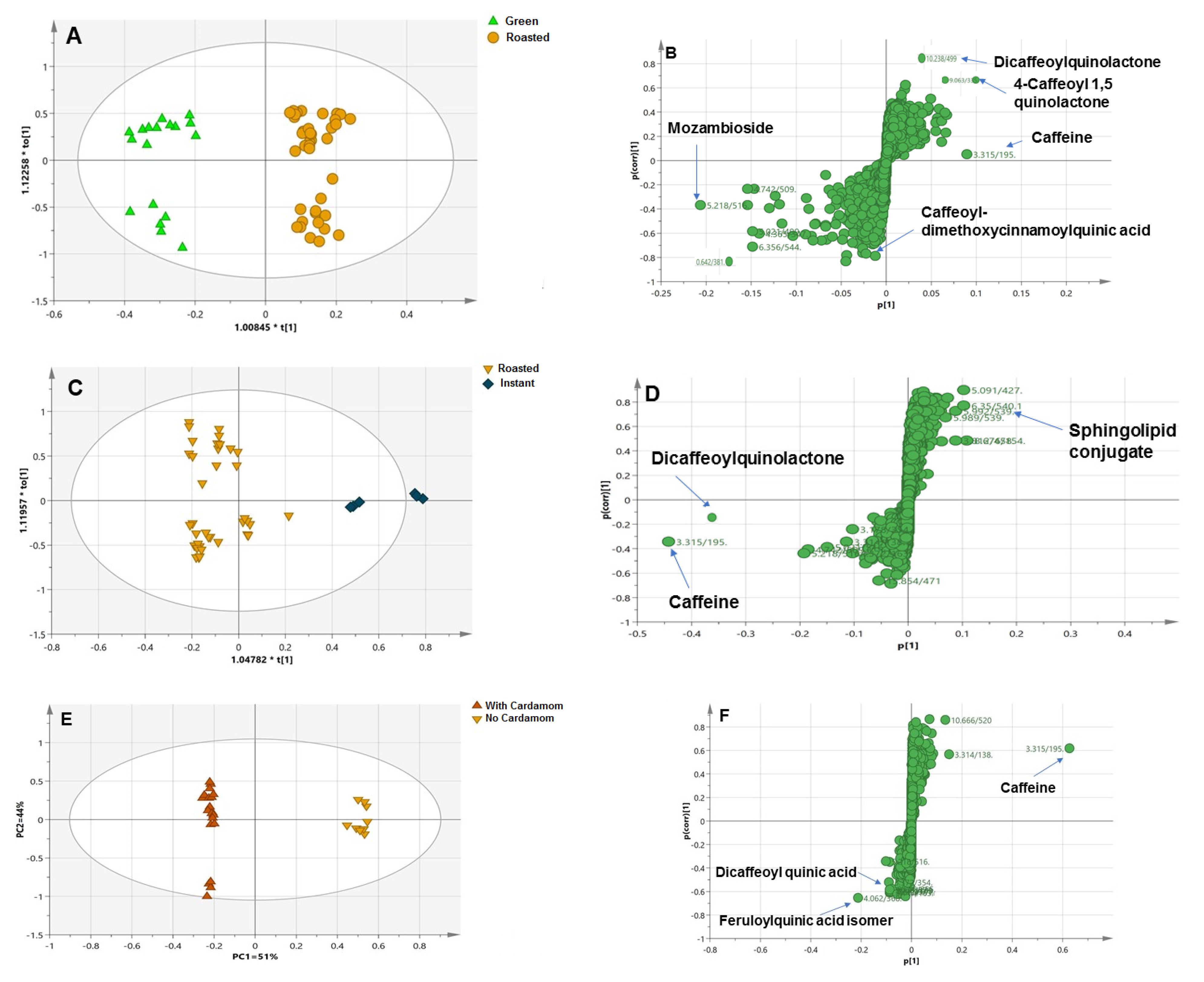
3.2.1. Roasted versus Green Coffee
3.2.2. Instant versus Roasted Coffee Samples
3.2.3. Blended versus Plain Samples
3.3. UV–Vis Fingerprinting of Coffee Seeds
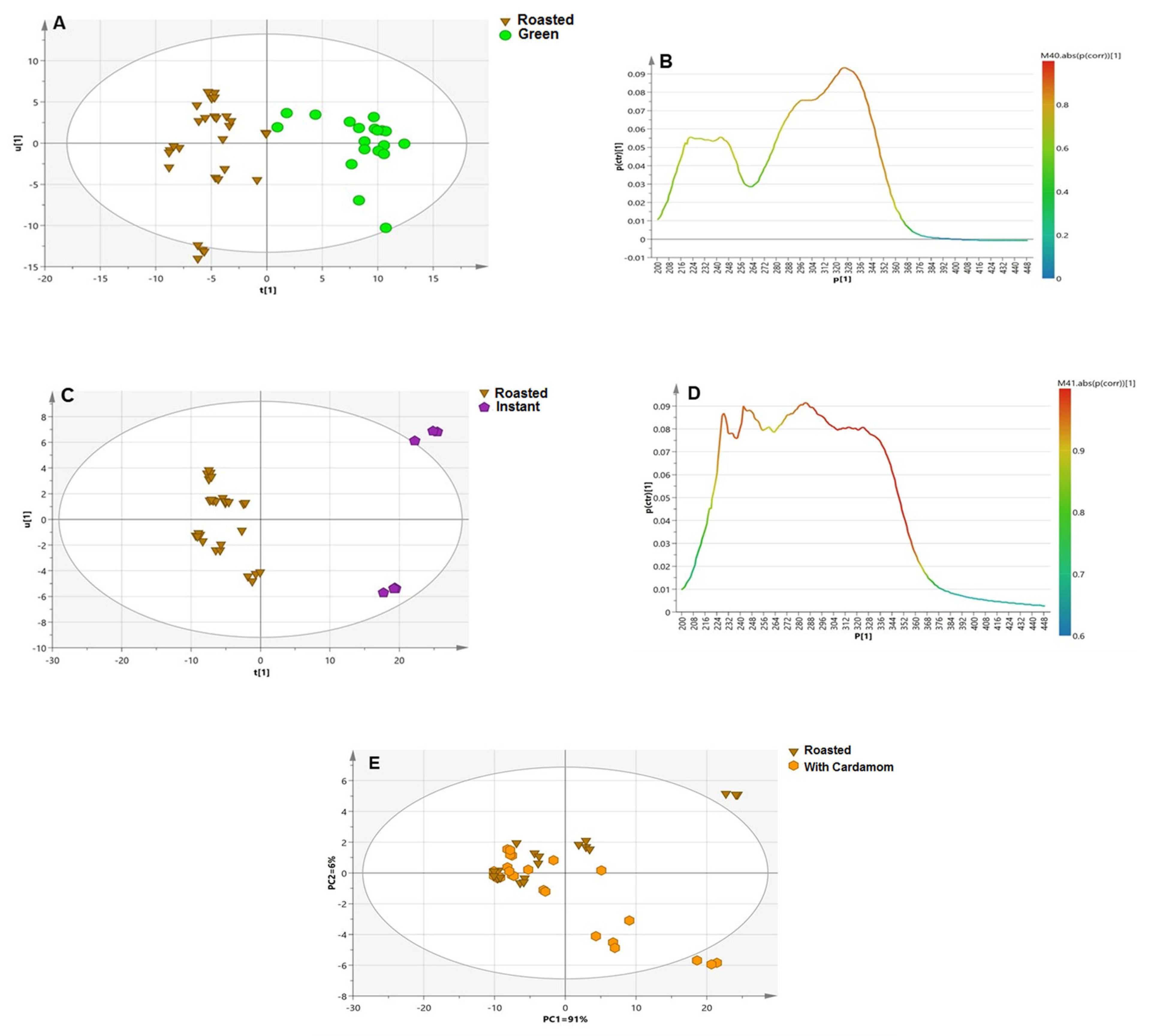
3.3.1. Roasted versus Green Coffee Specimens
3.3.2. Instant versus Roasted Samples
3.3.3. Blended versus Plain Coffee Samples
3.4. Comparison between UHPLC–MS and UV Fingerprinting Multivariate Data Analysis Models
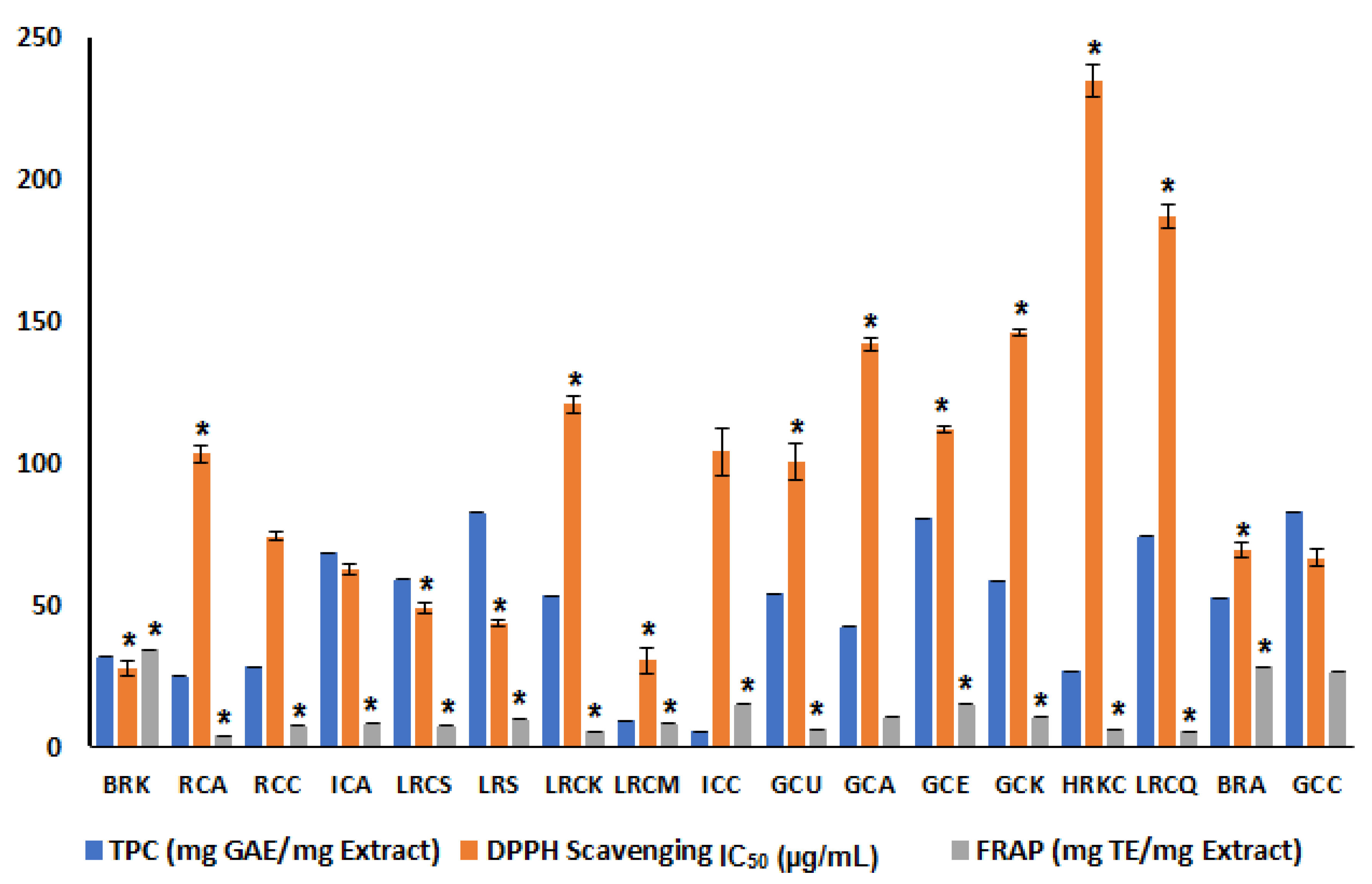
3.5. Determination of Total Phenolic Content of Coffee Species
3.6. In Vitro Antioxidant Activity
3.6.1. DPPH Assay
3.6.2. In Vitro FRAP Assay
3.7. Correlation between Biological Assays and UHPLC–MS Metabolite Profile
4. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Mahmud, M.M.C.; Shellie, R.A.; Keast, R. Unravelling the relationship between aroma compounds and consumer acceptance: Coffee as an example. Compr. Rev. Food Sci. Food Saf. 2020, 19, 2380–2420. [Google Scholar] [CrossRef] [PubMed]
- Samper, L.; Giovannucci, D.; Vieira, L. The Powerful Role of Intangibles in the Coffee Value Chain; World Intellectual Property Organization: Geneva, Switzerland, 2017; Volume 39. [Google Scholar]
- Shokouh, P.; Jeppesen, P.B.; Christiansen, C.B.; Mellbye, F.B.; Hermansen, K.; Gregersen, S. Efficacy of arabica versus robusta coffee in improving weight, insulin resistance, and liver steatosis in a rat model of type-2 diabetes. Nutrients 2019, 11, 2074. [Google Scholar] [CrossRef]
- De Rosso, M.; Colomban, S.; Flamini, R.; Navarini, L. UHPLC-ESI-QqTOF-MS/MS characterization of minor chlorogenic acids in roasted Coffea arabica from different geographical origin. J. Mass Spectrom. 2018, 53, 763–771. [Google Scholar] [CrossRef] [PubMed]
- Ren, Y.; Wang, C.; Xu, J.; Wang, S. Cafestol and kahweol: A review on their bioactivities and pharmacological properties. Int. J. Mol. Sci. 2019, 20, 4238. [Google Scholar] [CrossRef] [PubMed]
- Cavin, C.; Holzhaeuser, D.; Scharf, G.; Constable, A.; Huber, W.W.; Schilter, B. Cafestol and kahweol, two coffee specific diterpenes with anticarcinogenic activity. Food Chem. Toxicol. Int. J. Publ. Br. Ind. Biol. Res. Assoc. 2002, 40, 1155–1163. [Google Scholar] [CrossRef]
- Al Doghaither, H.A.; Al-Malki, E. The addition of herbal additives influences the antioxidant activity of traditional arabic coffee. World Appl. Sci. J. 2017, 35, 393–398. [Google Scholar]
- Abdelwareth, A.; Zayed, A.; Farag, M.A. Chemometrics-based aroma profiling for revealing origin, roasting indices, and brewing method in coffee seeds and its commercial blends in the Middle East. Food Chem. 2021, 349, 129162. [Google Scholar] [CrossRef]
- Sonam, K.S.; Guleria, S. Synergistic antioxidant activity of natural products. Ann. Pharmacol. Pharm. 2017, 2, 1086. [Google Scholar]
- Yashin, A.; Yashin, Y.; Wang, J.Y.; Nemzer, B. Antioxidant and antiradical activity of coffee. Antioxidants 2013, 2, 230–245. [Google Scholar] [CrossRef]
- Otify, A.M.; El-Sayed, A.M.; Michel, C.G.; Farag, M.A. Metabolites profiling of date palm (Phoenix dactylifera L.) commercial by-products (pits and pollen) in relation to its antioxidant effect: A multiplex approach of MS and NMR metabolomics. Metab. Off. J. Metab. Soc. 2019, 15, 119. [Google Scholar] [CrossRef] [PubMed]
- Nunez, N.; Martinez, C.; Saurina, J.; Nunez, O. High-performance liquid chromatography with fluorescence detection fingerprints as chemical descriptors to authenticate the origin, variety and roasting degree of coffee by multivariate chemometric methods. J. Sci. Food Agric. 2020, 101, 65–73. [Google Scholar] [CrossRef]
- Mattoli, L.; Cangi, F.; Maidecchi, A.; Ghiara, C.; Ragazzi, E.; Tubaro, M.; Stella, L.; Tisato, F.; Traldi, P. Metabolomic fingerprinting of plant extracts. J. Mass Spectrom. JMS 2006, 41, 1534–1545. [Google Scholar] [CrossRef] [PubMed]
- Liang, X.; Zhao, C.; Su, W. A Fast and Reliable UPLC-PAD Fingerprint Analysis of Chimonanthus salicifolius Combined with Chemometrics Methods. J. Chromatogr. Sci. 2016, 54, bmw053. [Google Scholar] [CrossRef][Green Version]
- Terrile, A.E.; Marcheafave, G.G.; Oliveira, G.S.; Rakocevic, M.; Bruns, R.E.; Scarminio, I.S. Chemometric analysis of UV characteristic profile and infrared fingerprint variations of Coffea arabica green beans under different space management treatments. J. Braz. Chem. Soc. 2016, 27, 1254–1263. [Google Scholar]
- Bobková, A.; Hudáček, M.; Jakabová, S.; Belej, L’.; Capcarová, M.; Čurlej, J.; Bobko, M.; Árvay, J.; Jakab, I.; Čapla, J.; et al. The effect of roasting on the total polyphenols and antioxidant activity of coffee. J. Environ. Sci. Health Part B 2020, 55, 495–500. [Google Scholar] [CrossRef] [PubMed]
- Zayed, A.; Abdelwareth, A.; Mohamed, T.A.; Fahmy, H.A.; Porzel, A.; Wessjohann, L.A.; Farag, M.A. Dissecting coffee seeds metabolome in context of genotype, roasting degree, and blending in the Middle East using NMR and GC/MS techniques. Food Chem. 2022, 373, 131452. [Google Scholar] [CrossRef]
- Farag, M.A.; Gad, H.A.; Heiss, A.G.; Wessjohann, L.A. Metabolomics driven analysis of six Nigella species seeds via UPLC-qTOF-MS and GC-MS coupled to chemometrics. Food Chem. 2014, 151, 333–342. [Google Scholar] [CrossRef]
- Farag, M.A.; Shakour, Z.T.; Lübken, T.; Frolov, A.; Wessjohann, L.A.; Mahrous, E. Unraveling the metabolome composition and its implication for Salvadora persica L. use as dental brush via a multiplex approach of NMR and LC-MS metabolomics. J. Pharm. Biomed. Anal. 2021, 193, 113727. [Google Scholar] [CrossRef] [PubMed]
- Wang, M.; Carver, J.J.; Phelan, V.V.; Sanchez, L.M.; Garg, N.; Peng, Y.; Nguyen, D.D.; Watrous, J.; Kapono, C.A.; Luzzatto-Knaan, T.; et al. Sharing and community curation of mass spectrometry data with Global Natural Products Social Molecular Networking. Nat. Biotechnol. 2016, 34, 828–837. [Google Scholar] [CrossRef]
- Zhang, Q.; Zhang, J.; Shen, J.; Silva, A.; Dennis, D.A.; Barrow, C.J. A simple 96-well microplate method for estimation of total polyphenol content in seaweeds. J. Appl. Phycol. 2006, 18, 445–450. [Google Scholar] [CrossRef]
- Hidalgo, M.; Sánchez-Moreno, C.; de Pascual-Teresa, S. Flavonoid–flavonoid interaction and its effect on their antioxidant activity. Food Chem. 2010, 121, 691–696. [Google Scholar] [CrossRef]
- Fernández-Poyatos, M.D.P.; Ruiz-Medina, A.; Zengin, G.; Llorent-Martínez, E.J. Phenolic characterization, antioxidant activity, and enzyme inhibitory properties of Berberis thunbergii DC. leaves: A valuable source of phenolic acids. Molecules 2019, 24, 4171. [Google Scholar] [CrossRef]
- Masike, K.; Mhlongo, M.I.; Mudau, S.P.; Nobela, O.; Ncube, E.N.; Tugizimana, F.; George, M.J.; Madala, N.E. Highlighting mass spectrometric fragmentation differences and similarities between hydroxycinnamoyl-quinic acids and hydroxycinnamoyl-isocitric acids. Chem. Cent. J. 2017, 11, 29. [Google Scholar] [CrossRef]
- Clifford, M.N.; Knight, S.; Surucu, B.; Kuhnert, N. Characterization by LC-MSn of four new classes of chlorogenic acids in green coffee beans: Dimethoxycinnamoylquinic acids, diferuloylquinic acids, caffeoyl-dimethoxycinnamoylquinic acids, and feruloyl-dimethoxycinnamoylquinic acids. J. Agric. Food Chem. 2006, 54, 1957–1969. [Google Scholar] [CrossRef]
- Bianco, G.; Abate, S.; Labella, C.; Cataldi, T.R. Identification and fragmentation pathways of caffeine metabolites in urine samples via liquid chromatography with positive electrospray ionization coupled to a hybrid quadrupole linear ion trap (LTQ) and Fourier transform ion cyclotron resonance mass spectrometry and tandem mass spectrometry. Rapid Commun. Mass Spectrom. RCM 2009, 23, 1065–1074. [Google Scholar] [CrossRef]
- Yashin, A.; Yashin, Y.; Xia, X.; Nemzer, B. Chromatographic Methods for Coffee Analysis: A Review. J. Food Res. 2017, 6, 60. [Google Scholar] [CrossRef]
- Ncube, E.N.; Mhlongo, M.I.; Piater, L.A.; Steenkamp, P.A.; Dubery, I.A.; Madala, N.E. Analyses of chlorogenic acids and related cinnamic acid derivatives from Nicotiana tabacum tissues with the aid of UPLC-QTOF-MS/MS based on the in-source collision-induced dissociation method. Chem. Cent. J. 2014, 8, 66. [Google Scholar] [CrossRef]
- Jaiswal, R.; Matei, M.F.; Subedi, P.; Kuhnert, N. Does roasted coffee contain chlorogenic acid lactones or/and cinnamoylshikimate esters? Food Res. Int. 2014, 61, 214–227. [Google Scholar] [CrossRef]
- Fang, N.; Yu, S.; Prior, R.L. LC/MS/MS characterization of phenolic constituents in dried plums. J. Agric. Food Chem. 2002, 50, 3579–3585. [Google Scholar] [CrossRef]
- Asamenew, G.; Kim, H.W.; Lee, M.K.; Lee, S.H.; Lee, S.; Cha, Y.S.; Lee, S.H.; Yoo, S.M.; Kim, J.B. Comprehensive characterization of hydroxycinnamoyl derivatives in green and roasted coffee beans: A new group of methyl hydroxycinnamoyl quinate. Food Chem. X 2019, 2, 100033. [Google Scholar] [CrossRef]
- Farah, A.; de Paulis, T.; Trugo, L.C.; Martin, P.R. Effect of roasting on the formation of chlorogenic acid lactones in coffee. J. Agric. Food Chem. 2005, 53, 1505–1513. [Google Scholar] [CrossRef]
- Jaiswal, R.; Patras, M.A.; Eravuchira, P.J.; Kuhnert, N. Profile and characterization of the chlorogenic acids in green Robusta coffee beans by LC-MS(n): Identification of seven new classes of compounds. J. Agric. Food Chem. 2010, 58, 8722–8737. [Google Scholar] [CrossRef]
- Panusa, A.; Petrucci, R.; Lavecchia, R.; Zuorro, A. UHPLC-PDA-ESI-TOF/MS metabolic profiling and antioxidant capacity of arabica and robusta coffee silverskin: Antioxidants vs phytotoxins. Food Res. Int. 2017, 99, 155–165. [Google Scholar] [CrossRef]
- Sun, X.; Zimmermann, M.L.; Campagne, J.-M.; Sneden, A.T. New Sucrose Phenylpropanoid Esters from Polygonum perfoliatum. J. Nat. Prod. 2000, 63, 1094–1097. [Google Scholar] [CrossRef]
- Panda, P.; Appalashetti, M.; Judeh, Z.M. Phenylpropanoid sucrose esters: Plant-derived natural products as potential leads for new therapeutics. Curr. Med. Chem. 2011, 18, 3234–3251. [Google Scholar] [CrossRef]
- Dias, R.C.; Faria, A.F.d.; Mercadante, A.Z.; Bragagnolo, N.; Benassi, M.d.T. Comparison of extraction methods for kahweol and cafestol analysis in roasted coffee. J. Braz. Chem. Soc. 2013, 24, 492–499. [Google Scholar] [CrossRef]
- Liu, M.; Wang, W.G.; Sun, H.-D.; Pu, J.-X. Diterpenoids from Isodon species: An update. Nat. Prod. Rep. 2017, 34, 1090–1140. [Google Scholar] [CrossRef] [PubMed]
- Lang, R.; Klade, S.; Beusch, A.; Dunkel, A.; Hofmann, T. Mozambioside is an arabica-specific bitter-tasting furokaurane glucoside in coffee beans. J. Agric. Food Chem. 2015, 63, 10492–10499. [Google Scholar] [CrossRef]
- Pérez-Míguez, R.; Sánchez-López, E.; Plaza, M.; Castro-Puyana, M.; Marina, M.L. A non-targeted metabolomic approach based on reversed-phase liquid chromatography–mass spectrometry to evaluate coffee roasting process. Anal. Bioanal. Chem. 2018, 410, 7859–7870. [Google Scholar] [CrossRef]
- Kurzrock, T.; Speer, K. Identification of kahweol fatty acid esters in Arabica coffee by means of LC/MS. J. Sep. Sci. 2001, 24, 843–848. [Google Scholar] [CrossRef]
- El-Hawary, S.S.; El-Kammar, H.A.; Farag, M.A.; Saleh, D.O.; El Dine, R.S. Metabolomic profiling of five Agave leaf taxa via UHPLC/PDA/ESI-MS inrelation to their anti-inflammatory, immunomodulatory and ulceroprotective activities. Steroids 2020, 160, 108648. [Google Scholar] [CrossRef]
- Farag, M.A.; Sallam, I.E.; Fekry, M.I.; Zaghloul, S.S.; El-Dine, R.S. Metabolite profiling of three Opuntia ficus-indica fruit cultivars using UPLC-QTOF-MS in relation to their antioxidant potential. Food Biosci. 2020, 36, 100673. [Google Scholar] [CrossRef]
- Speer, K.; Kölling-Speer, I. The lipid fraction of the coffee bean. Braz. J. Plant Physiol. 2006, 18, 201–216. [Google Scholar] [CrossRef]
- Lang, R.; Hofmann, T. A versatile method for the quantitative determination of β N-alkanoyl-5-hydroxytryptamides in roasted coffee. Eur. Food Res. Technol. 2005, 220, 638–643. [Google Scholar] [CrossRef]
- Berti, F.; Navarini, L.; Colomban, S.; Forzato, C. Hydroxycinnamoyl amino acids conjugates: A chiral pool to distinguish commercially exploited Coffea spp. Molecules 2020, 25, 1704. [Google Scholar] [CrossRef]
- Hegazi, N.M.; Khattab, A.R.; Frolov, A.; Wessjohann, L.A.; Farag, M.A. Authentication of saffron spice accessions from its common substitutes via a multiplex approach of UV/VIS fingerprints and UPLC/MS using molecular networking and chemometrics. Food Chem. 2022, 367, 130739. [Google Scholar] [CrossRef]
- Hegazi, N.M.; Saad, H.H.; Marzouk, M.M.; Abdel Rahman, M.F.; El Bishbishy, M.H.; Zayed, A.; Ulber, R.; Ezzat, S.M. Molecular Networking Leveraging the Secondary Metabolomes Space of Halophila stipulaceae (Forsk.) Aschers. and Thalassia hemprichii (Ehrenb. ex Solms) Asch. in Tandem with Their Chemosystematics and Antidiabetic Potentials. Mar. Drugs 2021, 19, 279. [Google Scholar] [CrossRef]
- Baky, M.H.; Badawy, M.T.; Bakr, A.F.; Hegazi, N.M.; Abdellatif, A.; Farag, M.A. Metabolome-based profiling of African baobab fruit (Adansonia digitata L.) using a multiplex approach of MS and NMR techniques in relation to its biological activity. RSC Adv. 2021, 11, 39680–39695. [Google Scholar] [CrossRef]
- Farag, M.A.; El-Ahmady, S.H.; Elian, F.S.; Wessjohann, L.A. Metabolomics driven analysis of artichoke leaf and its commercial products via UHPLC-q-TOF-MS and chemometrics. Phytochemistry 2013, 95, 177–187. [Google Scholar] [CrossRef]
- Amorim, J.L.; Lima, F.A.; Brand, A.L.M.; Cunha, S.; Rezende, C.M.; Fernandes, P.D. Two new (β)N-alkanoyl-5-hydroxytryptamides with relevant antinociceptive activity. Biomedicines 2021, 9, 455. [Google Scholar] [CrossRef]
- Andrzejewski, D.; Roach, J.A.G.; Gay, M.L.; Musser, S.M. Analysis of Coffee for the Presence of Acrylamide by LC-MS/MS. J. Agric. Food Chem. 2004, 52, 1996–2002. [Google Scholar] [CrossRef] [PubMed]
- Soares, C.; Cunha, S.; Fernandes, J. Determination of acrylamide in coffee and coffee products by GC-MS using an improved SPE clean-up. Food Addit. Contam. 2006, 23, 1276–1282. [Google Scholar] [CrossRef] [PubMed]
- Elguindy, N.M.; Yacout, G.A.; El Azab, E.F.; Maghraby, H.K. Chemoprotective effect of Elettaria cardamomum against chemically induced hepatocellular carcinoma in rats by inhibiting NF-κB, oxidative stress, and activity of ornithine decarboxylase. S. Afr. J. Bot. 2016, 105, 251–258. [Google Scholar] [CrossRef]
- Alfarhani, B. Analysis of acrylamide levels in various food types in the Iraqi markets using chromatography techniques. J. Mater. Environ. Sci. 2017, 8, 4902. [Google Scholar]
- Górecki, M.; Hallmann, E. The antioxidant content of coffee and its in vitro activity as an effect of its production method and roasting and brewing time. Antioxidants 2020, 9, 308. [Google Scholar] [CrossRef] [PubMed]
- Król, K.; Gantner, M.; Tatarak, A.; Hallmann, E. The content of polyphenols in coffee beans as roasting, origin and storage effect. Eur. Food Res. Technol. 2020, 246, 33–39. [Google Scholar] [CrossRef]
- Corso, M.P.; Vignoli, J.A.; Benassi, M.d.T. Development of an instant coffee enriched with chlorogenic acids. J. Food Sci. Technol. 2016, 53, 1380–1388. [Google Scholar] [CrossRef]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Sample Name | Supplier | Degree of Roasting * | Sample Code | |
|---|---|---|---|---|
| Authentic roasted samples | Roasted C. arabica | Mina Gerais, Brazil | 12.9 | RCA |
| Roasted C. canephora or C. robusta | 6.6 | RCC | ||
| Authentic green samples | Green C. arabica | 7.3 | GCA | |
| Green C. canephora or C. robusta | 1.6 | GCC | ||
| Commercial samples | Lightly roasted blended with cardamom | Maatouk, Saudi Arabia | 1.8 | LRCM |
| Lightly roasted coffee | Shahi, Saudi Arabia | 2.8 | LRS | |
| Heavily roasted blended with cardamom | Alameed coffee, Kuwait | 6.0 | HRKC | |
| Lab-roasted green coffee | 1.6 | BRK | ||
| Lightly roasted blended with cardamom | 1.0 | LRCK | ||
| Lab-roasted green coffee | Aswan, Egypt | 1.0 | BRA | |
| Lightly roasted blended with Qassim blend | Saudi Arabia | figure--- ** | LRSQ | |
| Lightly roasted blended with cardamom | Shahi, Saudi Arabia | 4.1 | LRCS | |
| Qatar | 4.2 | LRCQ | ||
| Instant C. arabica | Maxima coffee | 32.3 | ICA | |
| Instant Arabian coffee blended with cardamom | NESCAFE Arabiana | 7.1 | ICC | |
| Green coffee | Bayara, United Arab Emirates | 4.4 | GCU | |
| Aswan, Egypt | 3.5 | GCE | ||
| Saudi Arabia | 1.3 | GCS | ||
| Alameed coffee, Kuwait | 5.1 | GCK | ||
| Peak No. | Metabolite | Rt (Min) | UV Max (nm) | Mass Error (ppm) | Mol. Formula | [M − H]− (m/z) | [M + H]+ (m/z) | MSn Fragmentation Ions (m/z) | Method of Annotation | Coffee Specimen |
|---|---|---|---|---|---|---|---|---|---|---|
| Organic acids | ||||||||||
| P1 | Quinic acid * | 0.67 | 265 | −0.11 | C7H11O6− | 191.05589 | ** | 111, 173 | [24] | RCA RCC GCA GCC |
| P2 | Isocitric acid | 0.89 | 372 | −0.13 | C6H7O7− | 191.01965 | ** | 111, 173, 155, 127 | RCA RCC GCA GCC | |
| Phenolic acid glycosides | ||||||||||
| P3 | Unknown phenolic acid glycoside | 0.66 | 265 | 7.92 | C17H17O10+ | ** | 381.07895 | 219, 201 | RCA RCC GCA GCC | |
| P4 | Dihydroferulic acid 4-O-glucuronide | 0.94 | ** | −1.11 | C16H19O10− | 371.09756 | ** | 353, 191, 135 | [25] | GCC |
| Alkaloids | ||||||||||
| P5 | Trigonelline * | 1.37 | ** | −1.0 | C7H7NO2+ | ** | 138.08978 | 120, 110, 69, 90 | [26] | RCA RCC GCA GCC |
| P6 | Caffeine * | 3.54 | ** | −5.4 | C8H10N4O2+ | ** | 195.19037 | 137 | RCA RCC GCA GCC | |
| Hydroxycinnamate esters and lactones | ||||||||||
| P7 | 3-O-caffeoylquinic acid * | 1.9 | 221, 325 | −0.1 | C16H17O9− | 353.08777 | ** | 191, 179, 135 | [27,28] | RCA RCC GCA GCC |
| P8 | 5-O-caffeoylquinic acid * | 3.69 | 221, 325 | −1.06 | C16H17O9− | 353.08771 | ** | 191, 179 | RCA RCC GCA GCC | |
| P9 | 4-O-caffeoylquinic acid | 5.09 | 221,325 | −1.06 | C16H17O9− | 353.08844 | ** | 173, 179, 191 | RCA RCC GCA GCC | |
| P10 | Caffeoyl shikimic acid | 6.15 | 301,284 | −2.18 | C16H15O8− | 335.07687 | ** | 179, 161, 135 | [29] | RCA RCC |
| P11 | Feruloyl quinic acid isomer | 7.66 | 221,325 | −1.07 | C17H19O9− | 367.10324 | ** | 161, 193, 135 | [29,30] | RCA RCC GCA GCC |
| P12 | p-Coumaroyl quinic acid | 7.86 | 221,325 | −1.69 | C16H17O8− | 337.09256 | ** | 191, 163 | RCA RCC GCA GCC | |
| P13 | Caffeoyl-quinolactone | 8.99 | 221,325 | −1.55 | C16H15O8− | 335.07648 | ** | 161, 135, 179 | [29] | RCA RCC |
| P14 | Unknown chlorogenic acid derivative | 9.66 | 218,322 | −0.76 | C24H25O11− | 489.13986 | ** | 353, 315, 255, 191, 297 | **** | RCC |
| P15 | Unknown diacyl chlorogenic acid derivative | 9.7 | 220,320 | −2.3 | C21H25O11− | 453.13919 | ** | 353, 335, 291 | **** | GCC |
| P16 | Methyl-O-feruloyl quinic acid | 10.01 | 218,326 | −2.06 | C18H21O9− | 381.11981 | ** | 175, 160, 193 | [31] | RCC RCA |
| P17 | Feruloyl-quinolactone | 10.15 | 218,326 | −0.51 | C17H17O8− | 349.09271 | ** | 175, 193, 149, 134 | [32] | RCC RCA |
| P18 | Dicaffeoyl-quinolactone ** | 10.19 | 325 | −0.27 | C25H23O11− | 497.1075 | ** | 335 | **** | RCA RCC |
| P19 | Dicaffeoyl quinic acid * | 10.23 | 220,325 | −1.76 | C25H23O12− | 515.11859 | ** | 353, 335 | [30] | RCA RCC GCA GCC |
| P20 | Caffeoyl-feruloylquinic acid | 10.82 | 325 | −1.75 | C26H25O12− | 529.13428 | ** | 367, 353 | [25,31] | RCA RCC GCA GCC |
| P21 | Caffeoyl-feruloyl quinolactone *** | 11.74 | 220,325 | −1.8 | C26H23O11− | 511.12366 | ** | 335, 179.161 | [31] | RCC |
| P22 | Sinapoyl-feruoylquinic acid | 11.28 | 221,324 | −1.57 | C28H29O13− | 573.16046 | ** | 349, 397 | GCA GCC | |
| P23 | Di-feruloylquinic acid | 11.31 | 221,324 | −1.87 | C27H27O12− | 543.1499 | ** | 367, 349 | [25,33,34] | RCC GCC |
| P24 | Caffeoyl-dimethoxy cinnamoylquinic acid | 11.30 | 222,324 | −1.57 | C27H27O12− | 543.1500 | ** | 381.367, 335 | GCA | |
| P25 | Unknown quinolactone derivative | 11.6 | 221 | −2.37 | C25H21O10− | 481.11288 | ** | 335, 179, 161 | **** | RCC RCA |
| P26 | Feruloyl -dimethoxycinnamoylquinic acid | 11.75 | 222 | 1.62 | C28H29O12− | 557.1684 | ** | 381, 349 | [25,33,34] | GCA GCC |
| P27 | Triacyl-O-caffeoyl-O-feruloyl-O-sinapoylquinic acid *** | 11.9 | 222 | −1.91 | C37H35O16− | 735.19165 | ** | 573, 529 | [31,33] | GCC |
| P28 | Di-O-feruolyl-O-caffeoylquinic acid | 11.98 | 222 | −3.9 | C36H33O15− | 705.17969 | ** | 543, 529 | GCC | |
| P29 | Caffeoyl-feruloylquinic acid lactone | 12.1 | 222 | −2.13 | C26H23O11− | 437.14474 | ** | 335, 193, 179 | **** | RCC |
| P30 | Unknown quinolactone derivative | 12.23 | 223 | −2.8 | C21H23O9− | 419.13358 | ** | 335, 317, 255, 179 | RCA RCC | |
| P31 | Unknown chlorogenic acid | 13.15 | 222 | −1.23 | C21H25O10− | 437.1444 | ** | 173, 275 | GCA GCC | |
| Sugars and sugar derivatives | ||||||||||
| P32 | Di-O-hexoside | 0.7 | ** | −0.3 | C12H21O11− | 341.10883 | ** | ** | GCA GCC | |
| P33 | Acetyl-diferuloyl sucrose | 8.85 | 221,325 | −1.01 | C34H39O18− | 735.21344 | ** | 367 | [35,36] | GCC GCA |
| P34 | Acyl sucroses dihydroxycinnamoyl | 9.73 | 220,327 | −0.34 | C29H36O18− | 671.29065 | ** | 627 | GCA | |
| Diterpenes | ||||||||||
| P35 | Cafestol | 9.39 | 222 | −1.54 | C20H29O3+ | ** | 317.21063 | 299, 271, 253 | [37] | GCA |
| P36 | Trihydroxy-kauradienolide *** | 9.45 | 217 | −2.1 | C20H27O5+ | ** | 347.18457 | 329, 285 | [38] | GCA |
| P37 | Dehydrocafestol | 9.49 | 220 | −1.32 | C20H27O2+ | ** | 299.20016 | 145, 191, 281, 253 | [37] | GCA RCA |
| P38 | Mozambioside | 10.03 | 298 | −1.38 | C26H37O10+ | ** | 509.224 | 347, 329, 311 | [39,40] | RCA GCA |
| P39 | Bengalensol-O-hexoside | 11.75 | 221 | −1.38 | C26H35O9+ | ** | 491.2417 | 329, 311 | **** | RCA |
| P40 | Trihydroxy-kauranoic acid | 10.67 | 220 | −2.21 | C20H31O5− | 351.2171 | ** | 289, 321 | [38] | RCA |
| P41 | Bengalensol | 11.63 | 221 | −1.35 | C20H25O4+ | ** | 329.17429 | 293, 311, 237 | **** | RCA |
| P42 | Dihydroxy-kauren-oic acid | 12.64 | 223 | −0.218 | C20H29O4− | 333.20706 | ** | 303 | RCA | |
| P43 | 16-methyl kahweol | 12.78 | 222 | −1.79 | C20H27O4+ | ** | 331.19006 | 314, 296, 145, 279 | [37] | RCC RCA |
| P44 | Dehydro-kahweol | 13.6 | 222 | −1.33 | C20H25O2+ | ** | 297.18451 | 279, 145 | GCA | |
| P45 | Dehydrocafestol derivative | 15.28 | 225 | −1.62 | C20H25O+ | ** | 281.18954 | 263, 173, 131 | [41] | RCA RCC |
| P46 | Carboxyatractyligenin-O-hexoside | 9.57 | 324,221 | −1.74 | C26H37O11− | 525.23322 | ** | 396, 203 | GCA GCC | |
| P47 | Atracyligenin-O-hexoside | 9.82 | 219,311 | −1.1 | C25H37O9− | 481.24377 | ** | 301 | [34] | GCA GCC |
| P48 | Desoxycarboxyatractyligenin-O-hexoside | 10.5 | 220 | 0.05 | C37H55O17− | 771.34338 | ** | 727 | **** | GCA |
| P49 | Desoxyatractyligenin-O-hexoside | 11.21 | ** | 0.45 | C36H55O15− | 727.35376 | ** | 643, 625 | RCA GCA | |
| P50 | Carboxyatractyligenin-O-hexoside | 11.72 | 221 | −1.64 | C31H45O12− | 609.29065 | ** | 565 | GCA | |
| P51 | Isovaleryl- atractyligenin-O-hexoside derivative | 11.82 | 221 | −1.23 | C30H45O10− | 565.3009 | ** | 481, 463, 303 | RCA GCA | |
| Fatty acids and sphingolipids | ||||||||||
| P52 | Trihydroxy-octadecaenoic acid | 12.2 | 223 | −1.87 | C18H33O5− | 329.23273 | ** | 311, 293, 229, 171 | [11,42] | RCA GCA |
| P53 | Hexosyl-2-(pentanoyloxy) propyl dodecenoate | 13.03 | 223 | −1.29 | C26H45O10− | 517.302 | ** | 473, 367 | RCC | |
| P54 | Linoleic acid methyl ester *** | 13.19 | 223 | −1.47 | C17H25O4− | 293.17523 | ** | 236, 221 | [11] | RCC GCC |
| P55 | Unknown fatty acid | 13.6 | 222 | 4.18 | C14H29O8− | 325.18228 | ** | 183 | RCA RCC GCA GCC | |
| P56 | Sphingolipid conjugate I *** | 14.00 | 222 | 1.39 | C27H49NO9P− | 562.31610 | ** | 502 | **** | RCA RCC GCA GCC |
| P57 | Sphingolipid conjugate II *** | 14.55 | 222 | −2.18 | C27H51NO9P− | 564.32947 | ** | 504 | RCA RCC GCA GCC | |
| P58 | Phosphatidyl inositol hexanoic acid derivative | 14.68 | 224 | −3.21 | C25H48O12P− | 571.28705 | ** | 391, 315, 255, 241 | [11] | RCA RCC GCA GCC |
| P59 | Ceramide conjugate I *** | 14.75 | 224 | −0.45 | C22H49O6N4P+ | ** | 496.33881 | 478, 184 | RCA RCC GCA GCC | |
| P60 | Sphingolipid conjugate III *** | 14.78 | 222 | −1.9 | C25H51NO9P− | 540.32941 | ** | 480 | **** | RCA RCC GCA GCC |
| P61 | Unknown fatty acid | 14.85 | 224 | −7.34 | C13H27O8− | 311.16809 | ** | 183 | RCA RCC GCA GCC | |
| P62 | Sphingolipid conjugate IV *** | 14.97 | 224 | −1.45 | C27H53NO9P− | 566.34552 | ** | 506 | **** | RCA RCC GCA GCC |
| P63 | Ceramide conjugate II | 15.08 | 221 | −0.98 | C21H45O2N9P2+ | ** | 522.35461 | 504, 184 | RCA RCC GCA GCC | |
| P64 | Unknown fatty acid ester | 15.48 | 222 | 8.46 | C24H51O10− | 499.35391 | ** | 481, 455, 322, 279 | RCA RCC GCA GCC | |
| P65 | Ceramide conjugate III *** | 15.7 | 222 | −1.42 | C23 H46O8 N5+ | ** | 520.37042 | 502, 184 | RCC GCC GCC GCA | |
| P66 | Unknown hydroxy fatty acid | 16.22 | 225 | −8.13 | C28H31O3− | 415.22482 | ** | 279 | RCC | |
| P67 | Unknown fatty acid | 16.3 | 225 | −8.08 | C26H55O10− | 527.38416 | ** | 509, 350, 307 | RCC RCA | |
| P68 | Unknown fatty acid | 16.49 | 225 | −9.23 | C15H31O8− | 339.19919 | ** | 183 | RCA RCC GCA GCC | |
| P69 | Dimethyl octadecanedioate | 19.18 | 226 | 0.83 | C20H37O4− | 341.26892 | ** | 313, 269 | [11] | RCA RCC GCA GCC |
| P70 | Hydroxy-docosanoic acid | 19.35 | 224 | −2.97 | C22H43O3− | 355.32135 | ** | 309 | RCA RCC GCA GCC | |
| P71 | Hydroxy-tetracosanoic acid | 20.58 | 222 | −2.5 | C24H47O3− | 383.35208 | ** | 337 | RCA RCC GCA GCC | |
| P72 | Unknown fatty acid ester | 20.97 | 227 | −0.03 | C38H55O3− | 559.41382 | ** | 541, 279, 223, 183 | RCC GCC | |
| Fatty acyl amides | ||||||||||
| P73 | Unknown fatty acid amide | 15.77 | 224 | −0.06 | C20H38O2N+ | ** | 324.28943 | 307, 263, 245 | RCC RCA | |
| P74 | Docosenamide *** | 16.69 | 226 | −1.06 | C22H44NO+ | ** | 338.34137 | 321,303 | [43] | RCA RCC GCC GCA |
| Nitrogenous compounds (hydroxytryptamine derivatives) | ||||||||||
| P75 | N-heneicosanoyl- hydroxytryptamine | 15.92 | 226 | −1.4 | C30H51N2 O3+ | ** | 487.39 | 469, 177, 160 | RCA GCA RCC GCC | |
| P76 | N-tricosanoyl-hydroxytryptamine | 16.69 | 226 | −0.65 | C23H55N2O3+ | ** | 515.42 | 497, 177, 160 | [44,45] | RCA RCC GCA GCC |
| P77 | N-docosanoyl-hydroxytryptamide | 19.3 | 226 | −1.09 | C32H55N2O5+ | ** | 499.42 | 482, 177, 160 | RCA RCC GCA GCC | |
| P78 | N-octadecanoyl-5-hydroxytryptamide | 17.46 | 226 | −1.81 | C28H46N2O2+ | ** | 443.36185 | 426, 177, 160 | RCA RCC GCA GCC | |
| P79 | N-eicosanoyl-hydroxytryptamide | 18.32 | 226 | −1.68 | C30H51N2O2+ | ** | 471.39349 | 454, 177, 160 | RCA RCC GCA GCC | |
| P80 | N-tetracosanoyl-hydroxytryptamide | 20.32 | 226 | −0.95 | C34H59N2O2+ | ** | 527.46 | 510,177,160 | RCA RCC GCA GCC | |
| Hydroxycinnamoyl amides | ||||||||||
| P81 | Unknown caffeoyl amide | 9.8 | 220 | 9.75 | C18H16NO6− | 342.09775 | ** | 206 | [25] | GCA GCC |
| P82 | Caffeoyl-N- tryptophan | 11.08 | 221 | −0.61 | C20H17N2O5− | 365.11395 | ** | 135,229 | [25,46] | GCC RCC |
| P83 | p-Coumaroyl-N-tryptophan | 11.49 | 221 | −2.0 | C20H17N2O4− | 349.11868 | ** | 229 | RCC GCC | |
| P84 | Unknown feruloyl amide | 11.66 | 222 | 9.14 | C21H19N2O5− | 379.12805 | ** | 335, 229 | GCC | |
| P85 | Unknown hydroxy cinnamic acid amide | 12.1 | 229 | −6.59 | C25H21O6N2− | 445.1405 | ** | 309, 161, 229 | **** | RCA RCC |
| Unknowns | ||||||||||
| P86 | Unknown | 9.69 | 222 | −1.07 | C19H25O2+ | ** | 285.18472 | 267, 239, 229 | RCA | |
| P87 | 11.81 | 222 | −0.64 | C22H27O10− | 451.15973 | ** | 349, 275, 173 | GCA GCC | ||
| P88 | 15.3 | 222 | −2.3 | C28H33O4− | 433.23483 | ** | 153 | GCA | ||
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
El-Hawary, E.A.; Zayed, A.; Laub, A.; Modolo, L.V.; Wessjohann, L.; Farag, M.A. How Does LC/MS Compare to UV in Coffee Authentication and Determination of Antioxidant Effects? Brazilian and Middle Eastern Coffee as Case Studies. Antioxidants 2022, 11, 131. https://doi.org/10.3390/antiox11010131
El-Hawary EA, Zayed A, Laub A, Modolo LV, Wessjohann L, Farag MA. How Does LC/MS Compare to UV in Coffee Authentication and Determination of Antioxidant Effects? Brazilian and Middle Eastern Coffee as Case Studies. Antioxidants. 2022; 11(1):131. https://doi.org/10.3390/antiox11010131
Chicago/Turabian StyleEl-Hawary, Enas A., Ahmed Zayed, Annegret Laub, Luzia V. Modolo, Ludger Wessjohann, and Mohamed A. Farag. 2022. "How Does LC/MS Compare to UV in Coffee Authentication and Determination of Antioxidant Effects? Brazilian and Middle Eastern Coffee as Case Studies" Antioxidants 11, no. 1: 131. https://doi.org/10.3390/antiox11010131
APA StyleEl-Hawary, E. A., Zayed, A., Laub, A., Modolo, L. V., Wessjohann, L., & Farag, M. A. (2022). How Does LC/MS Compare to UV in Coffee Authentication and Determination of Antioxidant Effects? Brazilian and Middle Eastern Coffee as Case Studies. Antioxidants, 11(1), 131. https://doi.org/10.3390/antiox11010131