
Effects of Water Deficit Irrigation on Phenolic Composition and Antioxidant Activity of Monastrell Grapes under Semiarid Conditions


 ,
,
Abstract
:1. Introduction
2. Materials and Methods
2.1. Vineyard Conditions and Grape Sampling
2.2. Extraction of Grape Phenolic Compounds and Determination of Phenolic Composition by HPLC
2.3. Determination of Total Phenolic Content (TPC)
2.4. Determination of Free Radical Scavenging
2.4.1. DPPH Assay
2.4.2. ORAC Assay
2.4.3. TEAC Assay
2.5. Statistical Analysis
3. Results and Discussion
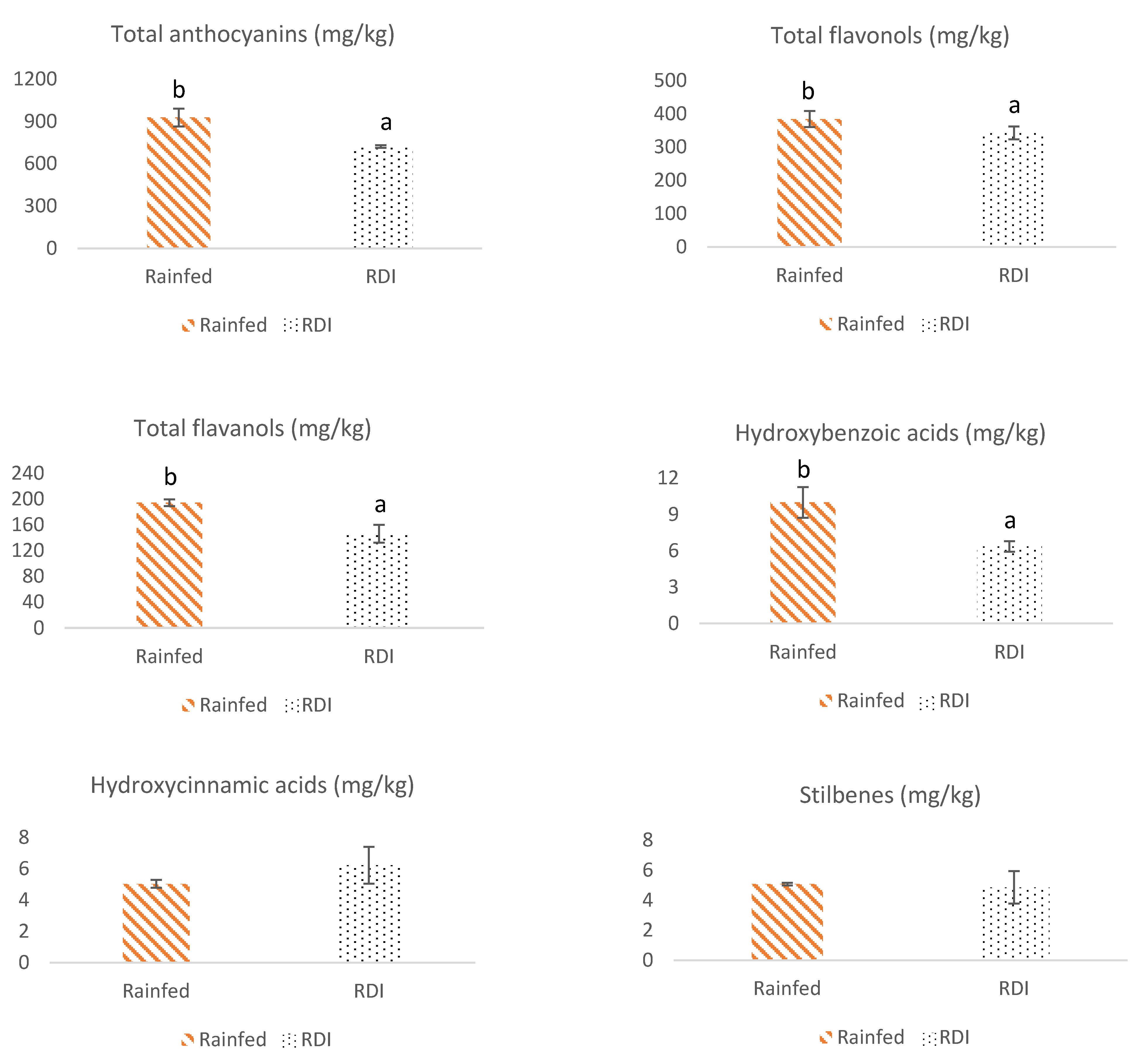
3.1. Flavonoid Composition of Monastrell Grapes from Grapevines under Non-Irrigated (Rainfed) and Regulated Deficit Irrigation (RDI) Watering Regimes
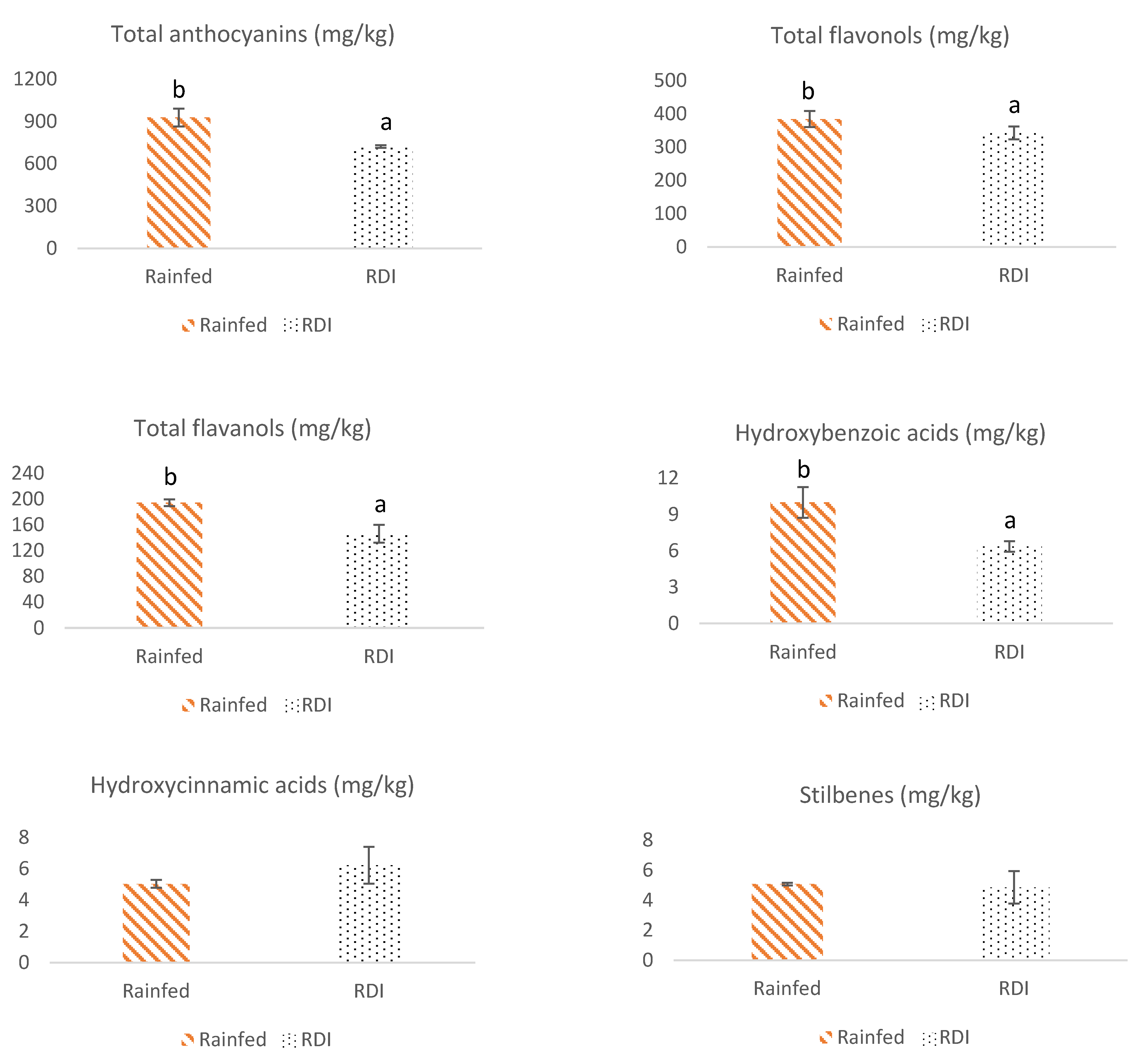
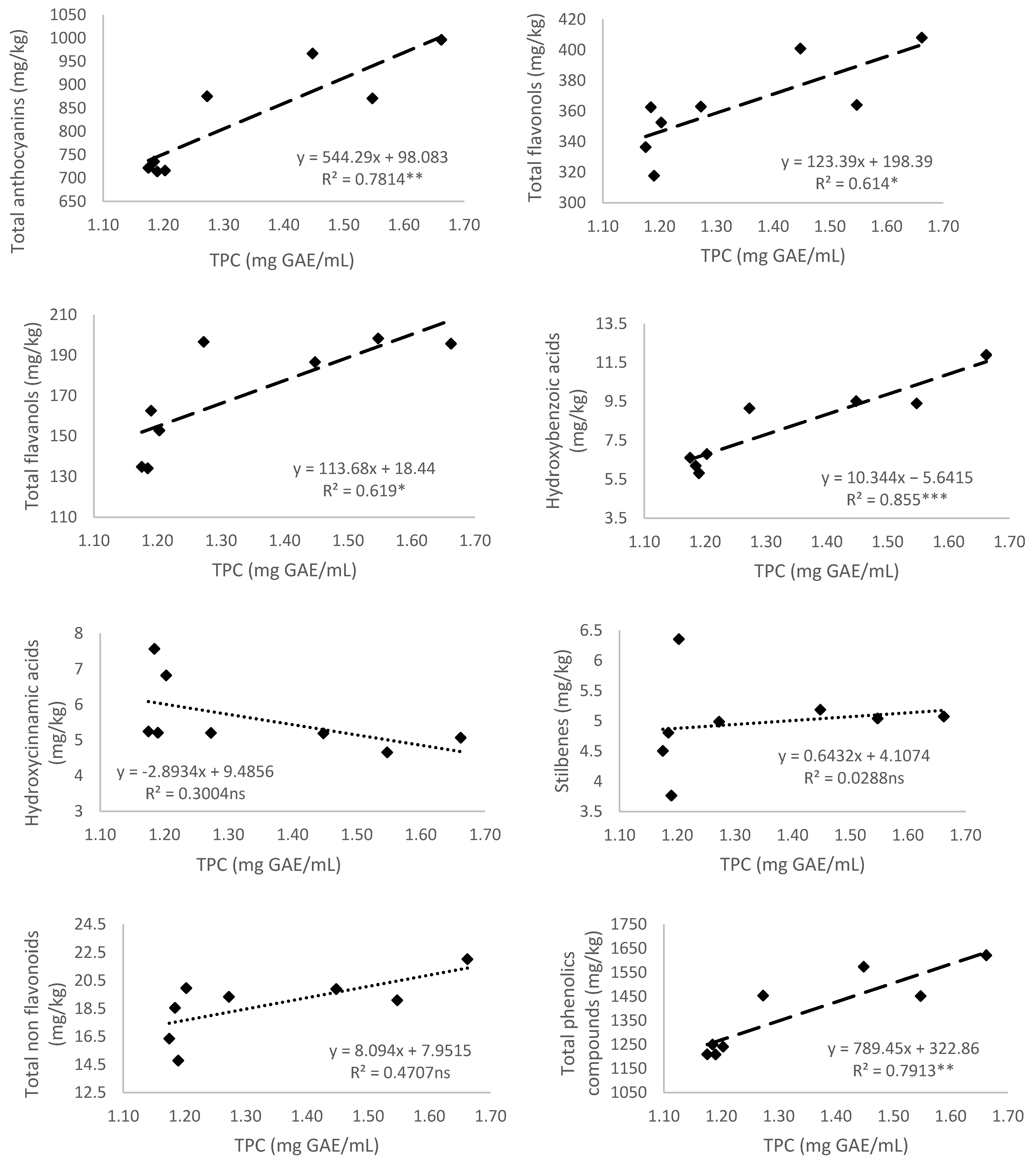
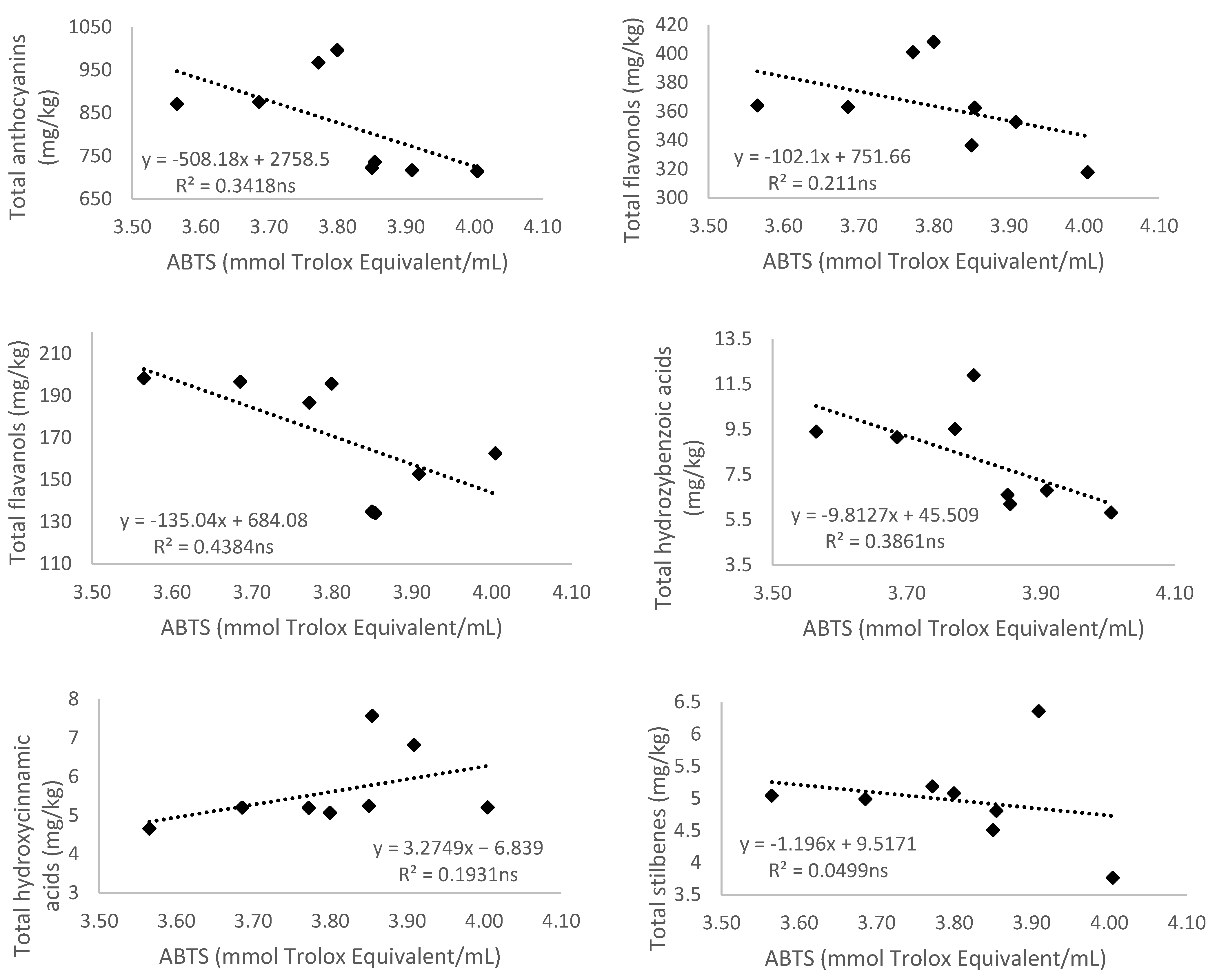
3.1.1. Anthocyanins
3.1.2. Flavonols
3.1.3. Flavanols
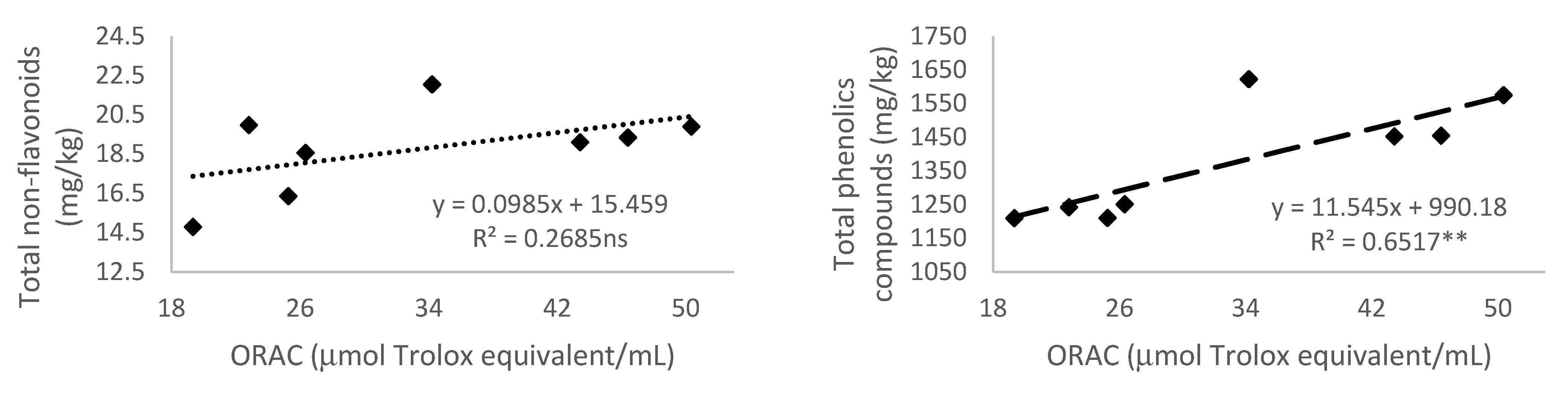
3.2. Non-Flavonoid Composition of Monastrell Grapes from Grapevines under Non-Irrigated (Rainfed) and Regulated Deficit Irrigation (RDI) Watering Regimes
3.3. Total Phenolic Compounds
3.4. Antioxidant Capacity
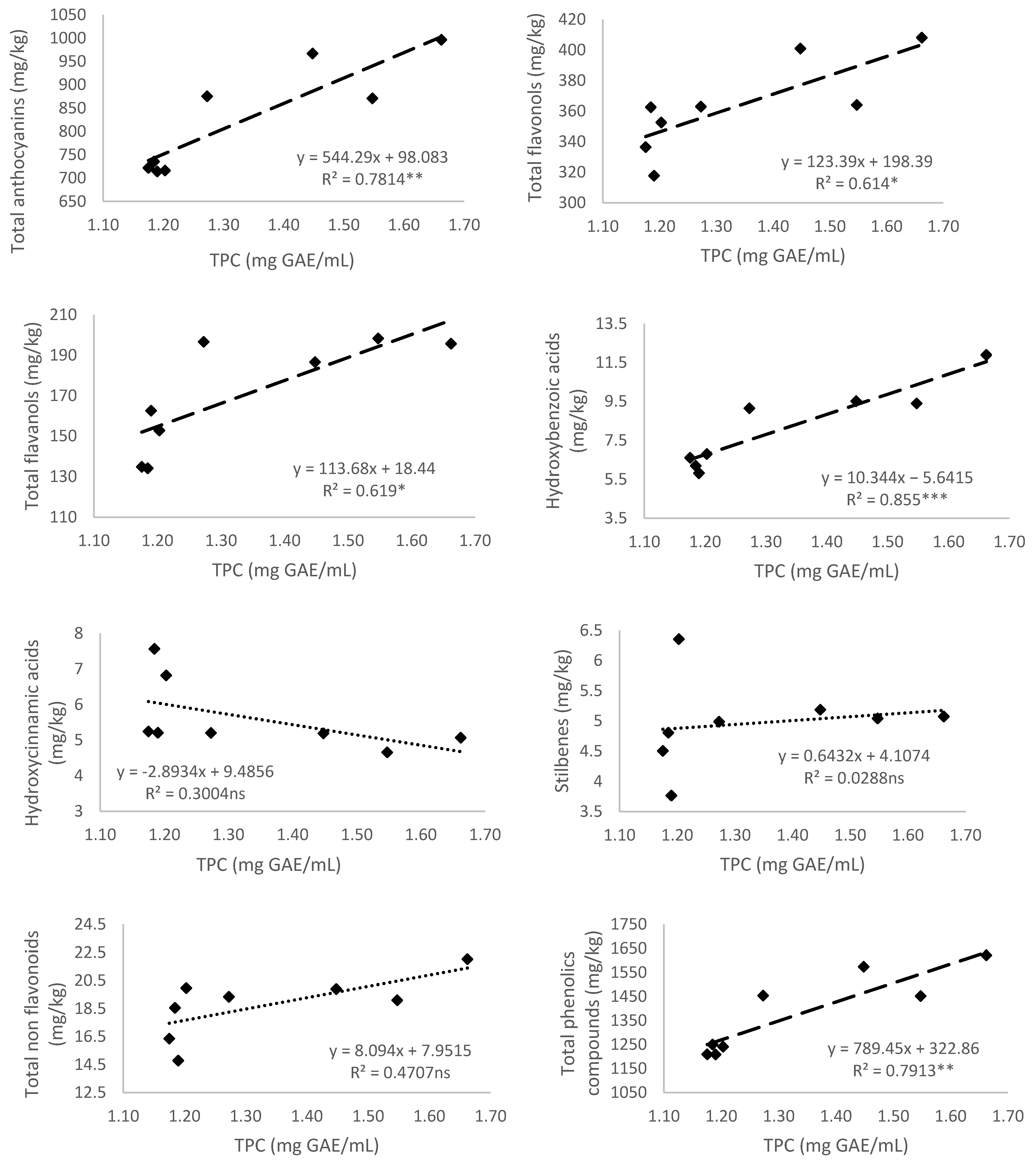
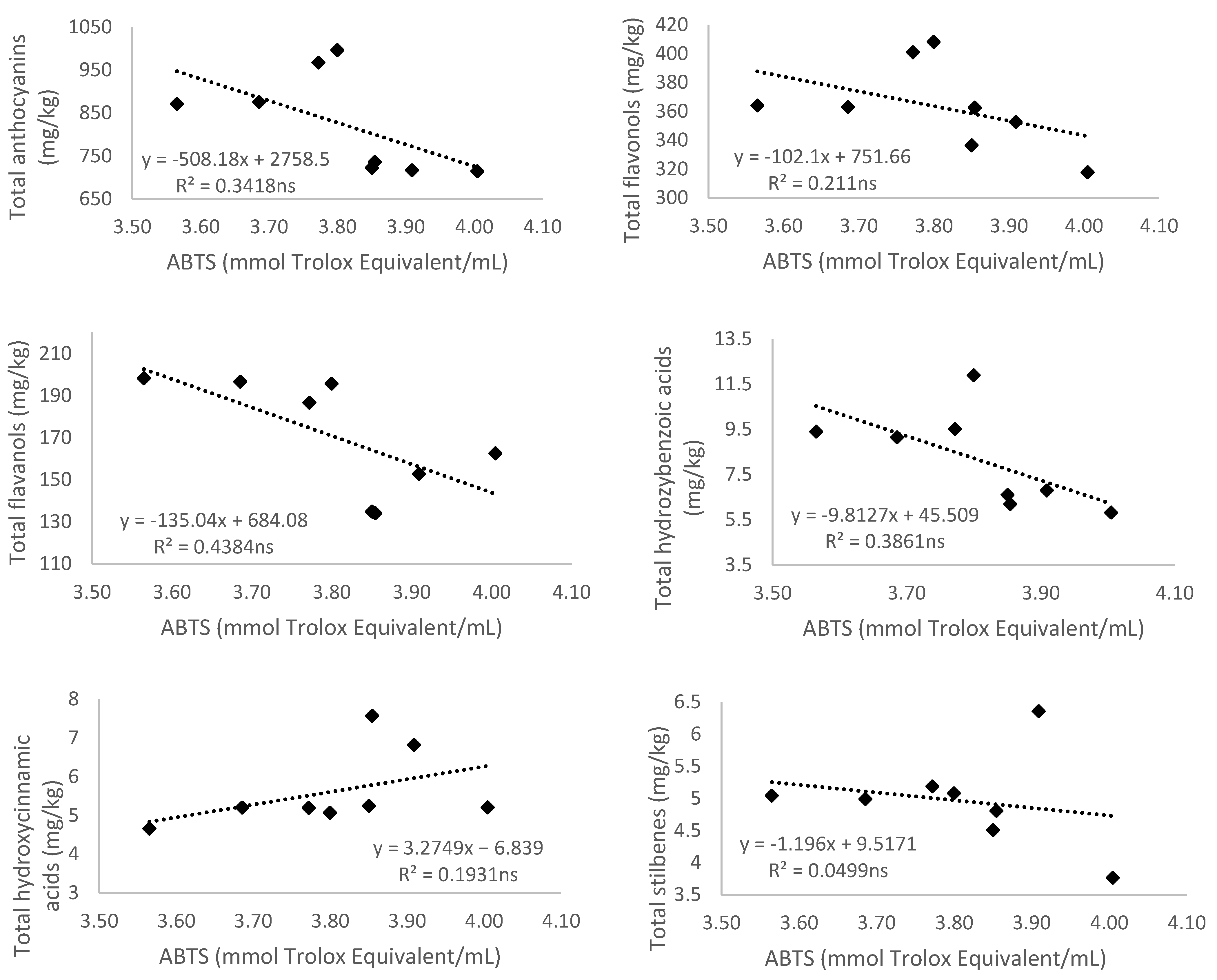
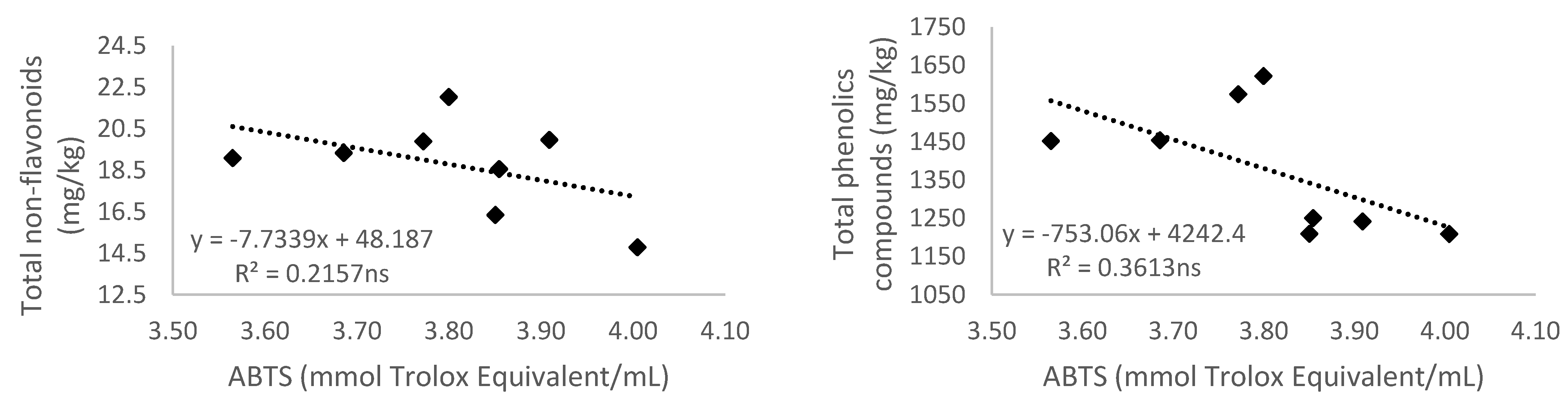
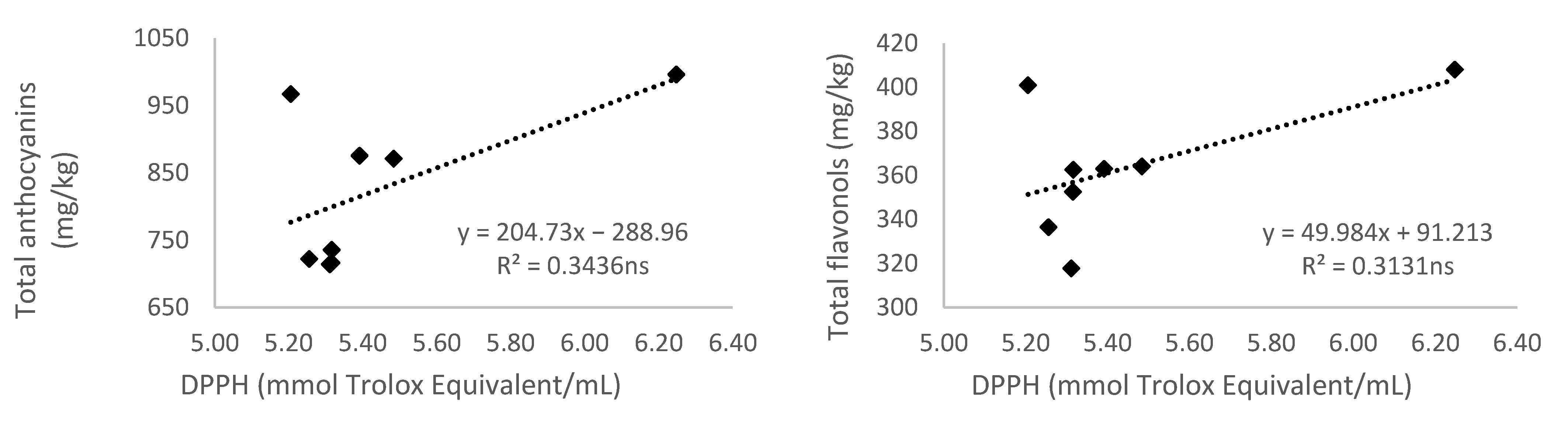
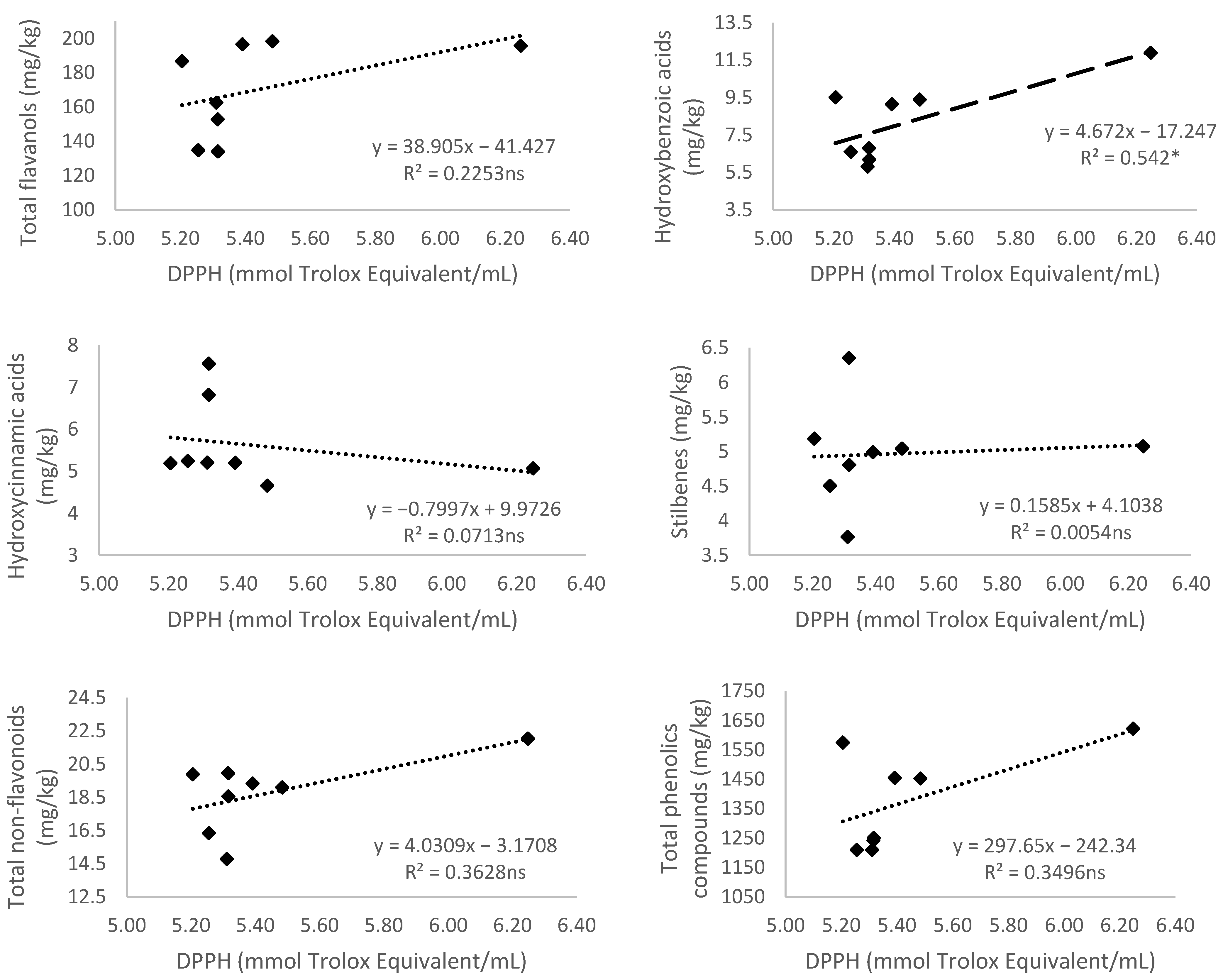
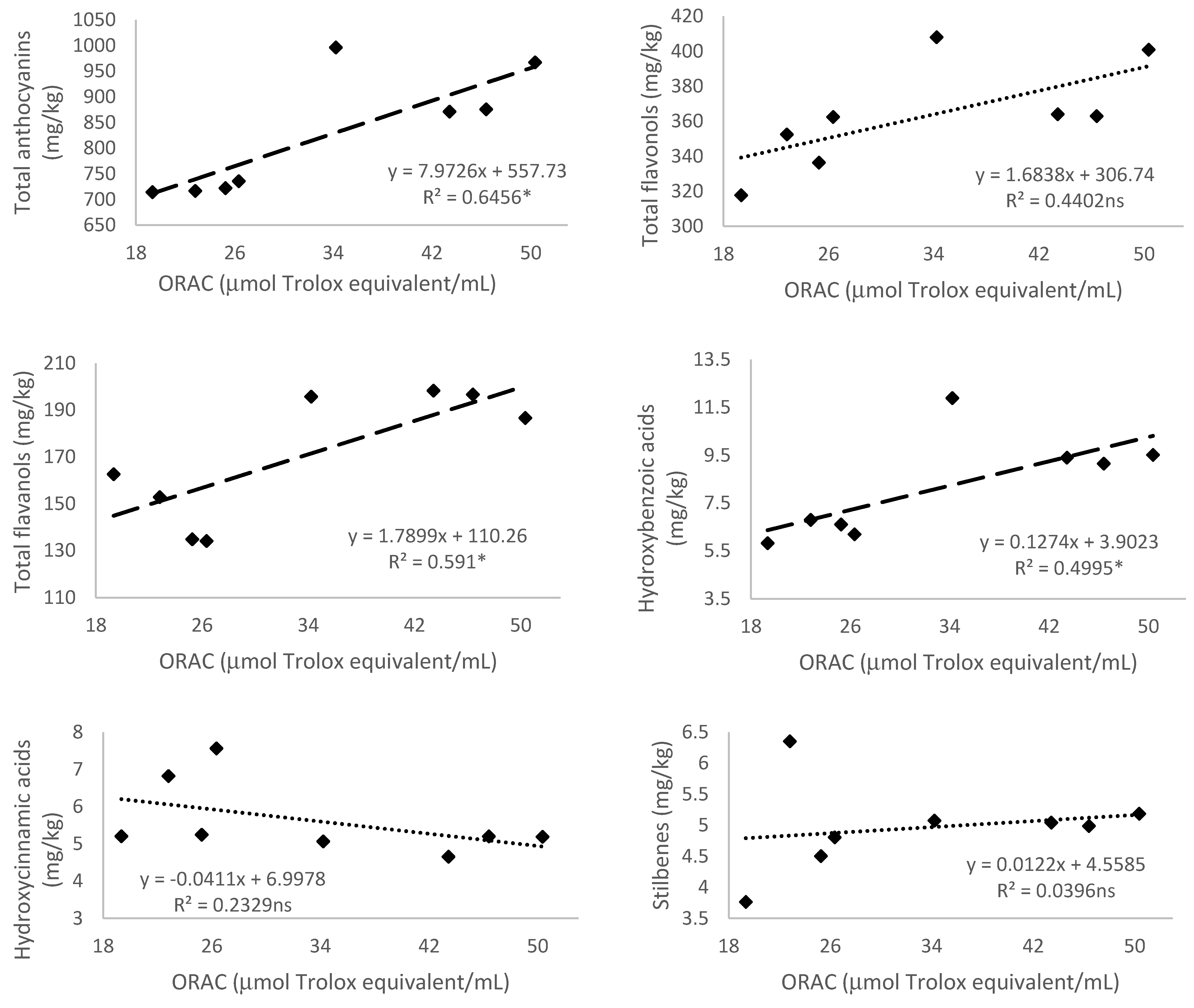
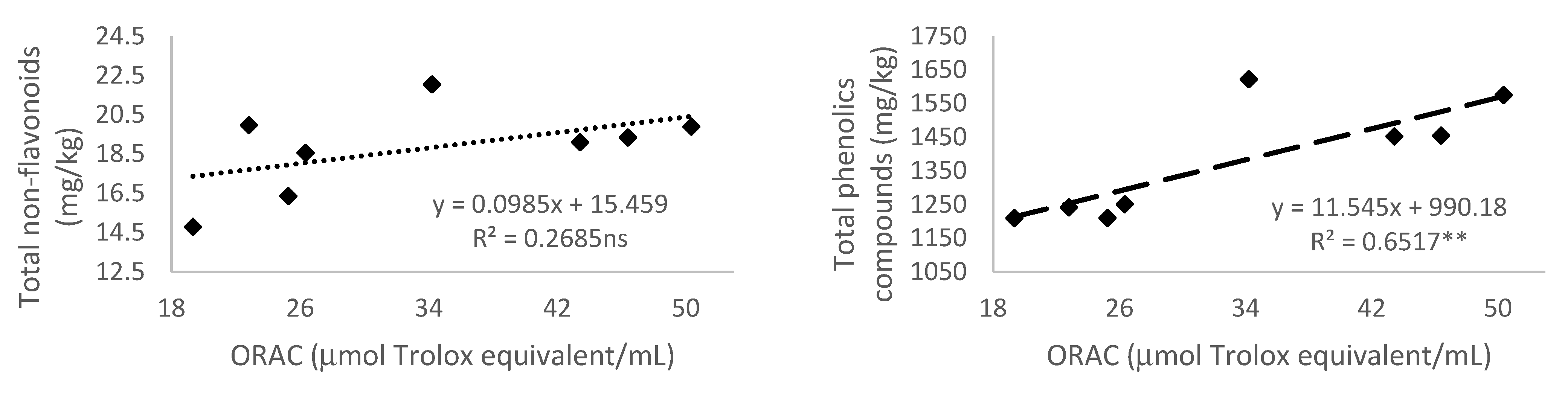
3.5. Relationship of Total Phenolic Content and Different Phenolic Groups According to Their Antioxidant Activity
3.5.1. Total Polyphenol Content
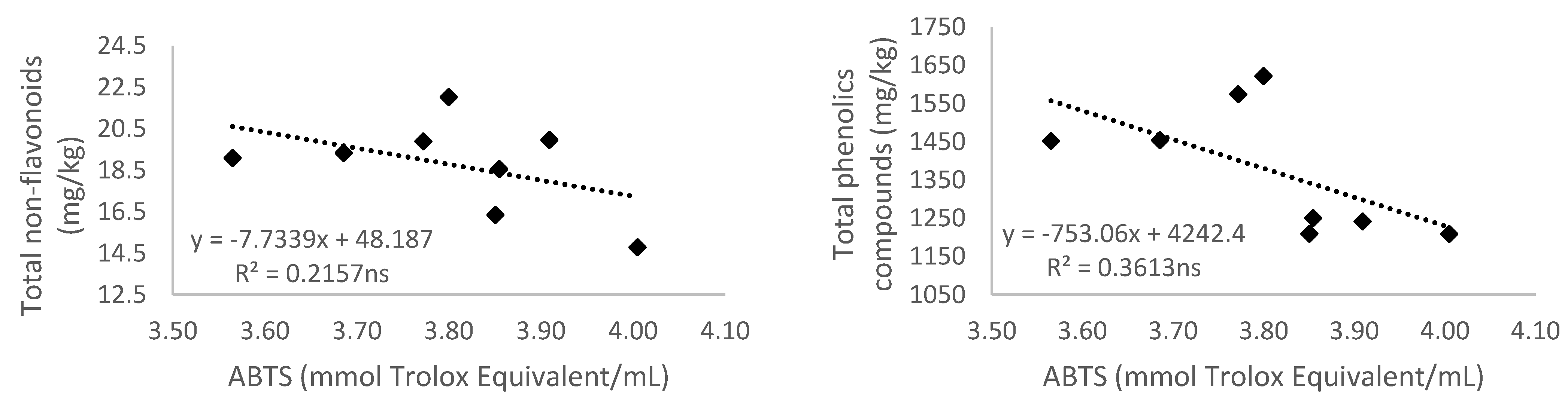
3.5.2. Antioxidant Activity Measured by 2,2′-Azino-bis(3-ethylbenzthiazoline-6-sulfonic acid) (ABTS)
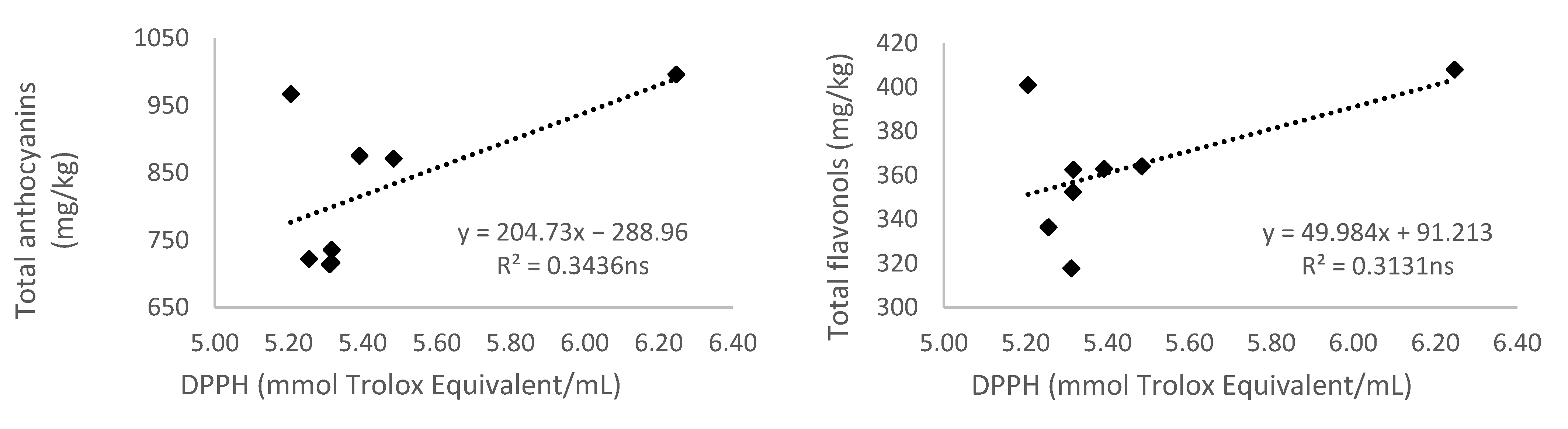
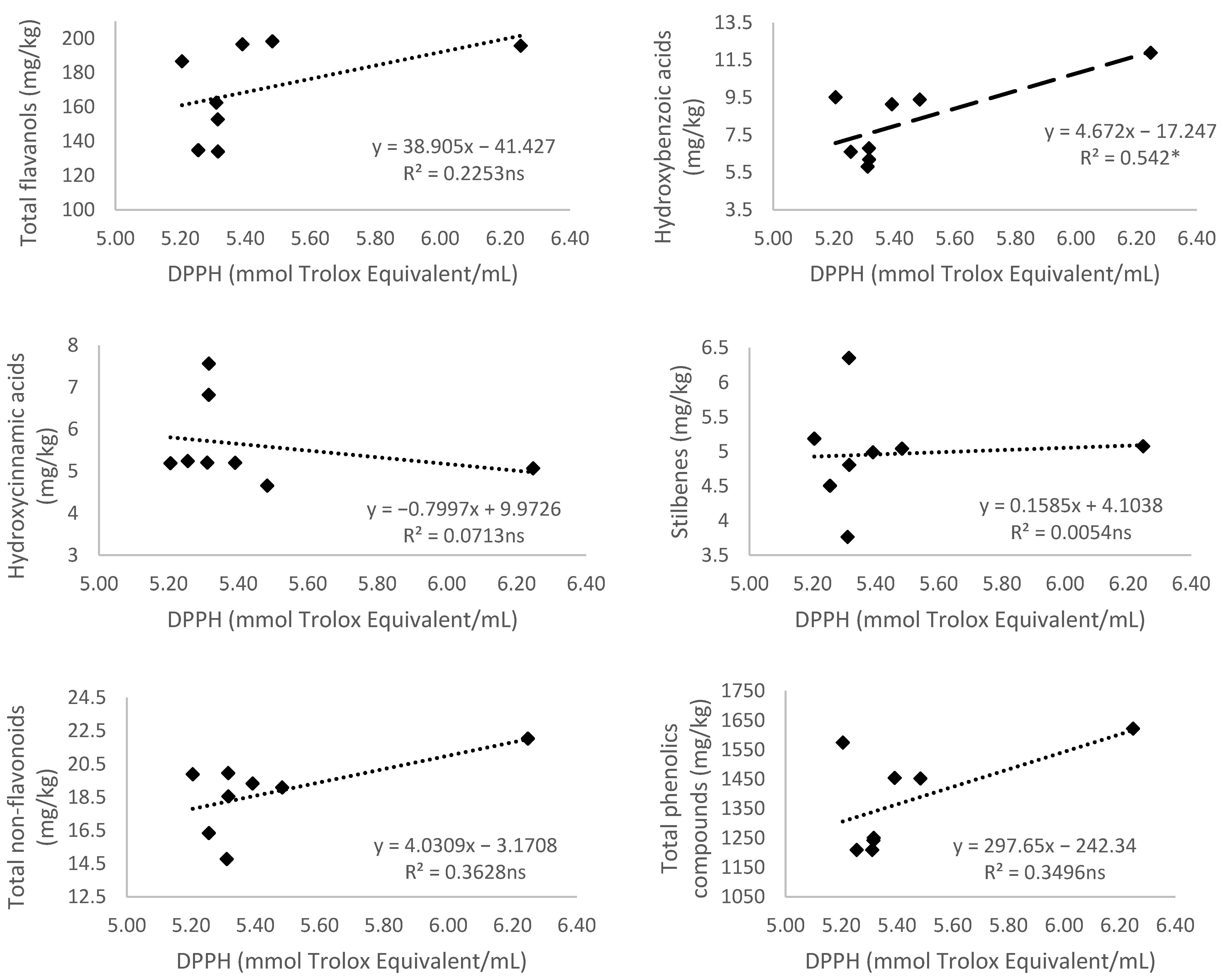
3.5.3. Antioxidant Activity Measured by 2,2-diphenyl-1-picryl-hydrazyl-hydrate (DPPH)
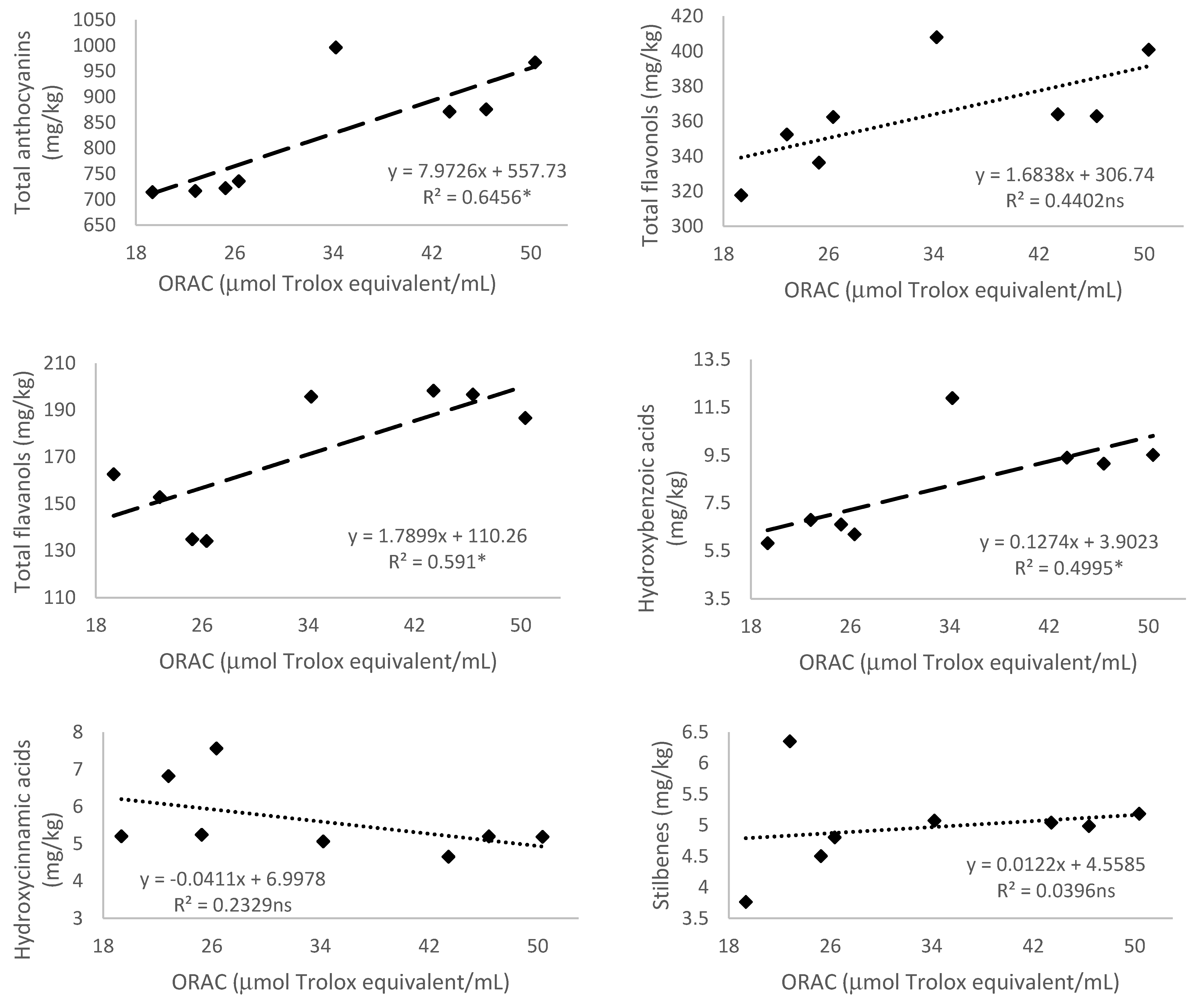
3.5.4. Antioxidant Activity Measured by 2,2′-Azobis(2-amidinopropane) dihydrochloride (AAPH)—Oxygen Radical Absorbance Capacity (ORAC)
4. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Li, L.; Sun, B. Grape and wine polymeric polyphenols: Their importance in enology. Crit. Rev. Food Sci. Nutr. 2017, 59, 563–579. [Google Scholar] [CrossRef] [PubMed]
- Elejalde, E.; Villarán, M.C.; Alonso, R.M. Grape polyphenols supplementation for exercise-induced oxidative stress. J. Int. Soc. Sports Nutr. 2021, 18, 1–12. [Google Scholar] [CrossRef] [PubMed]
- Luo, J.; Mills, K.; Le Cessie, S.; Noordam, R.; Van Heemst, D. Ageing, age-related diseases and oxidative stress: What to do next? Ageing Res. Rev. 2019, 57, 100982. [Google Scholar] [CrossRef] [PubMed]
- Poprac, P.; Jomova, K.; Simunkova, M.; Kollar, V.; Rhodes, C.J.; Valko, M. Targeting Free Radicals in Oxidative Stress-Related Human Diseases. Trends Pharmacol. Sci. 2017, 38, 592–607. [Google Scholar] [CrossRef] [PubMed]
- Sureda, A.; Tejada, S.; Bibiloni, M.D.M.; Tur, J.A.; Pons, A. Polyphenols: Well beyond the antioxidant capacity: Polyphenol supplementation and exercise-induced oxidative stress and inflammation. Curr. Pharm. Biotechnol. 2014, 15, 373–379. [Google Scholar] [CrossRef]
- García-Flores, L.A.; Medina, S.; Gómez, C.; Wheelock, C.E.; Cejuela, R.; Martínez-Sanz, J.M.; Oger, C.; Galano, J.-M.; Durand, T.; Hernández-Sáez, A.; et al. Aronia—Citrusjuice (polyphenol-rich juice) intake and elite triathlon training: A lipidomic approach using representative oxylipins in urine. Food Funct. 2017, 9, 463–475. [Google Scholar] [CrossRef] [Green Version]
- Portu, J.; Santamaría, P.; Alfaro, I.L.; López, R.; Garde-Cerdán, T. Methyl Jasmonate Foliar Application to Tempranillo Vineyard Improved Grape and Wine Phenolic Content. J. Agric. Food Chem. 2015, 63, 2328–2337. [Google Scholar] [CrossRef]
- Fanzone, M.; Zamora, F.; Jofré, V.; Assof, M.; Neira, P. Phenolic composition of Malbec grape skins and seeds from Valle de Uco (Mendoza, Argentina) during ripening. Effect of cluster thinning. J. Agric. Food Chem. 2011, 59, 6120–6136. [Google Scholar] [CrossRef]
- Aubert, C.; Chalot, G. Chemical composition, bioactive compounds, and volatiles of six table grape varieties (Vitis vinifera L.). Food Chem. 2018, 240, 524–533. [Google Scholar] [CrossRef]
- Bustamante, L.; Sáez, V.; Hinrichsen, P.; Castro, M.H.; Vergara, C.; Von Baer, D.; Mardones, C. Differences in Vvufgt and VvmybA1 Gene Expression Levels and Phenolic Composition in Table Grape (Vitis vinifera L.) ‘Red Globe’ and Its Somaclonal Variant ‘Pink Globe’. J. Agric. Food Chem. 2017, 65, 2793–2804. [Google Scholar] [CrossRef]
- Garde-Cerdán, T.; Portu, J.; López, R.; Santamaría, P. Effect of Foliar Applications of Proline, Phenylalanine, Urea, and Commercial Nitrogen Fertilizers on Stilbene Concentrations in Tempranillo Musts and Wines. Am. J. Enol. Vitic. 2015, 66, 542–547. [Google Scholar] [CrossRef]
- Pérez-Álvarez, E.P.; Martínez-Vidaurre, J.M.; Garde-Cerdán, T. Anthocyanin composition of grapes from three different soil types in cv. Tempranillo, A.O.C. Rioja vineyards. J. Sci. Food Agric. 2019, 99, 4833–4841. [Google Scholar] [CrossRef]
- Ojeda, H.; Andary, C.; Kraeva, E.; Carbonneau, A.; Deloire, A. Influence of pre- and postveraison water deficit on syn-thesis and concentration of skin phenolic compounds during berry growth of Vitis vinifera cv. Shiraz. Am. J. Enol. Vitic. 2002, 53, 261–267. [Google Scholar]
- Rusjan, D.; Veberic, R.; Mikulič-Petkovšek, M. The response of phenolic compounds in grapes of the variety ‘Chardonnay’ (Vitis vinifera L.) to the infection by phytoplasma Bois noir. Eur. J. Plant Pathol. 2012, 133, 965–974. [Google Scholar] [CrossRef]
- Perez-Magariño, S.; José, M.L.G.-S. Evolution of Flavanols, Anthocyanins, and Their Derivatives during the Aging of Red Wines Elaborated from Grapes Harvested at Different Stages of Ripening. J. Agric. Food Chem. 2004, 52, 1181–1189. [Google Scholar] [CrossRef] [PubMed]
- Kovalenko, Y.; Tindjau, R.; Madilao, L.L.; Castellarin, S.D. Regulated deficit irrigation strategies affect the terpene accumulation in Gewürztraminer (Vitis vinifera L.) grapes grown in the Okanagan Valley. Food Chem. 2020, 341, 128172. [Google Scholar] [CrossRef] [PubMed]
- Gambetta, G.A.; Herrera, J.C.; Dayer, S.; Feng, Q.; Hochberg, U.; Castellarin, S.D. The physiology of drought stress in grapevine: Towards an integrative definition of drought tolerance. J. Exp. Bot. 2020, 71, 4658–4676. [Google Scholar] [CrossRef] [PubMed]
- Romero, P.; Botía, P.; del Amor, F.M.; Gil-Muñoz, R.; Flores, P.; Navarro, J.M. Interactive effects of the rootstock and the deficit irrigation technique on wine composition, nutraceutical potential, aromatic profile, and sensory attributes under semiarid and water limiting conditions. Agric. Water Manag. 2019, 225, 105733. [Google Scholar] [CrossRef]
- Mirás-Avalos, J.M.; Buesa, I.; Llacer, E.; Bello, M.A.J.; Risco, D.; Castel, J.R.; Intrigliolo, D.S. Water Versus Source–Sink Relationships in a Semiarid Tempranillo Vineyard: Vine Performance and Fruit Composition. Am. J. Enol. Vitic. 2016, 68, 11–22. [Google Scholar] [CrossRef]
- Jacob, J.K.; Hakimuddin, F.; Paliyath, G.; Fisher, H. Antioxidant and antiproliferative activity of polyphenols in novel high-polyphenol grape lines. Food Res. Int. 2008, 41, 419–428. [Google Scholar] [CrossRef]
- Meng, J.; Fang, Y.; Zhang, A.; Chen, S.; Xu, T.; Ren, Z.; Han, G.; Liu, J.; Li, H.; Zhang, Z.; et al. Phenolic content and antioxidant capacity of Chinese raisins produced in Xinjiang Province. Food Res. Int. 2011, 44, 2830–2836. [Google Scholar] [CrossRef]
- De Castilhos, M.B.M.; Gómez-Alonso, S.; García-Romero, E.; del Bianchi, V.L.; Hermosín-Gutiérrez, I. Isabel red wines produced from grape pre-drying and submerged cap winemaking: A phenolic and sensory approach. LWT Food Sci. Technol. 2017, 81, 58–66. [Google Scholar] [CrossRef]
- Toscano, L.T.; Silva, A.S.; Toscano, L.T.; Tavares, R.L.; Biasoto, A.; de Camargo, A.C.; da Silva, C.S.O.; Gonçalves, M.D.C.R.; Shahidi, F. Phenolics from purple grape juice increase serum antioxidant status and improve lipid profile and blood pressure in healthy adults under intense physical training. J. Funct. Foods 2017, 33, 419–424. [Google Scholar] [CrossRef]
- Fernández-Fernández, A.M.; De Hond, A.I.; Dellacassa, E.; Medrano, A.; Del Castillo, M.D. Assessment of antioxidant, antidiabetic, antiobesity, and anti-inflammatory properties of a Tannat winemaking by-product. Eur. Food Res. Technol. 2019, 245, 1539–1551. [Google Scholar] [CrossRef]
- Li, F.-X.; Yang, Y.-X.; Yin, R.; Ming, J. Comparison of phenolic profiles and antioxidant activities in skins and pulps of eleven grape cultivars (Vitis vinifera L.). J. Integr. Agric. 2019, 18, 1148–1158. [Google Scholar] [CrossRef]
- Allen, R.G.; Pereira, L.S.; Raes, D.; Smith, M. Crop evapotranspiration—Guidelines for computing crop water requirements—FAO irrigation and drainage paper 56. Rome 1998, 300, D05109. [Google Scholar]
- Portu, J.; López, R.; Baroja, E.; Santamaría, P.; Garde-Cerdán, T. Improvement of grape and wine phenolic content by foliar application to grapevine of three different elicitors: Methyl jasmonate, chitosan, and yeast extract. Food Chem. 2016, 201, 213–221. [Google Scholar] [CrossRef]
- Castillo-Muñoz, N.; Fernández-González, M.; Gómez-Alonso, S.; García-Romero, E.; Hermosín-Gutiérrez, I.; García-Romero, E. Red-Color Related Phenolic Composition of Garnacha Tintorera (Vitis vinifera L.) Grapes and Red Wines. J. Agric. Food Chem. 2009, 57, 7883–7891. [Google Scholar] [CrossRef]
- Castillo-Muñoz, N.; Gómez-Alonso, S.; García-Romero, E.; Hermosín-Gutiérrez, I. Flavonol Profiles of Vitis vinifera Red Grapes and Their Single-Cultivar Wines. J. Agric. Food Chem. 2007, 55, 992–1002. [Google Scholar] [CrossRef] [PubMed]
- Mosca, F.; Hidalgo, G.I.; Villasante, J.; Almajano, M.P. Continuous or Batch Solid-Liquid Extraction of Antioxidant Compounds from Seeds of Sterculia apetala Plant and Kinetic Release Study. Molecules 2018, 23, 1759. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Villasante, J.; Pérez-Carrillo, E.; Heredia-Olea, E.; Metón, I.; Almajano, M.P. In Vitro Antioxidant Activity Optimization of Nut Shell (Carya illinoinensis) by Extrusion Using Response Surface Methods. Biomolecules 2019, 9, 883. [Google Scholar] [CrossRef] [Green Version]
- Azman, N.A.M.; Gallego, M.G.; Segovia, F.; Abdullah, S.; Shaarani, S.M.; Pablos, M.P.A. Study of the Properties of Bearberry Leaf Extract as a Natural Antioxidant in Model Foods. Antioxidants 2016, 5, 11. [Google Scholar] [CrossRef] [Green Version]
- Segovia, F.J.; Corral-Pérez, J.J.; Almajano, M. Avocado seed: Modeling extraction of bioactive compounds. Ind. Crop. Prod. 2016, 85, 213–220. [Google Scholar] [CrossRef]
- Garde-Cerdán, T.; Gutiérrez-Gamboa, G.; Ayestarán, B.; González-Lázaro, M.; Rubio-Bretón, P.; Pérez-Álvarez, E. Influence of seaweed foliar application to Tempranillo grapevines on grape and wine phenolic compounds over two vintages. Food Chem. 2020, 345, 128843. [Google Scholar] [CrossRef] [PubMed]
- Zarrouk, O.; Brunetti, C.; Egipto, R.; Pinheiro, C.; Genebra, T.; Gori, A.; Lopes, C.; Tattini, M.; Chaves, M.M. Grape Ripening Is Regulated by Deficit Irrigation/Elevated Temperatures According to Cluster Position in the Canopy. Front. Plant Sci. 2016, 7, 1640. [Google Scholar] [CrossRef] [Green Version]
- Yu, R.; Cook, M.G.; Yacco, R.S.; Watrelot, A.; Gambetta, G.; Kennedy, J.A.; Kurtural, S.K. Effects of Leaf Removal and Applied Water on Flavonoid Accumulation in Grapevine (Vitis viniferaL. cv. Merlot) Berry in a Hot Climate. J. Agric. Food Chem. 2016, 64, 8118–8127. [Google Scholar] [CrossRef] [PubMed]
- Savoi, S.; Herrera, J.C.; Carlin, S.; Lotti, C.; Bucchetti, B.; Peterlunger, E.; Castellarin, S.D.; Mattivi, F. From grape berries to wines: Drought impacts on key secondary metabolites. OENO One 2020, 54, 569–582. [Google Scholar] [CrossRef]
- Spayd, S.E.; Tarara, J.M.; Mee, D.L.; Ferguson, J.C. Separation of sunlight and temperature effects on the composition of Vitis vinifera cv. Merlot berries. Am. J. Enol. Vitic. 2002, 53, 171–182. [Google Scholar]
- Tarara, J.M.; Lee, J.; Spayd, S.E.; Scagel, C.F. Berry temperature and solar radiation alter acylation, proportion, and con-centration of anthocyanin in Merlot grapes. Am. J. Enol. Vitic. 2008, 59, 235–247. [Google Scholar]
- Grimplet, J.; Deluc, L.G.; Tillett, R.L.; Wheatley, M.D.; Schlauch, K.A.; Cramer, G.R.; Cushman, J.C. Tissue-specific mRNA expression profiling in grape berry tissues. BMC Genom. 2007, 8, 187. [Google Scholar] [CrossRef] [Green Version]
- Kennedy, J.A.; Matthews, M.A.; Waterhouse, A.L. Effect of maturity and vine water status on grape skin and wine fla-vonoids. Am. J. Enol. Vitic. 2002, 53, 268–274. [Google Scholar]
- Castellarin, S.D.; Matthews, M.A.; di Gaspero, G.; Gambetta, G.A. Water deficits accelerate ripening and induce changes in gene expression regulating flavonoid biosynthesis in grape berries. Planta 2007, 227, 101–112. [Google Scholar] [CrossRef] [PubMed]
- Intrigliolo, D.S.; Castel, J.R. Response of grapevine cv. ‘Tempranillo’ to timing and amount of irrigation: Water relations, vine growth, yield and berry and wine composition. Irrig. Sci. 2009, 28, 113–125. [Google Scholar] [CrossRef]
- Roby, G.; Harbertson, J.F.; Adams, D.A.; Matthews, M.A. Berry size and vine water deficits as factors in winegrape composition: Anthocyanins and tannins. Aust. J. Grape Wine Res. 2004, 10, 100–107. [Google Scholar] [CrossRef]
- Martínez-Lüscher, J.; Brillante, L.; Kurtural, S.K. Flavonol Profile Is a Reliable Indicator to Assess Canopy Architecture and the Exposure of Red Wine Grapes to Solar Radiation. Front. Plant Sci. 2019, 10, 10. [Google Scholar] [CrossRef] [Green Version]
- Luzio, A.; Bernardo, S.; Correia, C.; Moutinho-Pereira, J.; Dinis, L.-T. Phytochemical screening and antioxidant activity on berry, skin, pulp and seed from seven red Mediterranean grapevine varieties (Vitis vinifera L.) treated with kaolin foliar sunscreen. Sci. Hortic. 2021, 281, 109962. [Google Scholar] [CrossRef]
- Cerda-Carrasco, A.; López-Solís, R.; Nuñez-Kalasic, H.; Peña-Neira, Á.; Obreque-Slier, E. Phenolic composition and antioxidant capacity of pomaces from four grape varieties (Vitis vinifera L.). J. Sci. Food Agric. 2014, 95, 1521–1527. [Google Scholar] [CrossRef]
- Gironés-Vilaplana, A.; Moreno, D.A.; Garcia-Viguera, C. Phytochemistry and biological activity of Spanish Citrus fruits. Food Funct. 2014, 5, 764–772. [Google Scholar] [CrossRef] [PubMed]
- Zacarías-García, J.; Rey, F.; Gil, J.-V.; Rodrigo, M.J.; Zacarías, L. Antioxidant capacity in fruit of Citrus cultivars with marked differences in pulp coloration: Contribution of carotenoids and vitamin C. Food Sci. Technol. Int. 2020, 27, 210–222. [Google Scholar] [CrossRef]
- Zarrouk, O.; Francisco, R.; Pintó-Marijuan, M.; Brossa, R.; Santos, R.R.; Pinheiro, C.; Costa, J.; Lopes, C.; Chaves, M.M. Impact of irrigation regime on berry development and flavonoids composition in Aragonez (Syn. Tempranillo) grapevine. Agric. Water Manag. 2012, 114, 18–29. [Google Scholar] [CrossRef]
- Genebra, T.; Santos, R.R.; Francisco, R.; Pinto-Marijuan, M.; Brossa, R.; Serra, A.T.; Duarte, C.M.M.; Chaves, M.M.; Zarrouk, O. Proanthocyanidin Accumulation and Biosynthesis Are Modulated by the Irrigation Regime in Tempranillo Seeds. Int. J. Mol. Sci. 2014, 15, 11862–11877. [Google Scholar] [CrossRef] [PubMed]
- Kennedy, J.A.; Troup, G.J.; Pilbrow, J.R.; Hutton, D.R.; Hewitt, D.; Hunter, C.R.; Ristic, R.; Iland, P.G.; Jones, G.P. Development of seed polyphenols in berries from Vitis vinifera L. cv. Shiraz. Aust. J. Grape Wine Res. 2000, 6, 244–254. [Google Scholar] [CrossRef]
- Castellarin, S.D.; Pfeiffer, A.; Sivilotti, P.; Degan, M.; Peterlunger, E.; Di Gaspero, G. Transcriptional regulation of anthocyanin biosynthesis in ripening fruits of grapevine under seasonal water deficit. Plant Cell Environ. 2007, 30, 1381–1399. [Google Scholar] [CrossRef] [Green Version]
- Tkacz, K.; Wojdyło, A.; Turkiewicz, I.P.; Nowicka, P. Anti-diabetic, anti-cholinesterase, and antioxidant potential, chemical composition and sensory evaluation of novel sea buckthorn-based smoothies. Food Chem. 2020, 338, 128105. [Google Scholar] [CrossRef]
- González-Tejedor, G.A.; Martínez-Hernández, G.B.; Garre, A.; Egea, J.A.; Fernandez, P.S.; Artés-Hernández, F. Quality Changes and Shelf-Life Prediction of a Fresh Fruit and Vegetable Purple Smoothie. Food Bioprocess Technol. 2017, 10, 1892–1904. [Google Scholar] [CrossRef]
- Jara-Palacios, M.J.; Gonçalves, S.; Heredia, F.J.; Hernanz, D.; Romano, A. Extraction of Antioxidants from Winemaking Byproducts: Effect of the Solvent on Phenolic Composition, Antioxidant and Anti-Cholinesterase Activities, and Electrochemical Behaviour. Antioxidants 2020, 9, 675. [Google Scholar] [CrossRef]
- Teleszko, M.; Wojdyło, A. Bioactive compounds vs. organoleptic assessment of ‘smoothies’-type products prepared from selected fruit species. Int. J. Food Sci. Technol. 2013, 49, 98–106. [Google Scholar] [CrossRef]
- Wojdyło, A.; Samoticha, J.; Chmielewska, J. Effect of different pre-treatment maceration techniques on the content of phenolic compounds and color of Dornfelder wines elaborated in cold climate. Food Chem. 2020, 339, 127888. [Google Scholar] [CrossRef]
- Karaman, H.T.; Küskü, D.Y.; Söylemezoğlu, G. Phenolic compounds and antioxidant capacities in grape berry skin, seed and stems of six winegrape varieties grown in Turkey. Acta Sci. Pol. Hortorum Cultus 2021, 20, 15–25. [Google Scholar] [CrossRef]
- Xu, C.; Zhang, Y.; Cao, L.; Lu, J. Phenolic compounds and antioxidant properties of different grape cultivars grown in China. Food Chem. 2010, 119, 1557–1565. [Google Scholar] [CrossRef]
- Nowicka, P.; Wojdyło, A.; Samoticha, J. Evaluation of phytochemicals, antioxidant capacity, and antidiabetic activity of novel smoothies from selected Prunus fruits. J. Funct. Foods 2016, 25, 397–407. [Google Scholar] [CrossRef]
- Matos, M.S.; Romero-Díez, R.; Álvarez, A.; Bronze, M.R.; Rodríguez-Rojo, S.; Mato, R.B.; Cocero, M.J.; Matias, A.A. Polyphenol-Rich extracts obtained from winemaking waste streams as natural ingredients with cosmeceutical potential. Antioxidants 2019, 8, 355. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Faria, A.; Calhau, C.; Freitas, V.; Mateus, N. Procyanidins as Antioxidants and Tumor Cell Growth Modulators. J. Agric. Food Chem. 2006, 54, 2392–2397. [Google Scholar] [CrossRef] [PubMed]
- Hidalgo, M.; Sanchez-Moreno, C.; de Pascual-Teresa, S. Flavonoid–flavonoid interaction and its effect on their antioxidant activity. Food Chem. 2010, 121, 691–696. [Google Scholar] [CrossRef]
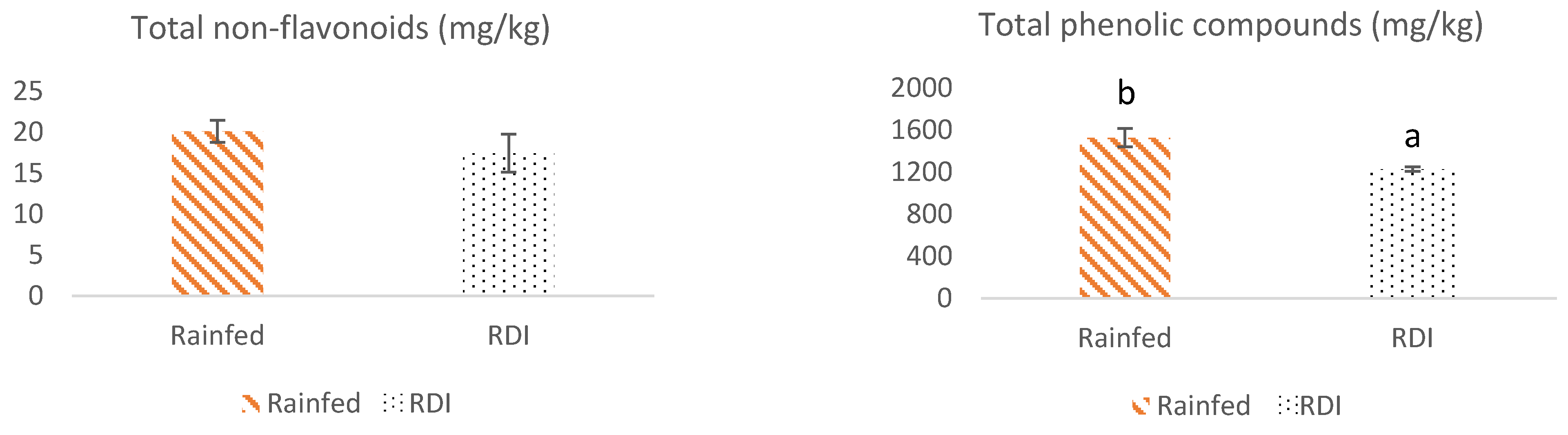

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Treatments (T) | |||
|---|---|---|---|
| Anthocyanins | Rainfed | RDI | p-Value 1 |
| Delphinidin-3-glc | 101.5 ± 18.5 | 86.1 ± 5.8 | 0.224 |
| Cyanidin-3-glc | 78.0 ± 5.7 | 72.3 ± 7.4 | 0.350 |
| Petunidin-3-glc | 108.9 ± 17.0 | 87.7 ± 4.0 | 0.088 |
| Peonidin-3-glc | 106.7 ± 5.6 b | 93.4 ± 2.9 a | * |
| Malvidin-3-glc | 370.1 ± 27.3 b | 273.2 ± 2.7 a | ** |
| Total non-acylated | 765.2 ± 60.4 b | 612.6 ± 12.8 a | *** |
| Delphinidin-3-acglc | 5.3 ± 0.4 b | 4.1 ± 0.4 a | * |
| Cyanidin-3-acglc | 3.3 ± 0.2 | 2.8 ± 0.3 | 0.437 |
| Petunidin-3-acglc | 5.0 ± 0.4 b | 4.1 ± 0.2 a | ** |
| Peonidin-3-acglc | 2.4 ± 0.3 | 2.3 ± 0.1 | 0.203 |
| Malvidin-3-acglc | 14.0 ± 0.8 b | 10.4 ± 0.9 a | ** |
| Delphinidin-3-cmglc | 10.9 ± 1.5 b | 8.5 ± 1.1 a | * |
| Petunidin-3-cmglc | 16.3 ± 2.3 b | 10.9 ± 0.2 a | ** |
| Cyanidin-3-cmglc | 11.0 ± 0.1 b | 10.4 ± 0.1 a | *** |
| Peonidin-3-cmglc | 19.0 ± 1.1 b | 11.4 ± 0.5 a | *** |
| Malvidin-3-cis-cmglc | 3.6 ± 0.7 | 2.6 ± 0.1 | 0.203 |
| Malvidin-3-trans-cmglc | 68.9 ± 9.7 b | 40.1 ± 1.7 a | ** |
| Malvidin-3-cfglc | 2.5 ± 0.3 | 2.0 ± 0.1 | 0.286 |
| Total acylated | 162.3 ± 14.3 b | 109.6 ± 3.2 a | ** |
| Treatments (T) | |||
|---|---|---|---|
| Flavonols | Rainfed | RDI | p-Value 1 |
| Myricetin-3-glcU | 12.0 ± 1.8 b | 6.5 ± 0.5 a | ** |
| Myricetin-3-gal | 8.8 ± 1.6 | 6.7 ± 1.0 | 0.066 |
| Myricetin-3-glc | 77.3 ± 8.2 | 68.4 ± 2.4 | 0.139 |
| Total myricetins | 98.1 ± 7.9 b | 81.5 ± 2.6 a | ** |
| Quercetin-3-glcU | 101.9 ± 17.7 | 77.5 ± 4.5 | 0.080 |
| Quercetin-3-glc | 129.4 ± 14.4 | 140.4 ± 15.3 | 0.425 |
| Total quercetins | 231.4 ± 16.7 | 218.0 ± 14.6 | 0.279 |
| Laricitrin-3-glc | 13.8 ± 0.8 b | 9.0 ± 0.5 a | *** |
| Kaempferol-3-gal | 3.1 ± 0.6 | 1.5 ± 0.3 | 0.067 |
| Kaempferol-3-glc | 25.0 ± 0.9 | 21.8 ± 4.8 | 0.297 |
| Total kaempferols | 28.1 ± 0.6 | 23.3 ± 4.9 | 0.120 |
| Isorhamnetin-3-gal | 2.2 ± 0.2 b | 1.7 ± 0.1 a | * |
| Isorhamnetin-3-glc | 4.2 ± 0.4 | 4.8 ± 0.6 | 0.519 |
| Total isorhamnetins | 6.5 ± 0.4 | 6.2 ± 0.4 | 0.537 |
| Syringetin-3-glc | 6.0 ± 0.1 b | 4.3 ± 0.2 a | ** |
| Treatments (T) | |||
|---|---|---|---|
| Flavanols | Rainfed | RDI | p-Value 1 |
| Catechin | 48.1 ± 3.3 | 38.6 ± 8.1 | 0.131 |
| Epicatechin | 58.1 ± 1.1 b | 37.6 ± 2.7 a | *** |
| Epicatechin-3-gallate | 68.8 ± 4.3 b | 50.4 ± 5.8 a | ** |
| Procyanidin B1 | 10.0 ± 1.4 | 8.8 ± 1.5 | 0.242 |
| Procyanidin B2 | 9.5 ± 1.3 | 10.8 ± 1.2 | 0.322 |
| Treatments (T) | |||
|---|---|---|---|
| Non-Flavonoids | Rainfed | RDI | p-Value 1 |
| Hydroxybenzoic acids | |||
| Syringic acid | 3.9 ± 0.6 | 1.5 ± 0.2 | 0.058 |
| Gallic acid | 6.1 ± 1.4 | 4.9 ± 0.5 | 0.147 |
| Hydroxycinnamic acids | |||
| trans-Caftaric acid | 1.5 ± 0.3 | 2.5 ± 0.8 | 0.132 |
| trans + cis-Coutaric acids | 1.0 ± 0.1 | 1.6 ± 0.4 | 0.116 |
| Caffeic acid | 1.0 ± 0.1 | 1.0 ± 0.1 | 0.489 |
| trans-Fertaric acid | 1.0 ± 0.1 b | 0.9 ± 0.1 a | ** |
| p-Coumaric acid | 0.4 ± 0.1 | 0.4 ± 0.1 | 0.718 |
| Stilbenes | |||
| trans-Piceid | 4.1 ± 0.2 | 3.9 ± 1.4 | 0.822 |
| cis-Piceid | 0.4 ± 0.1 | 0.4 ± 0.1 | 0.866 |
| trans-Resveratrol | 0.3 ± 0.1 | 0.3 ± 0.1 | 0.558 |
| cis-Resveratrol | 0.3 ± 0.1 | 0.2 ± 0.1 | 0.064 |
| TPC (mg GAE/mL) | ABTS 1 (mmol TE/mL) | DPPH (mmol TE/mL) | ORAC (μmol TE/mL) | |
|---|---|---|---|---|
| Rainfed | 1.5 ± 0.2 b | 3.7 ± 0.1 a | 5.6 ± 0.5 | 43.6 ± 6.9 b |
| RDI | 1.2 ± 0.1 a | 3.9 ± 0.1 a | 5.3 ± 0.1 | 23.4 ± 3.1 a |
| p-value 2 | ** | * | 0.265 | ** |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Pérez-Álvarez, E.P.; Intrigliolo, D.S.; Almajano, M.P.; Rubio-Bretón, P.; Garde-Cerdán, T. Effects of Water Deficit Irrigation on Phenolic Composition and Antioxidant Activity of Monastrell Grapes under Semiarid Conditions. Antioxidants 2021, 10, 1301. https://doi.org/10.3390/antiox10081301
Pérez-Álvarez EP, Intrigliolo DS, Almajano MP, Rubio-Bretón P, Garde-Cerdán T. Effects of Water Deficit Irrigation on Phenolic Composition and Antioxidant Activity of Monastrell Grapes under Semiarid Conditions. Antioxidants. 2021; 10(8):1301. https://doi.org/10.3390/antiox10081301
Chicago/Turabian StylePérez-Álvarez, Eva P., Diego S. Intrigliolo, María Pilar Almajano, Pilar Rubio-Bretón, and Teresa Garde-Cerdán. 2021. "Effects of Water Deficit Irrigation on Phenolic Composition and Antioxidant Activity of Monastrell Grapes under Semiarid Conditions" Antioxidants 10, no. 8: 1301. https://doi.org/10.3390/antiox10081301
APA StylePérez-Álvarez, E. P., Intrigliolo, D. S., Almajano, M. P., Rubio-Bretón, P., & Garde-Cerdán, T. (2021). Effects of Water Deficit Irrigation on Phenolic Composition and Antioxidant Activity of Monastrell Grapes under Semiarid Conditions. Antioxidants, 10(8), 1301. https://doi.org/10.3390/antiox10081301