
Oxidative Stress and Male Fertility: Role of Antioxidants and Inositols


 ,
,  and
and 
Abstract
1. Introduction
2. Dual Role of ROS and Antioxidant System in Male Fertility
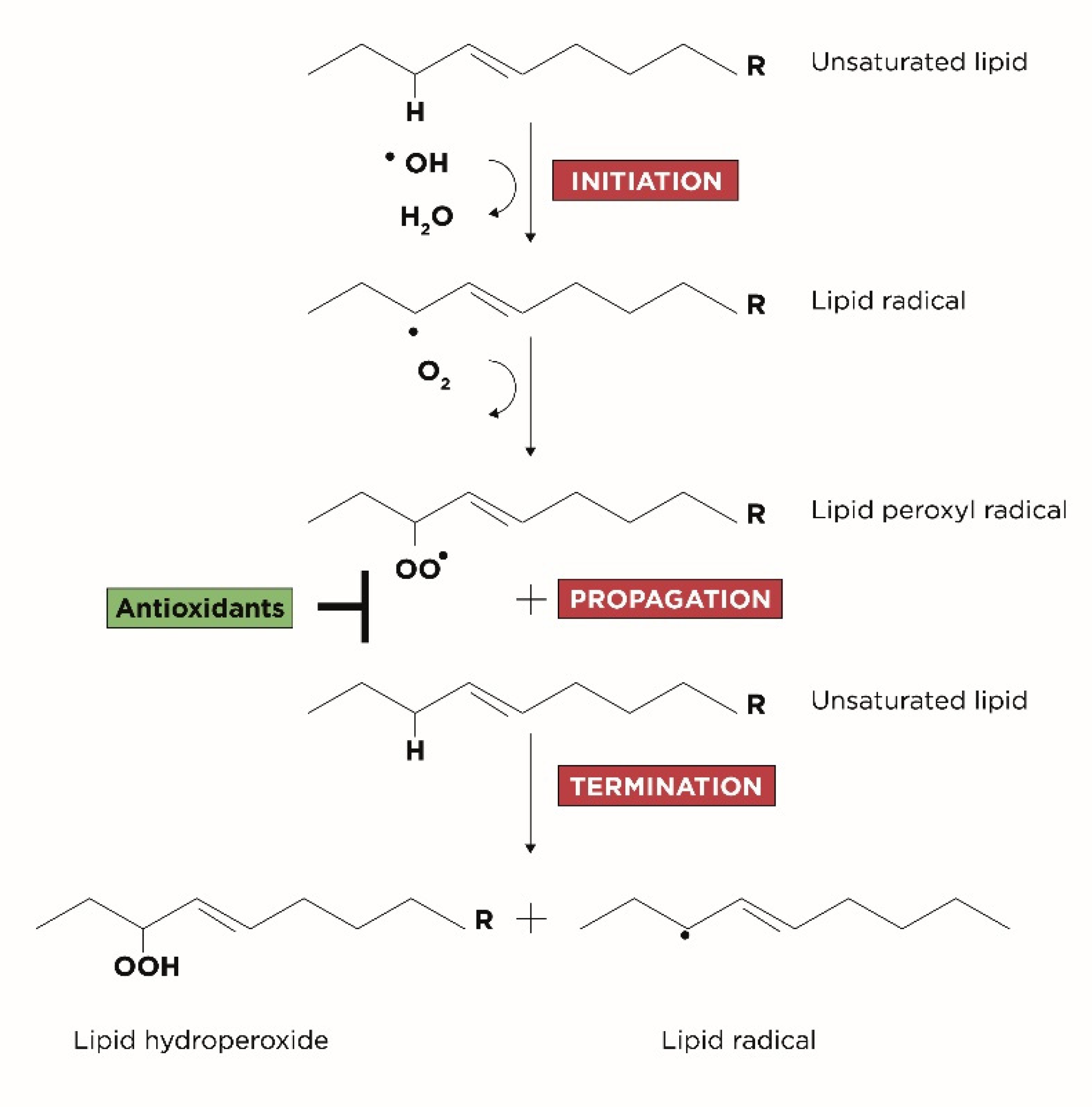
Genesis of Oxidative Stress, Lipid Peroxidation and DNA Damage
- Initiation: corresponds to the extraction of hydrogen atoms from the carbon-carbon double bonds of an unsaturated fatty acid to generate free radicals;
- Propagation: corresponds to the formation of lipid radicals followed by their rapid reaction with oxygen to form peroxyl radicals;
- Termination: corresponds to the last phase in which the radicals formed react with other lipids generating different cytotoxic adducts such as aldehydes [25].
3. Role of Antioxidant in Male Fertility
3.1. Folic Acid
3.2. L-carnitine
3.3. L-arginine
3.4. N-acetylcystenine
- (1)
- A direct antioxidant effect toward certain oxidant species including NO2 and hypohalous acids (HOX). HOX, due to their high reactivity, are not specific oxidants and also react with many biologically important molecules, thus inducing a cytotoxic effect [77].
- (2)
- As NAC acts as a predecessor of cysteine and is part of important step to glutathione synthesis has an indirect antioxidant effect. Then GSH is engaged in different detoxification processes as elimination of by-product of lipid peroxidation and hydroperoxides [78].
- (3)
3.5. Resveratrol
4. Role of Inositols in Male Fertility
4.1. MI: In Vitro Studies
4.2. MI: In Vivo Studies
4.3. DCI in Male Fertility
5. Conclusions
Author Contributions
Funding
Conflicts of Interest
References
- Comhaire, F.H.; World Health Organization. Towards More Objectivity in Diagnosis and Management of Male Infertility; Blackwell Scientific: Oxford, UK, 1987; Volume 10, pp. 1–53. [Google Scholar]
- Agarwal, A.; Majzoub, A.; Parekh, N.; Henkel, R. A schematic overview of the current status of male infertility practice. World J. Men’s Health 2020, 38, 308–322. [Google Scholar] [CrossRef]
- Iwasaki, A.; Gagnon, C. Formation of reactive oxygen species in spermatozoa of infertile patients. Fertil. Steril. 1992, 57, 409–416. [Google Scholar] [CrossRef]
- Tremellen, K. Oxidative stress and male infertility—A clinical perspective. Hum. Reprod Update 2008, 14, 243–258. [Google Scholar] [CrossRef] [PubMed]
- Wu, P.Y.; Scarlata, E.; O’Flaherty, C. Long-term adverse effects of oxidative stress on rat epididymis and spermatozoa. Antioxidants 2020, 9, 170. [Google Scholar] [CrossRef]
- Ghaleno, L.R.; Alizadeh, A.; Drevet, J.R.; Shahverdi, A.; Valojerdi, M.R. Oxidation of sperm DNA and male infertility. Antioxidants 2021, 10, 97. [Google Scholar] [CrossRef] [PubMed]
- Durairajanayagam, D. Lifestyle causes of male infertility. Arab. J. Urol. 2018, 16, 10–20. [Google Scholar] [CrossRef]
- Plaseska-Karanfilska, D.; Noveski, P.; Plaseski, T.; Maleva, I.; Madjunkova, S.; Moneva, Z. Genetic causes of male infertility. Balk. J. Med. Genet. 2012, 15, 31–34. [Google Scholar] [CrossRef]
- Anifandis, G.; Katsanaki, K.; Lagodonti, G.; Messini, C.; Simopoulou, M.; Dafopoulos, K.; Daponte, A. The effect of glyphosate on human sperm motility and sperm DNA fragmentation. Int. J. Environ. Res. Public Health 2018, 15, 1117. [Google Scholar] [CrossRef]
- Ochsendorf, F.R. Infections in the male gential tract and reactive oxygen species. Hum. Reprod. Update 1999, 5, 399–420. [Google Scholar] [CrossRef]
- Dutta, S.; Sengupta, P. SARS-CoV-2 and male infertility: Possible multifaced pathology. Reprod. Sci. 2021, 28, 23–26. [Google Scholar] [CrossRef]
- Fave, R.F.D.; Polisini, G.; Giglioni, G.; Parlavecchio, A.; Dell’Atti, L.; Galosi, A.B. COVID-19 and male fertility: Taking stock of one year after the outbreak began. Arch. Ital. Urol. Androl. 2021, 93, 115–119. [Google Scholar] [CrossRef]
- Falahieh, F.M.; Zarabadipour, M.; Mirani, M.; Abdiyan, M.; Dinparvar, M.; Alizadeh, H.; Paktinat, S.; Hosseinirad, H. Effects of moderate COVID-19 infection on semen oxidative status and parameters 14 and 120 days after diagnosis. Reprod. Fertil. Dev. 2021. [Google Scholar] [CrossRef]
- Arafa, M.; Agarwal, A.; Majzoub, A.; Selvam, M.K.P.; Baskaran, S.; Henkel, R.; ElBardisi, H. Efficacy of antioxidant supplementation on conventional and advanced sperm function tests in patients with idiopathic male infertility. Antioxidants 2020, 9, 219. [Google Scholar] [CrossRef]
- Calogero, A.E.; Condorelli, R.A.; Russo, G.I.; La Vignera, S. Conservative nonhormonal options for the treatment of male infertility: Antibiotics, anti-inflammatory drugs, and antioxidants. BioMed Res. Int. 2017, 2017, 4650182. [Google Scholar] [CrossRef] [PubMed]
- Oliva, M.M.; Buonomo, G.; Carra, M.C.; Lippa, A.; Lisi, F. Myo-inositol impact on sperm motility in vagina and evaluation of its effects on foetal development. Eur. Rev. Med. Pharmacol. Sci. 2020, 24, 2704–2709. [Google Scholar]
- Gambioli, R.; Forte, G.; Aragona, C.; Bevilacqua, A.; Bizzarri, M.; Unfer, V. The use of D-chiro-Inositol in clinical practice. Eur. Rev. Med. Pharmacol. Sci. 2021, 25, 438–446. [Google Scholar] [PubMed]
- O’Flaherty, C. Reactive oxygen species and male fertility. Antioxidants 2020, 9, 287. [Google Scholar] [CrossRef] [PubMed]
- Martin-Hidalgo, D.; Bragado, M.J.; Batista, A.R.; Oliveira, P.F.; Alves, M.G. Antioxidants and male fertility: From molecular studies to clinical evidence. Antioxidants 2019, 8, 89. [Google Scholar] [CrossRef] [PubMed]
- Dobrakowski, M.; Kasperczyk, S.; Horak, S.; Chyra-Jach, D.; Birkner, E. Oxidative stress and motility impairment in the semen of fertile males. Andrologia 2017, 49, e12783. [Google Scholar] [CrossRef]
- Cocuzza, M.; Sikka, S.C.; Athayde, K.S.; Agarwal, A. Clinical relevance of oxidative stress and sperm chromatin damage in male infertility: An evidence based analysis. Int. Braz. J. Urol. 2007, 33, 603–621. [Google Scholar] [CrossRef]
- Rehman, R.; Amjad, S.; Tariq, H.; Zahid, N.; Akhter, M.; Ashraf, M. Oxidative stress and male infertility: A cross sectional study. J. Pakistan Med. Assoc. 2020, 70, 461–466. [Google Scholar] [CrossRef]
- Walczak–Jedrzejowska, R.; Wolski, J.K.; Slowikowska–Hilczer, J. The role of oxidative stress and antioxidants in male fertility. Central Eur. J. Urol. 2013, 65, 60–67. [Google Scholar] [CrossRef]
- Imai, H.; Suzuki, K.; Ishizaka, K.; Ichinose, S.; Oshima, H.; Okayasu, I.; Emoto, K.; Umeda, M.; Nakagawa, Y. Failure of the expression of phospholipid hydroperoxide glutathione peroxidase in the spermatozoa of human infertile males1. Biol. Reprod. 2001, 64, 674–683. [Google Scholar] [CrossRef]
- Agarwal, A.; Rana, M.; Qiu, E.; Albunni, H.; Bui, A.D.; Henkel, R. Role of oxidative stress, infection and inflammation in male infertility. Andrologia 2018, 50, e13126. [Google Scholar] [CrossRef] [PubMed]
- Sanocka, D.; Kurpisz, M. Reactive oxygen species and sperm cells. Reprod. Biol. Endocrinol. 2004, 2, 12. [Google Scholar] [CrossRef]
- Lucock, M. Folic acid: Nutritional biochemistry, molecular biology, and role in disease processes. Mol. Genet. Metab. 2000, 71, 121–138. [Google Scholar] [CrossRef] [PubMed]
- Nematollahi-Mahani, S.N.; Azizollahi, G.H.; Baneshi, M.R.; Safari, Z.; Azizollahi, S. Effect of folic acid and zinc sulphate on endocrine parameters and seminal antioxidant level after varicocelectomy. Andrologia 2014, 46, 240–245. [Google Scholar] [CrossRef] [PubMed]
- Joshi, R.; Adhikari, S.; Patro, B.S.; Chattopadhyay, S.; Mukherjee, T. Free radical scavenging behavior of folic acid: Evi-dence for possible antioxidant activity. Free Radic. Biol. Med. 2001, 30, 1390–1399. [Google Scholar] [CrossRef]
- Moorthy, P.N.; Hayon, E. Intermediates produced from the one-electron reduction of nitrogen heterocyclic compounds in solution. J. Phys. Chem. 1974, 78, 2615–2620. [Google Scholar] [CrossRef]
- Moorthy, P.N.; Hayon, E. One-electron redox reactions of water soluble vitamins II. Pterin and folic acid. J. Org. Chem. 1976, 41, 1607–1613. [Google Scholar] [CrossRef] [PubMed]
- Vanderschitt, D.J.; Scimgeour, K.C. Reaction of Cu2+ and Fe3+ with tetrahydropteridines. Biochem. Biophys. Res. Commun. 1967, 28, 302–308. [Google Scholar] [CrossRef]
- Hemmerich, P. Free radicals of biological interest as studied by electron spin resonance. Proc. R. Soc. London. Ser. A Math. Phys. Sci. 1968, 302, 335–350. [Google Scholar] [CrossRef]
- Bellamy, C.O. p53 and apoptosis. Br. Med. Bull. 1997, 53, 522–538. [Google Scholar] [CrossRef] [PubMed]
- Chern, C.L.; Huang, R.F.; Chen, Y.H.; Cheng, J.T.; Liu, T.Z. Folate deficiency-induced oxidative stress and apoptosis are mediated via homocysteine-dependent overproduction of hydrogen peroxide and enhanced activation of NF-kappaB in human Hep G2 cells. Biomed. Pharmacother. 2001, 55, 434–442. [Google Scholar] [CrossRef]
- Bentivoglio, G.; Melica, F.; Cristoforoni, P. Folinic acid in the treatment of human male infertility. Fertil. Steril. 1993, 60, 698–701. [Google Scholar] [CrossRef]
- Pierik, F.H.; Vreeburg, J.T.; Stijnen, T.; De Jong, F.H.; Weber, R.F. Serum inhibin B as a marker of spermatogenesis. J. Clin. Endocrinol. Metab. 1998, 83, 3110–3114. [Google Scholar] [CrossRef]
- Andersson, A.-M. Inhibin B in the assessment of seminiferous tubular function. Best Pract. Res. Clin. Endocrinol. Metab. 2000, 14, 389–397. [Google Scholar] [CrossRef]
- Anderson, R.A.; Sharpe, R.M. Regulation of inhibin production in the human male and its clinical applications. Int. J. Androl. 2000, 23, 136–144. [Google Scholar] [CrossRef]
- Irani, M.; Amirian, M.; Sadeghi, R.; Le Lez, J.; Roudsari, R.L. The effect of folate and folate plus zinc supplementation on endocrine parameters and sperm characteristics in sub-fertile men: A systematic review and meta-analysis. Urol. J. 2017, 14, 4069–4078. [Google Scholar] [PubMed]
- Jeulin, C.; Lewin, L.M. Role of free L-carnitine and acetyl-L-carnitine in post-gonadal maturation of mammalian spermatozoa. Hum. Reprod. Update 1996, 2, 87–102. [Google Scholar] [CrossRef]
- Bahl, J.J. Bressler, R. The pharmacology of carnitine. Annu. Rev. Pharmacol. Toxicol. 1987, 27, 257–277. [Google Scholar] [CrossRef] [PubMed]
- Kerner, J.; Hoppel, C. Generic disorders of carnitine metabolism and their nutritional management. Annu. Rev. Nutr. 1998, 18, 179–206. [Google Scholar] [CrossRef] [PubMed]
- Adeva-Andany, M.M.; Calvo-Castro, I.; Fernández-Fernández, C.; Donapetry-García, C.; Pedre-Piñeiro, A.M. Signifi-cance of l-carnitine for human health. IUBMB Life 2017, 69, 578–594. [Google Scholar] [CrossRef]
- Cabral, R.E.L.; Mendes, T.B.; Vendramini, V.; Miraglia, S.M. Carnitine partially improves oxidative stress, acrosome integrity, and reproductive competence in doxorubicin-treated rats. Andrology 2018, 6, 236–246. [Google Scholar] [CrossRef] [PubMed]
- Vardiyan, R.; Ezati, D.; Anvari, M.; Ghasemi, N.; Talebi, A. Effect of L-carnitine on the expression of the apoptotic genes Bcl-2 and Bax. Clin. Exp. Reprod. Med. 2020, 47, 155–160. [Google Scholar] [CrossRef] [PubMed]
- Vicari, E.; Calogero, A.E. Effects of treatment with carnitines in infertile patients with pros-tato-vesiculo-epididymitis. Hum. Reprod. 2001, 16, 2338–2342. [Google Scholar] [CrossRef]
- Vicari, E.; Rubino, C.; De Palma, A.; Longo, G.; Lauretta, M.; Consoli, S.; Arancio, A. Antioxidant therapeutic effi-ciency after the use of carnitine in infertile patients with bacterial or non bacterial prostato-vesiculo-epididymitis. Arch. Ital. Urol Androl. 2001, 73, 15–25. [Google Scholar]
- Vicari, E.; La Vignera, S.; Calogero, A.E. Antioxidant treatment with carnitines is effective in infertile patients with prostatovesiculoepididymitis and elevated seminal leukocyte concentrations after treatment with nonsteroidal anti-inflammatory compounds. Fertil. Steril. 2002, 78, 1203–1208. [Google Scholar] [CrossRef]
- Mehni, N.M.; Ketabchi, A.A.; Hosseini, E. Combination effect of Pentoxifylline and L-carnitine on idiopathic oligoasthenoteratozoospermia. Iran. J. Reprod. Med. 2014, 12, 817–824. [Google Scholar]
- Lenzi, A.; Lombardo, F.; Sgr o, P.; Salacone, P.; Caponecchia, L.; Dondero, F.; Gandini, L. Use of carnitine therapy in selected cases of male factor infertility: A double-blind crossover trial. Fertil. Steril. 2003, 79, 292–300. [Google Scholar] [CrossRef]
- Lenzi, A.; Sgro, P.; Salacone, P.; Paoli, D.; Gilio, B.; Lombardo, F.; Santulli, M.; Agarwal, A.; Gandini, L. A placebo controlled double-blind randomized trial of the use of combined l-carnitine and l-acetylcarnitine treatment in men with asthenozoospermia. Fertil. Steril. 2004, 81, 1578–1584. [Google Scholar] [CrossRef] [PubMed]
- Balercia, G.; Regoli, F.; Armeni, T.; Koverech, A.; Mantero, F.; Boscaro, M. Placebo-controlled double-blind randomized trial on the use of L-carnitine, L-acetylcarnitine, or combined L-carnitine and Lacetylcarnitine in men with idiopathic asthenozoospermia. Fertil. Steril. 2005, 84, 662–671. [Google Scholar] [CrossRef] [PubMed]
- Peivandi, S.; Abasali, K.; Narges, M. Effects of L-carnitine on infertile men’s spermogram; a randomised clinical trial. J. Reprod. Infertil. 2010, 10, 331. [Google Scholar]
- Costa, M.; Canale, D.; Filicori, M.; D’lddio, S.; Lenzi, A. Lcarnitine in idiopathic asthenozoospermia: A multi-center study. Italian study group on carnitine and male infertility. Andrologia 1994, 26, 155–159. [Google Scholar] [CrossRef]
- Moncada, M.-L.; Vicari, E.; Cimino, C.; Calogero, A.E.; Mongio, A.; D’Agata, R. Effect of acetylcarnitine treatment in oligoasthenospermic patients. Acta Eur. Fertil. 1992, 23, 221–224. [Google Scholar] [PubMed]
- Garolla, A.; Maiorino, M.; Roverato, A.; Roveri, A.; Ursini, F.; Foresta, C. Oral carnitine supplementation in-creases sperm motility in asthenozoospermic men with normal sperm phospholipid hydroperoxide glutathione peroxidase levels. Fertil. Steril. 2005, 83, 355–361. [Google Scholar] [CrossRef]
- Zhou, X.; Liu, F.; Zhai, S. Effect of L-carnitine and/or L-acetyl-carnitine in nutrition treatment for male infertility: A systematic review. Asia Pac. J. Clin. Nutr. 2007, 16, 383–390. [Google Scholar] [PubMed]
- Miroueh, A. Effect of arginine on oligospermia. Fertil. Steril. 1970, 21, 217–219. [Google Scholar] [CrossRef]
- Govil, G.; Phadke, R.S.; Srivastava, S. Lipid Soluble Antioxidants: Biochemistry and Clinical Applications; Ong, A.S.H., Packer, L., Eds.; Birkhauser Verlag: Basel, Switzerland, 1992; pp. 27–46. [Google Scholar]
- Palmer, R.M.J.; Ashton, D.S.; Moncada, S. Vascular endothelial cells synthesize nitric oxide from L-arginine. Nature 1988, 333, 664–666. [Google Scholar] [CrossRef]
- Moncada, S.; Palmer, R.M.; Higgs, E.A. Nitric oxide: Physiology, pathophysiology, and pharmacology. Pharmacol. Rev. 1991, 43, 109–142. [Google Scholar]
- Zini, A.; De Lamirande, E.; Gagnon, C. Low levels of nitric oxide promote human sperm capacitation in vitro. J. Androl. 1995, 16, 424–431. [Google Scholar] [PubMed]
- Aitken, R.J.; Paterson, M.; Fisher, H.; Buckingham, D.; van Duin, M. Redox regulation of tyrosine phosphorylation in human spermatozoa and its role in the control of human sperm function. J. Cell Sci. 1995, 108, 2017–2025. [Google Scholar] [CrossRef] [PubMed]
- Srivastava, S.; Desai, P.; Coutinho, E.; Govil, G. Mechanism of action of L-arginine on the vitality of spermatozoa is primarily through increased biosynthesis of nitric oxide. Biol. Reprod. 2006, 74, 954–958. [Google Scholar] [CrossRef] [PubMed][Green Version]
- Keller, D.W.; Polakoski, K.L. L-arginine stimulation of human sperm motility in vitro. Biol. Reprod. 1975, 13, 154–157. [Google Scholar] [CrossRef]
- Srivastava, S.; Desai, P.; Coutinho, E.; Govil, G. Protective effect of L-arginine against lipid peroxidation in goat epi-didymal spermatozoa. Physiol. Chem. Phys. Med. NMR 2000, 32, 127–135. [Google Scholar]
- Scibona, M.; Meschini, P.; Capparelli, S.; Pecori, C.; Rossi, P.; Fabris, G.F.M. Arginine and male infertility. Minerva Urol. Nefrol. 1994, 46, 251–253. [Google Scholar]
- Aydin, S.; Inci, O.; Alagol, B. The role of arginine indomethacin and kallikrein in the treatment of oligospermia. Int. Urol. Nephrol. 1995, 27, 199–202. [Google Scholar] [CrossRef]
- Stanislavov, R.; Nikolova, V.; Rohdewald, P. Improvement of seminal parameters with Prelox®: A randomized, double-blind, placebo-controlled, cross-over trial. Phytother. Res. 2009, 23, 297–302. [Google Scholar] [CrossRef]
- Dodd, S.; Dean, O.; Copolov, D.L.; Malhi, G.S.; Berk, M. N-acetylcysteine for antioxidant therapy: Pharmacology and clinical utility. Expert Opin. Biol. Ther. 2008, 8, 1955–1962. [Google Scholar] [CrossRef]
- Vanderbist, F.; Maes, P.; Neve, J. In vitro comparative assessment of the antioxidant activity of nacystelyn against three reactive oxygen species. Arzneimittelforschung 1996, 46, 783–788. [Google Scholar]
- Kondo, H.; Takahashi, M.; Niki, E. Peroxynitrite-induced hemolysis of human erythrocytes and its inhibition by antioxidants. FEBS Lett. 1997, 413, 236–238. [Google Scholar] [CrossRef]
- Sueishi, Y.; Hori, M.; Ishikawa, M.; Matsu-Ura, K.; Kamogawa, E.; Honda, Y.; Kita, M.; Ohara, K. Scavenging rate constants of hydrophilic antioxidants against multiple reactive oxygen species. J. Clin. Biochem. Nutr. 2014, 54, 67–74. [Google Scholar] [CrossRef]
- Ates, B.; Abraham, L.; Ercal, N. Antioxidant and free radical scavenging properties of N-acetylcysteine amide (NACA) and comparison with N-acetylcysteine (NAC). Free Radic. Res. 2008, 42, 372–377. [Google Scholar] [CrossRef] [PubMed]
- Sagrista, M.L.; Garcıa, A.E.; De Madariaga, M.A.; Mora, M. Antioxidant and pro-oxidant effect of the thiolic com-pounds N-acetyl-L-cysteine and glutathione against free radical-induced lipid peroxidation. Free Radic. Res. 2002, 36, 329–340. [Google Scholar] [CrossRef]
- Hoy, A.; Leininger-Muller, B.; Kutter, D.; Siest, G.; Visvikis, S. Growing significance of myeloperoxidase in non-infectious diseases. Clin. Chem. Lab. Med. 2002, 40, 2–8. [Google Scholar] [CrossRef]
- Deponte, M. Glutathione catalysis and the reaction mechanisms of glutathione-dependent enzymes. Biochim. Biophys. Acta BBA Gen. Subj. 2013, 1830, 3217–3266. [Google Scholar] [CrossRef]
- Nagy, P. Kinetics and mechanisms of thiol–disulfide exchange covering direct substitution and thiol oxidation-mediated pathways. Antioxid. Redox Signal. 2013, 18, 1623–1641. [Google Scholar] [CrossRef]
- Aldini, G.; Altomare, A.; Baron, G.; Vistoli, G.; Carini, M.; Borsani, L.; Sergio, F. N-Acetylcysteine as an antioxidant and disulphide breaking agent: The reasons why. Free Radic. Res. 2018, 52, 751–762. [Google Scholar] [CrossRef]
- Elnagar, A.M.B.; Ibrahim, A.; Soliman, A.M. Histopathological effects of titanium dioxide nanoparticles and the possible protective role of n-acetylcysteine on the testes of male albino rats. Int. J. Fertil. Steril. 2018, 12, 249–256. [Google Scholar] [CrossRef]
- Da Silva, R.F.; Borges Cdos, S.; Villela e Silva, P.; Missassi, G.; Kiguti, L.R.; Pupo, A.S.; Barbosa, F., Jr.; Anselmo-Franci, J.A.; Kempinas, W.D.G. The coadministration of n-acetylcysteine ameliorates the effects of arsenic trioxide on the male mouse genital system. Oxid. Med. Cell. Longev. 2016, 2016, 4257498. [Google Scholar] [CrossRef] [PubMed]
- Kheradmandi, R.; Jorsaraei, S.G.A.; Feizi, F.; Moghadamnia, A.A.; Neamati, N. Protective effect of n-acetyl cysteine on chlorpyrifos-induced testicular toxicity in mice. Int. J. Fertil. Steril. 2019, 13, 51–56. [Google Scholar] [PubMed]
- Sarafian, T.A.; Montes, C.; Imura, T.; Qi, J.; Coppola, G.; Geschwind, D.H.; Sofroniew, M.V. Disruption of astrocyte STAT3 signaling decreases mitochondrial function and increases oxidative stress in vitro. PLoS ONE 2010, 5, e9532. [Google Scholar] [CrossRef] [PubMed]
- Zhou, H.; Newnum, A.B.; Martin, J.R.; Li, P.; Nelson, M.T.; Moh, A.; Fu, X.Y.; Yokota, H.; Li, J. Osteo-blast/osteocyte-specific inactivation of STAT3 decreases load-driven bone formation and accumulates reactive oxygen species. Bone 2011, 49, 404–411. [Google Scholar] [CrossRef] [PubMed]
- Lachance, C.; Goupil, S.; Tremblay, R.R.; Leclerc, P. The immobilization of human spermatozoa by STAT3 inhibitory compound V results from an excessive intracellular amount of reactive oxygen species. Andrology 2015, 4, 133–142. [Google Scholar] [CrossRef] [PubMed]
- Jannatifar, R.; Parivar, K.; Roodbari, N.H.; Nasr-Esfahani, M.H. Effects of N-acetyl-cysteine supplementation on sperm quality, chromatin integrity and level of oxidative stress in infertile men. Reprod. Biol. Endocrinol. 2019, 17, 24. [Google Scholar] [CrossRef]
- Barekat, F.; Tavalaee, M.; Deemeh, M.R.; Bahreinian, M.; Azadi, L.; Abbasi, H.; Rozbahani, S.; Nasr-Esfahani, M.H. A pre-liminary study: N-acetyl-L-cysteine improves semen quality following varicocelectomy. Int. J. Fertil. Steril. 2016, 10, 120–126. [Google Scholar]
- Safarinejad, M.R.; Safarinejad, S. Efficacy of selenium and/or N-acetyl-cysteine for improving semen parameters in infertile men: A double-blind, placebo controlled, randomized study. J. Urol. 2009, 181, 741–751. [Google Scholar] [CrossRef]
- Ciftci, H.; Verit, A.; Savas, M.; Yeni, E.; Erel, O. Effects of N-acetylcysteine on semen parameters and oxidative/antioxidant status. Urology 2009, 74, 73–76. [Google Scholar] [CrossRef]
- Wolfram, T.; Schwarz, M.; Reuß, M.; Lossow, K.; Ost, M.; Klaus, S.; Schwerdtle, T.; Kipp, A.P. N-acetylcysteine as modulator of the essential trace elements copper and zinc. Antioxidants 2020, 9, 1117. [Google Scholar] [CrossRef]
- Frémont, L. Biological effects of resveratrol. Life Sci. 2000, 66, 663–673. [Google Scholar] [CrossRef]
- Baur, J.; Sinclair, D. Therapeutic potential of resveratrol: The in vivo evidence. Nat. Rev. Drug Discov. 2006, 5, 493–506. [Google Scholar] [CrossRef] [PubMed]
- Dolinsky, V.W.; Chan, A.Y.; Robillard Frayne, I.; Light, P.E.; Des Rosiers, C.; Dyck, J.R. Resveratrol prevents the prohypertrophic effects of oxidative stress on LKB1. Circulation 2009, 119, 1643–1652. [Google Scholar] [CrossRef] [PubMed]
- Spanier, G.; Xu, H.; Xia, N.; Tobias, S.; Deng, S.; Wojnowski, L.; Forstermann, U.; Li, H. Resveratrol reduces endothelial oxidative stress by modulating the gene expression of superoxide dismutase 1 (SOD1), glutathione peroxidase 1 (GPx1) and NADPH oxidase subunit (Nox4). J. Physiol. Pharmacol. 2009, 60, 111–116. [Google Scholar]
- Tanno, M.; Kuno, A.; Yano, T.; Miura, T.; Hisahara, S.; Ishikawa, S.; Shimamoto, K.; Horio, Y. Induction of manganese superoxide dismutase by nuclear translocation and activation of SIRT1 promotes cell survival in chronic heart failure. J. Biol. Chem. 2010, 285, 8375–8382. [Google Scholar] [CrossRef] [PubMed]
- Ungvari, Z.; Orosz, Z.; Rivera, A.; Labinskyy, N.; Xiangmin, Z.; Olson, S. Resveratrol increases vascular oxidative stress resistance. Am. J. Physiol. Heart Circ. Physiol. 2007, 292, 2417–2424. [Google Scholar] [CrossRef]
- Mojica-Villegas, M.A.; Izquierdo-Vega, J.A.; Chamorro-Cevallos, G.; Sánchez-Gutiérrez, M. Protective effect of resveratrol on biomarkers of oxidative stress induced by iron/ascorbate in mouse spermatozoa. Nutrients 2014, 6, 489–503. [Google Scholar] [CrossRef]
- Lv, Z.M.; Wang, Q.; Chen, Y.H.; Wang, S.H.; Huang, D.Q. Resveratrol attenuates inflammation and oxidative stress in epididymal white adipose tissue: Implications for its involvement in improving steroidogenesis in diet-induced obese mice. Mol. Reprod. Dev. 2015, 82, 321–328. [Google Scholar] [CrossRef]
- Albuquerque, R.V.; Malcher, N.S.; Amado, L.L.; Coleman, M.D.; Dos Santos, D.C.; Borges, R.S. In vitro protective effect and antioxidant mechanism of resveratrol induced by Dapsone hydroxylamine in human cells. PLoS ONE 2015, 10, e0134768. [Google Scholar]
- Zini, R.; Morin, C.; A Bertelli, A.; Tillement, J.-P. Effects of resveratrol on the rat brain respiratory chain. Drugs Exp. Clin. Res. 1999, 25, 87–97. [Google Scholar]
- Berrougui, H.; Grenier, G.; Loued, S.; Drouin, G.; Khalil, A. A new insight into resveratrol as an atheroprotective compound: Inhibition of lipid peroxidation and enhancement of cholesterol efflux. Atherosclerosis 2009, 207, 420–427. [Google Scholar] [CrossRef]
- López-Vélez, M.; Martinez-Martinez, F.; Del Valle-Ribes, C. The study of phenolic compounds as natural antioxidants in wine. Crit. Rev. Food Sci. Nutr. 2003, 43, 233–244. [Google Scholar] [CrossRef] [PubMed]
- Park, S.-J.; Ahmad, F.; Philp, A.; Baar, K.; Williams, T.; Luo, H.; Ke, H.; Rehmann, H.; Taussig, R.; Brown, A.L.; et al. Resveratrol ameliorates aging-related metabolicphenotypes by inhibiting cAMP phosphodiesterases. Cell 2012, 148, 421–433. [Google Scholar] [CrossRef]
- Ecroyd, H.; Jones, R.C.; Aitken, R.J. Endogenous redox activity in mouse spermatozoa and its role in regulating the tyrosine phosphorylation events associated with sperm capacitation1. Biol. Reprod. 2003, 69, 347–354. [Google Scholar] [CrossRef]
- Nashtaei, M.S.; Amidi, F.; Gilani, M.A.S.; Aleyasin, A.; Bakhshalizadeh, S.; Naji, M.; Nekoonam, S. Protective features of resveratrol on human spermatozoa cryopreservation may be mediated through 5′ AMP-activated protein kinase activation. Andrology 2017, 5, 313–326. [Google Scholar] [CrossRef]
- Garcez, M.E.; Branco, C.D.S.; Lara, L.V.; Pasqualotto, F.F.; Salvador, M. Effects of resveratrol supplementation on cryopreservation medium of human semen. Fertil. Steril. 2010, 94, 2118–2121. [Google Scholar] [CrossRef]
- Archana, D.; Supriya, C.; Girish, B.; Kishori, B.; Reddy, P.S. Alleviative effect of resveratrol on polyvinyl chloride-induced reproductive toxicity in male Wistar rats. Food Chem. Toxicol. 2018, 116, 173–181. [Google Scholar] [CrossRef] [PubMed]
- Katen, A.L.; Stanger, S.J.; Anderson, A.L.; Nixon, B.; Roman, S.D. Chronic acrylamide exposure in male mice induces DNA damage to spermatozoa; potential for amelioration by resveratrol. Reprod. Toxicol. 2016, 63, 1–12. [Google Scholar] [CrossRef]
- Ourique, G.M.; Pês, T.S.; Saccol, E.M.; Finamor, I.A.; Glanzner, W.G.; Baldisserotto, B.; Pavanato, M.A.; Gonçalves, P.B.; Barreto, K.P. Resveratrol prevents oxidative damage and loss of sperm motility induced by long-term treatment with valproic acid in Wistar rats. Exp. Toxicol. Pathol. 2016, 68, 435–443. [Google Scholar] [CrossRef] [PubMed]
- Simas, J.N.; Mendes, T.B.; Fischer, L.W.; Vendramini, V.; Miraglia, S.M. Resveratrol improves sperm DNA quality and reproductive capacity in type 1 diabetes. Andrology 2021, 9, 384–399. [Google Scholar] [CrossRef]
- Murthy, P.P.N. Structure and nomenclature of inositol phosphates, phosphoinositides, and glycosylphosphatidylinositols. Subcell. Biochem. 2006, 39, 1–19. [Google Scholar] [CrossRef] [PubMed]
- Thomas, M.P.; Potter, B.V.L. The enzymes of human diphosphoinositol polyphosphate metabolism. FEBS J. 2013, 281, 14–33. [Google Scholar] [CrossRef]
- Posternak, T. Recherches dans la serie des cyclites VI. Sut la configuration de la meso-inosite, de la scyllite et d’un inosose obtenu par voie biochimique (scyllo-ms-inosose). Helv. Chim. Acta 1942, 25, 746–752. [Google Scholar] [CrossRef]
- Clements, R.S., Jr.; Darnell, B. Myo-inositol content of common foods: Development of a high myo-inositol diet. Am. J. Clin. Nutr. 1980, 33, 1954–1967. [Google Scholar] [CrossRef] [PubMed]
- Lam, S.; McWilliams, A.; Leriche, J.; MacAulay, C.; Wattenberg, L.; Szabo, E. A phase i study of myo-inositol for lung cancer chemoprevention. Cancer Epidemiol. Biomark. Prev. 2006, 15, 1526–1531. [Google Scholar] [CrossRef] [PubMed]
- Reddy, N.R.; Sathe, S.K. Food Phytates; CRC Press: Boca Raton, FL, USA, 2002; pp. 25–51. [Google Scholar]
- Grases, F.; Simonet, B.M.; Prieto, R.M.; Costa-Bauzà, A.; March, J.G.; Shamsuddin, A.M. Absorption and excretion of orally ad-ministrered inositol hexaphosphate (IP (6) or phyteta) in humans. Biofactors 2001, 15, 53–61. [Google Scholar] [CrossRef] [PubMed]
- Vazquez-Levin, M.H.; Verón, G.L. Myo-inositol in health and disease: Its impact on semen parameters and male fertility. Andrology 2020, 8, 277–298. [Google Scholar] [CrossRef] [PubMed]
- Chauvin, T.R.; Griswold, M.D. Characterization of the expression and regulation of genes necessary for myo-inositol biosynthesis and transport in the seminiferous epithelium. Biol. Reprod. 2004, 70, 744–751. [Google Scholar] [CrossRef] [PubMed]
- Monastra, G.; Sambuy, Y.; Ferruzza, S.; Ferrari, D.; Ranaldi, G. Alpha-lactalbumin effect on myo-inositol intestinal absorption: In vivo and in vitro. Curr. Drug Deliv. 2018, 15, 1305–1311. [Google Scholar] [CrossRef] [PubMed]
- Kwon, H.; Yamauchi, A.; Uchida, S.; Preston, A.; Garcia-Perez, A.; Burg, M.; Handler, J. Cloning of the cDNa for a Na+/myo-inositol cotransporter, a hypertonicity stress protein. J. Biol. Chem. 1992, 267, 6297–6301. [Google Scholar] [CrossRef]
- Casarini, L.; Crépieux, P. Molecular mechanisms of action of FSH. Front. Endocrinol. 2019, 10, 305. [Google Scholar] [CrossRef]
- Naaby-Hansen, S.; Wolkowicz, M.J.; Klots, K.; Bush, L.A.; Westbrook, V.A.; Shibahara, H.; Shetty, J.; Coonrod, S.A.; Reddi, P.P.; Shannon, J.; et al. Co-localization of inositol triphosphate receptor and calreticulin in the equatorial segment and in membrabe bounded vesicles in the cutoplasmatic droplet of huma spermatozoa. Mol. Hum. Reprod. 2001, 7, 923–933. [Google Scholar] [CrossRef]
- Marchetti, P.; Ballot, C.; Jouy, N.; Thomas, P.; Marchetti, C. Influence of mitochondrial membrane potential of spermatozoa on in vitro fertilisation outcome. Andrologia 2012, 44, 136–141. [Google Scholar] [CrossRef]
- Condorelli, R.A.; La Vignera, S.; Bellanca, S.; Vicari, E.; Calogero, A.E. Myoinositol: Does it improve sperm mitochondrial function and sperm motility? Urology 2012, 79, 1290–1295. [Google Scholar] [CrossRef]
- Wassarman, P.M.; Litscher, E.S. 1 Sperm-egg recognition mechanisms in mammals. Curr. Top. Dev. Biol. 1995, 30, 1–19. [Google Scholar] [CrossRef] [PubMed]
- Tesarik, J. Appropriate timing of the acrosome reaction is a major requirement for the fertilizing spermatozoa. Hum. Reprod. 1989, 4, 957–961. [Google Scholar] [CrossRef]
- Condorelli, R.A.; Barbagallo, F.; Calogero, A.E.; Cannarella, R.; Crafa, A.; La Vignera, S. D-chiro-inositol improves sperm mitochondrial membrane potential: In vitro evidence. J. Clin. Med. 2020, 9, 1373. [Google Scholar] [CrossRef] [PubMed]
- Aquila, S.; Guido, C.; Santoro, A.; Gazzerro, P.; Laezza, C.; Baffa, M.; Ando, S.; Bifulco, M. Rimonabant (SR141716) induces metabolism and acquisition of fertilizing ability in human sperm. Br. J. Pharmacol. 2010, 159, 831–841. [Google Scholar] [CrossRef] [PubMed]
- Aquila, S.; Middea, E.; Catalano, S.; Marsico, S.; Lanzino, M.; Casaburi, I.; Barone, I.; Bruno, R.; Zupo, S.; Andò, S. Human sperm express a functional androgen receptor: Effects on PI3K/AKT pathway. Hum. Reprod. 2007, 22, 2594–2605. [Google Scholar] [CrossRef]
- Visconti, P.E.; Baley, J.L.; Moore, G.D.; Pan, D.; Olds-Clarke, P.; Kopf, G.S. Capacitation in mouse spermatozoa I. Correlation between the capacitation state and protein phosphorylation. Development 1995, 121, 1129–1137. [Google Scholar] [CrossRef]
- Guido, C.; Santoro, M.; De Amicis, F.; Perrotta, I.; Panza, S.; Rago, V.; Cesario, M.G.; Lanzino, M.; Aquila, S. Human sperm anatomy and endocrinology in varicocele: Role of androgen receptor. Reproduction 2014, 147, 589–598. [Google Scholar] [CrossRef]
- Montanaro, D.; Santoro, M.; Carpino, A.; Perrotta, I.; De Amicis, F.; Sirianni, R.; Rago, V.; Gervasi, S.; Aquila, S. Human sperm liver receptor homolog-1 (LRH-1) acts as a downstream target of the estrogen signaling pathway. J. Anat. 2015, 227, 541–549. [Google Scholar] [CrossRef] [PubMed]
- Orth, J.M. Proliferation of sertoli cells in fetal and postnatal rats: A quantitative autoradiographic study. Anat. Rec. Adv. Integr. Anat. Evol. Biol. 1982, 203, 485–492. [Google Scholar] [CrossRef]
- Griswold, M.D. The central role of Sertoli cells in spermatogenesis. Semin. Cell Dev. Biol. 1998, 9, 411–416. [Google Scholar] [CrossRef] [PubMed]
- De Kretser, D.M. Endocrinology of male infertility. Br. Med. Bull. 1979, 35, 187–192. [Google Scholar] [CrossRef] [PubMed]
- Lambert, A.; Talbot, J.A.; Mitchell, R.; Robertson, W.R. Inhibition of protein kinase C by staurosporine increases estrogen secretion by rat Sertoli cells. Acta Endocrinol. 1991, 125, 286–290. [Google Scholar] [CrossRef] [PubMed]
- Abramsson, L.; Duchek, M. Gonadotropins, testosterone and prolactin in men with abnormal semen findings and an evaluation of the hormone profile. Int. Urol. Nephrol. 1989, 21, 499–510. [Google Scholar] [CrossRef] [PubMed]
- Illingworth, P.J.; Groome, N.P.; Byrd, W.; E Rainey, W.; McNeilly, A.S.; Mather, J.P.; Bremner, W.J. Inhibin-B: A likely candidate for the physiologically important form of inhibin in men. J. Clin. Endocrinol. Metab. 1996, 81, 1321–1325. [Google Scholar] [CrossRef][Green Version]
- Hu, Y.A.; Huang, Y.F. A serum marker of spermatogenesis–inhibin B. Zhonghua Nan Ke Xue 2002, 8, 57–60. [Google Scholar] [PubMed]
- Colone, M.; Marelli, G.; Unfer, V.; Bozzuto, G.; Molinari, A.; Stringaro, A. Inositol activity in oligoasthenoteratospermia--an in vitro study. Eur. Rev. Med. Pharmacol. Sci. 2010, 14, 891–896. [Google Scholar]
- Condorelli, R.A.; La Vignera, S.; Di Bari, F.; Unfer, V.; E Calogero, A. Effects of myoinositol on sperm mitochondrial function in-vitro. Eur. Rev. Med. Pharmacol. Sci. 2011, 15, 129–134. [Google Scholar]
- Donnelly, E.T.; Lewis, S.E.; McNally, J.A.; Thompson, W. In vitro fertilization and pregnancy rates: The influence of sperm motility and morphology on IVF outcome. Fertil. Steril. 1998, 70, 305–314. [Google Scholar] [CrossRef]
- Hirano, Y.; Shibahara, H.; Obara, H. Relationships between sperm motility characteristics assessed by the computer-aided sperm analysis (CASA) and fertilization rates in vitro. J. Assist. Reprod. Genet. 2001, 18, 215–220. [Google Scholar] [CrossRef]
- Marchetti, C.; Obert, G.; Deffosez, A. Study of mitochondrial membrane potential, reactive oxygen species, DNA fragmentation and cell viability by fow cytometry in human sperm. Hum. Reprod. 2002, 17, 1257–1265. [Google Scholar] [CrossRef]
- Rubino, P.; Palini, S.; Chigioni, S.; Carlomagno, G.; Quagliariello, A.; De Stefani, S.; Baglioni, A.; Bulletti, C. Improving fertilization rate in ICSI cycles by adding myoinositol to the semen preparation procedures: A prospective, bicentric, randomized trial on sibling oocytes. J. Assist. Reprod. Genet. 2015, 32, 387–394. [Google Scholar] [CrossRef][Green Version]
- Artini, P.G.; Casarosa, E.; Carletti, E.; Monteleone, P.; Di Noia, A.; Di Berardino, O.M. In vitro effect of myo-inositol on sperm motility in normal and oligoasthenospermia patients undergoing in vitro fertilization. Gynecol. Endocrinol. 2016, 33, 109–112. [Google Scholar] [CrossRef] [PubMed]
- Scarselli, F.; Lobascio, A.M.; Terribile, M.; Casciani, V.; Greco, P.; Franco, G.; Minasi, M.G.; Greco, E. Analysis of MYO-Inositol effect on spermatozoa motility, in hyper viscous ejaculates and in patients with grades II and III varicocele. Arch. Ital. Urol. Androl. 2016, 88, 279–283. [Google Scholar] [CrossRef]
- Palmieri, M.; Papale, P.; Della Ragione, A.; Quaranta, G.; Russo, G.; Russo, S. In vitroantioxidant treatment of semen samples in assisted reproductive technology: Effects of myo-inositol on nemaspermic parameters. Int. J. Endocrinol. 2016, 2016, 2839041. [Google Scholar] [CrossRef]
- Mohammadi, F.; Varanloo, N.; Nasrabadi, M.H.; Vatannejad, A.; Amjadi, F.S.; Masroor, M.J.; Bajelan, L.; Mehdizadeh, M.; Aflatoonian, R.; Zandieh, Z. Supplementation of sperm freezing medium with myoinositol improve human sperm parameters and protects it against DNA fragmentation and apoptosis. Cell Tissue Bank. 2019, 20, 77–86. [Google Scholar] [CrossRef] [PubMed]
- Saleh, R.; Assaf, H.; El Maged, W.M.A.; Elsuity, M.; Fawzy, M. Increased cryo-survival rate in ejaculated human sperm from infertile men following pre-freeze in vitro myo-inositol supplementation. Clin. Exp. Reprod. Med. 2018, 45, 177–182. [Google Scholar] [CrossRef]
- Governini, L.; Ponchia, R.; Artini, P.G.; Casarosa, E.; Marzi, I.; Capaldo, A.; Luddi, A.; Piomboni, P. Respiratory mitochondrial efficiency and DNA oxidation in human sperm after in vitro myo-inositol treatment. J. Clin. Med. 2020, 9, 1638. [Google Scholar] [CrossRef]
- Calogero, A.E.; Gullo, G.; La Vignera, S.; Condorelli, R.A.; Vaiarelli, A. Myoinositol improves sperm parameters and serum reproductive hormones in patients with idiopathic infertility: A prospective double-blind randomized placebo-controlled study. Andrology 2015, 3, 491–495. [Google Scholar] [CrossRef] [PubMed]
- Gulino, F.A.; Leonardi, E.; Marilli, I.; Musmeci, G.; Vitale, S.G.; Leanza, V.; Palumbo, M.A. Effect of treatment with myo-inositol on semen parameters of patients undergoing an IVF cycle: In vivo study. Gynecol. Endocrinol. 2016, 32, 65–68. [Google Scholar] [CrossRef] [PubMed]
- Oliva, M.M.; Minutolo, E.; Lippa, A.; Iaconianni, P.; Vaiarelli, A. Effect of myoinositol and antioxidants on sperm quality in men with metabolic syndrome. Int. J. Endocrinol. 2016, 2016, 1674950. [Google Scholar] [CrossRef]
- Dinkova, A.; Martinov, D.; Konova, E. Efficacy of myo-inositol in the clinical management of patients with asthenozoospermia. Eur. Rev. Med. Pharmacol. Sci. 2017, 21, 62–65. [Google Scholar]
- Facchinetti, F.; Espinola, M.S.B.; Dewailly, D.; Ozay, A.C.; Prapas, N.; Vazquez-Levin, M.; Wdowiak, A.; Unfer, V.; Appetecchia, M.; Aragona, C.; et al. Breakthroughs in the use of inositols for assisted reproductive treatment (ART). Trends Endocrinol. Metab. 2020, 31, 570–579. [Google Scholar] [CrossRef] [PubMed]
- Facchinetti, F.; Unfer, V.; Dewailly, D.; Kamenov, Z.A.; Diamanti-Kandarakis, E.; Laganà, A.S.; Nestler, J.E.; Soulage, C.O. Inositols in polycystic ovary syndrome: An overview on the advances. Trends Endocrinol. Metab. 2020, 31, 435–447. [Google Scholar] [CrossRef] [PubMed]
- Unfer, V.; Carlomagno, G.; Papaleo, E.; Vailati, S.; Candiani, M.; Baillargeon, J.-P. Hyperinsulinemia alters myoinositol to d-chiroinositol ratio in the follicular fluid of patients with PCOS. Reprod. Sci. 2014, 21, 854–858. [Google Scholar] [CrossRef]
- Heimark, D.; McAllister, J.; Larner, J. Decreased myo-inositol to chiro-inositol (M/C) ratios and increased M/C epimerase activity in PCOS theca cells demonstrate increased insulin sensitivity compared to controls. Endocr. J. 2014, 61, 111–117. [Google Scholar] [CrossRef]
- Bevilacqua, A.; Carlomagno, G.; Gerli, S.; Montanino Oliva, M.; Devroey, P.; Lanzone, A.; Soulange, C.; Facchinetti, F.; Carlo Di Renzo, G.; Bizzarri, M. Results from the international consensus conference on myo-inositol and d-chiroinositol in obstetrics and gynecology–assisted reproduction technology. Gynecol. Endocrinol. 2015, 31, 441–446. [Google Scholar] [CrossRef]
- Huang, L.C.; Fonteles, M.C.; Houston, D.B.; Zhang, C.; Larner, J. Chiroinositol deficiency and insulin resistance. III. Acute glycogenic and hypoglycemic effects of two inositol phosphoglycan insulin mediators in normal and streptozotocin-diabetic rats in vivo. Endocrinology 1993, 132, 652–657. [Google Scholar] [CrossRef] [PubMed]
- Larner, J. D-chiro-inositol—Its functional role in insulin action and its deficit in insulin resistance. Int. J. Exp. Diabetes Res. 2002, 3, 47–60. [Google Scholar] [CrossRef] [PubMed]
- Sacchi, S.; Marinaro, F.; Tondelli, D.; Lui, J.; Xella, S.; Marsella, T.; Tagliasacchi, D.; Argento, C.; Tirelli, A.; Giulini, S.; et al. Modulation of gonadotrophin induced steroidogenic enzymes in granulosa cells by d-chiroinositol. Reprod. Biol. Endocrinol. 2016, 14, 52. [Google Scholar] [CrossRef]
- Stocco, C. Tissue physiology and pathology of aromatase. Steroids 2012, 77, 27–35. [Google Scholar] [CrossRef]
- Laganà, A.S.; Garzon, S.; Unfer, V. New clinical targets of d-chiro-inositol: Rationale and potential applications. Expert Opin. Drug Metab. Toxicol. 2020, 16, 703–710. [Google Scholar] [CrossRef]
- Oh, J.-N.; Hwang, J.Y.; Choi, K.-H.; Lee, C.-K. Treatment of aromatase (CYP19A1) inhibitor reduces fertility in porcine sperm. Zygote 2015, 24, 98–106. [Google Scholar] [CrossRef]
- Helo, S.; Ellen, J.; Mechlin, C.; Feustel, P.; Grossman, M.; Ditkoff, E.; McCullough, A. A randomized prospective double-blind comparison trial of clomiphene citrate and anastrozole in raising testosterone in hypogonadal infertile men. J. Sex. Med. 2015, 12, 1761–1769. [Google Scholar] [CrossRef] [PubMed]
- Clark, R.V.; Sherins, R.J. Treatment of men with idiopathic oligozoospermic infertility using the aromatase inhibitor, testolactone results of a double-blinded, randomized, placebo-controlled trial with crossover. J. Androl. 1989, 10, 240–247. [Google Scholar] [CrossRef]
- Shoshany, O.; Abhyankar, N.; Mufarreh, N.; Daniel, G.; Niederberger, C. Outcomes of anastrozole in oligozoospermic hypoandrogenic subfertile men. Fertil. Steril. 2017, 107, 589–594. [Google Scholar] [CrossRef] [PubMed]


{kind=link}
| Author and Publication Year | Samples | Treatments | Results |
|---|---|---|---|
| Colone et al., 2010 | OAT patients | Inositol 2 mg/mL and then submitted to scansion electron microscopy (SEM) and to transmission electron microscopy (TEM) | Absence of amorphous material and reduction of mitochondrial damage to the crests |
| Condorelli et al., 2011 | 5 normozoospermic and 7 OAT patients | Incubated in-vitro with 2 mg/mL of myo-inositol or placebo (control) for 2 h | Increased the number of spermatozoa with high MMP and decreased the number of those with low MMP in OAT patients |
| Condorelli et al., 2012 | 20 normozoospermic and 20 OAT patients | Incubated in vitro with 2 mg/mL of myo-inositol or phosphate-buffered saline as a control for 2 h | Increased sperm motility and the number of spermatozoa after swim-up and in OAT patients, the improvement was associated with sperm mitochondrial function. |
| Rubino et al., 2015 | Myo-inositol group (n = 262 oocytes), placebo group (n = 238 oocytes) | Washed and subjected to swim-up with 2 mg/mL of myo-inositol or placebo-supplemented medium for 30–60 min. Spermatozoa recovered used for ICSI. | Improved spermatozoa motility in swim-up selected samples, fertilization rate (%), grade A embryos on day 3. |
| Artini et al., 2017 | 31 normospermic e 32 OAT patients | 2 mg/mL MI and incubated 30 min at 37 °C | Improved total motile sperm concentration, progressive motile sperm concentration. |
| Scarselli et al., 2016 | 30 patients with grade II and III varicocele | Semen centrifuged at 1800 rpm/10 min, resuspended, and incubated with 2 mg/mL myo-inositol and 133 mg/mL myo-inositol in 9 mg/mL sodium chloride) for 15 min at 37 °C | Patients suffering from varicocele response in >60% of the samples |
| Palmieri et al., 2017 | 46 normospermic, 19 oligospermic, 15 asthenospermic patients | Semen supplemented with 15 µL/mL of myo-inositol incubated 15 min at 37 °C | Improved progressive and total motility |
| Mohammadi et al., 2019 | 40 normospermic patients | Semen divided into two aliquots ad cryopreserved: one with 2 mg/mL myo-inositol; one without myo-inositol (control) | Improved progressive and total motility, normal sperm morphology, reactive oxygen species, malondialdehyde, total antioxidant assay and DNA fragmentation |
| Saleh et al., 2018 | 41 samples: 15 normal and 26 abnormal | Semen samples supplemented with 1 mg myo-inositol to cryoprotectant | Total and progressive motility, cryo-Survival Rate |
| Pallotti et al., 2019 | 9 normokinetic semen samples with nonlinear progressive motility | Incubation with a solution of myo-inositol | Increased linear progressive motility, significant reduction in nonlinear progressive motility, increased curvilinear velocity |
| Governini et al., 2020 | 56 Caucasian males with possible causes of male infertility such as varicocele, cryptorchidism, endocrine disorders or systemic diseases | The aliquots were incubated with standard medium (untreated sample) or medium supplemented with myo-inositol at 20 mg/mL (treated sample) for 20 min. | Increase in sperm motility and in oxygen consumption, the main index of oxidative phosphorylation efficiency and ATP production, both in basal and in in vitro capacitated samples. |
| Author | Study Design and Patients | Treatments | Results |
|---|---|---|---|
| Calogero et al., 2015 | Double-blind, randomized, place-bo-controlled; 194 men with idiopathic infertility | Group 1 (n = 98) received 2 g of myo-inositol and 200 mcg of folic acid twice daily. Group 2 (n = 96) received one placebo sachet twice day for 3 months | MI significantly increased the percentage of acrosome-reacted spermatozoa, sperm concentration, and total count and progressive motility. In addition, reduced serum luteinizing hormone, follicle-stimulating hormone, and in-creased inhibin B concentration |
| Gulino et al., 2013 | Prospective study; 62 patients divided into three different groups: healthy fertile patients (Group A); patients with oligoasthenospermia (OA)–(Group B)–control group (CTR). | 4000 mg/die of MI and 400 µg of folic acid for 2 months | Increase of basal and after density-gradient separation method spermatozoa concentration in Group B, and a significant increase of spermatozoa count after density-gradient separation method in Group A |
| Montanino Oliva et al., 2016 | Prospective longitudinal study; 45 asthenospermic males | The patients were treated by a dietary supplement administered twice a day containing 1 g MI, 30 mg L-carnitine, L-arginine and vit-amin E, 55 μg selenium, and 200 μg folic acid | Improved spermatic, hormonal and metabolic parameters: HOMA index, SHBG, E2, LH, free and total testosterone, sperm concentration, motility and normal morphology |
| Dinkova et al. et al., 2017 | Prospective longitudinal study; 109 patients with astheno-zoospermia | 1 g myo-inositol, 30 mg of L-carnitine, L-arginine, and vitamin E, 55 mcg of selenium, and 200 mcg of folic acid twice a day for 3 months | A significant improvement in spermmotility was reported in 85.32% of the patients |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
De Luca, M.N.; Colone, M.; Gambioli, R.; Stringaro, A.; Unfer, V. Oxidative Stress and Male Fertility: Role of Antioxidants and Inositols. Antioxidants 2021, 10, 1283. https://doi.org/10.3390/antiox10081283
De Luca MN, Colone M, Gambioli R, Stringaro A, Unfer V. Oxidative Stress and Male Fertility: Role of Antioxidants and Inositols. Antioxidants. 2021; 10(8):1283. https://doi.org/10.3390/antiox10081283
Chicago/Turabian StyleDe Luca, Maria Nunzia, Marisa Colone, Riccardo Gambioli, Annarita Stringaro, and Vittorio Unfer. 2021. "Oxidative Stress and Male Fertility: Role of Antioxidants and Inositols" Antioxidants 10, no. 8: 1283. https://doi.org/10.3390/antiox10081283
APA StyleDe Luca, M. N., Colone, M., Gambioli, R., Stringaro, A., & Unfer, V. (2021). Oxidative Stress and Male Fertility: Role of Antioxidants and Inositols. Antioxidants, 10(8), 1283. https://doi.org/10.3390/antiox10081283