
Enhancement of the Antiobesity and Antioxidant Effect of Purple Sweet Potato Extracts and Enhancement of the Effects by Fermentation

 , ,
, ,  and
and 
Abstract
1. Introduction
2. Materials and Methods
2.1. Sample Preparation
2.2. Animals and PSP Administration
2.3. Histological Analysis
2.4. Measurement of Plasma Cholesterol
2.5. Glucose Tolerance Test
2.6. Cell Culture and Differentiation
2.7. Oil Red O Staining
2.8. Real-Time Reverse Transcription Polymerase Chain Reaction (RT-PCR)
2.9. Western Blot Analysis
2.10. Measurement of Total Polyphenolic, Flavonoid, and Anthocyanin Contents
2.10.1. Total Phenolic Content
2.10.2. Total Flavonoid Content
2.10.3. Total Anthocyanin Content
2.11. Antioxidant Analysis
2.12. Statistical Analysis
3. Results
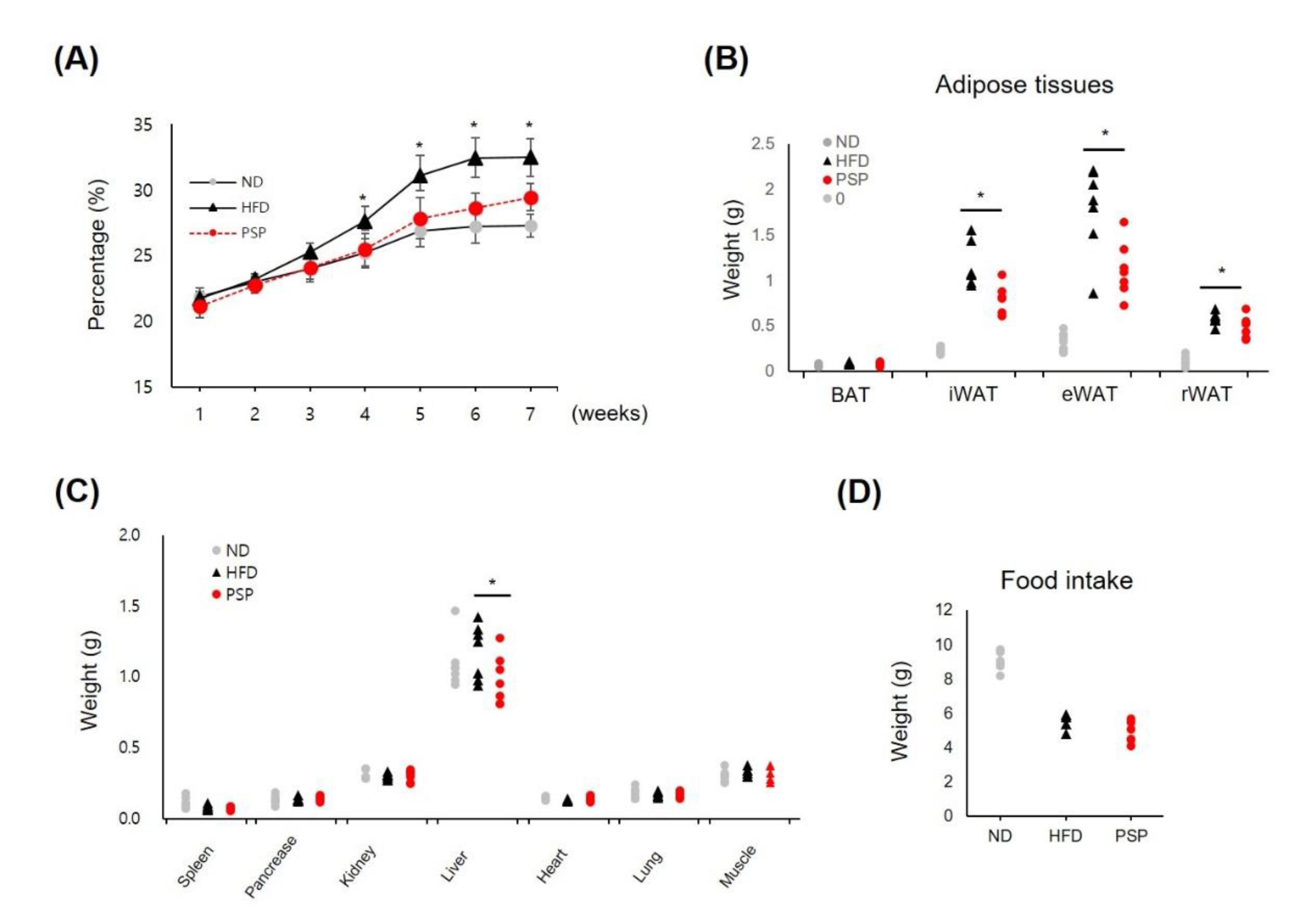
3.1. PSP Reduced Body Weight Gain and Suppressed Adipose Tissue Expansion in HFD-Induced Obese Mice
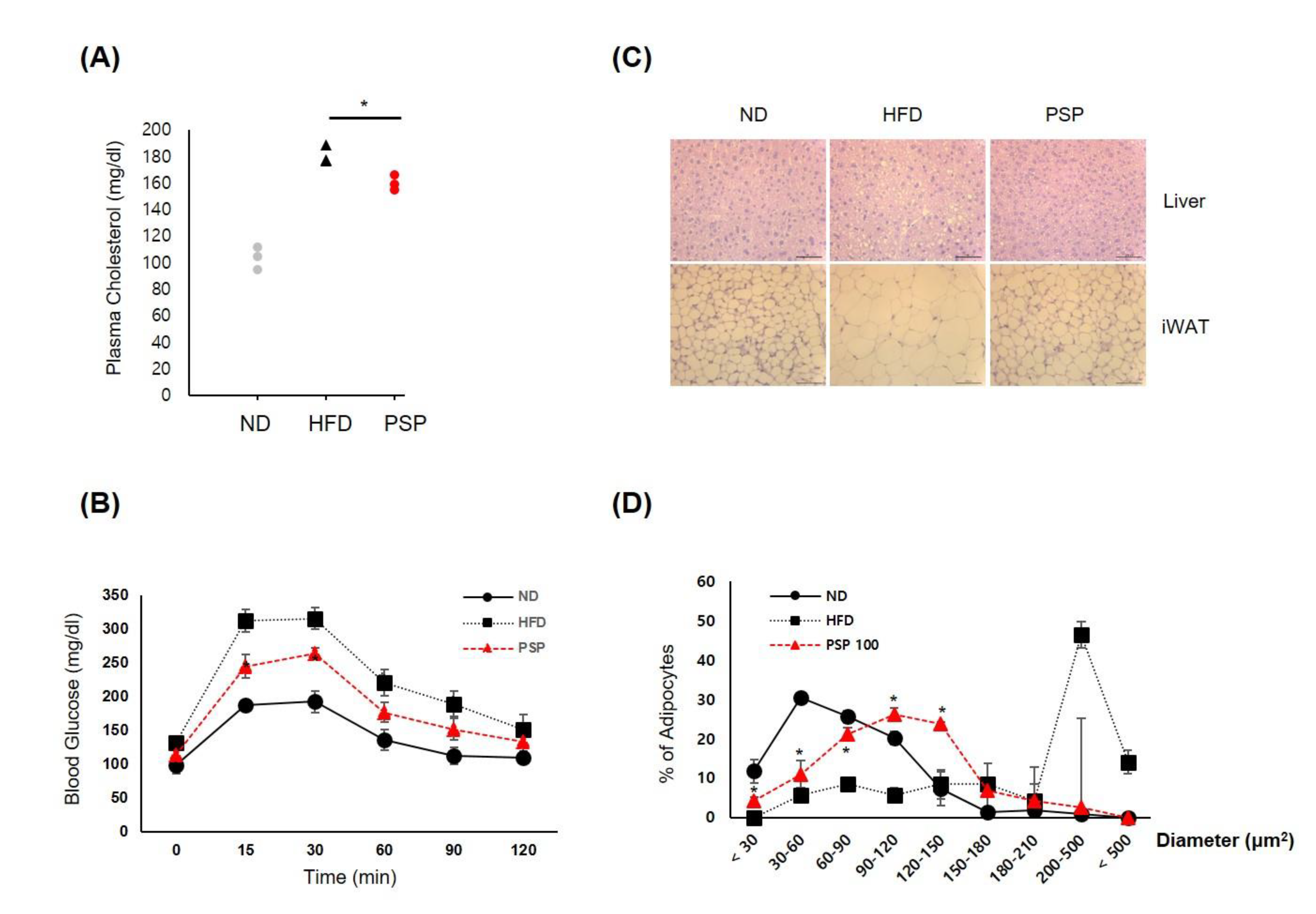
3.2. PSP Ameliorated the Metabolic Syndrome Associated with Obesity
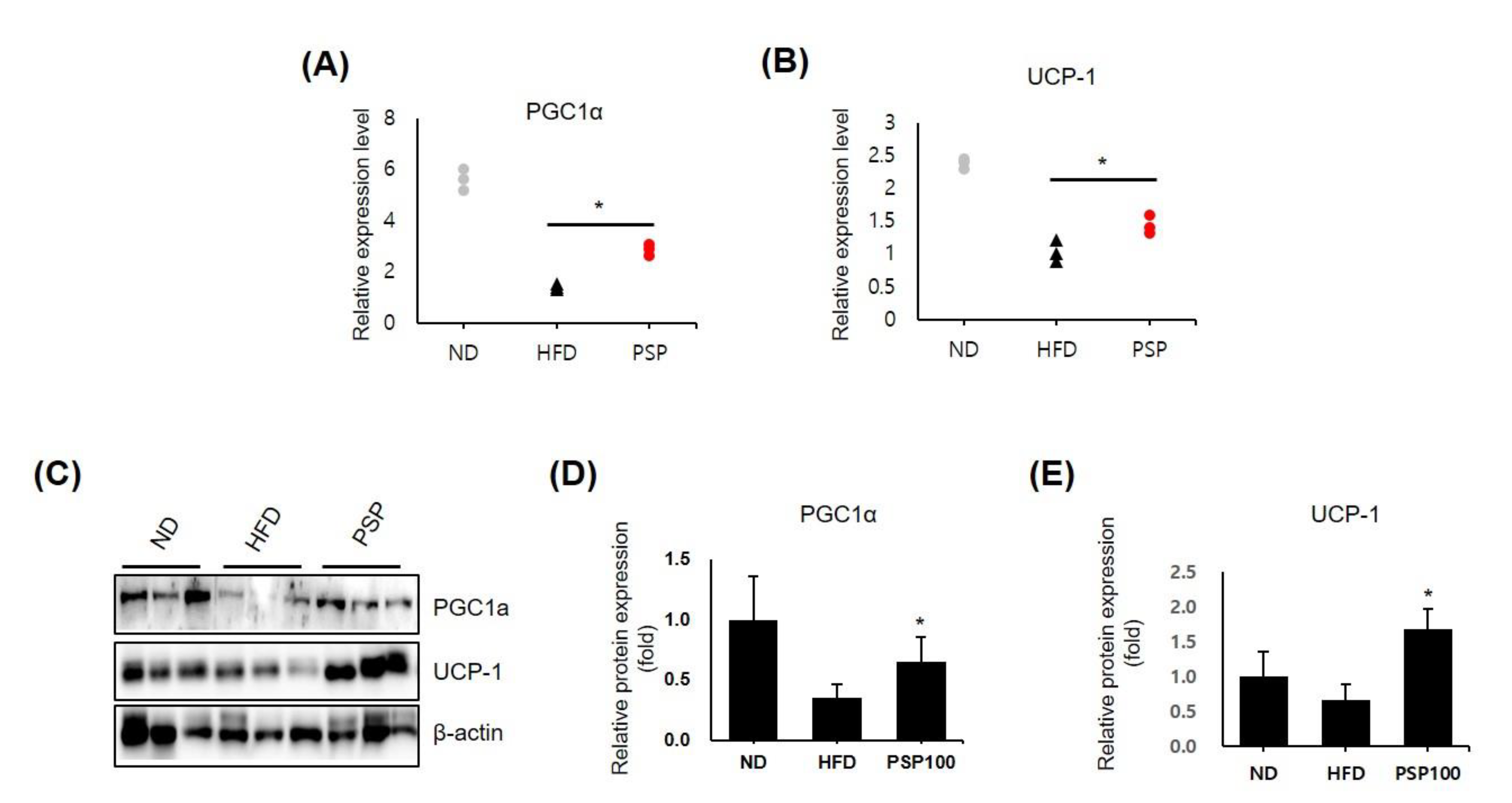
3.3. PSP Induced Browning Features in Adipose Tissue
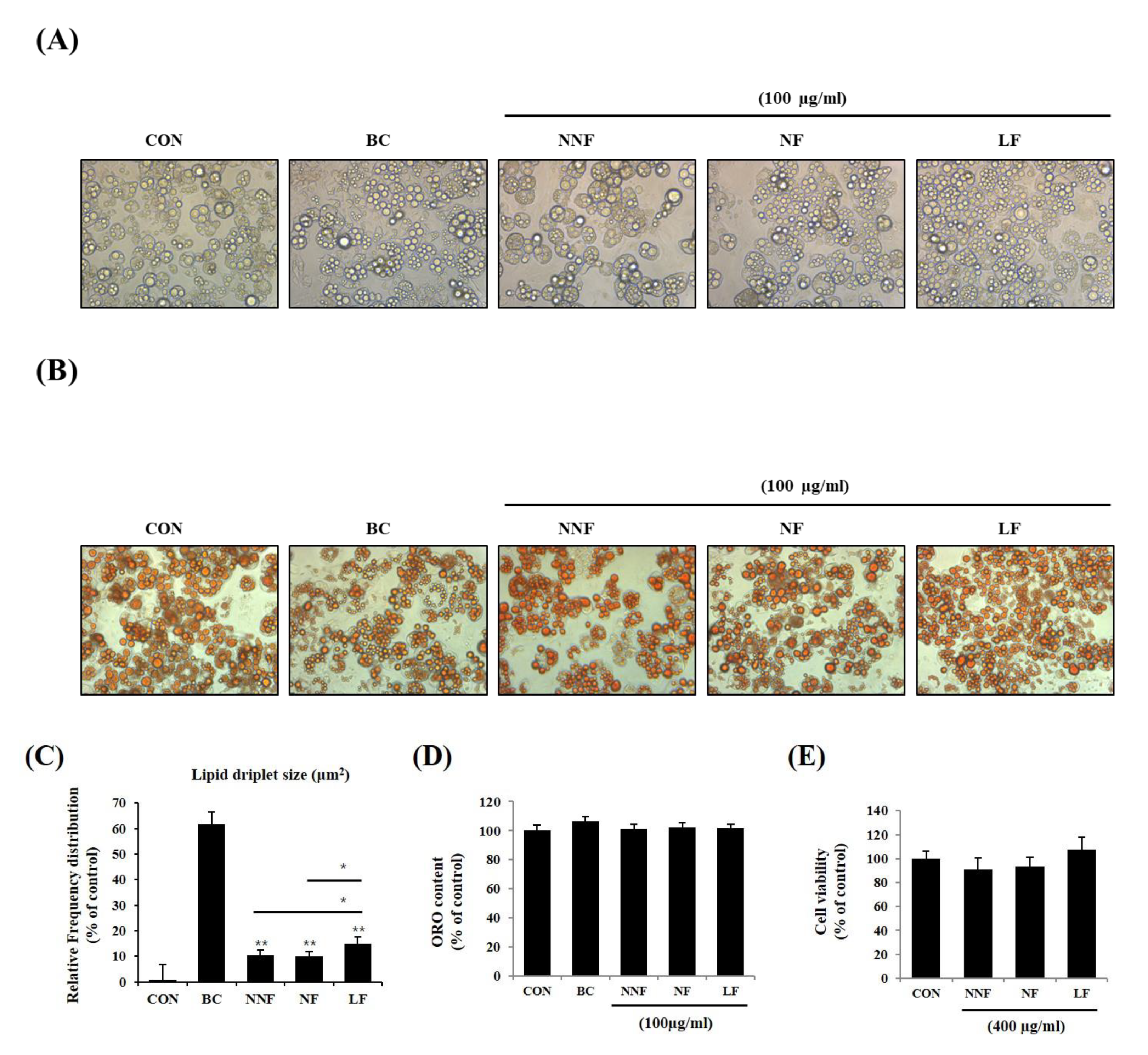
3.4. The Influence of PSP in the Differentiation of 3T3-L1 Adipocytes
3.5. PSPs Induce Brown Adipocyte-Like Phenotype in White Adipocytes
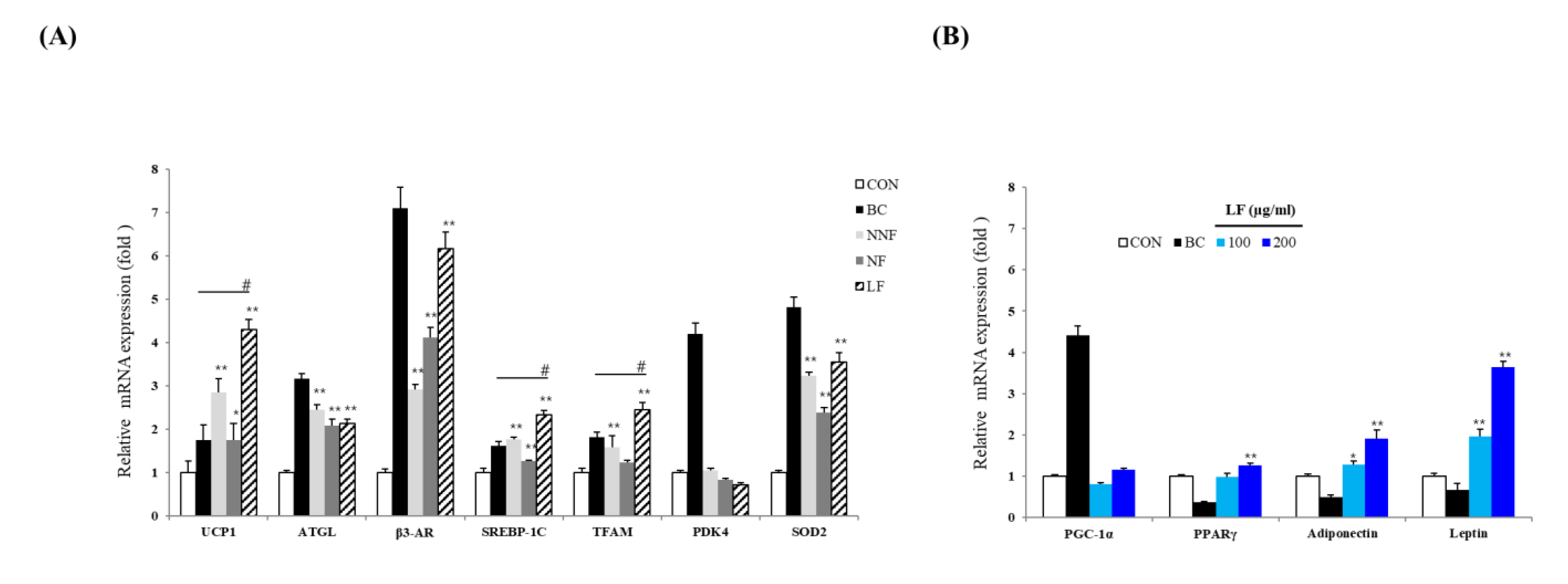
3.6. LF Increases mRNA Expression of Browning- and Adipogenesis-Related Genes in the 3T3-L1 Adipocytes
3.7. Effects of PSPs on Antioxidant Activities
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Vazquez-Vela, M.E.; Torres, N.; Tovar, A.R. White adipose tissue as endocrine organ and its role in obesity. Arch. Med. Res. 2008, 39, 715–728. [Google Scholar] [CrossRef]
- Townsend, K.L.; Tseng, Y.H. Brown fat fuel utilization and thermogenesis. Trends Endocrinol. Metab. 2014, 25, 168–177. [Google Scholar] [CrossRef] [PubMed]
- Choi, M.; Mukherjee, S.; Kang, N.H.; Barkat, J.L.; Parray, H.A.; Yun, J.W. L-rhamnose induces browning in 3T3-L1 white adipocytes and activates HIB1B brown adipocytes. IUBMB Life 2018, 70, 563–573. [Google Scholar] [CrossRef] [PubMed]
- Vijgen, G.H.; Bouvy, N.D.; Teule, G.J.; Brans, B.; Hoeks, J.; Schrauwen, P.; van Marken Lichtenbelt, W.D. Increase in brown adipose tissue activity after weight loss in morbidly obese subjects. J. Clin. Endocrinol. Metab. 2012, 97, E1229–E1233. [Google Scholar] [CrossRef] [PubMed]
- Lone, J.; Choi, J.H.; Kim, S.W.; Yun, J.W. Curcumin induces brown fat-like phenotype in 3T3-L1 and primary white adipocytes. J. Nutr. Biochem. 2016, 27, 193–202. [Google Scholar] [CrossRef] [PubMed]
- Matsukawa, T.; Villareal, M.O.; Motojima, H.; Isoda, H. Increasing cAMP levels of preadipocytes by cyanidin-3-glucoside treatment induces the formation of beige phenotypes in 3T3-L1 adipocytes. J. Nutr. Biochem. 2017, 40, 77–85. [Google Scholar] [CrossRef] [PubMed]
- Baskaran, P.; Krishnan, V.; Ren, J.; Thyagarajan, B. Capsaicin induces browning of white adipose tissue and counters obesity by activating TRPV1 channel-dependent mechanisms. Br. J. Pharmacol. 2016, 173, 2369–2389. [Google Scholar] [CrossRef] [PubMed]
- Hwang, Y.P.; Choi, J.H.; Han, E.H.; Kim, H.G.; Wee, J.H.; Jung, K.O.; Jung, K.H.; Kwon, K.I.; Jeong, T.C.; Chung, Y.C.; et al. Purple sweet potato anthocyanins attenuate hepatic lipid accumulation through activating adenosine monophosphate-activated protein kinase in human HepG2 cells and obese mice. Nutr. Res. 2011, 31, 896–906. [Google Scholar] [CrossRef]
- Zhang, Z.F.; Fan, S.H.; Zheng, Y.L.; Lu, J.; Wu, D.M.; Shan, Q.; Hu, B. Purple sweet potato color attenuates oxidative stress and inflammatory response induced by d-galactose in mouse liver. Food Chem. Toxicol. 2009, 47, 496–501. [Google Scholar] [CrossRef]
- Ju, J.H.; Yoon, H.S.; Park, H.J.; Kim, M.Y.; Shin, H.K.; Park, K.Y.; Yang, J.O.; Sohn, M.S.; Do, M.S. Anti-obesity and antioxidative effects of purple sweet potato extract in 3T3-L1 adipocytes in vitro. J. Med. Food 2011, 14, 1097–1106. [Google Scholar] [CrossRef]
- Ahmadian, M.; Abbott, M.J.; Tang, T.; Hudak, C.S.; Kim, Y.; Bruss, M.; Hellerstein, M.K.; Lee, H.Y.; Samuel, V.T.; Shulman, G.I.; et al. Desnutrin/ATGL is regulated by AMPK and is required for a brown adipose phenotype. Cell Metab. 2011, 13, 739–748. [Google Scholar] [CrossRef]
- Limon, R.I.; Penas, E.; Torino, M.I.; Martinez-Villaluenga, C.; Duenas, M.; Frias, J. Fermentation enhances the content of bioactive compounds in kidney bean extracts. Food Chem. 2015, 172, 343–352. [Google Scholar] [CrossRef] [PubMed]
- Katina, K.; Liukkonen, K.-H.; Kaukovirta-Norja, A.; Adlercreutz, H.; Heinonen, S.-M.; Lampi, A.-M.; Pihlava, J.-M.; Poutanen, K. Fermentation-induced changes in the nutritional value of native or germinated rye. J. Cereal Sci. 2007, 46, 348–355. [Google Scholar] [CrossRef]
- Mashmoul, M.; Azlan, A.; Khaza’ai, H.; Yusof, B.N.; Noor, S.M. Saffron: A Natural Potent Antioxidant as a Promising Anti-Obesity Drug. Antioxidants 2013, 2, 293–308. [Google Scholar] [CrossRef]
- Abdul Rahman, H.; Saari, N.; Abas, F.; Ismail, A.; Mumtaz, M.W.; Abdul Hamid, A. Anti-obesity and antioxidant activities of selected medicinal plants and phytochemical profiling of bioactive compounds. Int. J. Food Prop. 2017, 20, 2616–2629. [Google Scholar] [CrossRef]
- Lee, H.S.; Lim, W.C.; Lee, S.J.; Lee, S.H.; Lee, J.H.; Cho, H.Y. Antiobesity Effect of Garlic Extract Fermented by Lactobacillus plantarum BL2 in Diet-Induced Obese Mice. J. Med. Food 2016, 19, 823–829. [Google Scholar] [CrossRef] [PubMed]
- Lee, S.G.; Kim, J.S.; Lee, H.S.; Lim, Y.M.; So, J.H.; Hahn, D.; Ha, Y.S.; Nam, J.O. Bioconverted Orostachys japonicas Extracts Suppress Angiogenic Activity of Ms-1 Endothelial Cells. Int. J. Mol. Sci. 2017, 18, 2615. [Google Scholar] [CrossRef]
- Lee, S.G.; Kim, J.S.; Kim, H.J.; Schlaepfer, D.D.; Kim, I.S.; Nam, J.O. Endothelial angiogenic activity and adipose angiogenesis is controlled by extracellular matrix protein TGFBI. Sci. Rep. 2021, 11, 9644. [Google Scholar] [CrossRef]
- Lee, S.G.; Lee, Y.J.; Jang, M.H.; Kwon, T.R.; Nam, J.O. Panax ginseng Leaf Extracts Exert Anti-Obesity Effects in High-Fat Diet-Induced Obese Rats. Nutrients 2017, 9, 999. [Google Scholar] [CrossRef]
- Lee, S.G.; Kim, J.S.; Min, K.; Kwon, T.K.; Nam, J.O. Hispidulin inhibits adipogenesis in 3T3-L1 adipocytes through PPARgamma pathway. Chem. Biol. Interact. 2018, 293, 89–93. [Google Scholar] [CrossRef]
- Kim, J.S.; Lee, S.G.; Min, K.; Kwon, T.K.; Kim, H.J.; Nam, J.O. Eupatilin inhibits adipogenesis through suppression of PPARgamma activity in 3T3-L1 cells. Biomed. Pharmacother. 2018, 103, 135–139. [Google Scholar] [CrossRef]
- Singleton, V.L.; Orthofer, R.; Lamuela-Raventós, R.M. [14] Analysis of total phenols and other oxidation substrates and antioxidants by means of folin-ciocalteu reagent. In Methods in Enzymology; Elsevier: Amsterdam, The Netherlands, 1999; Volume 299, pp. 152–178. [Google Scholar]
- Teng, H.; Lee, W.Y.; Choi, Y.H. Optimization of ultrasonic-assisted extraction of polyphenols, anthocyanins, and antioxidants from raspberry (Rubus coreanus Miq.) using response surface methodology. Food Anal. Methods 2014, 7, 1536–1545. [Google Scholar] [CrossRef]
- Teng, H.; Ghafoor, K.; Choi, Y.H. Optimization of microwave-assisted extraction of active components from Chinese quince using response surface methodology. J. Korean Soc. Appl. Biol. Chem. 2009, 52, 694–701. [Google Scholar] [CrossRef]
- Blois, M.S. Antioxidant Determinations by the Use of a Stable Free Radical. Nature 1958, 181, 1199–1200. [Google Scholar] [CrossRef]
- Kroon, T.; Harms, M.; Maurer, S.; Bonnet, L.; Alexandersson, I.; Lindblom, A.; Ahnmark, A.; Nilsson, D.; Gennemark, P.; O’Mahony, G.; et al. PPARγ and PPARα synergize to induce robust browning of white fat in vivo. Mol. Metab. 2020, 36, 100964. [Google Scholar] [CrossRef]
- Ferrari, A.; Longo, R.; Fiorino, E.; Silva, R.; Mitro, N. HDAC3 is a molecular brake of the metabolic switch supporting white adipose tissue browning. Nat. Commun. 2017, 8, 93. [Google Scholar] [CrossRef]
- Demine, S.; Tejerina, S.; Bihin, B.; Thiry, M.; Reddy, N.; Renard, P.; Raes, M.; Jadot, M.; Arnould, T. Mild mitochondrial uncoupling induces HSL/ATGL-independent lipolysis relying on a form of autophagy in 3T3-L1 adipocytes. J. Cell Physiol. 2018, 233, 1247–1265. [Google Scholar] [CrossRef] [PubMed]
- Giordano, A.; Frontini, A.; Cinti, S. Convertible visceral fat as a therapeutic target to curb obesity. Nat. Rev. Drug Discov. 2016, 15, 405–424. [Google Scholar] [CrossRef] [PubMed]
- Yashin, A.; Yashin, Y.; Xia, X.; Nemzer, B. Antioxidant Activity of Spices and Their Impact on Human Health: A Review. Antioxidants 2017, 6, 70. [Google Scholar] [CrossRef]
- Wang, Q.A.; Zhang, F.; Jiang, L.; Ye, R.; An, Y.; Shao, M.; Tao, C.; Gupta, R.K.; Scherer, P.E. PPARgamma and its Role in Adipocyte Homeostasis and Thiazolidinedione-Mediated Insulin Sensitization. Mol. Cell Biol. 2018. [Google Scholar] [CrossRef] [PubMed]
- Coles, C.A.; Maksimovic, J.; Wadeson, J.; Fahri, F.T.; Webster, T.; Leyton, C.; McDonagh, M.B.; White, J.D. Knockdown of a disintegrin A metalloprotease 12 (ADAM12) during adipogenesis reduces cell numbers, delays differentiation, and increases lipid accumulation in 3T3-L1 cells. Mol. Biol. Cell 2018, 29, 1839–1855. [Google Scholar] [CrossRef]
- Morata, A.; Benito, S.; Loira, I.; Palomero, F.; Gonzalez, M.C.; Suarez-Lepe, J.A. Formation of pyranoanthocyanins by Schizosaccharomyces pombe during the fermentation of red must. Int. J. Food Microbiol. 2012, 159, 47–53. [Google Scholar] [CrossRef]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Gene Name | Accession No. | Forward Primer | Reverse Primer |
|---|---|---|---|
| ATGL | NM_001163689.1 | AACGCCACTCACATCTACGG | GGACACCTCAATAATGTTGGCAC |
| β3-AR | NM_013462.3 | CCTTCAACCCGGTCATCTACTG | CGCACCTTCATAGCCATCAAA |
| SREBP-1c | NM_011480.4 | GCTGTTGGCATCCTGCTATC | TAGCTGGAAGTGACGGTGGT |
| TFAM | NM_009360.4 | CAAAGGATGATTCGGCTCAG | AAGCTGAATATATGCCTGCTTTTC |
| PDK4 | NM_013743.2 | CCGCTGTCCATGAAGCA | GCAGAAAAGCAAAGGAC |
| SOD2 | NM_013671.3 | ACCTGCCTTACGACTATGGC | CCACCATTGAACTTCAGTGC |
| PGC1α | XM_006503778.3 | CCCTGCCATTGTTAAGACC | TGCTGCTGTTCCTGTTTTC |
| PPARγ | AB644275.1 | GGAAGACCACTCGCATTCCTT | GTAATCAGCAACCATTGGGTCA |
| UCP-1 | NM_009463.3 | CTGCCAGGACAGTACCCAAG | TCAGCTGTTCAAAGCACACA |
| Adiponectin | NM_009605.4 | GATGGCACTCCTGGAGAGAA | TCTCCAGGCTCTCCTTTCCT |
| Leptin | NM_008493.3 | GGGCTTCACCCCATTCTGA | TGGCTATCTGCAGCACATTTTG |
| β-actin | EF095208 | CGTGCGTGACATCAAAGAGAA | GCTCGTTGCCAATAGTGATGA |
| IC50 (mg/mL) | ||
|---|---|---|
| ABTS assay | Ascorbic acid (standard) | 0.0687 ± 0.01 |
| NNF | 0.9231 ± 0.046 | |
| NF | 1.2861 ± 0.059 | |
| LF | 1.0098 ± 0.009 | |
| DPPH assay | Ascorbic acid (standard) | 0.0505 ± 0.001 |
| NNF | 1.0642 ± 0.051 | |
| NF | 1.5641 ± 0.104 | |
| LF | 1.0451 ± 0.078 | |
| Sample (µg/mL) | TPC (GAE µg/mL) | TFC (µg RE/mL) | TAC (µg/mL) | |
|---|---|---|---|---|
| NNF | 100 | 3.90 ± 0.60 | 35.65 ± 6.55 | 5.57 ± 1.51 |
| 200 | 7.08 ± 0.70 | 131.07 ± 2.91 | 9.69 ± 0.58 | |
| NF | 100 | 2.80 ± 0.20 | 1.74 ± 0.26 | 2.89 ± 1.02 |
| 200 | 6.22 ± 0.44 | 49.69 ± 3.84 | 7.12 ± 0.70 | |
| LF | 100 | 3.21 ± 0.17 | 34.68 ± 8.39 | 3.34 ± 0.58 |
| 200 | 7.89 ± 1.08 | 135.43 ± 1.45 | 7.24 ± 0.19 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Lee, S.G.; Chae, J.; Kim, D.S.; Lee, J.-B.; Kwon, G.-S.; Kwon, T.K.; Nam, J.-O. Enhancement of the Antiobesity and Antioxidant Effect of Purple Sweet Potato Extracts and Enhancement of the Effects by Fermentation. Antioxidants 2021, 10, 888. https://doi.org/10.3390/antiox10060888
Lee SG, Chae J, Kim DS, Lee J-B, Kwon G-S, Kwon TK, Nam J-O. Enhancement of the Antiobesity and Antioxidant Effect of Purple Sweet Potato Extracts and Enhancement of the Effects by Fermentation. Antioxidants. 2021; 10(6):888. https://doi.org/10.3390/antiox10060888
Chicago/Turabian StyleLee, Seul Gi, Jongbeom Chae, Dong Se Kim, Jung-Bok Lee, Gi-Seok Kwon, Taeg Kyu Kwon, and Ju-Ock Nam. 2021. "Enhancement of the Antiobesity and Antioxidant Effect of Purple Sweet Potato Extracts and Enhancement of the Effects by Fermentation" Antioxidants 10, no. 6: 888. https://doi.org/10.3390/antiox10060888
APA StyleLee, S. G., Chae, J., Kim, D. S., Lee, J.-B., Kwon, G.-S., Kwon, T. K., & Nam, J.-O. (2021). Enhancement of the Antiobesity and Antioxidant Effect of Purple Sweet Potato Extracts and Enhancement of the Effects by Fermentation. Antioxidants, 10(6), 888. https://doi.org/10.3390/antiox10060888