
A Brief Overview of Oxidative Stress in Adipose Tissue with a Therapeutic Approach to Taking Antioxidant Supplements




Abstract
1. Introduction
2. Overview of ROS
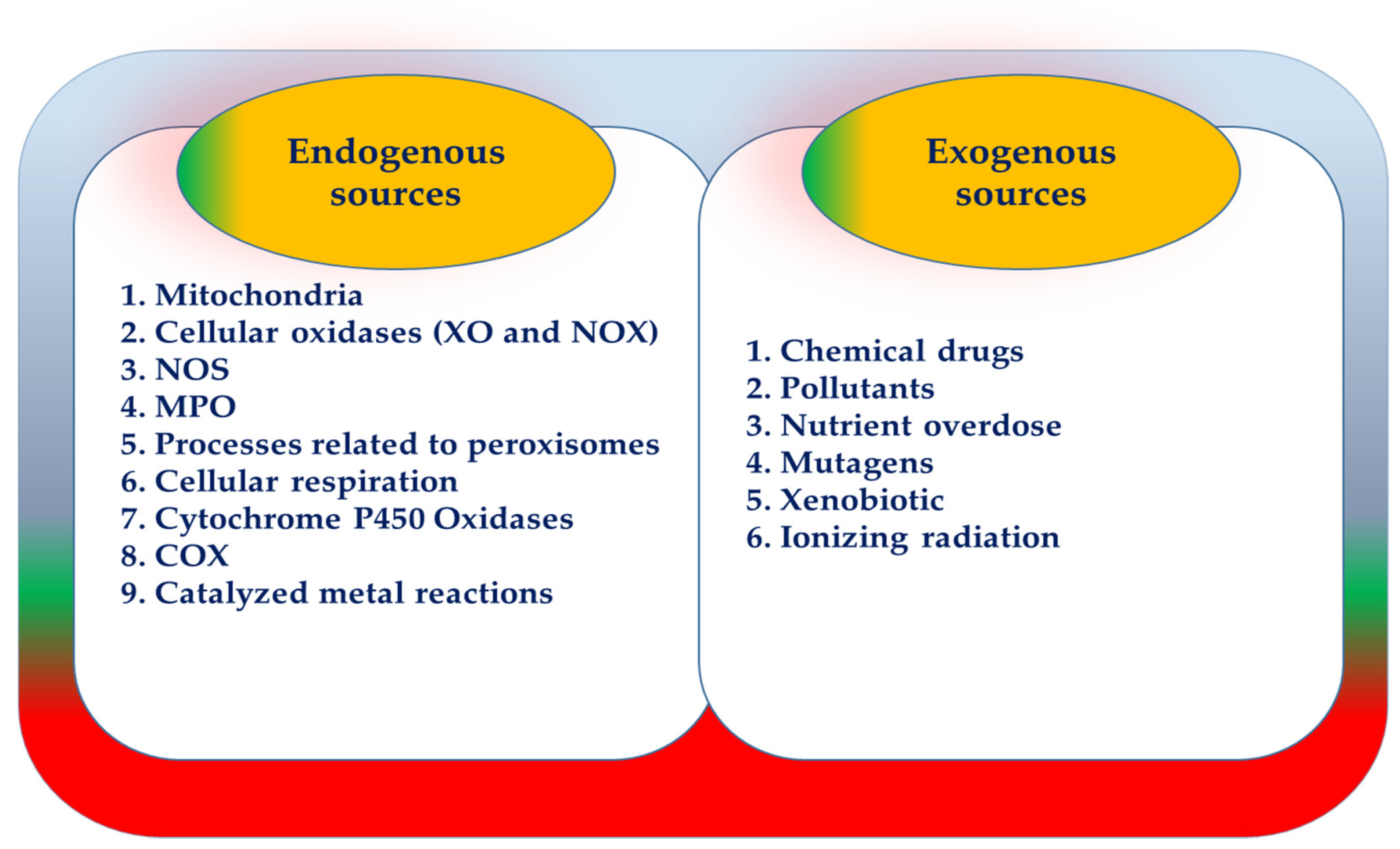
3. ROS Manufacturer Resources
3.1. ROS-Derived from Mitochondria
3.2. NOX
4. ROS Measuring Tools
4.1. MDA
4.2. 8-OHdG
4.3. 8-Nitroguanine (8-NO2-Gua)
4.4. Oxidative Products of Sugars
4.5. Reduced Glutathione (GSH): Oxidized Glutathione (GSSG) Ratio
5. A Brief Look at AT
6. Metabolic Disorders in AT
7. Antioxidants
7.1. SOD
7.2. GPx
7.3. CAT
8. Obesity, OS, and Antioxidant Supplementation
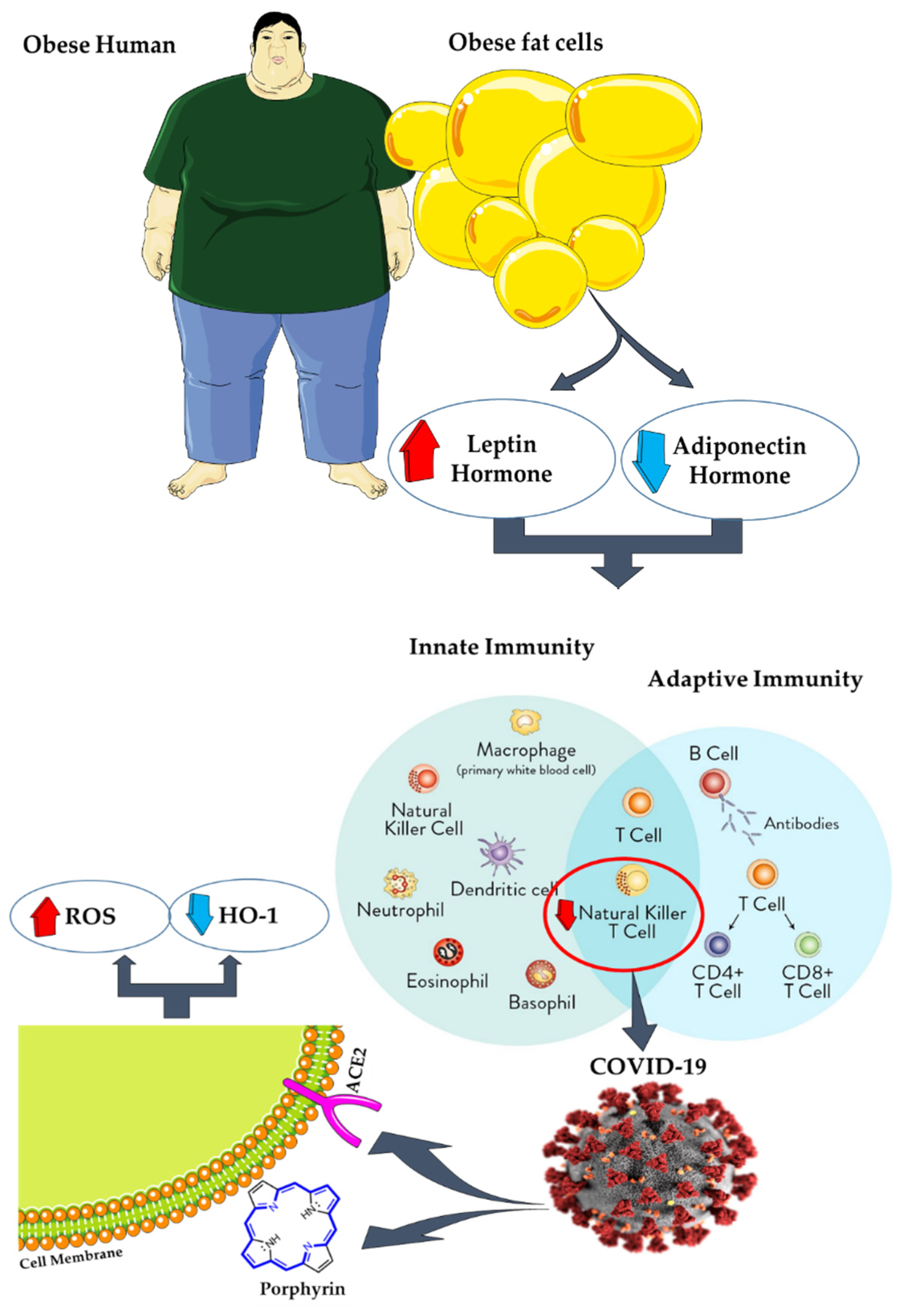
9. AT, Coronavirus Disease 2019 (COVID-19), and Antioxidants
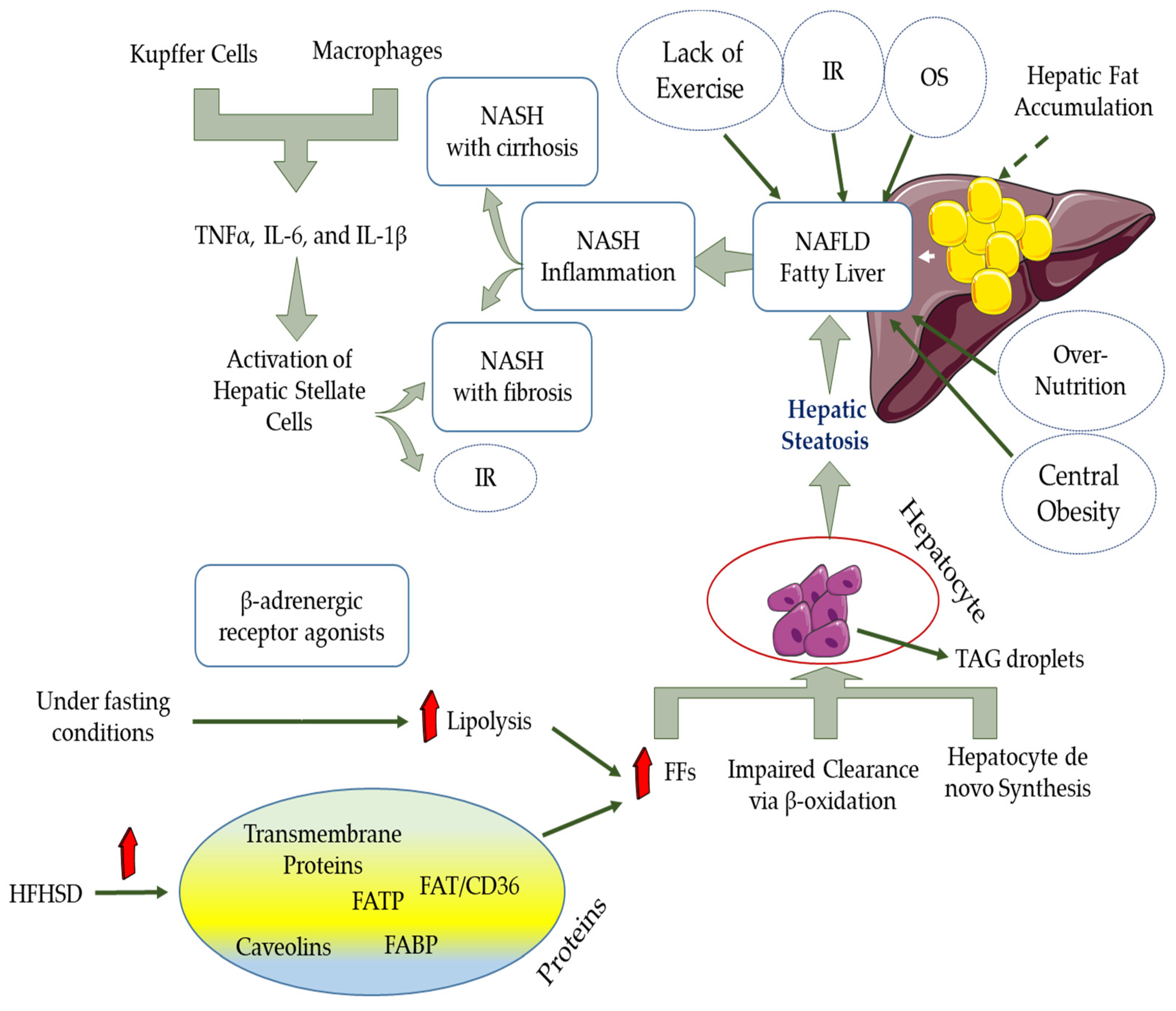
10. NAFLD and OS
11. Roles of Nutraceuticals as an Antioxidant in Reducing Oxidative Stress
12. Conclusions
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Tchang, B.G.; Saunders, K.H.; Igel, L.I. Best practices in the management of overweight and obesity. Med. Clin. N. Am. 2021, 105, 149–174. [Google Scholar] [CrossRef] [PubMed]
- Curley, S.; Gall, J.; Byrne, R.; Yvan-Charvet, L.; McGillicuddy, F.C. Metabolic inflammation in obesity—At the crossroads between fatty acid and cholesterol metabolism. Mol. Nutr. Food Res. 2020, 65, 1900482. [Google Scholar] [CrossRef] [PubMed]
- Goossens, G.H. The metabolic phenotype in obesity: Fat mass, body fat distribution, and adipose tissue function. Obes. Facts. 2017, 10, 207–215. [Google Scholar] [CrossRef]
- Payab, M.; Abedi, M.; Foroughi, H.N.; Hadavandkhani, M.; Arabi, M.; Tayanloo-Beik, A.; Sheikh Hosseini, M.; Gerami, H.; Khatami, F.; Larijani, B.; et al. Brown adipose tissue transplantation as a novel alternative to obesity treatment: A systematic review. Int. J. Obes. 2021, 45, 109–121. [Google Scholar] [CrossRef]
- Fernández-Sánchez, A.; Madrigal-Santillán, E.; Bautista, M.; Esquivel-Soto, J.; Morales-González, A.; Esquivel-Chirino, C.; Durante-Montiel, I.; Sánchez-Rivera, G.; Valadez-Vega, C.; Morales-González, J.A. Inflammation, oxidative stress, and obesity. Int. J. Mol. Sci. 2011, 12, 3117–3132. [Google Scholar] [CrossRef] [PubMed]
- Manna, P.; Jain, S.K. Obesity, oxidative stress, adipose tissue dysfunction, and the associated health risks: Causes and therapeutic strategies. Metab. Syndr. Relat. Disord. 2015, 13, 423–444. [Google Scholar] [CrossRef] [PubMed]
- Russo, L.; Lumeng, C.N. Properties and functions of adipose tissue macrophages in obesity. Immunology 2018, 155, 407–417. [Google Scholar] [CrossRef]
- Kardinaal, A.F.; van ‘t Veer, P.; Brants, H.A.; van den Berg, H.; van Schoonhoven, J.; Hermus, R.J. Relations between antioxidant vitamins in adipose tissue, plasma, and diet. Am. J. Epidemiol. 1995, 141, 440–450. [Google Scholar] [CrossRef]
- Boccellino, M.; D’Angelo, S. Anti-obesity effects of polyphenol intake: Current status and future possibilities. Int. J. Mol. Sci. 2020, 21, 5642. [Google Scholar] [CrossRef] [PubMed]
- Jakubczyk, K.; Dec, K.; Kałduńska, J.; Kawczuga, D.; Kochman, J.; Janda, K. Reactive oxygen species—Sources, functions, oxidative damage. Pol. Merkur. Lekarski. 2020, 48, 124–127. [Google Scholar]
- Bansal, M.; Kaushal, N. Oxidative Stress Mechanisms and Their Modulation; Springer: New Delhi, India, 2014. [Google Scholar] [CrossRef]
- Lü, J.M.; Lin, P.H.; Yao, Q.; Chen, C. Chemical and molecular mechanisms of antioxidants: Experimental approaches and model systems. J. Cell. Mol. Med. 2010, 14, 840–860. [Google Scholar] [CrossRef]
- Bloomer, R.J.; Goldfarb, A.H. Anaerobic exercise and oxidative stress: A review. Can. J. Appl. Physiol. 2004, 29, 245–263. [Google Scholar] [CrossRef]
- Oh, J.; Jung, S.R.; Lee, Y.J.; Park, K.W.; Han, J. Antioxidant and antiobesity activities of seed extract from campbell early grape as a functional ingredient. J. Food. Process. Pres. 2013, 37, 291–298. [Google Scholar] [CrossRef]
- Álvarez, E.; Rodiño-Janeiro, B.K.; Jerez, M.; Ucieda-Somoza, R.; Núñez, M.J.; González-Juanatey, J.R. Procyanidins from grape pomace are suitable inhibitors of human endothelial NADPH oxidase. J. Cell. Biochem. 2012, 113, 1386–1396. [Google Scholar] [CrossRef] [PubMed]
- Lefranc, C.; Friederich-Persson, M.; Palacios-Ramirez, R.; Cat, A.N.D. Mitochondrial oxidative stress in obesity: Role of the mineralocorticoid receptor. J. Endocrinol. 2018, 238, R143–R159. [Google Scholar] [CrossRef] [PubMed]
- Castro, J.P.; Grune, T.; Speckmann, B. The two faces of reactive oxygen species (ROS) in adipocyte function and dysfunction. Biol. Chem. 2016, 397, 709–724. [Google Scholar] [CrossRef]
- García-Sánchez, A.; Miranda-Díaz, A.G.; Cardona-Muñoz, E.G. The role of oxidative stress in physiopathology and pharmacological treatment with pro- and antioxidant properties in chronic diseases. Oxid. Med. Cell. Longev. 2020, 2020, 2082145. [Google Scholar] [CrossRef]
- Taherkhani, S.; Suzuki, K.; Castell, L. A short overview of changes in inflammatory cytokines and oxidative stress in response to physical activity and antioxidant supplementation. Antioxidants 2020, 9, 886. [Google Scholar] [CrossRef] [PubMed]
- Sharif, A.; Akhtar, N.; Khan, M.S.; Menaa, A.; Menaa, B.; Khan, B.A.; Menaa, F. Formulation and evaluation on human skin of a water-in-oil emulsion containing Muscat hamburg black grape seed extract. Int. J. Cosmet. Sci. 2015, 37, 253–258. [Google Scholar] [CrossRef]
- Le Lay, S.; Simard, G.; Martinez, M.C.; Andriantsitohaina, R. Oxidative stress and metabolic pathologies: From an adipocentric point of view. Oxid. Med. Cell. Longev. 2014, 2014, 908539. [Google Scholar] [CrossRef]
- Fernández-Iglesias, A.; Pajuelo, D.; Quesada, H.; Díaz, S.; Bladé, C.; Arola, L.; Salvadó, M.J.; Mulero, M. Grape seed proanthocyanidin extract improves the hepatic glutathione metabolism in obese Zucker rats. Mol. Nutr. Food. Res. 2014, 58, 727–737. [Google Scholar] [CrossRef] [PubMed]
- Santhakumar, A.B.; Bulmer, A.C.; Singh, I. A review of the mechanisms and effectiveness of dietary polyphenols in reducing oxidative stress and thrombotic risk. J. Hum. Nutr. Diet. 2014, 27, 1–21. [Google Scholar] [CrossRef] [PubMed]
- Sano, A.; Tokutake, S.; Seo, A. Proanthocyanidin-rich grape seed extract reduces leg swelling in healthy women during prolonged sitting. J. Sci. Food. Agric. 2013, 93, 457–462. [Google Scholar] [CrossRef] [PubMed]
- Saada, H.N.; Said, U.Z.; Meky, N.H.; Abd El Azime, A.S. Grape seed extract Vitis vinifera protects against radiation-induced oxidative damage and metabolic disorders in rats. Phytother. Res. 2009, 23, 434–438. [Google Scholar] [CrossRef]
- Valko, M.; Leibfritz, D.; Moncol, J.; Cronin, M.T.; Mazur, M.; Telser, J. Free radicals and antioxidants in normal physiological functions and human disease. Int. J. Biochem. Cell. Biol. 2007, 39, 44–84. [Google Scholar] [CrossRef]
- Friedenreich, C.M.; Pialoux, V.; Wang, Q.; Shaw, E.; Brenner, D.R.; Waltz, X.; Conroy, S.M.; Johnson, R.; Woolcott, C.G.; Poulin, M.J.; et al. Effects of exercise on markers of oxidative stress: An ancillary analysis of the Alberta physical activity and breast cancer prevention trial. BMJ. Open. Sport. Exerc. Med. 2016, 2, e000171. [Google Scholar] [CrossRef]
- Kurata, A.; Nishizawa, H.; Kihara, S.; Maeda, N.; Sonoda, M.; Okada, T.; Ohashi, K.; Hibuse, T.; Fujita, K.; Yasui, A.; et al. Blockade of Angiotensin II type-1 receptor reduces oxidative stress in adipose tissue and ameliorates adipocytokine dysregulation. Kidney Int. 2006, 70, 1717–1724. [Google Scholar] [CrossRef][Green Version]
- Candiracci, M.; Justo, M.L.; Castaño, A.; Rodriguez-Rodriguez, R.; Herrera, M.D. Rice bran enzymatic extract-supplemented diets modulate adipose tissue inflammation markers in Zucker rats. Nutrition 2014, 30, 466–472. [Google Scholar] [CrossRef] [PubMed]
- Alcalá, M.; Sánchez-Vera, I.; Sevillano, J.; Herrero, L.; Serra, D.; Ramos, M.P.; Viana, M. Vitamin E reduces adipose tissue fibrosis, inflammation, and oxidative stress and improves metabolic profile in obesity. Obesity 2015, 23, 1598–1606. [Google Scholar] [CrossRef]
- Preiser, J.C. Oxidative stress. JPEN. J. Parenter. Enteral. Nutr. 2012, 36, 147–154. [Google Scholar] [CrossRef]
- Vilchis-Landeros, M.M.; Matuz-Mares, D.; Vázquez-Meza, H. Regulation of metabolic processes by hydrogen peroxide generated by NADPH oxidases. Processes 2020, 8, 1424. [Google Scholar] [CrossRef]
- Sakurai, T.; Ogasawara, J.; Shirato, K.; Izawa, T.; Oh-Ishi, S.; Ishibashi, Y.; Radák, Z.; Ohno, H.; Kizaki, T. Exercise training attenuates the dysregulated expression of adipokines and oxidative stress in white adipose tissue. Oxid. Med. Cell. Longev. 2017, 2017, 9410954. [Google Scholar] [CrossRef]
- DeVallance, E.; Li, Y.; Jurczak, M.J.; Cifuentes-Pagano, E.; Pagano, P.J. The role of NADPH oxidases in the etiology of obesity and metabolic syndrome: Contribution of individual isoforms and cell biology. Antioxid. Redox. Signal. 2019, 31, 687–709. [Google Scholar] [CrossRef] [PubMed]
- Paredes, F.; Suster, I.; Martin, A.S. Poldip2 takes a central role in metabolic reprograming. Oncoscience 2018, 5, 130–131. [Google Scholar] [CrossRef] [PubMed]
- Tosetti, P.; Dunlap, K. Assays of RGS3 activation and modulation. Methods. Enzymol. 2004, 390, 99–119. [Google Scholar]
- Chazin, W.J. Relating form and function of EF-hand calcium binding proteins. Acc. Chem. Res. 2011, 44, 171–179. [Google Scholar] [CrossRef]
- Brownlee, M. The pathobiology of diabetic complications: A unifying mechanism. Diabetes 2005, 54, 1615–1625. [Google Scholar] [CrossRef]
- Matsuda, M.; Shimomura, I. Increased oxidative stress in obesity: Implications for metabolic syndrome, diabetes, hypertension, dyslipidemia, atherosclerosis, and cancer. Obes. Res. Clin. Pract. 2013, 7, e330–e341. [Google Scholar] [CrossRef]
- Dillard, C.J.; Litov, R.E.; Savin, W.M.; Dumelin, E.E.; Tappel, A.L. Effects of exercise, vitamin E, and ozone on pulmonary function and lipid peroxidation. J. Appl. Physiol. 1978, 45, 927–932. [Google Scholar] [CrossRef]
- Fisher-Wellman, K.; Bloomer, R.J. Acute exercise and oxidative stress: A 30 year history. Dyn. Med. 2009, 8, 1. [Google Scholar] [CrossRef]
- Halliwell, B.; Whiteman, M. Measuring reactive species and oxidative damage in vivo and in cell culture: How should you do it and what do the results mean? Br. J. Pharmacol. 2004, 142, 231–255. [Google Scholar] [CrossRef]
- Ohshima, H.; Sawa, T.; Akaike, T. 8-nitroguanine, a product of nitrative DNA damage caused by reactive nitrogen species: Formation, occurrence, and implications in inflammation and carcinogenesis. Antioxid. Redox. Signal. 2006, 8, 1033–1045. [Google Scholar] [CrossRef]
- Kilhovd, B.K.; Juutilainen, A.; Lehto, S.; Rönnemaa, T.; Torjesen, P.A.; Hanssen, K.F.; Laakso, M. Increased serum levels of advanced glycation endproducts predict total, cardiovascular and coronary mortality in women with type 2 diabetes: A population-based 18 year follow-up study. Diabetologia 2007, 50, 1409–1417. [Google Scholar] [CrossRef]
- Fang, Y.Z.; Yang, S.; Wu, G. Free radicals, antioxidants, and nutrition. Nutrition 2002, 18, 872–879. [Google Scholar] [CrossRef]
- Jones, D.P. Redox potential of GSH/GSSG couple: Assay and biological significance. Methods. Enzymol. 2002, 348, 93–112. [Google Scholar]
- Smas, C.M.; Sul, H.S. Control of adipocyte differentiation. Biochem. J. 1995, 309, 697–710. [Google Scholar] [CrossRef] [PubMed]
- Berry, R.; Church, C.D.; Gericke, M.T.; Jeffery, E.; Colman, L.; Rodeheffer, M.S. Imaging of adipose tissue. Methods Enzymol. 2014, 537, 47–73. [Google Scholar] [PubMed]
- Nakajima, I.; Yamaguchi, T.; Ozutsumi, K.; Aso, H. Adipose tissue extracellular matrix: Newly organized by adipocytes during differentiation. Differentiation 1998, 63, 193–200. [Google Scholar] [CrossRef] [PubMed]
- Luo, W.; Cao, J.; Li, J.; He, W. Adipose tissue-specific PPARgamma deficiency increases resistance to oxidative stress. Exp. Gerontol. 2008, 43, 154–163. [Google Scholar] [CrossRef]
- Sun, K.; Kusminski, C.M.; Scherer, P.E. Adipose tissue remodeling and obesity. J. Clin. Investig. 2011, 121, 2094–2101. [Google Scholar] [CrossRef]
- Trayhurn, P.; Beattie, J.H. Physiological role of adipose tissue: White adipose tissue as an endocrine and secretory organ. Proc. Nutr. Soc. 2001, 60, 329–339. [Google Scholar] [CrossRef]
- Choe, S.S.; Huh, J.Y.; Hwang, I.J.; Kim, J.I.; Kim, J.B. Adipose tissue remodeling: Its role in energy metabolism and metabolic disorders. Front. Endocrinol. 2016, 7, 30. [Google Scholar] [CrossRef]
- Siiteri, P.K. Adipose tissue as a source of hormones. Am. J. Clin. Nutr. 1987, 45, 277–282. [Google Scholar] [CrossRef]
- Kershaw, E.E.; Flier, J.S. Adipose tissue as an endocrine organ. J. Clin. Endocrinol. Metab. 2004, 89, 2548–2556. [Google Scholar] [CrossRef]
- Cook, K.S.; Min, H.Y.; Johnson, D.; Chaplinsky, R.J.; Flier, J.S.; Hunt, C.R.; Spiegelman, B.M. Adipsin: A circulating serine protease homolog secreted by adipose tissue and sciatic nerve. Science 1987, 237, 402–405. [Google Scholar] [CrossRef] [PubMed]
- Ryu, K.Y.; Jeon, E.J.; Leem, J.; Park, J.H.; Cho, H. Regulation of adipsin expression by endoplasmic reticulum stress in adipocytes. Biomolecules 2020, 10, 314. [Google Scholar] [CrossRef] [PubMed]
- Lo, J.C.; Ljubicic, S.; Leibiger, B.; Kern, M.; Leibiger, I.B.; Moede, T.; Kelly, M.E.; Bhowmick, D.C.; Murano, I.; Cohen, P.; et al. M. Adipsin is an adipokine that improves β cell function in diabetes. Cell 2014, 158, 41–53. [Google Scholar] [CrossRef] [PubMed]
- Liu, J.; DeYoung, S.M.; Zhang, M.; Zhang, M.; Cheng, A.; Saltiel, A.R. Changes in integrin expression during adipocyte differentiation. Cell. Metab. 2005, 2, 165–177. [Google Scholar] [CrossRef]
- Scheja, L.; Heeren, J. The endocrine function of adipose tissues in health and cardiometabolic disease. Nat. Rev. Endocrinol. 2019, 15, 507–524. [Google Scholar] [CrossRef] [PubMed]
- De Farias, J.M.; Bom, K.F.; Tromm, C.B.; Luciano, T.F.; Marques, S.O.; Tuon, T.; Silva, L.A.; Lira, F.S.; de Souza, C.T.; Pinho, R.A. Effect of physical training on the adipose tissue of diet-induced obesity mice: Interaction between reactive oxygen species and lipolysis. Horm. Metab. Res. 2013, 45, 190–196. [Google Scholar] [CrossRef][Green Version]
- Esteve Ràfols, M. Adipose tissue: Cell heterogeneity and functional diversity. Endocrinol. Nutr. 2014, 61, 100–112. [Google Scholar] [CrossRef] [PubMed]
- Lee, M.J.; Wu, Y.; Fried, S.K. Adipose tissue remodeling in pathophysiology of obesity. Curr. Opin. Clin. Nutr. Metab. Care 2010, 13, 371–376. [Google Scholar] [CrossRef] [PubMed]
- Zoico, E.; Rubele, S.; De Caro, A.; Nori, N.; Mazzali, G.; Fantin, F.; Rossi, A.; Zamboni, M. Brown and beige adipose tissue and aging. Front. Endocrinol. 2019, 10, 368. [Google Scholar] [CrossRef] [PubMed]
- Vegiopoulos, A.; Rohm, M.; Herzig, S. Adipose tissue: Between the extremes. EMBO J. 2017, 36, 1999–2017. [Google Scholar] [CrossRef]
- Berry, D.C.; Stenesen, D.; Zeve, D.; Graff, J.M. The developmental origins of adipose tissue. Development 2013, 140, 3939–3949. [Google Scholar] [CrossRef] [PubMed]
- Colaianni, G.; Colucci, S.; Grano, M. Anatomy and Physiology of Adipose Tissue; Springer: Cham, Switzerland, 2014. [Google Scholar] [CrossRef]
- Ro, S.H.; Nam, M.; Jang, I.; Park, H.W.; Park, H.; Semple, I.A.; Kim, M.; Kim, J.S.; Park, H.; Einat, P.; et al. Sestrin2 inhibits uncoupling protein 1 expression through suppressing reactive oxygen species. Proc. Natl. Acad. Sci. USA 2014, 111, 7849–7854. [Google Scholar] [CrossRef] [PubMed]
- Lee, J.H.; Budanov, A.V.; Talukdar, S.; Park, E.J.; Park, H.L.; Park, H.W.; Bandyopadhyay, G.; Li, N.; Aghajan, M.; Jang, I.; et al. Maintenance of metabolic homeostasis by Sestrin2 and Sestrin3. Cell. Metab. 2012, 16, 311–321. [Google Scholar] [CrossRef]
- Bae, S.H.; Sung, S.H.; Oh, S.Y.; Lim, J.M.; Lee, S.K.; Park, Y.N.; Lee, H.E.; Kang, D.; Rhee, S.G. Sestrins activate Nrf2 by promoting p62-dependent autophagic degradation of Keap1 and prevent oxidative liver damage. Cell. Metab. 2013, 17, 73–84. [Google Scholar] [CrossRef]
- Rothwell, N.J.; Stock, M.J. A role for brown adipose tissue in diet-induced thermogenesis. Obes. Res. 1997, 5, 650–656. [Google Scholar] [CrossRef] [PubMed]
- Turrens, J.F. Superoxide production by the mitochondrial respiratory chain. Biosci. Rep. 1997, 17, 3–8. [Google Scholar] [CrossRef]
- Krüger, K.; Mooren, F.C.; Eder, K.; Ringseis, R. Immune and inflammatory signaling pathways in exercise and obesity. Am. J. Lifestyle. Med. 2014, 10, 268–279. [Google Scholar] [CrossRef] [PubMed]
- Kaisanlahti, A.; Glumoff, T. Browning of white fat: Agents and implications for beige adipose tissue to type 2 diabetes. J. Physiol. Biochem. 2019, 75, 1–10. [Google Scholar] [CrossRef] [PubMed]
- Fain, J.N. Release of interleukins and other inflammatory cytokines by human adipose tissue is enhanced in obesity and primarily due to the nonfat cells. Vitam. Horm. 2006, 74, 443–477. [Google Scholar] [PubMed]
- Skurk, T.; Alberti-Huber, C.; Herder, C.; Hauner, H. Relationship between adipocyte size and adipokine expression and secretion. J. Clin. Endocrinol. Metab. 2007, 92, 1023–1033. [Google Scholar] [CrossRef] [PubMed]
- Zhao, J.; Zhai, L.; Liu, Z.; Wu, S.; Xu, L. Leptin level and oxidative stress contribute to obesity-induced low testosterone in murine testicular tissue. Oxid. Med. Cell. Longev. 2014, 2014, 190945. [Google Scholar] [CrossRef]
- Alexandre, E.C.; Calmasini, F.B.; Sponton, A.C.D.S.; de Oliveira, M.G.; André, D.M.; Silva, F.H.; Delbin, M.A.; Mónica, F.Z.; Antunes, E. Influence of the periprostatic adipose tissue in obesity-associated mouse urethral dysfunction and oxidative stress: Effect of resveratrol treatment. Eur. J. Pharmacol. 2018, 836, 25–33. [Google Scholar] [CrossRef] [PubMed]
- Ruhee, R.T.; Suzuki, K. Dietary fiber and its effect on obesity: A review article. Adv. Med. Res. 2018, 1, 1–13. [Google Scholar] [CrossRef]
- Bailey-Downs, L.C.; Tucsek, Z.; Toth, P.; Sosnowska, D.; Gautam, T.; Sonntag, W.E.; Csiszar, A.; Ungvari, Z. Aging exacerbates obesity-induced oxidative stress and inflammation in perivascular adipose tissue in mice: A paracrine mechanism contributing to vascular redox dysregulation and inflammation. J. Gerontol. A. Biol. Sci. Med. Sci. 2013, 68, 780–792. [Google Scholar] [CrossRef]
- Abdali, D.; Samson, S.E.; Grover, A.K. How effective are antioxidant supplements in obesity and diabetes? Med. Princ. Pract. 2015, 24, 201–215. [Google Scholar] [CrossRef]
- Bjørklund, G.; Chirumbolo, S. Role of oxidative stress and antioxidants in daily nutrition and human health. Nutrition 2017, 33, 311–321. [Google Scholar] [CrossRef]
- Ruiz-Ojeda, F.J.; Méndez-Gutiérrez, A.; Aguilera, C.M.; Plaza-Díaz, J. Extracellular matrix remodeling of adipose tissue in obesity and metabolic diseases. Int. J. Mol. Sci. 2019, 20, 4888. [Google Scholar] [CrossRef]
- Timmers, S.; de Vogel-van den Bosch, J.; Towler, M.C.; Schaart, G.; Moonen-Kornips, E.; Mensink, R.P.; Hesselink, M.K.; Hardie, D.G.; Schrauwen, P. Prevention of high-fat diet-induced muscular lipid accumulation in rats by alpha lipoic acid is not mediated by AMPK activation. J. Lipid. Res. 2010, 51, 352–359. [Google Scholar] [CrossRef]
- Martínez-Fernández, L.; Fernández-Galilea, M.; Felix-Soriano, E.; Escoté, X.; González-Muniesa, P.; Moreno-Aliaga, M.J. Inflammation and oxidative stress in adipose tissue: Nutritional regulation. In Obesity: Oxidative Stress and Dietary Antioxidants; del Moral, A.M., Garcia, C.M.A., Eds.; Elsevier: Amsterdam, The Netherlands, 2018; Chapter 4; pp. 63–92. [Google Scholar]
- Aldini, G.; Yeum, K.J.; Niki, E.; Russell, R.M. Biomarkers for Antioxidant Defense and Oxidative Damage: Principles and Practical Applications; Blackwell: Oxford, UK, 2010. [Google Scholar] [CrossRef]
- Brooker, S.; Martin, S.; Pearson, A.; Bagchi, D.; Earl, J.; Gothard, L.; Hall, E.; Porter, L.; Yarnold, J. Double-blind, placebo-controlled, randomised phase II trial of IH636 grape seed proanthocyanidin extract (GSPE) in patients with radiation-induced breast induration. Radiother. Oncol. 2006, 79, 45–51. [Google Scholar] [CrossRef] [PubMed]
- Han, Y.; Song, S.; Wu, H.; Zhang, J.; Ma, E. Antioxidant enzymes and their role in phoxim and carbaryl stress in Caenorhabditis elegans. Pestic. Biochem. Physiol. 2017, 138, 43–50. [Google Scholar] [CrossRef] [PubMed]
- Keong, C.C.; Singh, H.J.; Singh, R. Effects of palm vitamin e supplementation on exercise-induced oxidative stress and endurance performance in the heat. J. Sports Sci. Med. 2006, 5, 629–639. [Google Scholar]
- Leong, P.K.; Chen, J.; Chan, W.M.; Leung, H.Y.; Chan, L.; Ko, K.M. Acute pre-/post-treatment with 8th day SOD-like supreme (a free radical scavenging health product) protects against oxidant-induced injury in cultured cardiomyocytes and hepatocytes in vitro as well as in mouse myocardium and liver in vivo. Antioxidants 2017, 6, 28. [Google Scholar] [CrossRef] [PubMed]
- Li, Z.; Han, X.; Song, X.; Zhang, Y.; Jiang, J.; Han, Q.; Liu, M.; Qiao, G.; Zhuo, R. Overexpressing the sedum alfredii Cu/Zn superoxide dismutase increased resistance to oxidative stress in transgenic arabidopsis. Front. Plant. Sci. 2017, 8, 1010. [Google Scholar] [CrossRef]
- Staerck, C.; Vandeputte, P.; Gastebois, A.; Calenda, A.; Giraud, S.; Papon, N.; Bouchara, J.P.; Fleury, M.J.J. Enzymatic mechanisms involved in evasion of fungi to the oxidative stress: Focus on scedosporium apiospermum. Mycopathologia 2018, 183, 227–239. [Google Scholar] [CrossRef]
- Berwal, M.K.; Padmanabhan, S.; Vittal, N.; Hebbar, K.B. Variability in superoxide dismutase isoforms in tall and dwarf cultivars of coconut (Cocos nucifera L.) Leaves. Ind. J. Agric. Biochem. 2016, 29, 184. [Google Scholar] [CrossRef]
- Morrow, J.D.; Roberts, L.J. Mass spectrometric quantification of F2-isoprostanes in biological fluids and tissues as measure of oxidant stress. Methods. Enzymol. 1999, 300, 3–12. [Google Scholar]
- Kang, S.W. Superoxide dismutase 2 gene and cancer risk: Evidence from an updated meta-analysis. Int. J. Clin. Exp. Med. 2015, 8, 14647–14655. [Google Scholar]
- Halliwell, B.; Gutteridge, J.M.C. Free Radicals in Biology and Medicine; Oxford Scholarship Online: Oxford, UK, 2015. [Google Scholar] [CrossRef]
- Margis, R.; Dunand, C.; Teixeira, F.K.; Margis-Pinheiro, M. Glutathione peroxidase family—An evolutionary overview. FEBS J. 2008, 275, 3959–3970. [Google Scholar] [CrossRef] [PubMed]
- Chance, B.; Greenstein, D.S.; Roughton, F.J.W. The mechanism of catalase action. I. Steady-state analysis. Arch. Biochem. Biophys. 1952, 37, 301–321. [Google Scholar] [CrossRef]
- Kahn, S.E.; Hull, R.L.; Utzschneider, K.M. Mechanisms linking obesity to insulin resistance and type 2 diabetes. Nature 2006, 444, 840–846. [Google Scholar] [CrossRef] [PubMed]
- Suzuki, K. Chronic inflammation as an immunological abnormality and effectiveness of exercise. Biomolecules 2019, 9, 223. [Google Scholar] [CrossRef] [PubMed]
- Masschelin, P.M.; Cox, A.R.; Chernis, N.; Hartig, S.M. The impact of oxidative stress on adipose tissue energy balance. Front. Physiol. 2020, 10, 1638. [Google Scholar] [CrossRef] [PubMed]
- Simán, C.M.; Eriksson, U.J. Effect of butylated hydroxytoluene on alpha-tocopherol content in liver and adipose tissue of rats. Toxicol. Lett. 1996, 87, 103–138. [Google Scholar] [CrossRef]
- Rodrigues, P.; Ferreira, T.; Nascimento-Gonçalves, E.; Seixas, F.; Gil da Costa, R.M.; Martins, T.; Neuparth, M.J.; Pires, M.J.; Lanzarin, G.; Félix, L.; et al. Dietary supplementation with chestnut (Castanea sativa) reduces abdominal adiposity in FVB/n mice: A preliminary study. Biomedicines 2020, 8, 75. [Google Scholar] [CrossRef]
- Valls, V.; Goicoechea, M.; Muñiz, P.; Cabo, S.J.R. Effect of corn oil and vitamin E on the oxidative status of adipose tissues and liver in rat. Food. Chem. 2003, 81, 281–286. [Google Scholar] [CrossRef]
- Arias, N.; Macarulla, M.T.; Aguirre, L.; Martínez-Castaño, M.G.; Portillo, M.P. Quercetin can reduce insulin resistance without decreasing adipose tissue and skeletal muscle fat accumulation. Genes. Nutr. 2014, 9, 361. [Google Scholar] [CrossRef]
- Chen, J.W.; Lin, Y.L.; Chou, C.H.; Wu, Y.H.S.; Wang, S.Y.; Chen, Y.C. Antiobesity and hypolipidemic effects of protease A-digested crude-chalaza hydrolysates in a high-fat diet. J. Funct. Foods. 2020, 66, 103788. [Google Scholar] [CrossRef]
- Djurasevic, S.F.; Cvijic, G.; Djordjevic, J.; Davidovic, V. The influence of vitamin C supplementation on the oxidative status of rat interscapular brown adipose tissue. J. Therm. Biol. 2008, 33, 238–243. [Google Scholar] [CrossRef]
- Sung, Y.Y.; Yoon, T.; Yang, W.K.; Kim, S.J.; Kim, D.S.; Kim, H.K. The antiobesity effect of polygonum aviculare L. ethanol extract in high-fat diet-induced obese mice. Evid. Based. Complement. Altern. Med. 2013, 2013, 626397. [Google Scholar] [CrossRef]
- Maurya, R.; Bhattacharya, P.; Dey, R.; Nakhasi, H.L. Leptin functions in infectious diseases. Front. Immunol. 2018, 9, 2741. [Google Scholar] [CrossRef] [PubMed]
- Soeroto, A.Y.; Soetedjo, N.N.; Purwiga, A.; Santoso, P.; Kulsum, I.D.; Suryadinata, H.; Ferdian, F. Effect of increased BMI and obesity on the outcome of COVID-19 adult patients: A systematic review and meta-analysis. Diabetes. Metab. Syndr. 2020, 14, 1897–1904. [Google Scholar] [CrossRef]
- O’Shea, D.; Hogan, A.E. Dysregulation of natural killer cells in obesity. Cancers 2019, 11, 573. [Google Scholar] [CrossRef] [PubMed]
- Hinchliffe, N.; Bullen, V.; Haslam, D.; Feenie, J. COVID-19 and obesity. Pract. Diabetes 2020, 37, 149–151. [Google Scholar] [CrossRef]
- Khoramipour, K.; Basereh, A.; Hekmatikar, A.A.; Castell, L.; Ruhee, R.T.; Suzuki, K. Physical activity and nutrition guidelines to help with the fight against COVID-19. J. Sports Sci. 2021, 39, 101–107. [Google Scholar] [CrossRef]
- Fernández-Quintela, A.; Milton-Laskibar, I.; Trepiana, J.; Gómez-Zorita, S.; Kajarabille, N.; Léniz, A.; González, M.; Portillo, M.P. Key aspects in nutritional management of COVID-19 patients. J. Clin. Med. 2020, 9, 2589. [Google Scholar] [CrossRef]
- Mohammad, S.; Aziz, R.; Al Mahri, S.; Malik, S.S.; Haji, E.; Khan, A.H.; Khatlani, T.S.; Bouchama, A. Obesity and COVID-19: What makes obese host so vulnerable? Immun. Ageing 2021, 18, 1. [Google Scholar] [CrossRef]
- Fakhouri, E.W.; Peterson, S.J.; Kothari, J.; Alex, R.; Shapiro, J.I.; Abraham, N.G. Genetic polymorphisms complicate COVID-19 therapy: Pivotal role of HO-1 in cytokine storm. Antioxidants 2020, 9, 636. [Google Scholar] [CrossRef] [PubMed]
- Favre, G.; Legueult, K.; Pradier, C.; Raffaelli, C.; Ichai, C.; Iannelli, A.; Redheuil, A.; Lucidarme, O.; Esnault, V. Visceral fat is associated to the severity of COVID-19. Metabolism 2021, 115, 154440. [Google Scholar] [CrossRef] [PubMed]
- Gheblawi, M.; Wang, K.; Viveiros, A.; Nguyen, Q.; Zhong, J.C.; Turner, A.J.; Raizada, M.K.; Grant, M.B.; Oudit, G.Y. Angiotensin-Converting enzyme 2: SARS-CoV-2 receptor and regulator of the Renin-Angiotensin System: Celebrating the 20th anniversary of the discovery of ACE2. Circ. Res. 2020, 126, 1456–1474. [Google Scholar] [CrossRef]
- Xie, Z.J.; Novograd, J.; Itzkowitz, Y.; Sher, A.; Buchen, Y.D.; Sodhi, K.; Abraham, N.G.; Shapiro, J.I. The pivotal role of adipocyte-Na K peptide in reversing systemic inflammation in obesity and COVID-19 in the development of heart failure. Antioxidants 2020, 9, 1129. [Google Scholar] [CrossRef]
- Li, S.; Cao, Z.; Yang, H.; Zhang, Y.; Xu, F.; Wang, Y. Metabolic healthy obesity, vitamin D status, and risk of COVID-19. Aging. Dis. 2021, 12, 61–71. [Google Scholar] [CrossRef] [PubMed]
- Mohseni, H.; Amini, S.; Abiri, B.; Kalantar, M. Do body mass index (BMI) and history of nutritional supplementation play a role in the severity of COVID-19? A retrospective study. Nutr. Food. Sci. 2021. preprint. [Google Scholar] [CrossRef]
- Maggini, S.; Pierre, A.; Calder, P.C. Immune function and micronutrient requirements change over the life course. Nutrients 2018, 10, 1531. [Google Scholar] [CrossRef]
- Calder, P.C.; Carr, A.C.; Gombart, A.F.; Eggersdorfer, M. Optimal nutritional status for a well-functioning immune system is an important factor to protect against viral infections. Nutrients 2020, 12, 1181. [Google Scholar] [CrossRef]
- Grant, W.B.; Lahore, H.; McDonnell, S.L.; Baggerly, C.A.; French, C.B.; Aliano, J.L.; Bhattoa, H.P. Evidence that vitamin D supplementation could reduce risk of influenza and COVID-19 infections and deaths. Nutrients 2020, 12, 988. [Google Scholar] [CrossRef] [PubMed]
- Zhang, J.; Taylor, E.W.; Bennett, K.; Saad, R.; Rayman, M.P. Association between regional selenium status and reported outcome of COVID-19 cases in China. Am. J. Clin. Nutr. 2020, 111, 1297–1299. [Google Scholar] [CrossRef] [PubMed]
- Dornas, W.; Schuppan, D. Mitochondrial oxidative injury: A key player in nonalcoholic fatty liver disease. Am. J. Physiol. Gastrointest. Liver Physiol. 2020, 319, G400–G411. [Google Scholar] [CrossRef]
- Zhang, C.; Wang, P.; Li, Y.; Huang, C.; Ni, W.; Chen, Y.; Shi, J.; Chen, G.; Hu, X.; Ye, M.; et al. Role of MicroRNAs in the development of hepatocellular carcinoma in nonalcoholic fatty liver disease. Anat. Rec. 2019, 302, 193–200. [Google Scholar] [CrossRef] [PubMed]
- Rolo, A.P.; Teodoro, J.S.; Palmeira, C.M. Role of oxidative stress in the pathogenesis of nonalcoholic steatohepatitis. Free. Radic. Biol. Med. 2012, 52, 59–69. [Google Scholar] [CrossRef] [PubMed]
- He, Y.; Jiang, J.; He, B.; Shi, Z. Chemical activators of the Nrf2 signaling pathway in nonalcoholic fatty liver disease. Nat. Prod. Commun. 2021, 16, 1–9. [Google Scholar]
- Wang, J.; He, W.; Tsai, P.J.; Chen, P.H.; Ye, M.; Guo, J.; Su, Z.H. Mutual interaction between endoplasmic reticulum and mitochondria in nonalcoholic fatty liver disease. Lipids. Health Dis. 2020, 19, 72. [Google Scholar] [CrossRef] [PubMed]
- Kawanishi, N.; Mizokami, T.; Yada, K.; Suzuki, K. Exercise training suppresses scavenger receptor CD36 expression in kupffer cells of nonalcoholic steatohepatitis model mice. Physiol. Rep. 2018, 6, e13902. [Google Scholar] [CrossRef]
- Kawanishi, N.; Yano, H.; Mizokami, T.; Takahashi, M.; Oyanagi, E.; Suzuki, K. Exercise training attenuates hepatic inflammation, fibrosis and macrophage infiltration during diet induced-obesity in mice. Brain. Behav. Immun. 2012, 26, 931–941. [Google Scholar] [CrossRef]
- Simoes, I.C.M.; Janikiewicz, J.; Bauer, J.; Karkucinska-Wieckowska, A.; Kalinowski, P.; Dobrzyń, A.; Wolski, A.; Pronicki, M.; Zieniewicz, K.; Dobrzyń, P.; et al. Fat and sugar-A dangerous duet. A comparative review on metabolic remodeling in rodent models of nonalcoholic fatty liver disease. Nutrients 2019, 11, 2871. [Google Scholar] [CrossRef]
- Videla, L.A.; Rodrigo, R.; Orellana, M.; Fernandez, V.; Tapia, G.; Quiñones, L.; Varela, N.; Contreras, J.; Lazarte, R.; Csendes, A.; et al. Oxidative stress-related parameters in the liver of non-alcoholic fatty liver disease patients. Clin. Sci. 2004, 106, 261–268. [Google Scholar] [CrossRef] [PubMed]
- Yki-Järvinen, H. Non-alcoholic fatty liver disease as a cause and a consequence of metabolic syndrome. Lancet Diabetes Endocrinol. 2014, 2, 901–910. [Google Scholar] [CrossRef]
- Buzzetti, E.; Pinzani, M.; Tsochatzis, E.A. The multiple-hit pathogenesis of non-alcoholic fatty liver disease (NAFLD). Metabolism 2016, 65, 1038–1048. [Google Scholar] [CrossRef] [PubMed]
- Tarantino, G.; Citro, V.; Capone, D. Nonalcoholic fatty liver disease: A challenge from mechanisms to therapy. J. Clin. Med. 2019, 9, 15. [Google Scholar] [CrossRef] [PubMed]
- Chiu, S.; Mulligan, K.; Schwarz, J.M. Dietary carbohydrates and fatty liver disease: De novo lipogenesis. Curr. Opin. Clin. Nutr. Metab. Care. 2018, 21, 277–282. [Google Scholar] [CrossRef]
- Drygalski, K.; Berk, K.; Charytoniuk, T.; Iłowska, N.; Łukaszuk, B.; Chabowski, A.; Konstantynowicz-Nowicka, K. Does the enterolactone (ENL) affect fatty acid transporters and lipid metabolism in liver? Nutr. Metab 2020, 14, 69. [Google Scholar] [CrossRef]
- Pagano, C.; Soardo, G.; Esposito, W.; Fallo, F.; Basan, L.; Donnini, D.; Federspil, G.; Sechi, L.A.; Vettor, R. Plasma adiponectin is decreased in nonalcoholic fatty liver disease. Eur. J. Endocrinol. 2005, 152, 113–118. [Google Scholar] [CrossRef]
- Khoramipour, K.; Chamari, K.; Hekmatikar, A.A.; Ziyaiyan, A.; Taherkhani, S.; Elguindy, N.M.; Bragazzi, N.L. Adiponectin: Structure, physiological functions, role in diseases, and effects of nutrition. Nutrients 2021, 13, 1180. [Google Scholar] [CrossRef]
- Liu, W.; Baker, S.S.; Baker, R.D.; Zhu, L. Antioxidant mechanisms in nonalcoholic fatty liver disease. Curr. Drug Targets 2015, 16, 1301–1314. [Google Scholar] [CrossRef]
- Vanni, E.; Bugianesi, E.; Kotronen, A.; De Minicis, S.; Yki-Järvinen, H.; Svegliati-Baroni, G. From the metabolic syndrome to NAFLD or vice versa? Dig. Liver Dis. 2010, 42, 320–330. [Google Scholar] [CrossRef] [PubMed]
- Younossi, Z.M.; Loomba, R.; Rinella, M.E.; Bugianesi, E.; Marchesini, G.; Neuschwander-Tetri, B.A.; Serfaty, L.; Negro, F.; Caldwell, S.H.; Ratziu, V.; et al. Current and future therapeutic regimens for nonalcoholic fatty liver disease and nonalcoholic steatohepatitis. Hepatology 2018, 68, 361–371. [Google Scholar] [CrossRef] [PubMed]
- Brunt, E.M.; Wong, V.W.; Nobili, V.; Day, C.P.; Sookoian, S.; Maher, J.J.; Bugianesi, E.; Sirlin, C.B.; Neuschwander-Tetri, B.A.; Rinella, M.E. Nonalcoholic fatty liver disease. Nat. Rev. Dis. Primers 2015, 1, 15080. [Google Scholar] [CrossRef]
- Alisi, A.; Carpino, G.; Oliveira, F.L.; Panera, N.; Nobili, V.; Gaudio, E. The role of tissue macrophage-mediated inflammation on NAFLD pathogenesis and its clinical implications. Mediators. Inflamm. 2017, 2017, 8162421. [Google Scholar] [CrossRef] [PubMed]
- Maher, J.; Yamamoto, M. The rise of antioxidant signaling—The evolution and hormetic actions of Nrf2. Toxicol. Appl. Pharmacol. 2010, 244, 4–15. [Google Scholar] [CrossRef] [PubMed]
- Kobayashi, M.; Yamamoto, M. Molecular mechanisms activating the Nrf2-Keap1 pathway of antioxidant gene regulation. Antioxid. Redox. Signal. 2005, 7, 385–394. [Google Scholar] [CrossRef]
- Michalopoulos, G.K. Liver regeneration. J. Cell. Physiol. 2007, 213, 286–300. [Google Scholar] [CrossRef]
- McMahon, M.; Thomas, N.; Itoh, K.; Yamamoto, M.; Hayes, J.D. Redox-regulated turnover of Nrf2 is determined by at least two separate protein domains, the redox-sensitive Neh2 degron and the redox-insensitive Neh6 degron. J. Biol. Chem. 2004, 279, 31556–31567. [Google Scholar] [CrossRef]
- Yamamoto, T.; Suzuki, T.; Kobayashi, A.; Wakabayashi, J.; Maher, J.; Motohashi, H.; Yamamoto, M. Physiological significance of reactive cysteine residues of Keap1 in determining Nrf2 activity. Mol. Cell. Biol. 2008, 28, 2758–2770. [Google Scholar] [CrossRef] [PubMed]
- Sharma, R.S.; Harrison, D.J.; Kisielewski, D.; Cassidy, D.M.; McNeilly, A.D.; Gallagher, J.R.; Walsh, S.V.; Honda, T.; McCrimmon, R.J.; Dinkova-Kostova, A.T.; et al. Experimental nonalcoholic steatohepatitis and liver fibrosis are ameliorated by pharmacologic activation of Nrf2 (NF-E2 p45-related factor 2). Cell. Mol. Gastroenterol. Hepatol. 2017, 5, 367–398. [Google Scholar] [CrossRef]
- Surh, Y.J. Cancer chemoprevention with dietary phytochemicals. Nat. Rev. Cancer 2003, 3, 768–780. [Google Scholar] [CrossRef] [PubMed]
- Agrawal, D.K.; Mishra, P.K. Curcumin and its analogues: Potential anticancer agents. Med. Res. Rev. 2010, 30, 818–860. [Google Scholar] [CrossRef]
- Ma, S.; Yada, K.; Lee, H.; Fukuda, Y.; Iida, A.; Suzuki, K. Taheebo polyphenols attenuate free fatty acid-induced inflammation in murine and human macrophage cell lines as inhibitor of cyclooxygenase-2. Front. Nutr. 2017, 4, 63. [Google Scholar] [CrossRef]
- Yada, K.; Suzuki, K.; Oginome, N.; Ma, S.; Fukuda, Y.; Iida, A.; Radak, Z. Single dose administration of taheebo polyphenol enhances endurance capacity in mice. Sci. Rep. 2018, 8, 1–12. [Google Scholar] [CrossRef] [PubMed]
- Ruhee, R.T.; Ma, S.; Suzuki, K. Sulforaphane protects cells against lipopolysaccharide-stimulated inflammation in murine macrophages. Antioxidants 2019, 8, 577. [Google Scholar] [CrossRef] [PubMed]
- Ruhee, R.T.; Ma, S.; Suzuki, K. Protective effects of sulforaphane on exercise-induced organ damage via inducing antioxidant defense responses. Antioxidants 2020, 9, 136. [Google Scholar] [CrossRef] [PubMed]
- Espín, J.C.; García-Conesa, M.T.; Tomás-Barberán, F.A. Nutraceuticals: Facts and fiction. Phytochemistry 2007, 68, 2986–3008. [Google Scholar] [CrossRef] [PubMed]
- Tramontin, N.d.S.; Luciano, T.F.; Marques, S.d.O.; de Souza, C.T.; Muller, A.P. Ginger and avocado as nutraceuticals for obesity and its comorbidities. Phytother. Res. 2020, 34, 1282–1290. [Google Scholar] [CrossRef]
- Kota, S.; Jammula, S.; Kota, S.; Satya Krishna, S.; Meher, L.; Rao, E.; Modi, K. Nutraceuticals in pathogenic obesity; striking the right balance between energy imbalance and inflammation. J. Med. Nutr. Nutraceuticals 2012, 1, 63–76. [Google Scholar] [CrossRef]
- Suzuki, K.; Tominaga, T.; Ruhee, R.T.; Ma, S. Characterization and Modulation of Systemic Inflammatory Response to Exhaustive Exercise in Relation to Oxidative Stress. Antioxidants 2020, 9, 401. [Google Scholar] [CrossRef]
- Suzuki, K.; Hayashida, H. Effect of exercise intensity on cell-mediated immunity. Sports 2021, 9, 8. [Google Scholar] [CrossRef]




{kind=link}
{kind=link}
{kind=link}
| Reference | Subjects | Antioxidant Supplementation | Results |
|---|---|---|---|
| Simán et al. [102] | Sprague Dawley rats | BHT (0.5% and 1%) with or without vitamin E acetate (4%) for four weeks. | No change in the alpha-tocopherol concentration of abdominal AT with BHT supplementation. |
| Rodrigues et al. [103] | FVB/n male 7-month-old mice | Chestnut at a dose of 1.1%. | The reduction of serum cholesterol and AT deposition. |
| Candiracci et al. [29] | Obese and lean Zucker rats | Rice bran enzymatic extract (RBEE) for 20 weeks. | The reduction of overproduction of IL-6, TNF-α, IL-1β, and NOS in abdominal and epidermal visceral AT. Reducing the adipocyte size of abdominal and epidural visceral AT. |
| Valls et al. [104] | Male Wistar rats | Diet rich in corn oil with or without antioxidant supplementation of vitamin E (30 mg per day). | The reduction of activity of the antioxidant enzymes CAT and SOD. |
| Arias et al. [105] | Male Wistar rats | Quercetin (30 mg/kg body weight). | No impact on reducing AT size and body weight. No change in the activity of lipoprotein lipase and lipogenic enzymes. |
| Chen et al. [106] | Syrian male Golden Hamsters | Protease A-digested crude-chalaza hydrolysates (CCH-As). | The reduction adipose-perinatal/hepatic tissue size. The increase of lipolysis (unpaired carnitine palmitoyltransferase 1, hormone-sensitive lipase, and protein 2). |
| Djurasevic et al. [107] | Male Wistar rats | Vitamin C supplementation (low and high doses of 0.75 and 25 mg of ascorbic acid per kg of body weight, respectively) for 4 weeks. | Excessive consumption of this antioxidant supplement was able to strengthen antioxidant defenses (MnSOD, CuZnSOD, and CAT in AT |
| Sung et al. [108] | Male C57BL/6J mice | High-fat diet or a high-fat diet with PAE at a dose of 400 mg/kg body weight per day. | The reduction of AT weight, serum TG concentration, body weight, MDA and leptin concentrations, and fat cell area. |
| Alcalá et al. [30] | C57BL/6J mice | Vitamin E supplementation (150 mg twice daily). | The reduction in collagen deposition and OS in rat visceral AT. The increase of storage capacity and fat cells’ proliferation. |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Taherkhani, S.; Suzuki, K.; Ruhee, R.T. A Brief Overview of Oxidative Stress in Adipose Tissue with a Therapeutic Approach to Taking Antioxidant Supplements. Antioxidants 2021, 10, 594. https://doi.org/10.3390/antiox10040594
Taherkhani S, Suzuki K, Ruhee RT. A Brief Overview of Oxidative Stress in Adipose Tissue with a Therapeutic Approach to Taking Antioxidant Supplements. Antioxidants. 2021; 10(4):594. https://doi.org/10.3390/antiox10040594
Chicago/Turabian StyleTaherkhani, Shima, Katsuhiko Suzuki, and Ruheea Taskin Ruhee. 2021. "A Brief Overview of Oxidative Stress in Adipose Tissue with a Therapeutic Approach to Taking Antioxidant Supplements" Antioxidants 10, no. 4: 594. https://doi.org/10.3390/antiox10040594
APA StyleTaherkhani, S., Suzuki, K., & Ruhee, R. T. (2021). A Brief Overview of Oxidative Stress in Adipose Tissue with a Therapeutic Approach to Taking Antioxidant Supplements. Antioxidants, 10(4), 594. https://doi.org/10.3390/antiox10040594