
Exploring Molecular Insights of Cereal Peptidic Antioxidants in Metabolic Syndrome Prevention

 ,
, 
 and
and 
Abstract
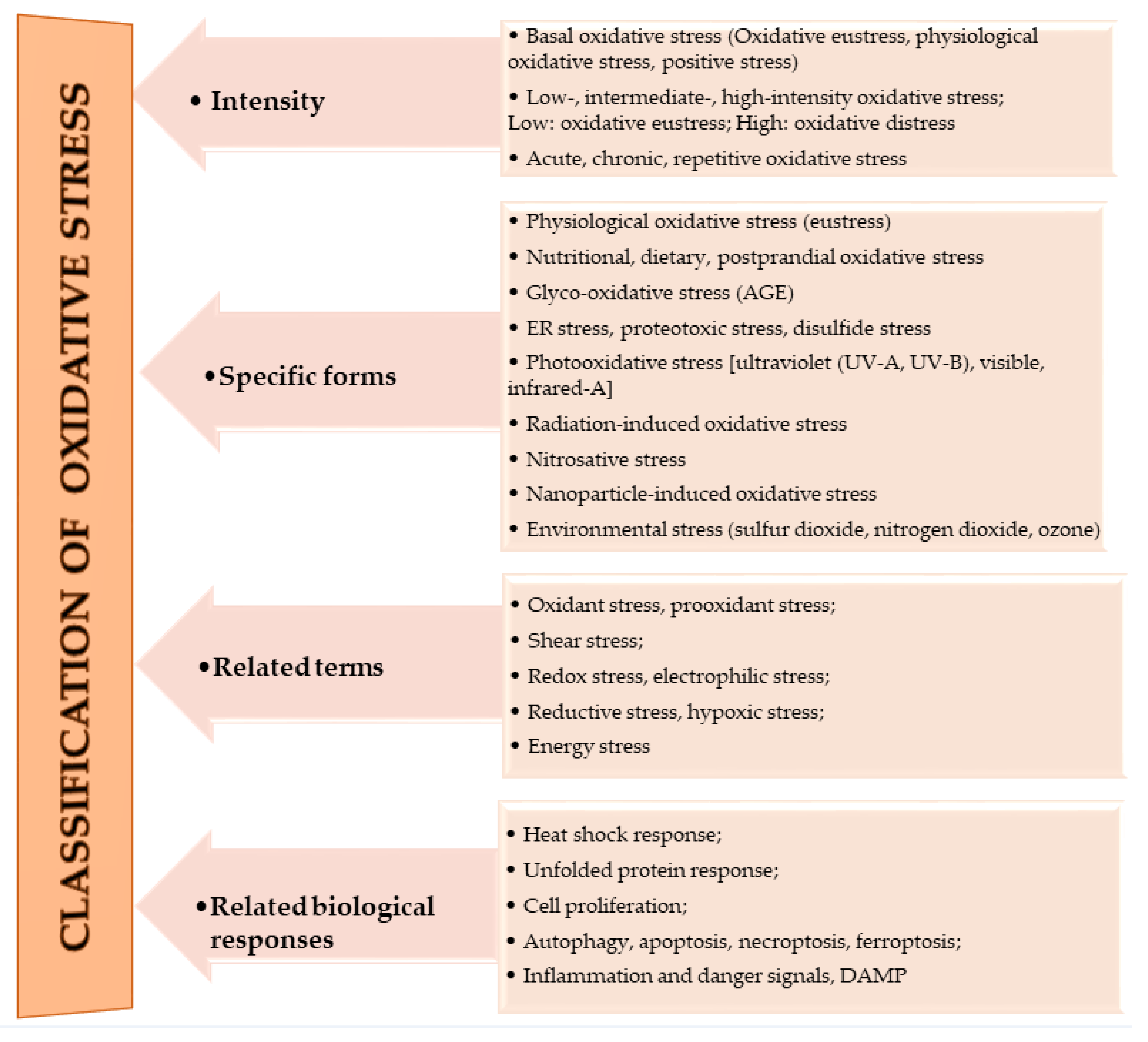
1. Oxidative Stress
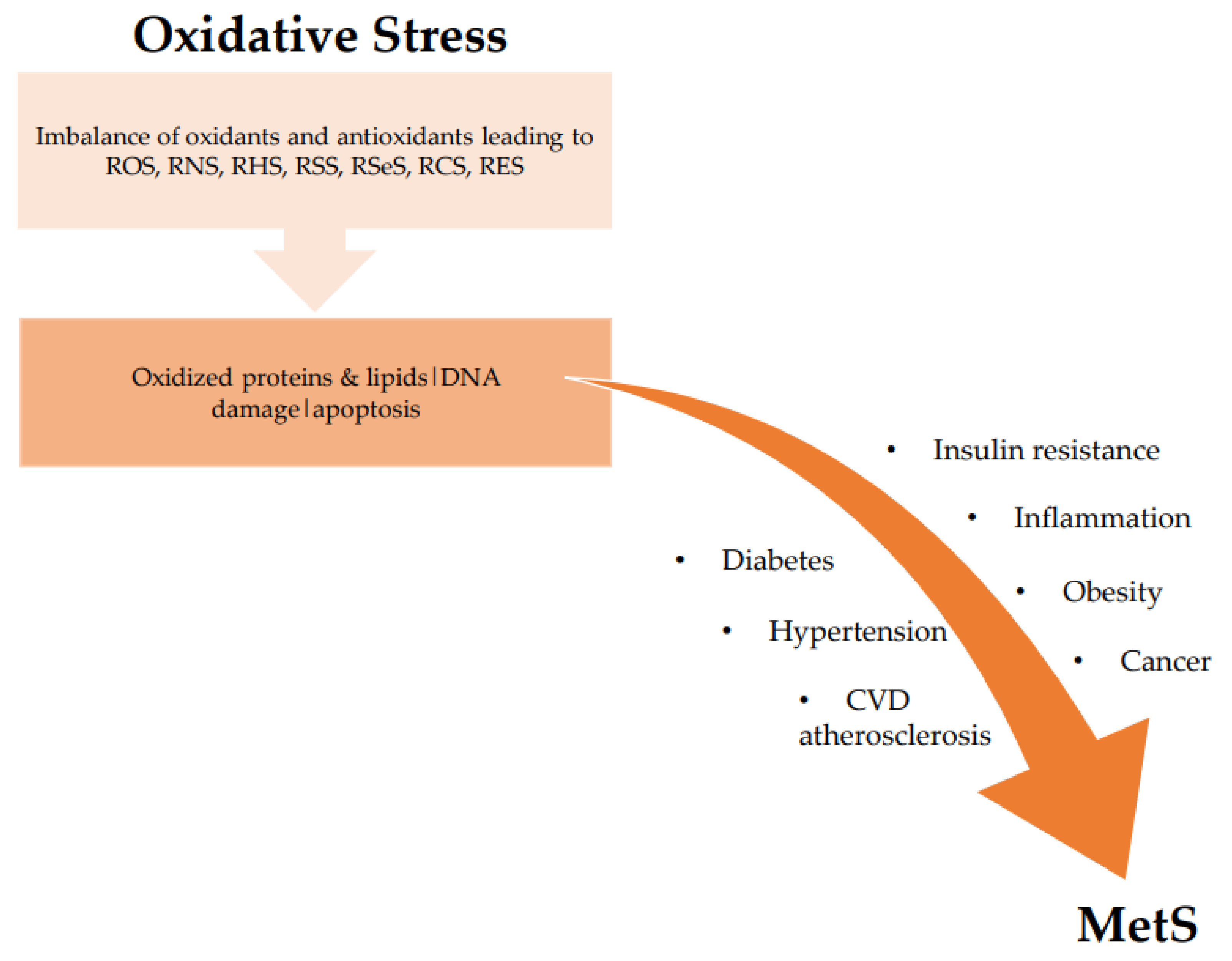
2. Metabolic Syndrome (MetS)
The Link: Oxidative Stress and Metabolic Syndrome
3. Antioxidative Peptides
4. Cereals Historical Significance and Assortments
4.1. Cereals as Sources of Antioxidant Peptides
4.1.1. Oats
4.1.2. Wheat
4.1.3. Rice
4.1.4. Barley
4.1.5. Rye
4.1.6. Maize
4.1.7. Millet
4.1.8. Sorghum
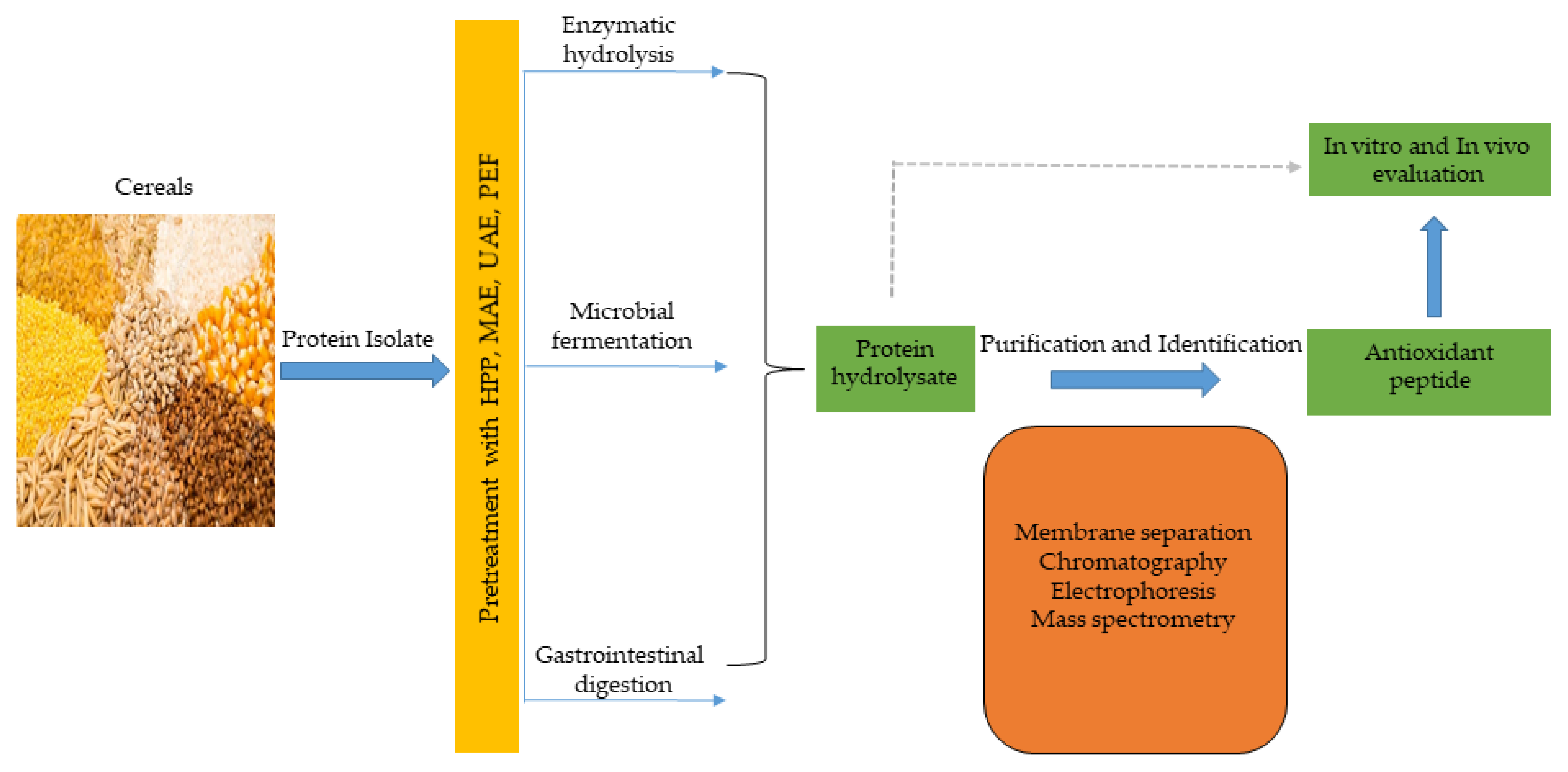
5. Production Strategies for Antioxidant Peptides
5.1. Conventional Approach
5.1.1. Enzymatic Hydrolysis
5.1.2. Microbial Fermentation
5.2. Bioinformatics Approach
5.3. Emerging Food Processing Technologies
6. Potential Mechanisms of Cereal Peptidic Antioxidants in MetS Prevention
7. Conclusions
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Breitenbach, M.; Eckl, P. Introduction to Oxidative Stress in Biomedical and Biological Research. Biomolecules 2015, 5, 1169–1170. [Google Scholar] [CrossRef]
- Sies, H. On the history of oxidative stress: Concept and some aspects of current development. Curr. Opin. Toxicol. 2018, 7, 122–126. [Google Scholar] [CrossRef]
- Salim, S. Oxidative stress and the central nervous system. J. Pharmacol. Exp. Ther. 2017, 360, 201–205. [Google Scholar] [CrossRef] [PubMed]
- Sies, H.; Berndt, C.; Jones, D.P. Oxidative stress. Annu. Rev. Biochem. 2017, 86, 715–748. [Google Scholar] [CrossRef] [PubMed]
- Sies, H. Oxidative stress: Concept and some practical aspects. Antioxidants 2020, 9, 852. [Google Scholar] [CrossRef] [PubMed]
- Sies, H.; Jones, D.P. Reactive oxygen species (ROS) as pleiotropic physiological signalling agents. Nat. Rev. Mol. Cell Biol. 2020, 21, 363–383. [Google Scholar] [CrossRef] [PubMed]
- Mitra, A.K. Antioxidants: A Masterpiece of Mother Nature to Prevent Illness. J. Chem. Rev. 2020, 2, 243–256. [Google Scholar]
- Rahal, A.; Kumar, A.; Singh, V.; Yadav, B.; Tiwari, R.; Chakraborty, S.; Dhama, K. Oxidative stress, prooxidants, and antioxidants: The interplay. Biomed. Res. Int. 2014, 2014, 761264. [Google Scholar] [CrossRef]
- Ofosu, F.K.; Elahi, F.; Daliri, E.B.-M.; Tyagi, A.; Chen, X.Q.; Chelliah, R.; Kim, J.-H.; Han, S.-I.; Oh, D.-H. UHPLC-ESI-QTOF-MS/MS characterization, antioxidant and antidiabetic properties of sorghum grains. Food Chem. 2021, 337, 127788. [Google Scholar] [CrossRef]
- Lu, Q.-B. Reaction Cycles of Halogen Species in the Immune Defense: Implications for Human Health and Diseases and the Pathology and Treatment of COVID-19. Cells 2020, 9, 1461. [Google Scholar] [CrossRef]
- Olson, K.R. Reactive oxygen species or reactive sulfur species: Why we should consider the latter. J. Exp. Biol. 2020, 223, jeb196352. [Google Scholar] [CrossRef]
- Parvez, S.; Long, M.J.; Poganik, J.R.; Aye, Y. Redox signaling by reactive electrophiles and oxidants. Chem. Rev. 2018, 118, 8798–8888. [Google Scholar] [CrossRef]
- Semchyshyn, H.M. Reactive carbonyl species in vivo: Generation and dual biological effects. Sci. World J. 2014, 2014, 417842. [Google Scholar] [CrossRef] [PubMed]
- Mano, J.I.; Biswas, M.; Sugimoto, K. Reactive carbonyl species: A missing link in ROS signaling. Plants 2019, 8, 391. [Google Scholar] [CrossRef] [PubMed]
- Lee, B.H.; Lopez-Hilfiker, F.D.; Veres, P.R.; McDuffie, E.E.; Fibiger, D.L.; Sparks, T.L.; Ebben, C.J.; Green, J.R.; Schroder, J.C.; Campuzano-Jost, P. Flight deployment of a high-resolution time-of-flight chemical ionization mass spectrometer: Observations of reactive halogen and nitrogen oxide species. J. Geophys. Res. Atmos. 2018, 123, 7670–7686. [Google Scholar] [CrossRef]
- Möller, M.N.; Rios, N.; Trujillo, M.; Radi, R.; Denicola, A.; Alvarez, B. Detection and quantification of nitric oxide–derived oxidants in biological systems. J. Biol. Chem. 2019, 294, 14776–14802. [Google Scholar] [CrossRef]
- Nakamura, T.; Lipton, S.A. Nitric oxide-dependent protein post-translational modifications impair mitochondrial function and metabolism to contribute to neurodegenerative diseases. Antioxid. Redox Signal. 2020, 32, 817–833. [Google Scholar] [CrossRef] [PubMed]
- Davies, M.J.; Hawkins, C.L. The role of myeloperoxidase in biomolecule modification, chronic inflammation, and disease. Antioxid. Redox Signal. 2020, 32, 957–981. [Google Scholar] [CrossRef]
- Yang, X.; Li, Y.; Li, Y.; Ren, X.; Zhang, X.; Hu, D.; Gao, Y.; Xing, Y.; Shang, H. Oxidative stress-mediated atherosclerosis: Mechanisms and therapies. Front. Physiol. 2017, 8, 600. [Google Scholar] [CrossRef]
- Lushchak, V.I. Free radicals, reactive oxygen species, oxidative stress and its classification. Chem. Biol. Interact. 2014, 224, 164–175. [Google Scholar] [CrossRef]
- Hermes-Lima, M. Oxygen in biology and biochemistry: Role of free radicals. Funct. Metab. Regul. Adapt. 2004, 1, 319–366. [Google Scholar]
- Lushchak, V.I. Environmentally induced oxidative stress in aquatic animals. Aquat. Toxicol. 2011, 101, 13–30. [Google Scholar] [CrossRef]
- Lushchak, V.I. Adaptive response to oxidative stress: Bacteria, fungi, plants and animals. Comp. Biochem. Physiol. Part C Toxicol. Pharmacol. 2011, 153, 175–190. [Google Scholar] [CrossRef]
- Sies, H. (Ed.) Oxidative stress: Introductory remarks. In Oxidative Stress, 1st ed.; Academic Press: London, UK, 1985; pp. 1–8. [Google Scholar]
- Roede, J.R.; Stewart, B.J.; Petersen, D.R. Hepatotoxicity of Reactive Aldehydes. In Comprehensive Toxicology, 2nd ed.; McQueen, C.A., Ed.; Elsevier: Oxford, MA, USA, 2010; Volume 9, pp. 581–594. [Google Scholar]
- El-Fawal, H.A.N. Neurotoxicology. In Encyclopedia of Environmental Health, 1st ed.; Nriagu, J.O., Ed.; Elsevier: Burlington, MA, USA, 2011; Volume 4, pp. 87–106. [Google Scholar]
- Ofosu, F.K.; Mensah, D.J.F.; Daliri, E.B.M.; Lee, B.H.; Oh, D.H. Probiotics, diet, and gut microbiome modulation in metabolic syndromes prevention. In Advances in Probiotics Microorganisms in Food and Health; Dhanasekaran, D., Narayanan, S., Eds.; Elsievier Academic Press: Philadelphia, PA, USA, 2021; in press. [Google Scholar]
- Shankar, K.; Mehendale, H. Encyclopedia of toxicology. Oxidative Stress 2014, 20, 735–737. [Google Scholar]
- Grundy, S.M. Metabolic syndrome. In Diabetes Complications, Comorbidities and Related Disorders, 1st ed.; Bonora, E., DeFronzo, R.A., Eds.; Springer: Cham, Switzerland, 2018; pp. 71–107. [Google Scholar]
- Shoushou, I.M.; Melebari, A.N.; Alalawi, H.A.; Alghaith, T.A.; Alaithan, M.S.; Albriman, M.H.A.; Hawsawi, H. Evaluation of Metabolic Syndrome in Primary Health Care. Int. J. Pharm. Res. Allied Sci. 2020, 9, 52–55. [Google Scholar]
- Swarup, S.; Goyal, A.; Grigorova, Y.; Zeltser, R. Metabolic syndrome. In Statpearls; StatPearls Publishing: Treasure Island, FL, USA, 2020. [Google Scholar]
- Di Daniele, N.; Petramala, L.; Di Renzo, L.; Sarlo, F.; Della Rocca, D.G.; Rizzo, M.; Fondacaro, V.; Iacopino, L.; Pepine, C.J.; De Lorenzo, A. Body composition changes and cardiometabolic benefits of a balanced Italian Mediterranean Diet in obese patients with metabolic syndrome. Acta Diabetol. 2013, 50, 409–416. [Google Scholar] [CrossRef]
- Aguilar, M.; Bhuket, T.; Torres, S.; Liu, B.; Wong, R.J. Prevalence of the metabolic syndrome in the United States, 2003–2012. JAMA 2015, 313, 1973–1974. [Google Scholar] [CrossRef]
- Sigit, F.S.; Tahapary, D.L.; Trompet, S.; Sartono, E.; Van Dijk, K.W.; Rosendaal, F.R.; De Mutsert, R. The prevalence of metabolic syndrome and its association with body fat distribution in middle-aged individuals from Indonesia and the Netherlands: A cross-sectional analysis of two population-based studies. Diabetol. Metab. Syndr. 2020, 12, 1–11. [Google Scholar] [CrossRef]
- Marcos-Delgado, A.; Hernández-Segura, N.; Fernández-Villa, T.; Molina, A.J.; Martín, V. The Effect of Lifestyle Intervention on Health-Related Quality of Life in Adults with Metabolic Syndrome: A Meta-Analysis. Int. J. Environ. Res. Public Health 2021, 18, 887. [Google Scholar] [CrossRef] [PubMed]
- Shin, D.; Kongpakpaisarn, K.; Bohra, C. Trends in the prevalence of metabolic syndrome and its components in the United States 2007–2014. Int. J. Cardiol. 2018, 259, 216–219. [Google Scholar] [CrossRef]
- Lesser, I.A.; Gasevic, D.; Lear, S.A. The effect of body fat distribution on ethnic differences in cardiometabolic risk factors of Chinese and Europeans. Appl. Physiol. Nutr. Metab. 2013, 38, 701–706. [Google Scholar] [CrossRef]
- Furukawa, S.; Fujita, T.; Shimabukuro, M.; Iwaki, M.; Yamada, Y.; Nakajima, Y.; Nakayama, O.; Makishima, M.; Matsuda, M.; Shimomura, I. Increased oxidative stress in obesity and its impact on metabolic syndrome. J. Clin. Investig. 2017, 114, 1752–1761. [Google Scholar] [CrossRef]
- Bonomini, F.; Rodella, L.F.; Rezzani, R. Metabolic syndrome, aging and involvement of oxidative stress. Aging Dis. 2015, 6, 109. [Google Scholar] [CrossRef] [PubMed]
- Li, L.; Yang, X. The essential element manganese, oxidative stress, and metabolic diseases: Links and interactions. Oxid. Med. Cell. Longev. 2018, 2018, 7580707. [Google Scholar] [CrossRef] [PubMed]
- Monserrat-Mesquida, M.; Quetglas-Llabrés, M.; Capó, X.; Bouzas, C.; Mateos, D.; Pons, A.; Tur, J.A.; Sureda, A. Metabolic syndrome is associated with oxidative stress and proinflammatory state. Antioxidants 2020, 9, 236. [Google Scholar] [CrossRef]
- Spahis, S.; Borys, J.-M.; Levy, E. Metabolic syndrome as a multifaceted risk factor for oxidative stress. Antioxid. Redox Signal. 2017, 26, 445–461. [Google Scholar] [CrossRef] [PubMed]
- Manna, P.; Jain, S.K. Obesity, oxidative stress, adipose tissue dysfunction, and the associated health risks: Causes and therapeutic strategies. Metab. Syndr. Relat. Disord. 2015, 13, 423–444. [Google Scholar] [CrossRef]
- Lemes, A.C.; Sala, L.; Ores, J.D.C.; Braga, A.R.C.; Egea, M.B.; Fernandes, K.F. A review of the latest advances in encrypted bioactive peptides from protein-rich waste. Int. J. Mol. Sci. 2016, 17, 950. [Google Scholar] [CrossRef]
- Sánchez, A.; Vázquez, A. Bioactive peptides: A review. Food Qual. Saf. 2017, 1, 29–46. [Google Scholar] [CrossRef]
- Mora, L.; Gallego, M.; Aristoy, M.-C.; Reig, M.; Toldrá, F. Bioactive peptides. In Innovative Thermal and Non-Thermal Processing, Bioaccessibility and Bioavailability of Nutrients and Bioactive Compounds; Barba, F.J., Saraiva, J.M.A., Cravotto, G., Lorenzo, J.M., Eds.; Elsevier: Duxford, UK, 2019; pp. 333–345. [Google Scholar]
- Amigo, L.; Hernández-Ledesma, B. Current evidence on the bioavailability of food bioactive peptides. Molecules 2020, 25, 4479. [Google Scholar] [CrossRef]
- Tadesse, S.A.; Emire, S.A. Production and processing of antioxidant bioactive peptides: A driving force for the functional food market. Heliyon 2020, 6, e04765. [Google Scholar] [CrossRef]
- Esfandi, R.; Walters, M.E.; Tsopmo, A. Antioxidant properties and potential mechanisms of hydrolyzed proteins and peptides from cereals. Heliyon 2019, 5, e01538. [Google Scholar] [CrossRef]
- Micewicz, E.D.; Bahattab, O.S.; Willars, G.B.; Waring, A.J.; Navab, M.; Whitelegge, J.P.; McBride, W.H.; Ruchala, P. Small lipidated anti-obesity compounds derived from neuromedin U. Eur. J. Med. Chem. 2015, 101, 616–626. [Google Scholar] [CrossRef] [PubMed][Green Version]
- Hajfathalian, M.; Ghelichi, S.; García-Moreno, P.J.; Moltke Sørensen, A.-D.; Jacobsen, C. Peptides: Production, bioactivity, functionality, and applications. Crit. Rev. Food Sci. Nutr. 2018, 58, 3097–3129. [Google Scholar] [CrossRef]
- Kumar, P.; Kizhakkedathu, J.N.; Straus, S.K. Antimicrobial peptides: Diversity, mechanism of action and strategies to improve the activity and biocompatibility in vivo. Biomolecules 2018, 8, 4. [Google Scholar] [CrossRef] [PubMed]
- Xu, S.; Fan, F.; Liu, H.; Cheng, S.; Tu, M.; Du, M. Novel anticoagulant peptide from lactoferrin binding thrombin at the active site and exosite-I. J. Agric. Food Chem. 2020, 68, 3132–3139. [Google Scholar] [CrossRef] [PubMed]
- Guha, S.; Majumder, K. Structural-features of food-derived bioactive peptides with anti-inflammatory activity: A brief review. J. Food Biochem. 2019, 43, e12531. [Google Scholar] [CrossRef] [PubMed]
- Liu, Y.-F.; Oey, I.; Bremer, P.; Carne, A.; Silcock, P. Bioactive peptides derived from egg proteins: A review. Crit. Rev. Food Sci. Nutr. 2018, 58, 2508–2530. [Google Scholar] [CrossRef]
- Lee, Y.-M.; Han, S.-I.; Song, B.C.; Yeum, K.-J. Bioactives in commonly consumed cereal grains: Implications for oxidative stress and inflammation. J. Med. Food 2015, 18, 1179–1186. [Google Scholar] [CrossRef]
- Luo, J.; Zhou, Z.; Yao, X.; Fu, Y. Mineral-chelating peptides derived from fish collagen: Preparation, bioactivity and bioavailability. LWT 2020, 134, 110209. [Google Scholar] [CrossRef]
- Amado, I.R.; González, M.P.; Murado, M.A.; Vázquez, J.A. Shrimp wastewater as a source of astaxanthin and bioactive peptides. J. Chem. Technol. Biotechnol. 2016, 91, 793–805. [Google Scholar] [CrossRef]
- Chen, Y.; Chen, J.; Chen, J.; Yu, H.; Zheng, Y.; Zhao, J.; Zhu, J. Recent advances in seafood bioactive peptides and their potential for managing osteoporosis. Crit. Rev. Food Sci. Nutr. 2020, 23, 1–17. [Google Scholar] [CrossRef]
- Gong, H.; Gao, J.; Wang, Y.; Luo, Q.; Guo, K.; Ren, F.; Mao, X. Identification of novel peptides from goat milk casein that ameliorate high-glucose-induced insulin resistance in HepG2 cells. J. Dairy Sci. 2020, 103, 4907–4918. [Google Scholar] [CrossRef] [PubMed]
- Padghan, P.; Mann, B.; Hati, S. Purification and characterization of antioxidative peptides derived from fermented milk (lassi) by lactic cultures. Int. J. Pept. Res. Ther. 2018, 24, 235–249. [Google Scholar] [CrossRef]
- Uraipong, C.; Zhao, J. Identification and functional characterisation of bioactive peptides in rice bran albumin hydrolysates. Int. J. Food Sci. Technol. 2016, 51, 2201–2208. [Google Scholar] [CrossRef]
- Chatterjee, C.; Gleddie, S.; Xiao, C.-W. Soybean bioactive peptides and their functional properties. Nutrients 2018, 10, 1211. [Google Scholar] [CrossRef] [PubMed]
- Montone, C.M.; Capriotti, A.L.; Cavaliere, C.; La Barbera, G.; Piovesana, S.; Chiozzi, R.Z.; Laganà, A. Characterization of antioxidant and angiotensin-converting enzyme inhibitory peptides derived from cauliflower by-products by multidimensional liquid chromatography and bioinformatics. J. Funct. Foods 2018, 44, 40–47. [Google Scholar] [CrossRef]
- Sila, A.; Bougatef, A. Antioxidant peptides from marine by-products: Isolation, identification and application in food systems. A review. J. Funct. Foods 2016, 21, 10–26. [Google Scholar] [CrossRef]
- Wong, F.-C.; Xiao, J.; Wang, S.; Ee, K.-Y.; Chai, T.-T. Advances on the antioxidant peptides from edible plant sources. Trends Food Sci. Technol. 2020, 99, 44–57. [Google Scholar] [CrossRef]
- Aluko, R. Amino acids, peptides, and proteins as antioxidants for food preservation. In Handbook of Antioxidants for Food Preservation, 1st ed.; Shahidi, F., Ed.; Woodhead Publishing: Cambridge, UK, 2015; pp. 105–140. [Google Scholar]
- Power, O.; Jakeman, P.; FitzGerald, R. Antioxidative peptides: Enzymatic production, in vitro and in vivo antioxidant activity and potential applications of milk-derived antioxidative peptides. Amino Acids 2013, 44, 797–820. [Google Scholar] [CrossRef]
- Selamassakul, O.; Laohakunjit, N.; Kerdchoechuen, O.; Yang, L.; Maier, C.S. Bioactive peptides from brown rice protein hydrolyzed by bromelain: Relationship between biofunctional activities and flavor characteristics. J. Food Sci. 2020, 85, 707–717. [Google Scholar] [CrossRef] [PubMed]
- Han, B.K.; Park, Y.; Choi, H.S.; Suh, H.J. Hepatoprotective effects of soluble rice protein in primary hepatocytes and in mice. J. Sci. Food Agric. 2016, 96, 685–694. [Google Scholar] [CrossRef] [PubMed]
- Ren, D.; Zhao, F.; Liu, C.; Wang, J.; Guo, Y.; Liu, J.; Min, W. Antioxidant hydrolyzed peptides from Manchurian walnut (Juglans mandshurica Maxim.) attenuate scopolamine-induced memory impairment in mice. J. Sci. Food Agric. 2018, 98, 5142–5152. [Google Scholar] [CrossRef]
- Wang, L.; Ding, L.; Wang, Y.; Zhang, Y.; Liu, J. Isolation and characterisation of in vitro and cellular free radical scavenging peptides from corn peptide fractions. Molecules 2015, 20, 3221–3237. [Google Scholar] [CrossRef]
- Wang, L.; Ding, L.; Yu, Z.; Zhang, T.; Ma, S.; Liu, J. Intracellular ROS scavenging and antioxidant enzyme regulating capacities of corn gluten meal-derived antioxidant peptides in HepG2 cells. Food Res. Int. 2016, 90, 33–41. [Google Scholar] [CrossRef]
- Sun, C.; Tang, X.; Ren, Y.; Wang, E.; Shi, L.; Wu, X.; Wu, H. Novel antioxidant peptides purified from mulberry (Morus Atropurpurea Roxb.) leaf protein hydrolysates with hemolysis inhibition ability and cellular antioxidant activity. J. Agric. Food Chem. 2019, 67, 7650–7659. [Google Scholar] [CrossRef]
- Chalé, F.G.H.; Ruiz, J.C.R.; Fernández, J.J.A.; Ancona, D.A.B.; Campos, M.R.S. ACE inhibitory, hypotensive and antioxidant peptide fractions from Mucuna pruriens proteins. Process Biochem. 2014, 49, 1691–1698. [Google Scholar] [CrossRef]
- Indiano-Romacho, P.; Fernández-Tomé, S.; Amigo, L.; Hernández-Ledesma, B. Multifunctionality of lunasin and peptides released during its simulated gastrointestinal digestion. Food Res. Int. 2019, 125, 108513. [Google Scholar] [CrossRef] [PubMed]
- Adebayo, J.O.; Adewole, K.E.; Krettli, A.U. Cysteine-stabilised peptide extract of Morinda lucida (Benth) leaf exhibits antimalarial activity and augments antioxidant defense system in P. berghei-infected mice. J. Ethnopharmacol. 2017, 207, 118–128. [Google Scholar] [CrossRef] [PubMed]
- Adewole, K.E.; Adebayo, J.O. Antioxidant defense system induced by cysteine-stabilized peptide fraction of aqueous extract of Morinda lucida leaf in selected tissues of Plasmodium berghei-infected mice. J. Integr. Med. 2017, 15, 388–397. [Google Scholar] [CrossRef]
- Malaguti, M.; Dinelli, G.; Leoncini, E.; Bregola, V.; Bosi, S.; Cicero, A.F.; Hrelia, S. Bioactive peptides in cereals and legumes: Agronomical, biochemical and clinical aspects. Int. J. Mol. Sci. 2014, 15, 21120–21135. [Google Scholar] [CrossRef] [PubMed]
- Nwachukwu, I.D.; Aluko, R.E. Structural and functional properties of food protein-derived antioxidant peptides. J. Food Biochem. 2019, 43, e12761. [Google Scholar] [CrossRef]
- Edelstein, S. Food Science: An Ecological Approach, 1st ed.; Jones & Bartlett Publishers: Burlington, MA, USA, 2014; pp. 1–495. [Google Scholar]
- Ortiz-Martinez, M.; Winkler, R.; García-Lara, S. Preventive and therapeutic potential of peptides from cereals against cancer. J. Proteom. 2014, 111, 165–183. [Google Scholar] [CrossRef]
- Díaz-Gómez, J.L.; Castorena-Torres, F.; Preciado-Ortiz, R.E.; García-Lara, S. Anti-cancer activity of maize bioactive peptides. Front. Chem. 2017, 5, 44. [Google Scholar] [CrossRef] [PubMed]
- Thielecke, F.; Lecerf, J.-M.; Nugent, A.P. Processing in the food chain: Do cereals have to be processed to add value to the human diet? Nutr. Res. Rev. 2020, 28, 1–15. [Google Scholar] [CrossRef] [PubMed]
- Walsh, J.; Haddock, J.; Blumberg, J.B.; McKay, D.L.; Wei, X.; Dolnikowski, G.; Chen, C.-Y.O. Identification of methylated metabolites of oat avenanthramides in human plasma using UHPLC QToF-MS. Int. J. Food Sci. Nutr. 2018, 69, 377–383. [Google Scholar] [CrossRef]
- Green, P.H.; Lebwohl, B.; Greywoode, R. Celiac disease. J. Allergy Clin. Immunol. 2015, 135, 1099–1106. [Google Scholar] [CrossRef]
- Syage, J.A.; Kelly, C.P.; Dickason, M.A.; Ramirez, A.C.; Leon, F.; Dominguez, R.; Sealey-Voyksner, J.A. Determination of gluten consumption in celiac disease patients on a gluten-free diet. Am. J. Clin. Nutr. 2018, 107, 201–207. [Google Scholar] [CrossRef]
- Cian, R.E.; Vioque, J.; Drago, S.R. Structure–mechanism relationship of antioxidant and ACE I inhibitory peptides from wheat gluten hydrolysate fractionated by pH. Food Res. Int. 2015, 69, 216–223. [Google Scholar] [CrossRef]
- Ngamsuk, S.; Hsu, J.-L.; Huang, T.-C.; Suwannaporn, P. Ultrasonication of milky stage rice milk with bioactive peptides from rice bran: Its bioactivities and absorption. Food Bioprocess Technol. 2020, 13, 462–474. [Google Scholar] [CrossRef]
- Bamdad, F.; Wu, J.; Chen, L. Effects of enzymatic hydrolysis on molecular structure and antioxidant activity of barley hordein. J. Cereal Sci. 2011, 54, 20–28. [Google Scholar] [CrossRef]
- Giriwono, P.E.; Hashimoto, T.; Ohsaki, Y.; Shirakawa, H.; Hokazono, H.; Komai, M. Extract of fermented barley attenuates chronic alcohol induced liver damage by increasing antioxidative activities. Food Res. Int. 2010, 43, 118–124. [Google Scholar] [CrossRef]
- Tok, K.; Moulahoum, H.; Kocadag Kocazorbaz, E.; Zihnioglu, F. Bioactive peptides with multiple activities extracted from Barley (Hordeum vulgare L.) grain protein hydrolysates: Biochemical analysis and computational identification. J. Food Process. Preserv. 2021, 45, e15024. [Google Scholar] [CrossRef]
- Leung, R.; Venus, C.; Zeng, T.; Tsopmo, A. Structure-function relationships of hydroxyl radical scavenging and chromium-VI reducing cysteine-tripeptides derived from rye secalin. Food Chem. 2018, 254, 165–169. [Google Scholar] [CrossRef]
- Coda, R.; Rizzello, C.G.; Pinto, D.; Gobbetti, M. Selected lactic acid bacteria synthesize antioxidant peptides during sourdough fermentation of cereal flours. Appl. Environ. Microbiol. 2012, 78, 1087–1096. [Google Scholar] [CrossRef]
- Tang, N.; Zhuang, H. Evaluation of antioxidant activities of zein protein fractions. J. Food Sci. 2014, 79, C2174–C2184. [Google Scholar] [CrossRef] [PubMed]
- Jin, D.-X.; Liu, X.-L.; Zheng, X.-Q.; Wang, X.-J.; He, J.-F. Preparation of antioxidative corn protein hydrolysates, purification and evaluation of three novel corn antioxidant peptides. Food Chem. 2016, 204, 427–436. [Google Scholar] [CrossRef]
- Majid, A.; Priyadarshini CG, P. Millet derived bioactive peptides: A review on their functional properties and health benefits. Crit. Rev. Food Sci. Nutr. 2020, 60, 3342–3351. [Google Scholar] [CrossRef]
- Bandyopadhyay, T.; Muthamilarasan, M.; Prasad, M. Millets for next generation climate-smart agriculture. Front. Plant Sci. 2017, 8, 1266. [Google Scholar] [CrossRef]
- Anitha, S.; Govindaraj, M.; Kane-Potaka, J. Balanced amino acid and higher micronutrients in millets complements legumes for improved human dietary nutrition. Cereal Chem. 2020, 97, 74–84. [Google Scholar] [CrossRef]
- Hu, S.; Yuan, J.; Gao, J.; Wu, Y.; Meng, X.; Tong, P.; Chen, H. Antioxidant and Anti-Inflammatory Potential of Peptides Derived from In Vitro Gastrointestinal Digestion of Germinated and Heat-Treated Foxtail Millet (Setaria italica) Proteins. J. Agric. Food Chem. 2020, 68, 9415–9426. [Google Scholar] [CrossRef] [PubMed]
- Agrawal, H.; Joshi, R.; Gupta, M. Purification, identification and characterization of two novel antioxidant peptides from finger millet (Eleusine coracana) protein hydrolysate. Food Res. Int. 2019, 120, 697–707. [Google Scholar] [CrossRef] [PubMed]
- Agrawal, H.; Joshi, R.; Gupta, M. Isolation and characterisation of enzymatic hydrolysed peptides with antioxidant activities from green tender sorghum. LWT Food Sci. Technol. 2017, 84, 608–616. [Google Scholar] [CrossRef]
- de Morais Cardoso, L.; Pinheiro, S.S.; Martino, H.S.D.; Pinheiro-Sant’Ana, H.M. Sorghum (Sorghum bicolor L.): Nutrients, bioactive compounds, and potential impact on human health. Crit. Rev. Food Sci. Nutr. 2017, 57, 372–390. [Google Scholar] [CrossRef] [PubMed]
- Mohajeri, M.H.; Eckert, G.P.; Pauly, J.R.; Butt, C.M. Pharmacology: The Pharmacodynamics of Nutrients and Nutrient Interactions in Biological Functions. Biomed Res. Int. 2015, 2015, 974572. [Google Scholar] [CrossRef]
- Stefoska-Needham, A.; Beck, E.J.; Johnson, S.K.; Tapsell, L.C. Sorghum: An underutilized cereal whole grain with the potential to assist in the prevention of chronic disease. Food Rev. Int. 2015, 31, 401–437. [Google Scholar] [CrossRef]
- Massoud, M.I.; Abd El-Razek, A.M. Suitability of Sorghum bicolor L. stalks and grains for bioproduction of ethanol. Ann. Agric. Sci. 2011, 56, 83–87. [Google Scholar] [CrossRef][Green Version]
- Moraes, É.A.; Natal, D.I.G.; Queiroz, V.A.V.; Schaffert, R.E.; Cecon, P.R.; de Paula, S.O.; dos Anjos Benjamim, L.; Ribeiro, S.M.R.; Martino, H.S.D. Sorghum genotype may reduce low-grade inflammatory response and oxidative stress and maintains jejunum morphology of rats fed a hyperlipidic diet. Food Res. Int. 2012, 49, 553–559. [Google Scholar] [CrossRef]
- Zou, Z.; Wang, M.; Wang, Z.; Aluko, R.E.; He, R. Antihypertensive and antioxidant activities of enzymatic wheat bran protein hydrolysates. J. Food Biochem. 2020, 44, e13090. [Google Scholar] [CrossRef]
- Zhao, Y.; Zhao, Q.; Lu, Q. Purification, structural analysis, and stability of antioxidant peptides from purple wheat bran. BMC Chem. 2020, 14, 1–12. [Google Scholar] [CrossRef]
- Karami, Z.; Peighambardoust, S.H.; Hesari, J.; Akbari-Adergani, B.; Andreu, D. Identification and synthesis of multifunctional peptides from wheat germ hydrolysate fractions obtained by proteinase K digestion. J. Food Biochem. 2019, 43, e12800. [Google Scholar] [CrossRef] [PubMed]
- Suetsuna, K.; Chen, J.-R. Isolation and characterization of peptides with antioxidant activity derived from wheat gluten. Food Sci. Technol. Res. 2002, 8, 227–230. [Google Scholar] [CrossRef]
- Karami, Z.; Peighambardoust, S.H.; Hesari, J.; Akbari-Adergani, B.; Andreu, D. Antioxidant, anticancer and ACE-inhibitory activities of bioactive peptides from wheat germ protein hydrolysates. Food Biosci. 2019, 32, 100450. [Google Scholar] [CrossRef]
- Elmalimadi, M.B.; Stefanović, A.B.; Šekuljica, N.Ž.; Žuža, M.G.; Luković, N.D.; Jovanović, J.R.; Knežević-Jugović, Z.D. The synergistic effect of heat treatment on alcalase-assisted hydrolysis of wheat gluten proteins: Functional and antioxidant properties. J. Food Process. Preserv. 2017, 41, e13207. [Google Scholar] [CrossRef]
- Zaky, A.A.; Chen, Z.; Liu, Y.; Li, S.; Jia, Y. Preparation and assessment of bioactive extracts having antioxidant activity from rice bran protein hydrolysates. J. Food Meas. Charact. 2019, 13, 2542–2548. [Google Scholar] [CrossRef]
- Wang, X.; Chen, H.; Fu, X.; Li, S.; Wei, J. A novel antioxidant and ACE inhibitory peptide from rice bran protein: Biochemical characterization and molecular docking study. Lwt 2017, 75, 93–99. [Google Scholar] [CrossRef]
- Selamassakul, O.; Laohakunjit, N.; Kerdchoechuen, O.; Yang, L.; Maier, C.S. Isolation and characterisation of antioxidative peptides from bromelain-hydrolysed brown rice protein by proteomic technique. Process Biochem. 2018, 70, 179–187. [Google Scholar] [CrossRef]
- Zaky, A.A.; Liu, Y.; Han, P.; Chen, Z.; Jia, Y. Effect of Pepsin–Trypsin In Vitro Gastro-Intestinal Digestion on the Antioxidant Capacities of Ultra-Filtrated Rice Bran Protein Hydrolysates (Molecular Weight >10 kDa; 3–10 kDa, and <3 kDa). Int. J. Pept. Res. Ther. 2020, 26, 1661–1667. [Google Scholar] [CrossRef]
- Pantoa, T.; Kubota, M.; Suwannaporn, P.; Kadowaki, M. Characterization and bioactivities of young rice protein hydrolysates. J. Cereal Sci. 2020, 95, 103049. [Google Scholar] [CrossRef]
- Li, H.; Wang, Z.; Liang, M.; Cai, L.; Yang, L. Methionine augments antioxidant activity of rice protein during gastrointestinal digestion. Int. J. Mol. Sci. 2019, 20, 868. [Google Scholar] [CrossRef]
- Phongthai, S.; D’Amico, S.; Schoenlechner, R.; Homthawornchoo, W.; Rawdkuen, S. Fractionation and antioxidant properties of rice bran protein hydrolysates stimulated by in vitro gastrointestinal digestion. Food Chem. 2018, 240, 156–164. [Google Scholar] [CrossRef] [PubMed]
- Gomes, M.H.G.; Kurozawa, L.E. Improvement of the functional and antioxidant properties of rice protein by enzymatic hydrolysis for the microencapsulation of linseed oil. J. Food Eng. 2020, 267, 109761. [Google Scholar] [CrossRef]
- Zhou, K.; Canning, C.; Sun, S. Effects of rice protein hydrolysates prepared by microbial proteases and ultrafiltration on free radicals and meat lipid oxidation. LWT Food Sci. Technol. 2013, 50, 331–335. [Google Scholar] [CrossRef]
- Yan, Q.-J.; Huang, L.-H.; Sun, Q.; Jiang, Z.-Q.; Wu, X. Isolation, identification and synthesis of four novel antioxidant peptides from rice residue protein hydrolyzed by multiple proteases. Food Chem. 2015, 179, 290–295. [Google Scholar] [CrossRef] [PubMed]
- Shen, Y.; Hu, R.; Li, Y. Antioxidant and Emulsifying Activities of Corn Gluten Meal Hydrolysates in Oil-in-Water Emulsions. J. Am. Oil Chem. Soc. 2020, 97, 175–185. [Google Scholar] [CrossRef]
- Zhang, S.; Zhang, M.; Yang, R.; Zhang, S.; Lin, S. Preparation, identification, and activity evaluation of antioxidant peptides from protein hydrolysate of corn germ meal. J. Food Process. Preserv. 2019, 43, e14160. [Google Scholar] [CrossRef]
- Hu, R.; Dunmire, K.M.; Truelock, C.N.; Paulk, C.B.; Aldrich, G.; Li, Y. Antioxidant performances of corn gluten meal and DDGS protein hydrolysates in food, pet food, and feed systems. J. Agric. Food Res. 2020, 2, 100030. [Google Scholar] [CrossRef]
- Zhou, K.; Sun, S.; Canning, C. Production and functional characterisation of antioxidative hydrolysates from corn protein via enzymatic hydrolysis and ultrafiltration. Food Chem. 2012, 135, 1192–1197. [Google Scholar] [CrossRef]
- Wang, X.-J.; Zheng, X.-Q.; Kopparapu, N.-K.; Cong, W.-S.; Deng, Y.-P.; Sun, X.-J.; Liu, X.-L. Purification and evaluation of a novel antioxidant peptide from corn protein hydrolysate. Process Biochem. 2014, 49, 1562–1569. [Google Scholar] [CrossRef]
- Ren, X.; Liang, Q.; Zhang, X.; Hou, T.; Li, S.; Ma, H. Stability and antioxidant activities of corn protein hydrolysates under simulated gastrointestinal digestion. Cereal Chem. 2018, 95, 760–769. [Google Scholar] [CrossRef]
- Hu, R.; Chen, G.; Li, Y. Production and Characterization of Antioxidative Hydrolysates and Peptides from Corn Gluten Meal Using Papain, Ficin, and Bromelain. Molecules 2020, 25, 4091. [Google Scholar] [CrossRef] [PubMed]
- Xu, S.; Shen, Y.; Li, Y. Antioxidant activities of sorghum kafirin alcalase hydrolysates and membrane/gel filtrated fractions. Antioxidants 2019, 8, 131. [Google Scholar] [CrossRef] [PubMed]
- Xu, S.; Shen, Y.; Xu, J.; Qi, G.; Chen, G.; Wang, W.; Sun, X.; Li, Y. Antioxidant and anticancer effects in human hepatocarcinoma (HepG2) cells of papain-hydrolyzed sorghum kafirin hydrolysates. J. Funct. Foods 2019, 58, 374–382. [Google Scholar] [CrossRef]
- Xu, S.; Shen, Y.; Chen, G.; Bean, S.; Li, Y. Antioxidant characteristics and identification of peptides from sorghum kafirin hydrolysates. J. Food Sci. 2019, 84, 2065–2076. [Google Scholar] [CrossRef] [PubMed]
- Marciniak, A.; Suwal, S.; Naderi, N.; Pouliot, Y.; Doyen, A. Enhancing enzymatic hydrolysis of food proteins and production of bioactive peptides using high hydrostatic pressure technology. Trends Food Sci. Technol. 2018, 80, 187–198. [Google Scholar] [CrossRef]
- Wen, C.; Zhang, J.; Zhang, H.; Duan, Y.; Ma, H. Plant protein-derived antioxidant peptides: Isolation, identification, mechanism of action and application in food systems: A review. Trends Food Sci. Technol. 2020, 105, 308–322. [Google Scholar] [CrossRef]
- Samaranayaka, A.G.; Li-Chan, E.C. Food-derived peptidic antioxidants: A review of their production, assessment, and potential applications. J. Funct. Foods 2011, 3, 229–254. [Google Scholar] [CrossRef]
- Di Bernardini, R.; Rai, D.K.; Bolton, D.; Kerry, J.; O’Neill, E.; Mullen, A.M.; Harnedy, P.; Hayes, M. Isolation, purification and characterization of antioxidant peptidic fractions from a bovine liver sarcoplasmic protein thermolysin hydrolyzate. Peptides 2011, 32, 388–400. [Google Scholar] [CrossRef] [PubMed]
- Karami, Z.; Akbari-Adergani, B. Bioactive food derived peptides: A review on correlation between structure of bioactive peptides and their functional properties. J. Food Sci. Technol. 2019, 56, 535–547. [Google Scholar] [CrossRef]
- Daliri, E.B.-M.; Lee, B.H.; Oh, D.H. Current trends and perspectives of bioactive peptides. Crit. Rev. Food Sci. Nutr. 2018, 58, 2273–2284. [Google Scholar] [CrossRef]
- Gänzle, M.G. Enzymatic and bacterial conversions during sourdough fermentation. Food Microbiol. 2014, 37, 2–10. [Google Scholar] [CrossRef]
- Raveschot, C.; Cudennec, B.; Coutte, F.; Flahaut, C.; Fremont, M.; Drider, D.; Dhulster, P. Production of bioactive peptides by Lactobacillus species: From gene to application. Front. Microbiol. 2018, 9, 2354. [Google Scholar] [CrossRef]
- Gobbetti, M.; De Angelis, M.; Corsetti, A.; Di Cagno, R. Biochemistry and physiology of sourdough lactic acid bacteria. Trends Food Sci. Technol. 2005, 16, 57–69. [Google Scholar] [CrossRef]
- Galli, V.; Mazzoli, L.; Luti, S.; Venturi, M.; Guerrini, S.; Paoli, P.; Vincenzini, M.; Granchi, L.; Pazzagli, L. Effect of selected strains of lactobacilli on the antioxidant and anti-inflammatory properties of sourdough. Int. J. Food Microbiol. 2018, 286, 55–65. [Google Scholar] [CrossRef] [PubMed]
- Galli, V.; Venturi, M.; Pini, N.; Guerrini, S.; Granchi, L.; Vincenzini, M. Liquid and firm sourdough fermentation: Microbial robustness and interactions during consecutive backsloppings. LWT 2019, 105, 9–15. [Google Scholar] [CrossRef]
- Luti, S.; Mazzoli, L.; Ramazzotti, M.; Galli, V.; Venturi, M.; Marino, G.; Lehmann, M.; Guerrini, S.; Granchi, L.; Paoli, P. Antioxidant and anti-inflammatory properties of sourdoughs containing selected Lactobacilli strains are retained in breads. Food Chem. 2020, 322, 126710. [Google Scholar] [CrossRef] [PubMed]
- Wu, J.; Cheng, Y.; Dong, Y. Antioxidant activity of Lactobacillus plantarum DY-1 fermented wheat germ extract and its influence on lipid oxidation and texture properties of emulsified sausages. J. Food Qual. 2020, 2020, 8885886. [Google Scholar] [CrossRef]
- Niu, L.-Y.; Jiang, S.-T.; Pan, L.-J. Preparation and evaluation of antioxidant activities of peptides obtained from defatted wheat germ by fermentation. J. Food Sci. Technol. 2013, 50, 53–61. [Google Scholar] [CrossRef]
- Wang, K.; Niu, M.; Song, D.; Liu, Y.; Wu, Y.; Zhao, J.; Li, S.; Lu, B. Evaluation of biochemical and antioxidant dynamics during the co-fermentation of dehusked barley with Rhizopus oryzae and Lactobacillus plantarum. J. Food Biochem. 2020, 44, e13106. [Google Scholar] [CrossRef]
- Sánchez-López, F.; Robles-Olvera, V.J.; Hidalgo-Morales, M.; Tsopmo, A. Characterization of Amaranthus hypochondriacus seed protein fractions, and their antioxidant activity after hydrolysis with lactic acid bacteria. J. Cereal Sci. 2020, 95, 103075. [Google Scholar] [CrossRef]
- Li-Chan, E.C. Bioactive peptides and protein hydrolysates: Research trends and challenges for application as nutraceuticals and functional food ingredients. Curr. Opin. Food Sci. 2015, 1, 28–37. [Google Scholar] [CrossRef]
- Tu, M.; Cheng, S.; Lu, W.; Du, M. Advancement and prospects of bioinformatics analysis for studying bioactive peptides from food-derived protein: Sequence, structure, and functions. Trac Trends Anal. Chem. 2018, 105, 7–17. [Google Scholar] [CrossRef]
- Agyei, D.; Pan, S.; Acquah, C.; Bekhit, A.E.D.A.; Danquah, M.K. Structure-informed detection and quantification of peptides in food and biological fluids. J. Food Biochem. 2019, 43, e12482. [Google Scholar] [CrossRef] [PubMed]
- Nongonierma, A.B.; Mazzocchi, C.; Paolella, S.; FitzGerald, R.J. Release of dipeptidyl peptidase IV (DPP-IV) inhibitory peptides from milk protein isolate (MPI) during enzymatic hydrolysis. Food Res. Int. 2017, 94, 79–89. [Google Scholar] [CrossRef]
- Guedes, I.A.; de Magalhães, C.S.; Dardenne, L.E. Receptor–ligand molecular docking. Biophys. Rev. 2014, 6, 75–87. [Google Scholar] [CrossRef]
- Yap, P.G.; Gan, C.Y. In vivo challenges of anti-diabetic peptide therapeutics: Gastrointestinal stability, toxicity and allergenicity. Trends Food Sci. Technol. 2020, 105, 161–175. [Google Scholar] [CrossRef]
- Nongonierma, A.B.; FitzGerald, R.J. Learnings from quantitative structure–activity relationship (QSAR) studies with respect to food protein-derived bioactive peptides: A review. RSC Adv. 2016, 6, 75400–75413. [Google Scholar] [CrossRef]
- Guo, H.; Wang, Y.; He, Q.; Zhang, Y.; Hu, Y.; Wang, Y.; Lin, Z. In silico rational design and virtual screening of antixoidant tripeptides based on 3D-QSAR modeling. J. Mol. Struct. 2019, 1193, 223–230. [Google Scholar] [CrossRef]
- Yan, W.; Lin, G.; Zhang, R.; Liang, Z.; Wu, W. Studies on the bioactivities and molecular mechanism of antioxidant peptides by 3D-QSAR, in vitro evaluation and molecular dynamic simulations. Food Funct. 2020, 11, 3043–3052. [Google Scholar] [CrossRef]
- Bermúdez-Aguirre, D.; Barbosa-Cánovas, G.V. An update on high hydrostatic pressure, from the laboratory to industrial applications. Food Eng. Rev. 2011, 3, 44–61. [Google Scholar] [CrossRef]
- Bonomi, F.; Fiocchi, A.; Frøkiær, H.; Gaiaschi, A.; Iametti, S.; Poiesi, C.; Rasmussen, P.; Restani, P.; Rovere, P. Reduction of immunoreactivity of bovine [Beta]-lactoglobulin upon combined physical and proteolytic treatment. J. Dairy Res. 2003, 70, 51. [Google Scholar] [CrossRef] [PubMed]
- Chemat, F.; Rombaut, N.; Sicaire, A.-G.; Meullemiestre, A.; Fabiano-Tixier, A.-S.; Abert-Vian, M. Ultrasound assisted extraction of food and natural products. Mechanisms, techniques, combinations, protocols and applications. A review. Ultrason. Sonochem. 2017, 34, 540–560. [Google Scholar] [CrossRef]
- Sullivan, A.C.; Pangloli, P.; Dia, V.P. Impact of ultrasonication on the physicochemical properties of sorghum kafirin and in vitro pepsin-pancreatin digestibility of sorghum gluten-like flour. Food Chem. 2018, 240, 1121–1130. [Google Scholar] [CrossRef] [PubMed]
- Kuan, Y.-H.; Bhat, R.; Patras, A.; Karim, A.A. Radiation processing of food proteins—A review on the recent developments. Trends Food Sci. Technol. 2013, 30, 105–120. [Google Scholar] [CrossRef]
- Wang, L.; Li, T.; Sun, D.; Tang, M.; Sun, Z.; Chen, L.; Luo, X.; Li, Y.; Wang, R.; Li, Y. Effect of electron beam irradiation on the functional properties and antioxidant activity of wheat germ protein hydrolysates. Innov. Food Sci. Emerg. Technol. 2019, 54, 192–199. [Google Scholar] [CrossRef]
- Li, T.; Wang, L.; Sun, D.; Li, Y.; Chen, Z. Effect of enzymolysis-assisted electron beam irradiation on structural characteristics and antioxidant activity of rice protein. J. Cereal Sci. 2019, 89, 102789. [Google Scholar] [CrossRef]
- Liu, F.; Chen, Z.; Shao, J.; Wang, C.; Zhan, C. Effect of fermentation on the peptide content, phenolics and antioxidant activity of defatted wheat germ. Food Biosci. 2017, 20, 141–148. [Google Scholar] [CrossRef]
- Babini, E.; Tagliazucchi, D.; Martini, S.; Dei Più, L.; Gianotti, A. LC-ESI-QTOF-MS identification of novel antioxidant peptides obtained by enzymatic and microbial hydrolysis of vegetable proteins. Food Chem. 2017, 228, 186–196. [Google Scholar] [CrossRef] [PubMed]
- Zhang, J.; Wen, C.; Li, C.; Duan, Y.; Zhang, H.; Ma, H. Antioxidant Peptide Fractions Isolated from Wheat Germ Protein with Subcritical Water Extraction and Its Transport Across Caco-2 Cells. J. Food Sci. 2019, 84, 2139–2146. [Google Scholar] [CrossRef]
- Chen, M.-L.; Ning, P.; Jiao, Y.; Xu, Z.; Cheng, Y.-H. Extraction of antioxidant peptides from rice dreg protein hydrolysate via an angling method. Food Chem. 2021, 337, 128069. [Google Scholar] [CrossRef]
- Liang, Q.; Ren, X.; Ma, H.; Li, S.; Xu, K.; Oladejo, A.O. Effect of low-frequency ultrasonic-assisted enzymolysis on the physicochemical and antioxidant properties of corn protein hydrolysates. J. Food Qual. 2017, 2017, 2784146. [Google Scholar] [CrossRef]
- Huang, D.; Ou, B.; Prior, R.L. The chemistry behind antioxidant capacity assays. J. Agric. Food Chem. 2005, 53, 1841–1856. [Google Scholar] [CrossRef]
- Wang, F.; Weng, Z.; Lyu, Y.; Bao, Y.; Liu, J.; Zhang, Y.; Sui, X.; Fang, Y.; Tang, X.; Shen, X. Wheat germ-derived peptide ADWGGPLPH abolishes high glucose-induced oxidative stress via modulation of the PKCζ/AMPK/NOX4 pathway. Food Funct. 2020, 11, 6843–6854. [Google Scholar] [CrossRef] [PubMed]
- Itoh, K.; Tong, K.I.; Yamamoto, M. Molecular mechanism activating Nrf2–Keap1 pathway in regulation of adaptive response to electrophiles. Free Radic. Biol. Med. 2004, 36, 1208–1213. [Google Scholar] [CrossRef]
- Kobayashi, M.; Yamamoto, M. Nrf2-Keap1 regulation of cellular defense mechanisms against electrophiles and reactive oxygen species. Adv. Enzym. Regul. 2006, 46, 113–140. [Google Scholar] [CrossRef] [PubMed]
- Moritani, C.; Kawakami, K.; Shimoda, H.; Hatanaka, T.; Suzaki, E.; Tsuboi, S. Protective Effects of Rice Peptide Oryza Peptide-P60 against Oxidative Injury through Activation of Nrf2 Signaling Pathway In Vitro and In Vivo. ACS Omega 2020, 5, 13096–13107. [Google Scholar] [CrossRef] [PubMed]
- Sullivan, A.C.; Pangloli, P.; Dia, V.P. Kafirin from Sorghum bicolor inhibition of inflammation in THP-1 human macrophages is associated with reduction of intracellular reactive oxygen species. Food Chem. Toxicol. 2018, 111, 503–510. [Google Scholar] [CrossRef] [PubMed]
- Liang, Y.; Lin, Q.; Huang, P.; Wang, Y.; Li, J.; Zhang, L.; Cao, J. Rice bioactive peptide binding with TLR4 to overcome H2O2-induced injury in human umbilical vein endothelial cells through NF-κB signaling. J. Agric. Food Chem. 2018, 66, 440–448. [Google Scholar] [CrossRef]
- Cruz-Chamorro, I.; Álvarez-Sánchez, N.; Santos-Sánchez, G.; Pedroche, J.; Fernández-Pachón, M.-S.; Millán, F.; Millán-Linares, M.C.; Lardone, P.J.; Bejarano, I.; Guerrero, J.M. Immunomodulatory and antioxidant properties of wheat gluten protein hydrolysates in human peripheral blood mononuclear cells. Nutrients 2020, 12, 1673. [Google Scholar] [CrossRef] [PubMed]
- Ji, Z.; Mao, J.; Chen, S.; Mao, J. Antioxidant and anti-inflammatory activity of peptides from foxtail millet (Setaria italica) prolamins in HaCaT cells and RAW264. 7 murine macrophages. Food Biosci. 2020, 36, 100636. [Google Scholar] [CrossRef]
- Wang, L.; Ding, L.; Xue, C.; Ma, S.; Du, Z.; Zhang, T.; Liu, J. Corn gluten hydrolysate regulates the expressions of antioxidant defense and ROS metabolism relevant genes in H2O2-induced HepG2 cells. J. Funct. Foods 2018, 42, 362–370. [Google Scholar] [CrossRef]
- Reagan-Shaw, S.; Nihal, M.; Ahmad, N. Dose translation from animal to human studies revisited. FASEB J. 2008, 22, 659–661. [Google Scholar] [CrossRef]
- Chen, S.; Lin, D.; Gao, Y.; Cao, X.; Shen, X. A novel antioxidant peptide derived from wheat germ prevents high glucose-induced oxidative stress in vascular smooth muscle cells in vitro. Food Funct. 2017, 8, 142–150. [Google Scholar] [CrossRef] [PubMed]
- Díaz-Gómez, J.L.; Ortíz-Martínez, M.; Aguilar, O.; García-Lara, S.; Castorena-Torres, F. Antioxidant activity of zein hydrolysates from Zea species and their cytotoxic effects in a hepatic cell culture. Molecules 2018, 23, 312. [Google Scholar] [CrossRef] [PubMed]
- Kawaguchi, T.; Ueno, T.; Nogata, Y.; Hayakawa, M.; Koga, H.; Torimura, T. Wheat-bran autolytic peptides containing a branched-chain amino acid attenuate non-alcoholic steatohepatitis via the suppression of oxidative stress and the upregulation of AMPK/ACC in high-fat diet-fed mice. Int. J. Mol. Med. 2017, 39, 407–414. [Google Scholar] [CrossRef]
- Jiang, X.; Cui, Z.; Wang, L.; Xu, H.; Zhang, Y. Production of bioactive peptides from corn gluten meal by solid-state fermentation with Bacillus subtilis MTCC5480 and evaluation of its antioxidant capacity in vivo. LWT 2020, 131, 109767. [Google Scholar] [CrossRef]
- Yu, Y.; Wang, L.; Wang, Y.; Lin, D.; Liu, J. Hepatoprotective effect of albumin peptides from corn germ meal on chronic alcohol-induced liver injury in mice. J. Food Sci. 2017, 82, 2997–3004. [Google Scholar] [CrossRef]




{kind=link}
{kind=link}
{kind=link}
| Definition of Oxidative Stress | Reference |
|---|---|
| Oxidative stress is a disturbance in the prooxidant–antioxidant balance in favor of the former. | [24] |
| Oxidative stress is defined as a disturbance in the prooxidant–antioxidant balance that leads to potential damage. | [25] |
| Oxidative stress is a situation when steady-state ROS concentration is transiently or chronically enhanced, disturbing cellular metabolism, its regulation, and damaging cellular constituents. | [20] |
| Oxidative stress is defined as excess production of reactive oxygen species (ROS) relative to antioxidant defense. | [26] |
| Oxidative stress refers to the imbalance between cellular antioxidant cascade and processes that generate reactive oxygen species (ROS), such as superoxide (O2.), hydrogen peroxide (H2O2), and hydroxyl anion (OH−). | [27] |
| Oxidative stress is defined as an imbalance between oxidants and antioxidants in favor of the oxidants, leading to a disruption of redox signaling and control and/or molecular damage. This implies a deviation to the opposite side of the balance (thus, “reductive stress”), as well as physiological and supraphysiological deviations, which tie into the overarching concept of “oxidative distress” and “oxidative eustress”, respectively. | [5] |
| Oxidative Stress | Definition | Reference |
|---|---|---|
| Acute | The inability of cells to neutralize enhanced ROS level over a period of time, such that the time is enough to result in specific physiological consequences. | [20,21,22,23] |
| Chronic | It occurs when acute oxidative stress progresses to significantly disturb homeostasis. Here, the ROS levels are elevated and very stable, and very potent in altering healthy cell components. | [20] |
| Quasi-stationery | It occurs when ROS levels are so elevated that it is almost impossible to return to ideal homeostatic levels, thus resulting in the need for a substantial reorganization of the entire homeostatic system. | [22,23] |
| Cereal | Family | Sub-Family | Tribe | Genus | Reference |
|---|---|---|---|---|---|
| Wheat | Gramineae or Poaccac | Pooideae Bentham | Triticeae | Triticale | [49,56] |
| Rye | Pooideae Bentham | Triticinae | Triticale | [49,56] | |
| Barley | Pooideae Bentham | Hordeinae | Triticale | [49,56] | |
| Oat | Pooideae Bentham | Aveneae | Aveneae | [56] | |
| Millet | Panicoideae Link | Paniceae | Panicease | [49] | |
| Sorghum | Panicoideae Link | Andropogoneae | Sorghum | [49] | |
| Maize | Panicoideae Link | Maydeae | Zea | [49,56] | |
| Rice | Ehrhartoideae Link | Oryzeae | Oryza | [49,56] |
| Protein Source | Enzyme | Peptide Sequence/Hydrolysate | Size | Assay Outcome | Reference |
|---|---|---|---|---|---|
| Wheat bran | Alcalase | NL, QL, FL, HAL, AAVL, AKTVF, and TPLTR | <1, 1–10 kDa, (membrane ultrafiltration) | Higher ORAC by <1 kDa fraction. | [108] |
| Wheat bran | Alcalase, favourzyme, papain, neutral enzyme, trypsin | CGFPGHC, QAC, RNF, SSC, WF | <1 kDa, (size-exclusion and and ion-exchange chromatography) | QAC and SSC from alcalase hydrolysate showed the highest superoxide anion radical scavenging and Trolox equivalent antioxidant capacity. | [109] |
| Wheat germ | Alcalase, pepsin or proteinase K | KELPPSDADW, SGGSYADELVSTAK, GNPIPREPGQVPAY | <2.5 kDa, (RP-HPLC) | Pepsin fraction showed the strongest ABTS and DPPH radical scavenging activity. | [110] |
| Wheat gluten | Pepsin | LQPGQGQQG, AQIPQQ | <5 kDa, (ion-exchange and gel filtration chromatography) | Fraction 3 showed stronger antioxidant activity using ferric thiocyanate method. | [111] |
| Wheat germ | proteinase K | SGGSYADELVSTAK, MDATALHYENQK | <3 kDa, (RP-HPLC) | This fraction displayed strong ABTS and DPPH radical scavenging activity. | [112] |
| Wheat gluten | Alcalase | Hydrolysate | Not determined | Heat and alcalase treated hydrolysate synergistically increased DPPH and ABTS radical scavenging inhibition. | [113] |
| Rice bran | Alcalase, Trypsin, Protamex, Flavourzyme | Hydrolysate | N.R | Flavourzyme hydrolysate exhibited the strongest DPPH, ABTS radical scavenging and metal chelating activity. | [114] |
| Rice bran | Trypsin | YSK | <1 kDa, (gel filtration, RP-HPLC) | F2-a fraction displayed strong DPPH radical scavenging and reducing power activity | [115] |
| Brown rice | Bromelain | GSGVGGAK, SSVGGGSAG, FGGSGGPGG, FGGGGAGAGG, GGGGGAAAAGA, AGGGGGGVVAG, SGPSGGGGAL, ESDVVSDL | <1 kDa, (ultra, gel filtration, RP-HPLC) | DPPH, ABTS, and hydroxyl radical-scavenging activities was higher in F4 fraction. | [69] |
| Brown rice | Bromelain | SPFWNINAHS, MPVDVIANAYR, VVYFDQTQAQA, AVYVYDVNNNANQ, YNILSGFDTEL, EFFDVSNELFQ | <1.5 kDa, (ultra, gel filtration, RP-HPLC) | F4 fraction of glutelin hydrolysates showed the highest ABTS radical scavenging and Cu2+ chelating activity. | [116] |
| Rice bran | Pepsin, Trypsin | N.R | 3–10 kDa, (ultra, gel filtration, RP-HPLC) | F2 sub-fraction displayed the highest DPPH, ABTS radical scavenging and Fe2+ chelating activity. | [117] |
| White and colored rice | Alcalase, Flavourzyme, Neutrase | Hydrolysate | <10 kDa | Alcalase hydrolysate of immature rice showed the highest DPPH and Fe2+ chelating activity compared to matured rice. | [118] |
| Rice | Pepsin-Pancreatin | Hydrolysate | N.R | Methionine-supplemented hydrolysates showed enhanced scavenging capacities for ABTS, superoxide, and nitric oxide. | [119] |
| Rice bran | pepsin-trypsin | N.R | <3 kDa, ultrafiltration | Higher ABTS radical scavenging and metal chelating activities in F1 fragment. | [120] |
| Rice | Alcalase, Flavourzyme | Hydrolysate | <3 kDa | Alcalase hydrolysate showed higher DPPH and FRAP activities and, hence, improved oxidative stability of linseed oil during storage. | [121] |
| Rice | Microbial proteases | Hydrolysate | 1–3 kDa, ultrafiltration | Validase and alkaline protease F3 fractions exerted remarkable DPPH and ABTS radical scavenging abilities. Validase-F3 | [122] |
| (Validase®, Neutral Protease, alkaline protease). | (500 µg/g) inhibited lipid oxidation by 19 and 15% at days 8 and 15 of storage, respectively. | ||||
| Rice residue protein | Alcalase, Flavourzyme, Protamex, Pepsin, Trypsin, Papain | RPNYTDA, TSQLLSDQ, TRTGDPFF, NFHPQ | <1 kDa, ultrafiltration, SEC, RP-FPLC | RRPB3 fraction showed the strongest DPPH, ABTS radical scavenging and FRAP-Fe3+ reducing activities. Synergistic activity was displayed by RRPB3 I and III. | [123] |
| Corn gluten meal | Neutrase | Hydrolysate | - | Hydrolysate (5 mg mL−1) inhibited lipid oxidation in emulsion by reducing hydroperoxides and TBARS formation. | [124] |
| Corn germ meal | Alcalase | MGGN, MNN, MEN | <1 kDa, (ultra, gel filtration) | F3 fraction from hydrolysate-5 displayed higher DPPH, ABTS radical scavenging, and ORAC activities. | [125] |
| Corn gluten meal and distillers’- dried grains with solubles (DDGS) | Neutrase, | Hydrolysate | - | Neutrase-hydrolyzed corn gluten meal (2%) displayed the strongest oxidation prevention in ground meat and significantly reduced TBARS in pig feed. Alcalase-hydrolyzed DDGS retarded the lipid oxidation by 37.8%. | [126] |
| Alcalase | |||||
| Corn protein isolate | Microbial protease (Validase, Alkaline, Neutral) | Hydrolysate | <1–10 kDa, (ultrafiltration) | Neutral protease fraction (NP-F3) demonstrated the highest ORAC peroxyl radicals scavenging activity. NP-F3 (250 and 500 µg/g) remarkably inhibited lipid oxidation by 52.9% in ground beef. | [127] |
| Corn gluten meal | Alcalase, Protamex | QQPQPW | <1 kDa (ultra, gel filtration and RP-HPLC). | Alcalase hydrolysate displayed the highest antioxidant activity. Hexapeptide showed remarkable DPPH, ABTS, and hydroxyl radical scavenging activity, but weaker Fe2+-chelating capacity. | [128] |
| Corn gluten meal | Alcalase, | YPQ, AYL | <1 kDa (ultra, gel filtration and RP-HPLC) | Alcalase hydrolysates and GI-resistant peptides exhibited higher DPPH, hydroxyl radical scavenging and reduced power activity. | [129] |
| Pepsin, pancreatin | |||||
| Corn gluten meal | Papain, Ficin, Bromelain | Hydrolysate | <1 kDa, >10 kDa (ultra, gel filtration) | Papain F4 fraction, ficin F1 fraction, and bromelain F3 fraction showed the strongest DPPH and ABTS radical scavenging and metal chelating activity. Papain F4 fraction (1000 mg/kg) reduced lipid oxidation in ground meat by 30.45%. | [130] |
| Corn gluten meal | Alcalase, Flavourzyme, Alcalase + Flavourzyme and Flavourzyme + Alcalase | CSQAPLA, YPKLAPN, YPQLLPNE | <1 kDa (ultra, gel filtration and RP-HPLC | Alcalase + Flavourzyme hydrolysate exhibited better antioxidant activities. CSQAPLA displayed the highest DPPH and superoxide anion radical scavenging capacities, and good reducing power activity. | [96] |
| Sorghum | Alcalase | VAITLTMK, VSKSVLVK | <1 kDa, (ultra, gel filtration, RP-UFLC) | These peptides displayed the highest DPPH radical scavenging activity. | [102] |
| Sorghum | Alcalase | QQWQ, QWQQ | <5–10 kDa, (ultra, gel filtration) | Medium-sized hydrolysate fraction exhibited stronger antioxidant activities based on DPPH, ABTS, metal chelating, and reducing power assay. The fraction also exerted remarkable lipid oxidation inhibition, both in emulsion and in the ground meat system. | [131] |
| Finger millet | Trypsin, pepsin | TSSSLNMAVRGGLTR, STTVGLGISMRSASVR | <3 kDa, (ultra, gel filtration and RP-UFLC) | Trypsin fraction GFB exhibited the highest DPPH and ABTS radical scavenging activity. In silico interaction of serine and threonine residues with free radicals potentially resulted in antioxidant activity. | [101] |
| Sorghum | Papain | LRQQ, QLQGV, WQPN, GLQDL, AMCGVV, YLRQ, TPCATS, QGVAAA, AQVAQ, QQLQ | 1–3 kDa (ultrafiltration and gel filtration) | Kafirin hydrolysates (1.0 mg/g) inhibited TBARS formation by 32.1% in meat samples. Fraction F3 also exhibited the highest % of DPPH radical scavenging activity, metal chelating and ORAC activity. | [132] |
| Sorghum | Alcalase, flavourzyme, neutrase, everlase, protamex, papain, ficin, bromelain, trypsin, Pepsin | Neutrase hydrolysate fractions | 3–10 kDa (ultra and gel filtration) | Neutrase F2 hydrolysates fraction displayed higher antioxidant activities (DPPH, ORAC, metal chelating) and retarded lipid autoxidation and peroxidation in the meat model. | [133] |
| Protein Source | Processing Method | Peptide Sequence/Size/Hydrolysate | Assay Outcome | Reference |
|---|---|---|---|---|
| Wheat germ protein hydrolysates | Electron beam irradiation (EBI) | <1 kDa | Irradiation at 50 kGy increased DPPH and ABTS+ radical scavenging activity by 45.77% and 52.52%, respectively. | [164] |
| Wheat germ | Fermentation with Bacillus subtilis | N.R | Peptide content increased in fermented samples compared to non-fermented samples and resulted in an increase in DPPH radical scavenging, Fe2+ chelating and Fe3+ reducing power activities. | [166] |
| KAMUT® Wheat | Combination of enzyme hydrolysis (Alcalase, Neutrase, Flavourzyme) and fermentation with (Lactobacillus spp. strains) | VLPPQQQY | Stronger superoxide anion, hydroxyl radicals, organic nitro-radicals (ABTS, DPPH) scavenging, and lipid peroxidation inhibition was observed. | [167] |
| Wheat germ | Subcritical water extraction | GPFGPE, FGE, <1 kDa | Peptide fraction 4 showed the strongest DPPH radical scavenging activity and could effectively cross Caco-2 intestinal epithelium cells. | [168] |
| Sorghum | Ultrasonication combined with pepsin-pancreatin hydrolysis | <1 kDa | Ultrasonication increased DH, DPPH scavenging activity and ORAC values. However, there was no improvement in NO scavenging activity. | [162] |
| Rice dregs | Angling method using metal-organic framework combined with alcalase hydrolysis. | GDMNP, LLLRW | Strong DPPH, superoxide anion, hydroxyl radical scavenging and Fe2+ chelating activity was exhibited by peptides. | [169] |
| Rice | Alcalase-assisted electron beam irradiation (EBI) | Hydrolysate | EBI treatment at 50 kGy improved DPPH (96.81%) and ABTS (92.04%) radical scavenging activity. | [165] |
| Corn gluten meal | Ultrasonication assisted alcalase hydrolysis | SGV, LPF, LLPH, LLPF, FLPF, AHL, LGV (<1 kDa) | Ultrasonic pretreatment (5 W/L, 2 s/2 s on/off, 50 °C, and 25 min) significantly increased DH, DPPH and hydroxyl radical scavenging activity and enhanced formation of small size peptides. | [170] |
| Protein Source | Enzyme | Peptide Sequence/Hydrolysate | Cellular Model | Cellular Outcome | Reference |
|---|---|---|---|---|---|
| Wheat germ | Trypsin, Alcalase | AREGETVVPG | Vascular smooth muscle cells | High glucose-induced cell growth and generation of intracellular ROS was significantly decreased by AOP (5 µM). Suppression of PKCζ, AKT and Erk1/2 phosphorylation, and inhibition of Nox4 protein expression by AOP (5 µM). | [182] |
| Wheat germ | neutral protease | ADWGGPLPH | Vascular smooth muscle cells | High glucose-induced cell proliferation and intracellular ROS generation was significantly reduced by peptide at 10 µM and 20 µM. Stimulation of AMPK activity, inhibition of PKCζ, AKT and Erk1/2 phosphorylation, and suppression of NOX4 protein expression. | [172] |
| Wheat gluten protein | Alcalase | Protein hydrolysate | Human peripheral blood mononuclear cells | Hydrolysate (0.5 mg/mL) directly scavenged free radicals, increased GSH levels, reduced NO overproduction, and, thus, enhanced cells’ antioxidant capacity. Also, cell proliferation, Th1 and Th17 pro-inflammatory cytokines IFN-γ and and IL-17 were reduced. | [178] |
| Foxtail millet | Alcalase | PFLF, IALLIPF | Human keratinocyte HaCaT cells | ROS, MDA production was effectively reduced and GSH levels increased by MPP (300 µg/mL) in H2O2-induced HaCaT cells. | [179] |
| Sorghum | Pepsin-pancreatin | Kafirin hydrolysate | THP-1 human macrophages | Kafirin (100 μg/mL) reduced LPS-induced intracellular ROS production, inflammatory cytokines (IL-1β, IL-6 and TNF-α) production, and nuclear translocation of p65 and c-JUN. | [176] |
| Rice bran | - | KHNRGDEF | Human umbilical vein endothelial cells (HUVECs) | H2O2-induced HUVECs oxidant injury was protected by rice bran peptide (0.1 mM) supplementation via TLR4 binding, pathway inhibition, and suppression of NF-κB activation. | [177] |
| Rice | - | OP60 commercial peptide | HepG2 cells | H2O2- or APAP-induced HepG2 cytotoxicity was reduced by 5 mg/mL OP60 pretreatment via glutathione homeostasis restoration and increased mRNA expression of antioxidant enzymes. | [175] |
| Corn | Alcalase | Zein hydrolysate/peptides | HepG2 cells | Hydrolysate showed higher ORAC activity than native proteins. Peptides (1155.56–1781.63 ng/mL IC50) induced apoptosis at 24 hr by increasing caspase 3 expression. | [183] |
| Corn germ meal | Alcalase | MGGN, MNN, MEN | HepG2 cells | Peptides (0.2 mM) significantly reduced ROS generation in H2O2-induced HepG2 cells. MNN showed the highest cellular antioxidant activity. | [125] |
| Corn gluten meal | Alcalase | corn gluten hydrolysate (CGH1) <1 kDa | HepG2 cells | CGH1-pretreated cells at 2.5 mg/mL upregulated the genes GPX3, GPX5, SOD3, CYGB, SEPP1, and MT3 involved in antioxidant defense. CGH1 suppressed EPHX2 expression, increased cellular EETs, EET-phospholipids formation, and, thus, protected against H2O2-induced HepG2 cell damage. | [180] |
| Corn gluten meal | Alcalase | GLLLPH | HepG2 cells | Corn peptide fractions (CPF) at 2.50 mg/mL exhibited high cellular antioxidant activities and increased the levels of intracellular antioxidant enzymes (SOD, CAT, GR and GSH) in oxidized HepG2 cells. | [73] |
| Protein Source | Peptide/Hydrolysate | Purpose of Study | In Vivo Outcome | BSA-Based Human Equivalent Dose (mg/kg) | Reference |
|---|---|---|---|---|---|
| Wheat bran | NL, QL, FL, HAL, AAVL, AKTVF, and TPLTR | Investigated the blood pressure lowering effects of wheat protein hydrolysates and peptides in spontaneous hypertensive rats (SHR). | Oral administration of <1 kDa peptides at 100 mg/kg showed a better reduction of systolic blood pressure (−35 mmHg) after 6 h compared to hydrolysate (−20 mmHg). | 16.22 | [108] |
| Wheat bran | LRP, LQP | Evaluated the effects of peptides on oxidative stress and the AMPK signaling pathway in HFD-induced NASH C57BL/6 mice. | NASH mice treated with 0.20% LRP showed a remarkable decrease in serum d-ROM and a significant increase in BAP levels. Administration of 0.20% LQP increased phospho-AMPK expression and decreased phospho-ACC expression, thus alleviating the severity of NASH. | 7.46 | [184] |
| Wheat bran | ADWGGPLPH | Assessed the antioxidative and antidiabetic vascular dysfunction effects of peptide in STZ-induced C57BL/6 mice. | Administered peptide at 4mg/kg enhanced SOD expression and total antioxidant capacity, and also attenuated hyperglycemia-induced inflammatory factors, such as TNF-α and IL-1β. | 0.32 | [172] |
| Rice | OP60 commercial peptide | Evaluated the protective effect of OP60 against APAP-induced hepatic injury in mice. | GSH synthesis and antioxidant enzymes were induced by OP60 (500 mg/kg) via activation of the Nrf2 pathway. | 40.54 | [175] |
| Corn Gluten meal (CGM) | CGM peptides (<10 kDa) | Investigated the antioxidant capacity of CGMP produced by solid–state fermentation with Bacillus subtilis MTCC5480 in aging rats induced with D-galactose. | Serum and liver antioxidant enzymes (SOD, catalase, glutathione peroxidase) and total antioxidant capacity activities increased, with a decrease in MDA in D-galactose-induced aging rats fed CGMP (250 mg/kg bw). | 40.54 | [185] |
| Corn germ meal | Albumin peptides fractions (APF-4) | Examined the hepatoprotective effects of APF-4 in alcohol-induced liver injury in mice. | APF-4 (800 mg/kg bw) administration significantly reduced activities and levels of hepatic (AST), ALT and MDA, and increased activities of SOD, CAT and GSH. | 64.86 | [186] |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Ofosu, F.K.; Mensah, D.-J.F.; Daliri, E.B.-M.; Oh, D.-H. Exploring Molecular Insights of Cereal Peptidic Antioxidants in Metabolic Syndrome Prevention. Antioxidants 2021, 10, 518. https://doi.org/10.3390/antiox10040518
Ofosu FK, Mensah D-JF, Daliri EB-M, Oh D-H. Exploring Molecular Insights of Cereal Peptidic Antioxidants in Metabolic Syndrome Prevention. Antioxidants. 2021; 10(4):518. https://doi.org/10.3390/antiox10040518
Chicago/Turabian StyleOfosu, Fred Kwame, Dylis-Judith Fafa Mensah, Eric Banan-Mwine Daliri, and Deog-Hwan Oh. 2021. "Exploring Molecular Insights of Cereal Peptidic Antioxidants in Metabolic Syndrome Prevention" Antioxidants 10, no. 4: 518. https://doi.org/10.3390/antiox10040518
APA StyleOfosu, F. K., Mensah, D.-J. F., Daliri, E. B.-M., & Oh, D.-H. (2021). Exploring Molecular Insights of Cereal Peptidic Antioxidants in Metabolic Syndrome Prevention. Antioxidants, 10(4), 518. https://doi.org/10.3390/antiox10040518