
Antioxidant and Anti-Inflammaging Ability of Prune (Prunus Spinosa L.) Extract Result in Improved Wound Healing Efficacy

 , , ,
, , ,  , ,
, ,  ,
,  ,
, 

Abstract
1. Introduction
2. Materials and Methods
2.1. Chemicals and Reagents
2.2. Plant Material and Extract Preparation
2.3. Determination of Phenolic Compounds in Prunus Extract by HPLC/MS Analysis
2.4. Cell Culture and Treatments
2.5. In Vitro Wound Healing Assay
2.6. HUVEC Replicative Senescence Analysis
2.7. Quantitative Real Time PCR (RT-qPCR) of Mature MicroRNAs and mRNA
2.8. DPPH Radical Assay
2.9. C. elegans Cultivation
2.10. E. coli Strains and Growth
2.11. Prunus Extract Treatment in C. elegans
2.12. Life and Healthspan
2.13. Pharyngeal Pumping Rate and Body Bends Assay
2.14. Statistical Analysis
3. Results
3.1. Chemical Characterization

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Peak No | tR (min) | λ max (nm) | M+ or [M+Na]+ (m/z) | [M-H]− or [M+Na]− (m/z) | HPLC-ESI/MSn m/z (% base Peak) | Tentative Assignment | Ref. |
|---|---|---|---|---|---|---|---|
| 1 | 15.2 | 300sh, 325 | 355 a | 353 | O-Caffeoylquinic acid isomer | [1] | |
| 2 | 18.5 | 281, 514 | 595 | MS2[595]: 287 (100), 449 (65) | Cyanidin-3-O-rutinoside | [25,26,27] | |
| 3 | 19.4 | 282, 517 | 609 | MS2[609]: 301 (50), 463 (40) | Peonidin-3-O-rutinoside | [25,26,27] | |
| 4 | 25.9 | 260, 352 | 611 a | 609 | MS2[611]: 303 (100), 465 (30) | Rutin (quercetin-3-O-rutinoside) | [1] |
| 5 | 26.5 | 258,347 | 597 a | 595 | MS2[597]: 303 (100), 465 a(50) | Quercetin 3-O-hexoside-O-pentoside | [1] |
| 6 | 27.4 | 258, 350 | 465 a | 463 | MS2[463]: 301 20) MS2[465]: 303 (100) | Quercetin glucoside or galactoside | [1] |
| 7 | 29.0 | 275, 350 | 625 a | 623 a | MS2[623]: 433 (100), MS2[625]: 303 (100), 435 a (20) | Quercetin xyloside or arabinoside derivative | [1] |
| 8 | 29.8 | 260, 352 | 435 a | 433 | MS2[435]: 303 (100) | Quercetin arabinoside or xyloside | [1] |
| 9 | 30.6 | 278, 352 | 433 | MS2[435]: 303 (100) | Quercetin pentoside | [28] | |
| 10 | 31.4 | 278, 352 | 447 | MS2[449]: 303 (100) | Quercetin-3-O-rhamnoside | [1] |
3.2. In Vitro Analyses
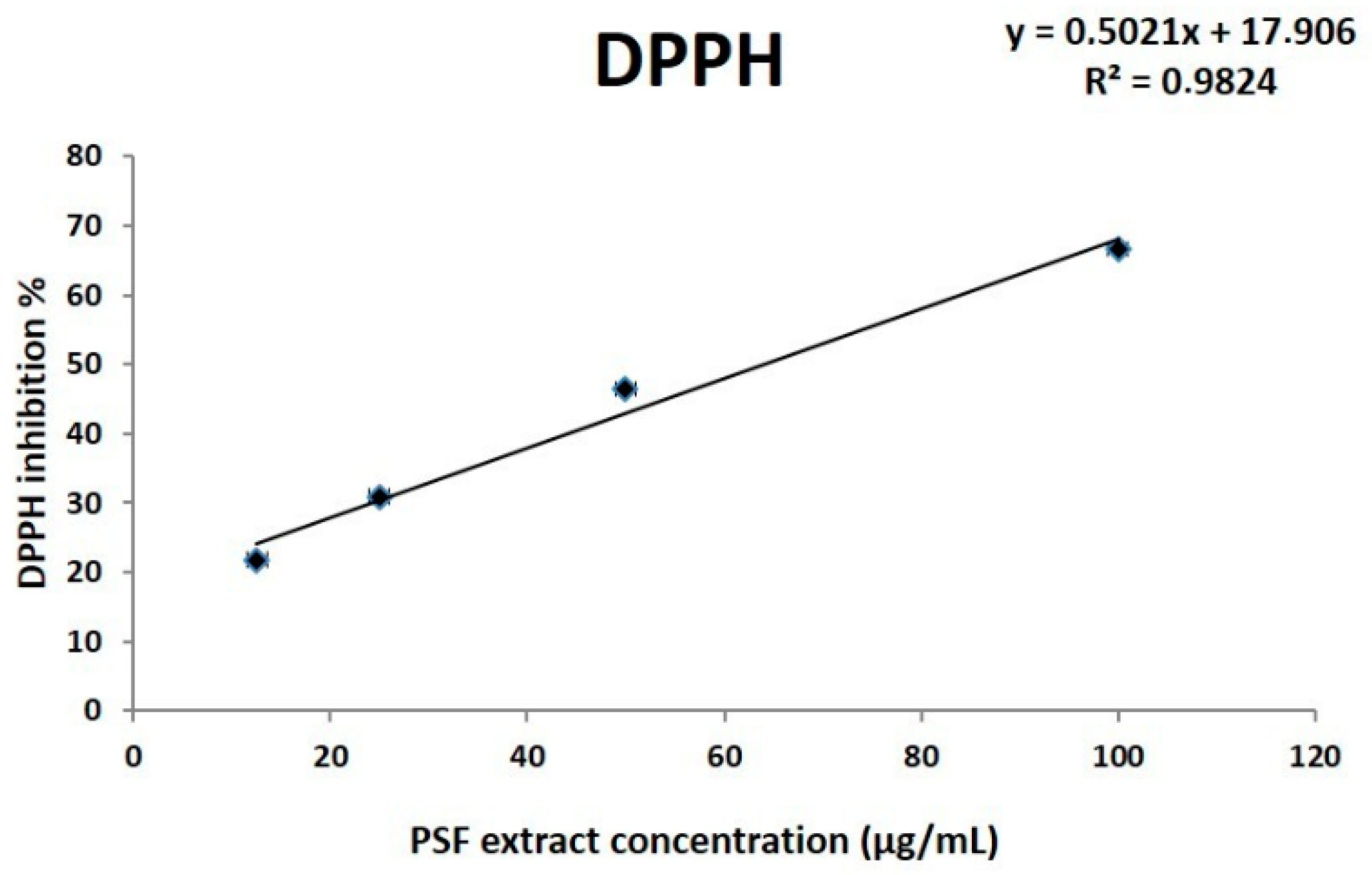
3.2.1. In Vitro Evaluation of Antioxidant Properties
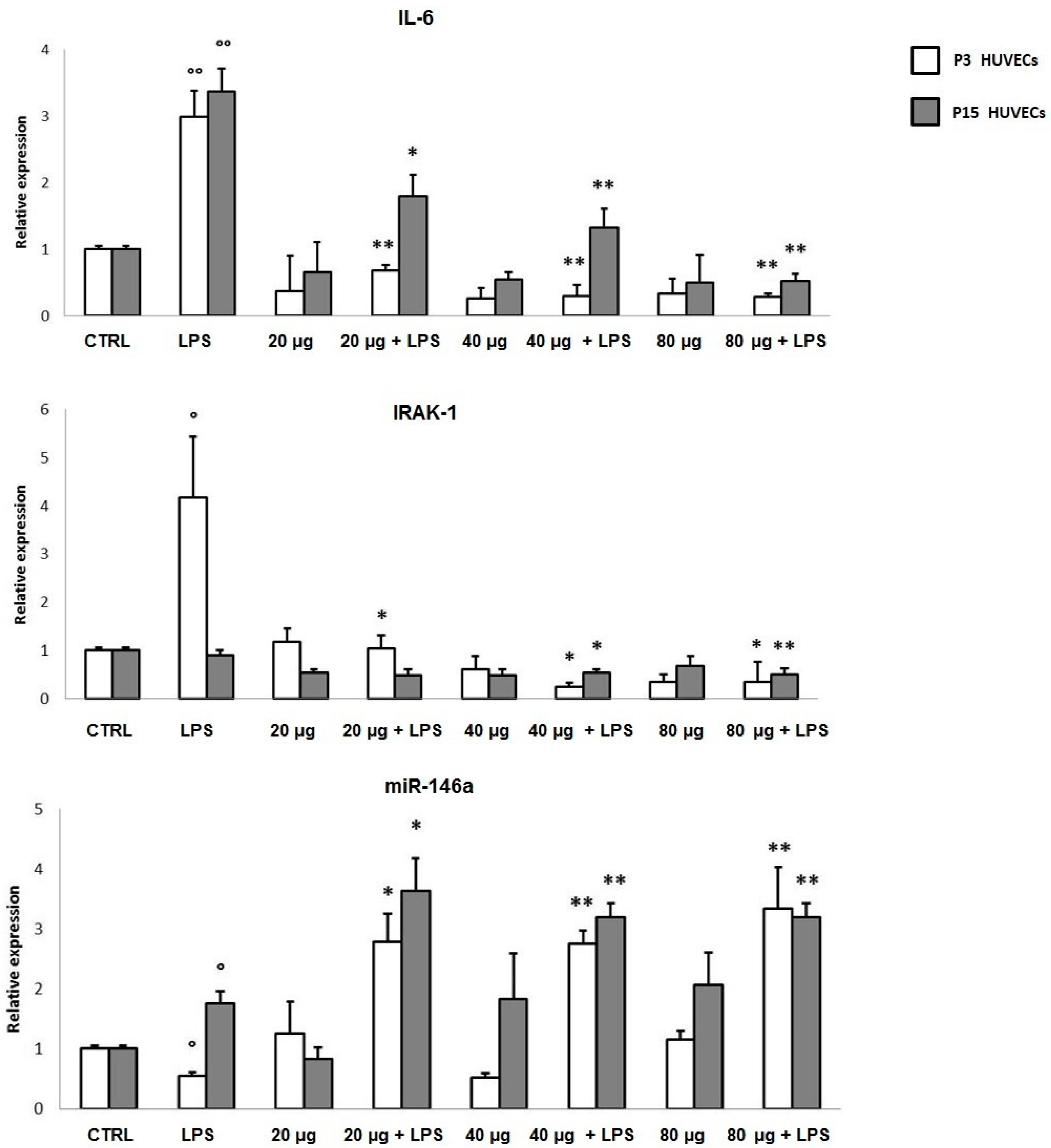
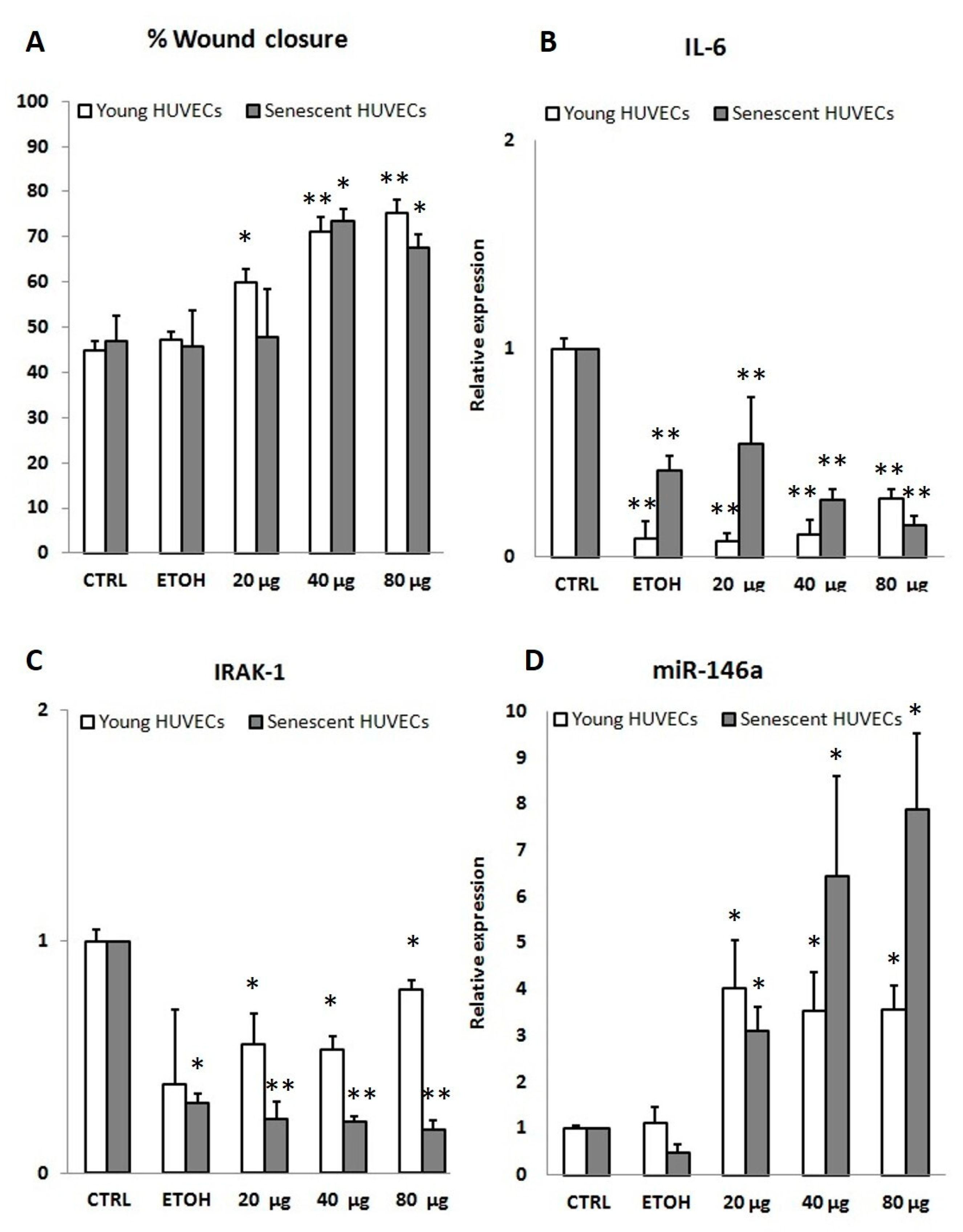
3.2.2. In Vitro Anti-inflammatory Activity
3.3. In Vivo Analyses
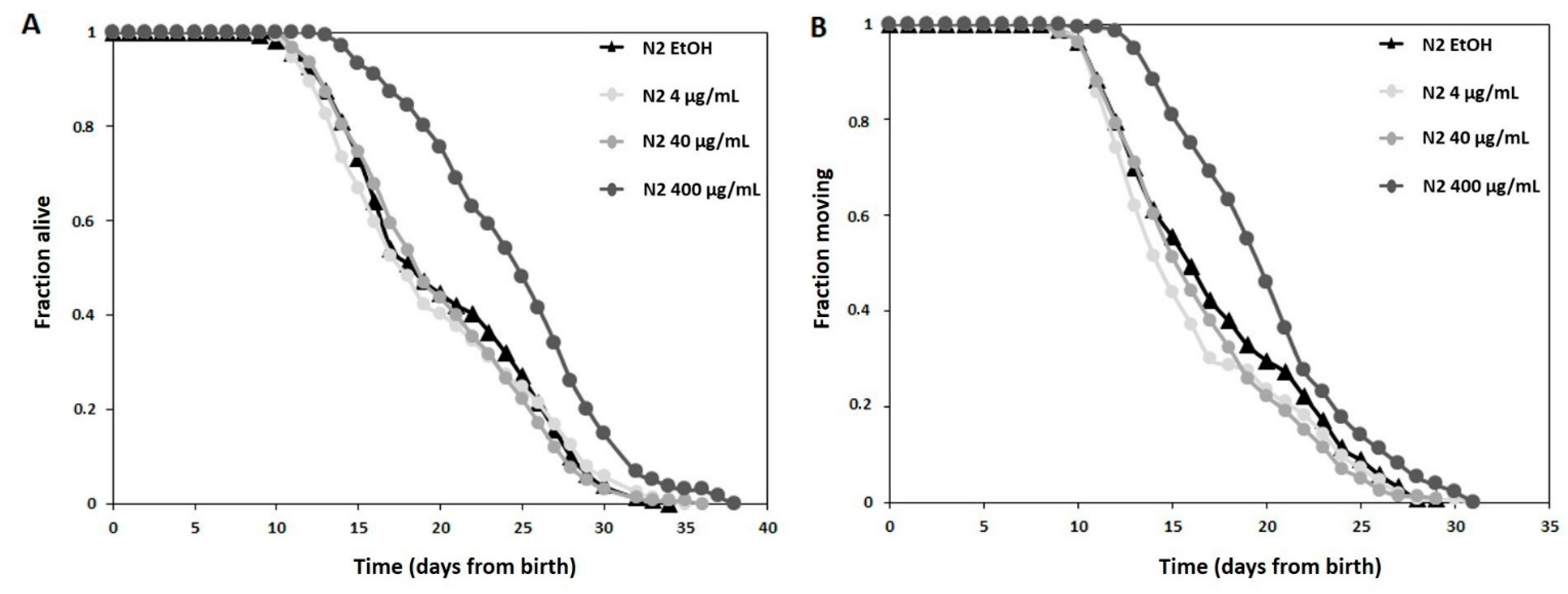
3.3.1. Longevity Study
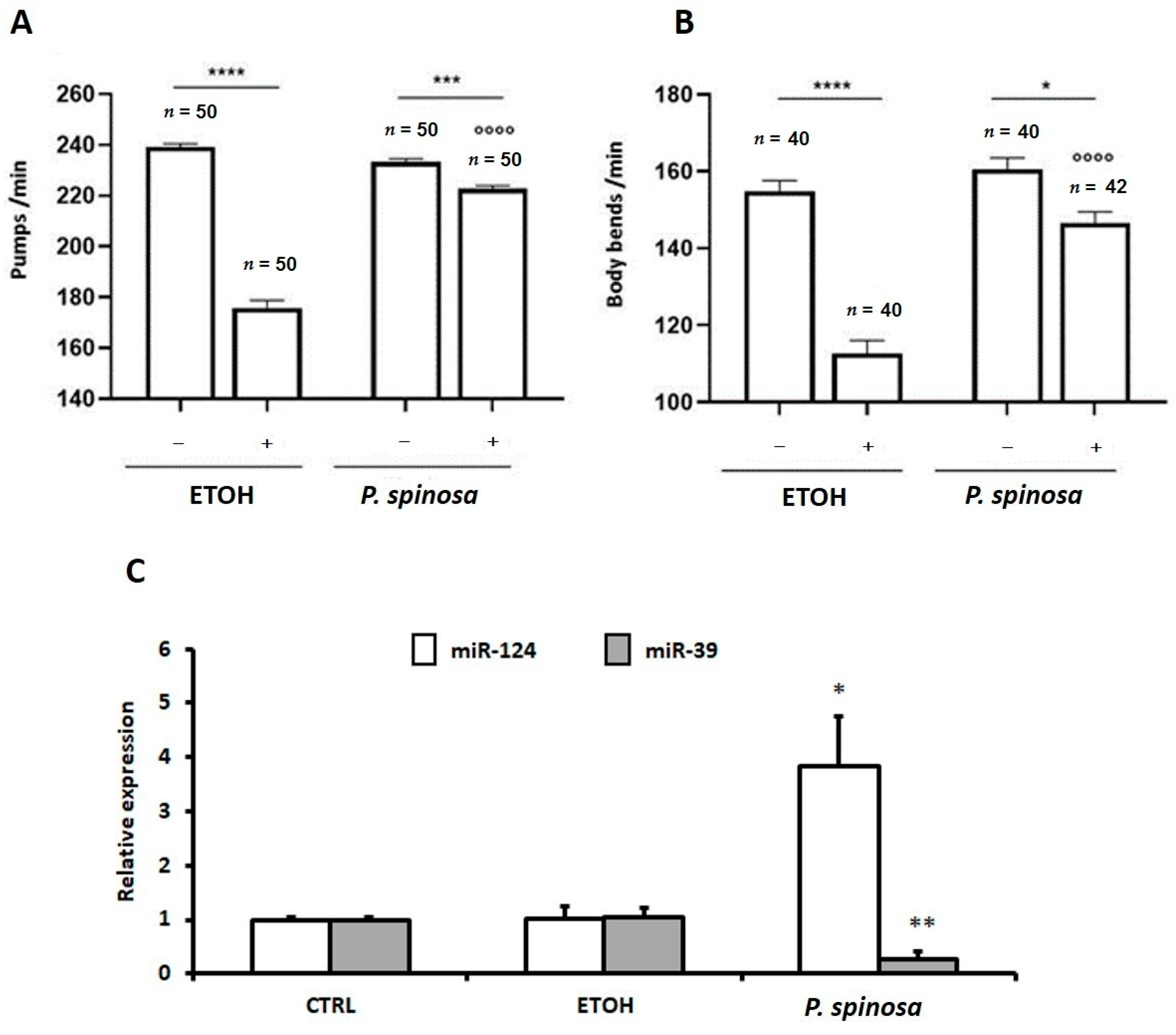
3.3.2. In Vivo Evaluation of Antioxidant Properties
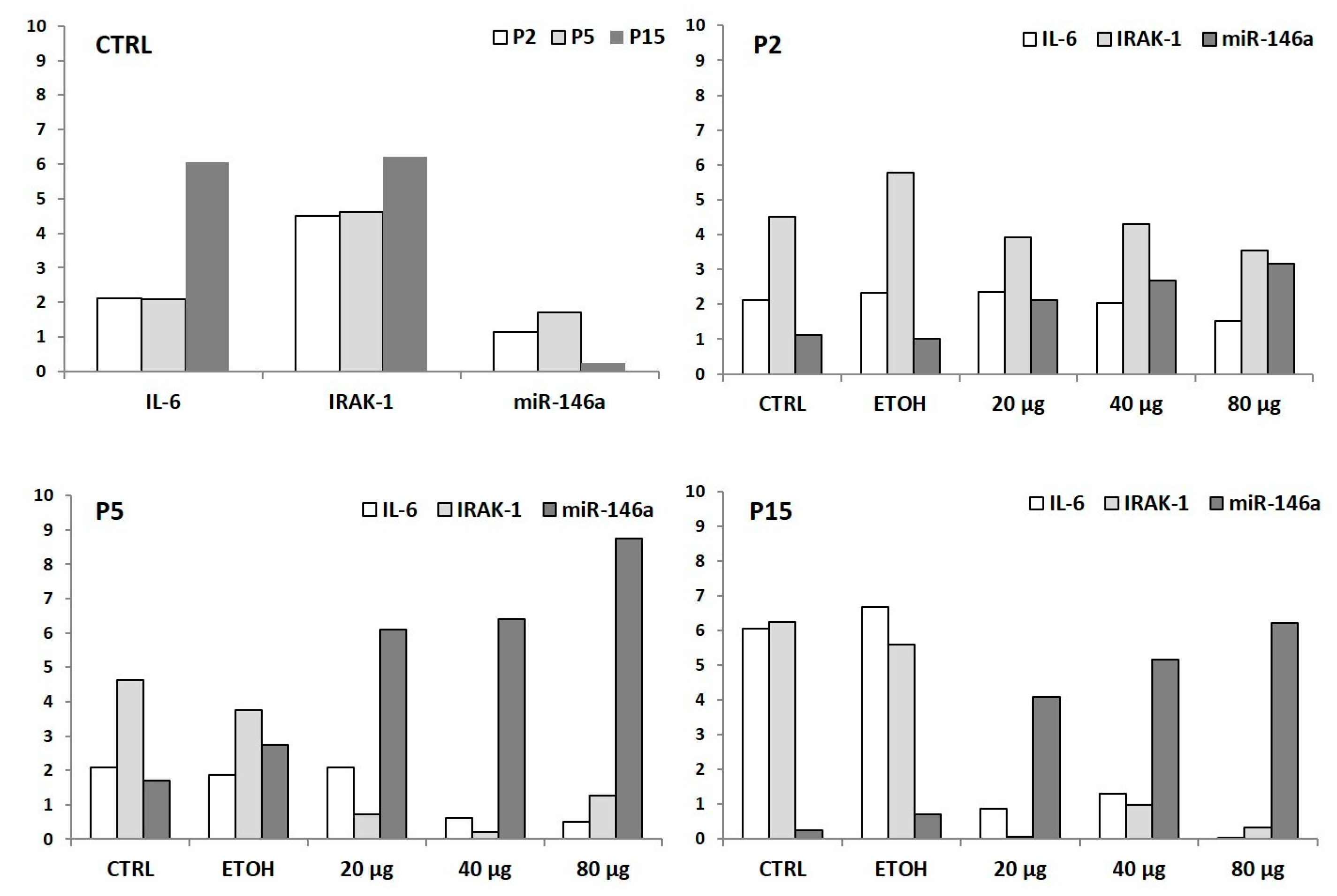
3.3.3. In Vivo MicroRNAs Modulation Associated to Prunus Antioxidant Activity
3.4. PSF Extract Effect on Wound Healing
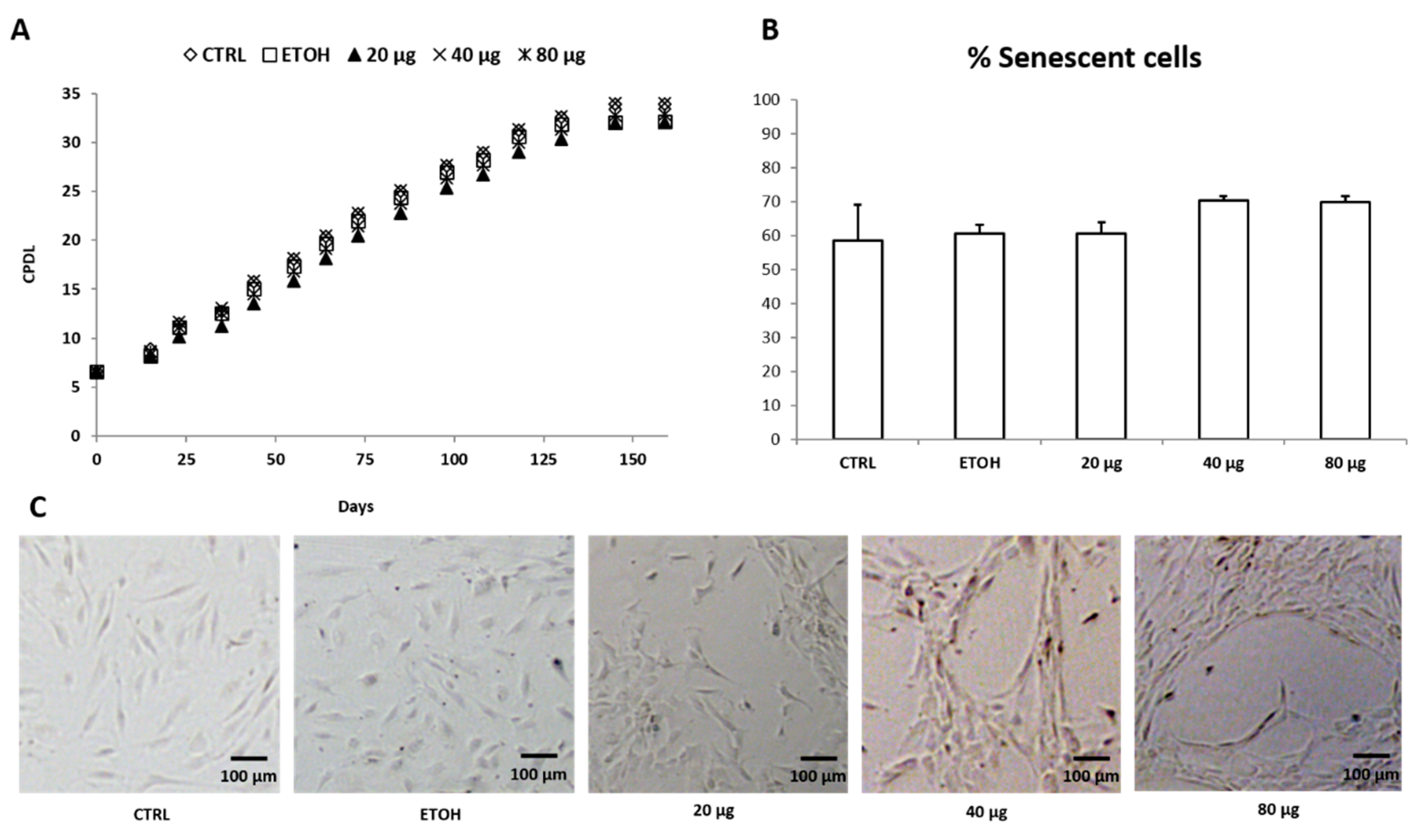
3.5. HUVEC Replicative Senescence Analysis and PSF Effect on Cell Biological Aging
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Tiboni, M.; Coppari, S.; Casettari, L.; Guescini, M.; Colomba, M.; Fraternale, D.; Gorassini, A.; Verardo, G.; Ramakrishna, S.; Guidi, L.; et al. Prunus spinosa Extract Loaded in Biomimetic Nanoparticles Evokes In Vitro Anti-Inflammatory and Wound Healing Activities. Nanomaterials 2020, 11, 36. [Google Scholar] [CrossRef]
- Franceschi, C.; Bonafè, M.; Valensin, S.; Olivieri, F.; De Luca, M.; Ottaviani, E.; De Benedictis, G. Inflamm-aging. An evolutionary perspective on immunosenescence. Ann. N. Y. Acad. Sci. 2000, 908, 244–254. [Google Scholar] [CrossRef] [PubMed]
- Vasa-Nicotera, M.; Chen, H.; Tucci, P.; Yang, A.L.; Saintigny, G.; Menghini, R.; Mahè, C.; Agostini, M.; Knight, R.A.; Melino, G.; et al. MiR-146a is modulated in human endothelial cell with aging. Atherosclerosis 2011, 217, 326–330. [Google Scholar] [CrossRef]
- Olivieri, F.; Lazzarini, R.; Recchioni, R.; Marcheselli, F.; Rippo, M.R.; Di Nuzzo, S.; Albertini, M.C.; Graciotti, L.; Babini, L.; Mariotti, S.; et al. MiR-146a as marker of senescence-associated pro-inflammatory status in cells involved in vascular remodelling. Age (Omaha) 2013, 35, 1157–1172. [Google Scholar] [CrossRef] [PubMed]
- Nour, V.; Trandafir, I.; Cosmulescu, S. Bioactive compounds, antioxidant activity and nutritional quality of different culinary aromatic herbs. Not. Bot. Horti Agrobot. Cluj-Napoca 2017, 45, 179–184. [Google Scholar] [CrossRef]
- Fraternale, D.; Giamperi, L.; Bucchini, A.; Sestili, P.; Paolillo, M.; Ricci, D. Prunus spinosa fresh fruit juice: Antioxidant activity in cell-free and cellular systems. Nat. Prod. Commun. 2009, 4, 1665–1670. [Google Scholar] [CrossRef] [PubMed]
- Ganhão, R.; Estévez, M.; Kylli, P.; Heinonen, M.; Morcuende, D. Characterization of selected wild mediterranean fruits and comparative efficacy as inhibitors of oxidative reactions in emulsified raw pork burger patties. J. Agric. Food Chem. 2010, 58, 8854–8861. [Google Scholar] [CrossRef]
- Radovanović, B.C.; Andelković, A.S.M.; Radovanović, A.B.; Andelković, M.Z. Antioxidant and antimicrobial activity of polyphenol extracts from wild berry fruits grown in Southeast Serbia. Trop. J. Pharm. Res. 2013, 12, 813–819. [Google Scholar] [CrossRef]
- Karabegović, I.T.; Stojičević, S.S.; Veličković, D.T.; Todorović, Z.B.; Nikolić, N.Č.; Lazić, M.L. The effect of different extraction techniques on the composition and antioxidant activity of cherry laurel (Prunus laurocerasus) leaf and fruit extracts. Ind. Crops Prod. 2014, 54, 142–148. [Google Scholar] [CrossRef]
- Veličković, J.M.; Kostić, D.A.; Stojanović, G.S.; Mitić, S.S.; Mitić, M.N.; Ranđelović, S.S.; Đorđević, A.S. Phenolic composition, antioxidant and antimicrobial activity of the extracts from Prunus spinosa L. fruit. Hem. Ind. 2014, 68, 297–303. [Google Scholar] [CrossRef]
- Walraven, M.; Hinz, B. Therapeutic approaches to control tissue repair and fibrosis: Extracellular matrix as a game changer. Matrix Biol. 2018, 71–72, 205–224. [Google Scholar] [CrossRef]
- Albertini, M.C.; Fraternale, D.; Semprucci, F.; Cecchini, S.; Colomba, M.; Rocchi, M.B.L.; Sisti, D.; Di Giacomo, B.; Mari, M.; Sabatini, L.; et al. Bioeffects of Prunus spinosa L. Fruit ethanol extract on reproduction and phenotypic plasticity of Trichoplax adhaerens Schulze, 1883 (Placozoa). PeerJ 2019, 7. [Google Scholar] [CrossRef]
- Sabatini, L.; Fraternale, D.; Di Giacomo, B.; Mari, M.; Albertini, M.C.; Gordillo, B.; Rocchi, M.B.L.; Sisti, D.; Coppari, S.; Semprucci, F.; et al. Chemical composition, antioxidant, antimicrobial and anti-inflammatory activity of Prunus spinosa L. fruit ethanol extract. J. Funct. Foods 2020, 67, 103885. [Google Scholar] [CrossRef]
- Neveu, V.; Perez-Jiménez, J.; Vos, F.; Crespy, V.; du Chaffaut, L.; Mennen, L.; Knox, C.; Eisner, R.; Cruz, J.; Wishart, D.; et al. Phenol-Explorer: An online comprehensive database on polyphenol contents in foods. Database (Oxford) 2010, 2010. [Google Scholar] [CrossRef] [PubMed]
- Prattichizzo, F.; Giuliani, A.; Recchioni, R.; Bonafè, M.; Marcheselli, F.; De Carolis, S.; Campanati, A.; Giuliodori, K.; Rippo, M.R.; Brugè, F.; et al. Anti-TNF-α treatment modulates SASP and SASP-related microRNAs in endothelial cells and in circulating angiogenic cells. Oncotarget 2016, 7, 11945–11958. [Google Scholar] [CrossRef] [PubMed]
- Angel-Morales, G.; Noratto, G.; Mertens-Talcott, S.U. Standardized curcuminoid extract (Curcuma longa L.) decreases gene expression related to inflammation and interacts with associated microRNAs in human umbilical vein endothelial cells (HUVEC). Food Funct. 2012, 3, 1286–1293. [Google Scholar] [CrossRef] [PubMed]
- Fraternale, D.; Giamperi, L.; Bucchini, A.; Ricci, D.; Epifano, F.; Burini, G.; Curini, M. Chemical Composition, Antifungal and In Vitro Antioxidant Properties of Monarda didyma L. Essential Oil. J. Essent. Oil Res. J. Essent. Oil Res 2006, 18, 581–585. [Google Scholar] [CrossRef]
- Kamath, R.S.; Ahringer, J. Genome-wide RNAi screening in Caenorhabditis elegans. Methods 2003, 30, 313–321. [Google Scholar] [CrossRef]
- Han, S.K.; Lee, D.; Lee, H.; Kim, D.; Son, H.G.; Yang, J.S.; Lee, S.J.V.; Kim, S. OASIS 2: Online application for survival analysis 2 with features for the analysis of maximal lifespan and healthspan in aging research. Oncotarget 2016, 7, 56147–56152. [Google Scholar] [CrossRef] [PubMed]
- Schiavi, A.; Maglioni, S.; Palikaras, K.; Shaik, A.; Strappazzon, F.; Brinkmann, V.; Torgovnick, A.; Castelein, N.; De Henau, S.; Braeckman, B.P.; et al. Iron-Starvation-Induced Mitophagy Mediates Lifespan Extension upon Mitochondrial Stress in C. elegans. Curr. Biol. 2015, 25, 1810–1822. [Google Scholar] [CrossRef]
- Shaham, S. Methods in cell biology. WormBook 2006. [Google Scholar] [CrossRef]
- Diomede, L.; Romeo, M.; Rognoni, P.; Beeg, M.; Foray, C.; Ghibaudi, E.; Palladini, G.; Cherny, R.A.; Verga, L.; Capello, G.L.; et al. Cardiac Light Chain Amyloidosis: The Role of Metal Ions in Oxidative Stress and Mitochondrial Damage. Antioxid. Redox Signal. 2017, 27, 567–582. [Google Scholar] [CrossRef]
- Diomede, L.; Rognoni, P.; Lavatelli, F.; Romeo, M.; Del Favero, E.; Cantù, L.; Ghibaudi, E.; Di Fonzo, A.; Corbelli, A.; Fiordaliso, F.; et al. A Caenorhabditis elegans-based assay recognizes immunoglobulin light chains causing heart amyloidosis. Blood 2014, 123, 3543–3552. [Google Scholar] [CrossRef] [PubMed]
- Diomede, L.; Soria, C.; Romeo, M.; Giorgetti, S.; Marchese, L.; Mangione, P.P.; Porcari, R.; Zorzoli, I.; Salmona, M.; Bellotti, V.; et al. C. elegans Expressing Human β2-Microglobulin: A Novel Model for Studying the Relationship between the Molecular Assembly and the Toxic Phenotype. PLoS ONE 2012, 7, e52314. [Google Scholar] [CrossRef]
- Deineka, V.I.; Deineka, L.A.; Sirotin, A.A. Anthocyans from fruit of certain plants of the genus Prunus. Chem. Nat. Compd. 2005, 41, 230–231. [Google Scholar] [CrossRef]
- Ruiz, A.; Hermosín-Gutiérrez, I.; Vergara, C.; von Baer, D.; Zapata, M.; Hitschfeld, A.; Obando, L.; Mardones, C. Anthocyanin profiles in south Patagonian wild berries by HPLC-DAD-ESI-MS/MS. Food Res. Int. 2013, 51, 706–713. [Google Scholar] [CrossRef]
- Wu, X.; Prior, R.L. Systematic identification and characterization of anthocyanins by HPLC-ESI-MS/MS in common foods in the United States: Fruits and berries. J. Agric. Food Chem. 2005, 53, 2589–2599. [Google Scholar] [CrossRef]
- Zhuang, B.; Bi, Z.M.; Wang, Z.Y.; Duan, L.; Lai, C.J.S.; Liu, E.H. Chemical profiling and quantitation of bioactive compounds in Platycladi Cacumen by UPLC-Q-TOF-MS/MS and UPLC-DAD. J. Pharm. Biomed. Anal. 2018, 154, 207–215. [Google Scholar] [CrossRef] [PubMed]
- Torgovnick, A.; Schiavi, A.; Maglioni, S.; Ventura, N. Gesundes Altern: Was können wir von Caenorhabditis elegans lernen? Z. Gerontol. Geriatr. 2013, 46, 623–628. [Google Scholar] [CrossRef]
- Maglioni, S.; Arsalan, N.; Ventura, N. C. elegans screening strategies to identify pro-longevity interventions. Mech. Ageing Dev. 2016, 157, 60–69. [Google Scholar] [CrossRef] [PubMed]
- Wilson, M.A.; Shukitt-Hale, B.; Kalt, W.; Ingram, D.K.; Joseph, J.A.; Wolkow, C.A. Blueberry polyphenols increase lifespan and thermotolerance in Caenorhabditis elegans. Aging Cell 2006, 5, 59–68. [Google Scholar] [CrossRef]
- Wang, N.; Liu, J.; Xie, F.; Gao, X.; Ye, J.H.; Sun, L.Y.; Wei, R.; Ai, J. miR-124/ATF-6, a novel lifespan extension pathway of Astragalus polysaccharide in Caenorhabditis elegans. J. Cell. Biochem. 2015, 116, 242–251. [Google Scholar] [CrossRef]
- Dallaire, A.; Garand, C.; Paquet, E.R.; Mitchell, S.J.; de Cabo, R.D.; Simard, M.J.; Lebel, M. Down regulation of miR-124 in both Werner syndrome DNA helicase mutant mice and mutant Caenorhabditis elegans wrn-1 reveals the importance of this microRNA in accelerated aging. Aging (Albany NY) 2012, 4, 636–647. [Google Scholar] [CrossRef]
- Qu, M.; Luo, L.; Yang, Y.; Kong, Y.; Wang, D. Nanopolystyrene-induced microRNAs response in Caenorhabditis elegans after long-term and lose-dose exposure. Sci. Total Environ. 2019, 697, 134131. [Google Scholar] [CrossRef] [PubMed]
- Hayflick, L. The limited in vitro lifetime of human diploid cell strains. Exp. Cell Res. 1965, 37, 614–636. [Google Scholar] [CrossRef]
- Campisi, J. Cellular senescence as a tumor-suppressor mechanism. Trends Cell Biol. 2001, 11, S27–S31. [Google Scholar] [CrossRef]
- Demaria, M.; Ohtani, N.; Youssef, S.A.; Rodier, F.; Toussaint, W.; Mitchell, J.R.; Laberge, R.M.; Vijg, J.; VanSteeg, H.; Dollé, M.E.T.; et al. An essential role for senescent cells in optimal wound healing through secretion of PDGF-AA. Dev. Cell 2014, 31, 722–733. [Google Scholar] [CrossRef]
- Storer, M.; Mas, A.; Robert-Moreno, A.; Pecoraro, M.; Ortells, M.C.; Di Giacomo, V.; Yosef, R.; Pilpel, N.; Krizhanovsky, V.; Sharpe, J.; et al. Senescence is a developmental mechanism that contributes to embryonic growth and patterning. Cell 2013, 155, 1119. [Google Scholar] [CrossRef]
- Boccardi, V.; Pelini, L.; Ercolani, S.; Ruggiero, C.; Mecocci, P. From cellular senescence to Alzheimer’s disease: The role of telomere shortening. Ageing Res. Rev. 2015, 22, 1–8. [Google Scholar] [CrossRef] [PubMed]
- Chinta, S.J.; Lieu, C.A.; DeMaria, M.; Laberge, R.-M.; Campisi, J.; Andersen, J.K. Environmental stress, ageing and glial cell senescence: A novel mechanistic link to Parkinson’s disease? J. Intern. Med. 2013, 273, 429–436. [Google Scholar] [CrossRef] [PubMed]
- Kostyuk, V.A.; Potapovich, A.I.; Suhan, T.O.; De Luca, C.; Korkina, L.G. Antioxidant and signal modulation properties of plant polyphenols in controlling vascular inflammation. Eur. J. Pharmacol. 2011, 658, 248–256. [Google Scholar] [CrossRef]
- Benavente-García, O.; Castillo, J. Update on uses and properties of citrus flavonoids: New findings in anticancer, cardiovascular, and anti-inflammatory activity. J. Agric. Food Chem. 2008, 56, 6185–6205. [Google Scholar] [CrossRef] [PubMed]
- Carmona-Gutierrez, D.; Zimmermann, A.; Kainz, K.; Pietrocola, F.; Chen, G.; Maglioni, S.; Schiavi, A.; Nah, J.; Mertel, S.; Beuschel, C.B.; et al. The flavonoid 4,4′-dimethoxychalcone promotes autophagy-dependent longevity across species. Nat. Commun. 2019, 10. [Google Scholar] [CrossRef] [PubMed]
- Voinnet, O. MicroRNA and autophagy - C. elegans joins the crew. EMBO Rep. 2013, 14, 485–487. [Google Scholar] [CrossRef] [PubMed]
- Zhu, H.-Y.; Bai, W.-D.; Liu, J.-Q.; Zheng, Z.; Guan, H.; Zhou, Q.; Su, L.-L.; Xie, S.-T.; Wang, Y.-C.; Li, J.; et al. Up-regulation of FGFBP1 signaling contributes to miR-146a-induced angiogenesis in human umbilical vein endothelial cells. Sci. Rep. 2016. [Google Scholar] [CrossRef]
- Buachan, P.; Chularojmontri, L.; Wattanapitayakul, S. Selected activities of Citrus maxima merr. fruits on human endothelial cells: Enhancing cell migration and delaying cellular aging. Nutrients 2014, 6, 1618–1634. [Google Scholar] [CrossRef] [PubMed]
- Han, K.T.; Kim, D.W.; Kim, S.J.; Kim, S.J. Biological age is associated with the active use of nutrition data. Int. J. Environ. Res. Public Health 2018, 15, 2431. [Google Scholar] [CrossRef]








Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Coppari, S.; Colomba, M.; Fraternale, D.; Brinkmann, V.; Romeo, M.; Rocchi, M.B.L.; Di Giacomo, B.; Mari, M.; Guidi, L.; Ramakrishna, S.; et al. Antioxidant and Anti-Inflammaging Ability of Prune (Prunus Spinosa L.) Extract Result in Improved Wound Healing Efficacy. Antioxidants 2021, 10, 374. https://doi.org/10.3390/antiox10030374
Coppari S, Colomba M, Fraternale D, Brinkmann V, Romeo M, Rocchi MBL, Di Giacomo B, Mari M, Guidi L, Ramakrishna S, et al. Antioxidant and Anti-Inflammaging Ability of Prune (Prunus Spinosa L.) Extract Result in Improved Wound Healing Efficacy. Antioxidants. 2021; 10(3):374. https://doi.org/10.3390/antiox10030374
Chicago/Turabian StyleCoppari, Sofia, Mariastella Colomba, Daniele Fraternale, Vanessa Brinkmann, Margherita Romeo, Marco Bruno Luigi Rocchi, Barbara Di Giacomo, Michele Mari, Loretta Guidi, Seeram Ramakrishna, and et al. 2021. "Antioxidant and Anti-Inflammaging Ability of Prune (Prunus Spinosa L.) Extract Result in Improved Wound Healing Efficacy" Antioxidants 10, no. 3: 374. https://doi.org/10.3390/antiox10030374
APA StyleCoppari, S., Colomba, M., Fraternale, D., Brinkmann, V., Romeo, M., Rocchi, M. B. L., Di Giacomo, B., Mari, M., Guidi, L., Ramakrishna, S., Ventura, N., & Albertini, M. C. (2021). Antioxidant and Anti-Inflammaging Ability of Prune (Prunus Spinosa L.) Extract Result in Improved Wound Healing Efficacy. Antioxidants, 10(3), 374. https://doi.org/10.3390/antiox10030374