
Planarians as an In Vivo Experimental Model for the Study of New Radioprotective Substances



{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Materials and Methods
2.1. Animals
2.2. Computer-Assisted Morphometry In Vivo
2.3. Whole-Mount Immunocytochemical Study of Planarian Stem Cell Mitotic Activity
2.4. Experimental Testing Substance
2.5. Planarian X-ray Irradiation
2.6. RAPD PCR for Genotoxicity Analysis
2.7. RT-PCR for Gene Expression Analysis
2.8. ROS Measurement in the Planarian Body
2.9. Statistical Data Analysis
2.10. Ethical Standards
3. Results and Discussion
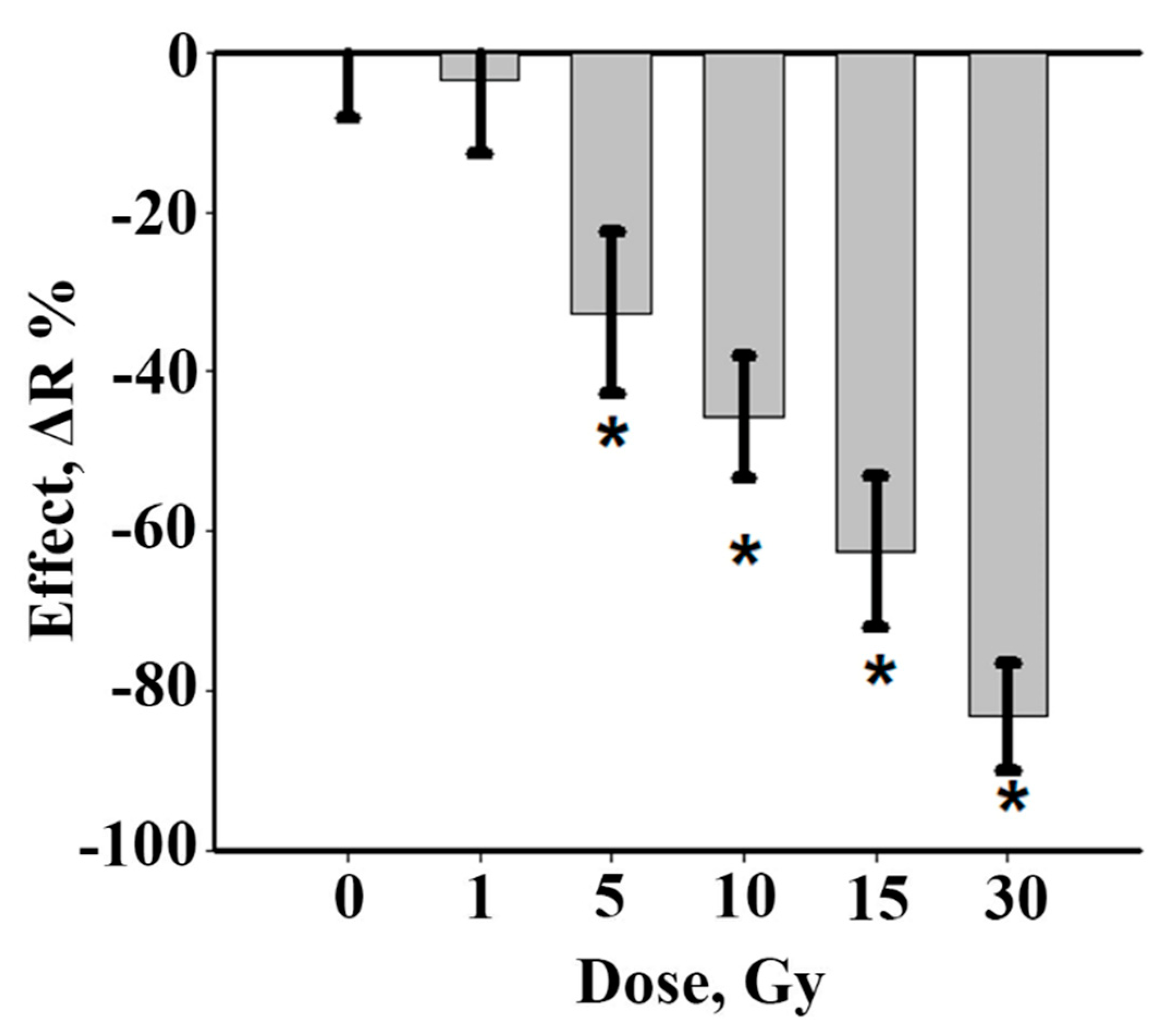
3.1. Working Dose Selection
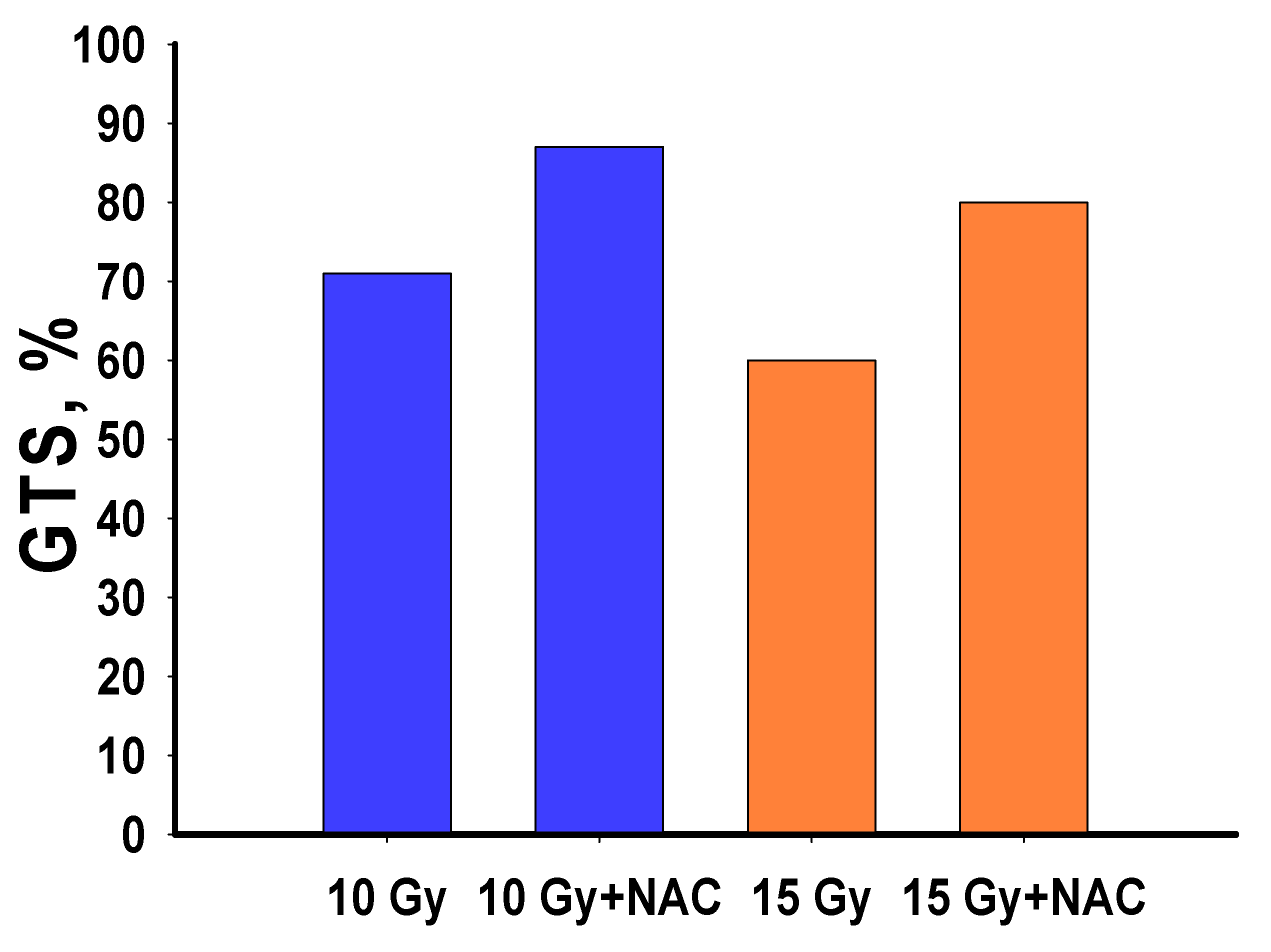
3.2. Blastema Growth Rate in Irradiated Planarians Was Increased by NAC
3.3. Neoblast Survival and Mitotic Activity Was Increased by NAC
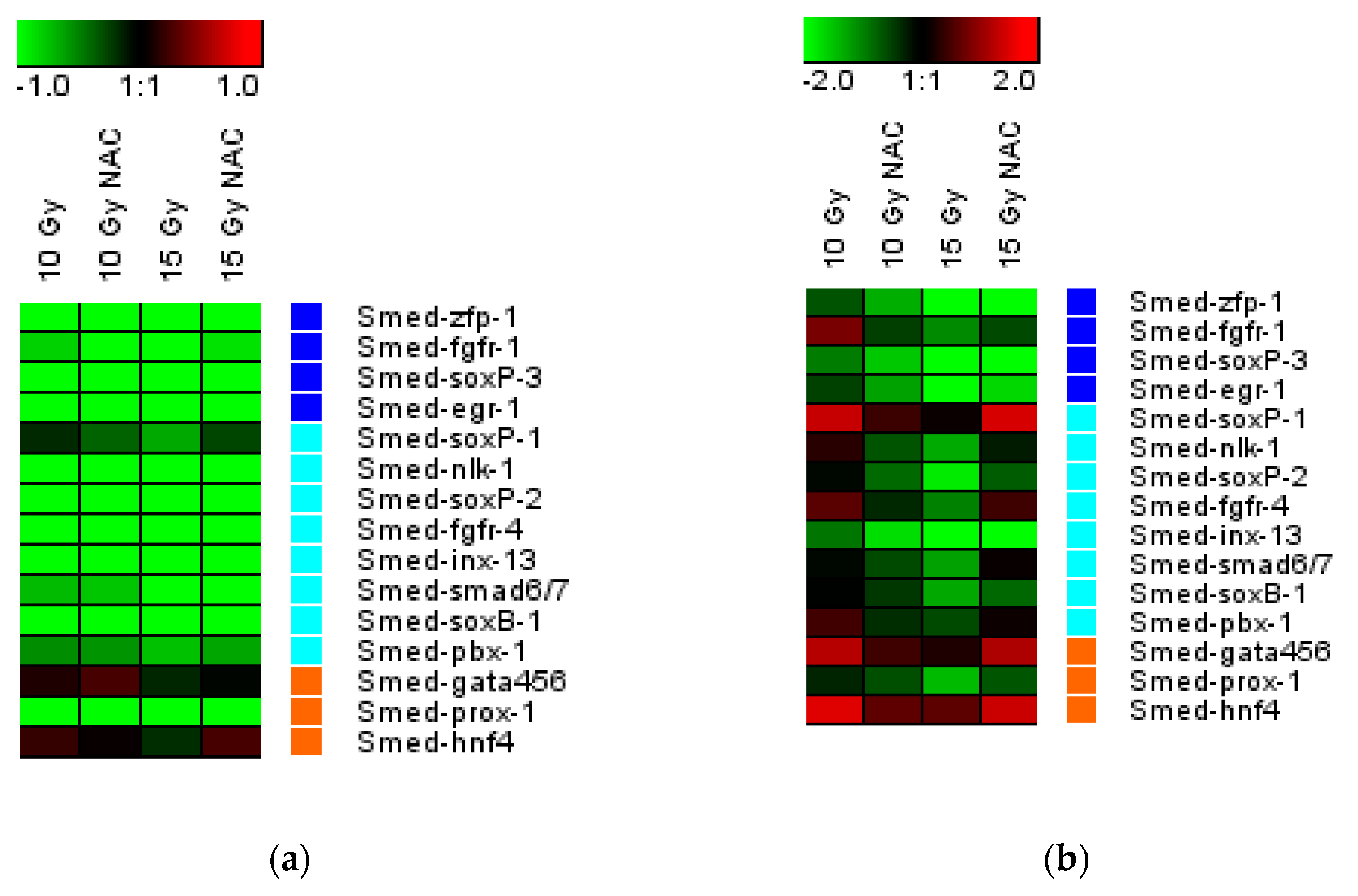
3.4. NAC Increased the Neoblast Markers Expression and Reduced DNA Damage
3.5. The Amount of ROS Generated after Irradiation Was Reduced by NAC
4. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Singh, V.K.; Hauer-Jensen, M. γ-Tocotrienol as a Promising Countermeasure for Acute Radiation Syndrome: Current Status. Int. J. Mol. Sci. 2016, 17, 663. [Google Scholar] [CrossRef] [Green Version]
- Mishra, K.; Alsbeih, G. Appraisal of biochemical classes of radioprotectors: Evidence, current status and guidelines for future development. 3 Biotech 2017, 7, 292. [Google Scholar] [CrossRef] [PubMed]
- Kamran, M.Z.; Ranjan, A.; Kaur, N.; Sur, S.; Tandon, V. Radioprotective Agents: Strategies and Translational Advances. Med. Res. Rev. 2016, 36, 461–493. [Google Scholar] [CrossRef] [PubMed]
- Samuni, Y.; Goldstein, S.; Dean, O.M.; Berk, M. The chemistry and biological activities of N-acetylcysteine. Biochim. Biophys. Acta 2013, 1830, 4117–4129. [Google Scholar] [CrossRef]
- Azzam, E.I.; Jay-Gerin, J.-P.; Pain, D. Ionizing radiation-induced metabolic oxidative stress and prolonged cell injury. Cancer Lett. 2012, 327, 48–60. [Google Scholar] [CrossRef] [Green Version]
- Reisz, J.A.; Bansal, N.; Qian, J.; Zhao, W.; Furdui, C.M. Effects of ionizing radiation on biological molecules--mechanisms of damage and emerging methods of detection. Antioxid. Redox Signal. 2014, 21, 260–292. [Google Scholar] [CrossRef] [PubMed]
- Benrahmoune, M.; Thérond, P.; Abedinzadeh, Z. The reaction of superoxide radical with N-acetylcysteine. Free Radic. Biol. Med. 2000, 29, 775–782. [Google Scholar] [CrossRef]
- Winterbourn, C.C.; Metodiewa, D. Reactivity of biologically important thiol compounds with superoxide and hydrogen peroxide. Free Radic. Biol. Med. 1999, 27, 322–328. [Google Scholar] [CrossRef]
- Gao, X.; Lampraki, E.M.; Al-Khalidi, S.; Qureshi, M.A.; Desai, R.; Wilson, J.B. N-acetylcysteine (NAC) ameliorates Epstein-Barr virus latent membrane protein 1 induced chronic inflammation. PLoS ONE 2017, 12, e0189167. [Google Scholar]
- Ono, K.; Jung, M.; Zhang, T.; Tsutsuki, H.; Sezaki, H.; Ihara, H.; Wei, F.Y.; Tomizawa, K.; Akaike, T.; Sawa, T. Synthesis of l-cysteine derivatives containing stable sulfur isotopes and application of this synthesis to reactive sulfur metabolome. Free Radic. Biol. Med. 2017, 106, 69–79. [Google Scholar] [CrossRef] [PubMed]
- Aldini, G.; Altomare, A.; Baron, G.; Vistoli, G.; Carini, M.; Borsani, L.; Sergio, F. N-Acetylcysteine as an antioxidant and disulphide breaking agent: The reasons why. Free Radic. Res. 2018, 52, 751–762. [Google Scholar] [CrossRef]
- Chouchani, E.T.; Kazak, L.; Jedrychowski, M.P.; Lu, G.Z.; Erickson, B.K.; Szpyt, J.; Pierce, K.A.; Laznik-Bogoslavski, D.; Vetrivelan, R.; Clish, C.B.; et al. Mitochondrial ROS regulate thermogenic energy expenditure and sulfenylation of UCP1. Nature 2016, 532, 112. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Park, E.J.; Min, K.J.; Lee, T.J.; Yoo, Y.H.; Kim, Y.S.; Kwon, T.K. β-Lapachone induces programmed necrosis through the RIP1-PARP-AIF-dependent pathway in human hepatocellular carcinoma SK-Hep1 cells. Cell Death Dis. 2014, 5, e1230. [Google Scholar] [CrossRef] [Green Version]
- Wang, C.; Wong, W.-F. Observational wear leveling: An efficient algorithm for flash memory management. In Proceedings of the 49th Annual Design Automation Conference; ACM: San Francisco, CA, USA, 2012; pp. 235–242. [Google Scholar]
- Kouvaris, J.R.; Kouloulias, V.E.; Vlahos, L.J. Amifostine: The first selective-target and broad-spectrum radioprotector. Oncologist 2007, 12, 738–747. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Singh, V.K.; Seed, T.M. The efficacy and safety of amifostine for the acute radiation syndrome. Expert Opin. Drug Saf. 2019, 18, 1077–1090. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Du, J.; Zhang, P.; Cheng, Y.; Liu, R.; Liu, H.; Gao, F.; Shi, C.; Liu, C. General principles of developing novel radioprotective agents for nuclear emergency. Radiat. Med. Prot. 2020, 1, 120–126. [Google Scholar] [CrossRef]
- Rosen, E.M.; Day, R.; Singh, V.K. New approaches to radiation protection. Front. Oncol. 2015, 4, 381. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Liu, Y.; Zhang, P.; Li, F.; Jin, X.; Li, J.; Chen, W.; Li, Q. Metal-based NanoEnhancers for Future Radiotherapy: Radiosensitizing and Synergistic Effects on Tumor Cells. Theranostics 2018, 8, 1824–1849. [Google Scholar] [CrossRef]
- Rink, J.C. Stem cell systems and regeneration in planaria. Dev. Genes Evol. 2013, 223, 67–84. [Google Scholar] [CrossRef] [Green Version]
- Reddien, P.W.; Sanchez Alvarado, A. Fundamentals of planarian regeneration. Annu. Rev. Cell Dev. Biol. 2004, 20, 725–757. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Roberts-Galbraith, R.H.; Newmark, P.A. On the organ trail: Insights into organ regeneration in the freshwater flatworms. Curr. Opin. Genet. Dev. 2015, 32, 37–46. [Google Scholar] [CrossRef] [PubMed]
- Gentile, L.; Cebrià, F.; Bartscherer, K. The planarian flatworm: An in vivo model for stem cell biology and nervous system regeneration. Dis. Model. Mech. 2011, 4, 12–19. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pagan, O.R. Planaria: An animal model that integrates development, regeneration and pharmacology. Int. J. Dev. Biol. 2017, 61, 519–529. [Google Scholar] [CrossRef]
- Pellettieri, J.; Alvarado, S.A. Cell turnover and adult tissue homeostasis: From humans to planarians. Annu. Rev. Genet. 2007, 41, 83–105. [Google Scholar] [CrossRef] [PubMed]
- Salvetti, A.; Rossi, L.; Bonuccelli, L.; Lena, A.; Pugliesi, C.; Rainaldi, G.; Evangelista, M.; Gremigni, V. Adult stem cell plasticity: Neoblast repopulation in non-lethally irradiated planarians. Dev. Biol. 2009, 328, 305–314. [Google Scholar] [CrossRef]
- Reddien, P.W.; Oviedo, N.J.; Jennings, J.R.; Jenkin, J.C.; Alvarado, S.A. SMEDWI-2 is a PIWI-like protein that regulates planarian stem cells. Science 2005, 310, 1327–1330. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rossi, L.; Cassella, L.; Iacopetti, P.; Ghezzani, C.; Tana, L.; Gimenez, G.; Ghigo, E.; Salvetti, A. Insight into stem cell regulation from sub-lethally irradiated worms. Gene 2018, 662, 37–45. [Google Scholar] [CrossRef] [PubMed]
- Goupil, L.S.; Ivry, S.L.; Hsieh, I.; Suzuki, B.M.; Craik, C.S.; O’Donoghue, A.J.; McKerrow, J.H. Cysteine and Aspartyl Proteases Contribute to Protein Digestion in the Gut of Freshwater Planaria. PLoS Negl. Trop. Dis. 2016, 10, e0004893. [Google Scholar] [CrossRef]
- Ermakov, A.M.; Ermakova, O.N.; Mayevsky, E.I. The role of some intracellular signaling cascades in the activation of planarian regeneration upon irradiation with low-temperature argon plasma. Biophysics 2014, 59, 552–557. [Google Scholar] [CrossRef]
- Newmark, P.A.; Sanchez Alvarado, A. Bromodeoxyuridine specifically labels the regenerative stem cells of planarians. Dev. Biol. 2000, 220, 142–153. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wenemoser, D.; Reddien, P.W. Planarian regeneration involves distinct stem cell responses to wounds and tissue absence. Dev Biol. 2010, 344, 979–991. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hijioka, M.; Ikemoto, Y.; Fukao, K.; Inoue, T.; Kobayakawa, T.; Nishimura, K.; Takata, K.; Agata, K.; Kitamura, Y. MEK/ERK Signaling Regulates Reconstitution of the Dopaminergic Nerve Circuit in the Planarian Dugesia japonica. Neurochem. Res. 2021. [Google Scholar] [CrossRef] [PubMed]
- Hubert, A.; Henderson, J.M.; Cowles, M.W.; Ross, K.G.; Hagen, M.; Anderson, C.; Szeterlak, C.J.; Zayas, R.M. A functional genomics screen identifies an Importin-α homolog as a regulator of stem cell function and tissue patterning during planarian regeneration. BMC Genom. 2015, 16, 769. [Google Scholar] [CrossRef] [Green Version]
- Ermakova, O.N.; Ermakov, A.M.; Tiras, H.P.; Lednev, V.V. The effect of melatonin on the regeneration of planaria Girardia tigrine. Ontogenesis 2009, 6, 466–469. [Google Scholar]
- Ermakov, A.; Popov, A.; Ermakova, O.; Ivanova, O.; Baranchikov, A.; Kamenskikh, K.; Scherbakov, A.; Popova, N.; Ivanov, V. The first inorganic mitogens: Cerium oxide and cerium fluoride nanoparticles stimulate planarian regeneration via neoblastic activation. Mater. Sci. Eng. C 2019, 104, 109924. [Google Scholar] [CrossRef] [PubMed]
- Zhang, H.C.; Shi, C.Y.; Yang, H.H.; Chen, G.W.; Liu, D. Z: Genotoxicity evaluation of ionic liquid 1-octyl-3-methylimidazolium bromide in freshwater planarian Dugesia japonica using RAPD assay. Ecotoxicol. Environ. Saf. 2016, 134, 17–22. [Google Scholar] [CrossRef]
- Ermakov, A.M.; Ermakova, O.N.; Afanasyeva, V.A.; Popov, A.L. Dose-Dependent Effects of Cold Atmospheric Argon Plasma on the Mesenchymal Stem and Osteosarcoma Cells In Vitro. Int. J. Mol. Sci. 2021, 22, 6797. [Google Scholar] [CrossRef]
- van Wolfswinkel, J.C.; Wagner, D.E.; Reddien, P.W. Single-Cell Analysis Reveals Functionally Distinct Classes within the Planarian Stem Cell Compartment. Cell Stem Cell 2014, 15, 326–339. [Google Scholar] [CrossRef] [Green Version]
- Sturn, A.; Quackenbush, J.; Trajanoski, Z. Genesis: Cluster analysis of microarray data. Bioinformatics 2002, 18, 207–208. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lee, J.G.; Noh, W.J.; Kim, H.; Lee, M.-Y. Generation of Reactive Oxygen Species Contributes to the Development of Carbon Black Cytotoxicity to Vascular Cells. Toxicol Res. 2011, 27, 161–166. [Google Scholar] [CrossRef]
- Guedelhoefer, O.C.; Alvarado, S. Amputation induces stem cell mobilization to sites of injury during planarian regeneration. Development 2012, 139, 3510–3520. [Google Scholar] [CrossRef] [Green Version]
- Hayashi, T.; Asami, M.; Higuchi, S.; Shibata, N.; Agata, K. Isolation of planarian X-ray-sensitive stem cells by fluorescence-activated cell sorting. Dev. Growth Differ. 2006, 48, 371–380. [Google Scholar] [CrossRef]
- Isolani, M.E.; Abril, J.F.; Saló, E.; Deri, P.; Bianucci, A.M.; Batistoni, R. Planarians as a Model to Assess In Vivo the Role of Matrix Metalloproteinase Genes during Homeostasis and Regeneration. PLoS ONE 2013, 8, e55649. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Fabbrizi, M.R.; Warshowsky, K.E.; Zobel, C.L.; Hallahan, D.E.; Sharma, G.G. Molecular and epigenetic regulatory mechanisms of normal stem cell radiosensitivity. Cell Death Discov. 2018, 4, 117. [Google Scholar] [CrossRef]
- Sahu, S.; Sridhar, D.; Abnave, P.; Kosaka, N.; Dattani, A.; Thompson, J.M.; Hill, M.A.; Aboobaker, A. Ongoing repair of migration-coupled DNA damage allows planarian adult stem cells to reach wound sites. eLife 2021, 10, e63779. [Google Scholar] [CrossRef] [PubMed]
- Shiroor, D.A.; Bohr, T.E.; Adler, C.E. Injury delays stem cell apoptosis after radiation in planarians. Curr. Biol. 2020, 30, 2166–2174. [Google Scholar] [CrossRef] [PubMed]
- Juan, J.Y.; Zhu, J.Q.; Zhao, C.Z.; Kang, Q.; Zhang, X.; Suo, K.; Cao, N.; Hao, L.; Lu, J. Potential of natural products as radioprotectors and radiosensitizers: Opportunities and challenges. Food Funct. 2021, 12, 5204–5218. [Google Scholar]
- Tiwari, P.; Mishra, K.P. Flavonoids sensitize tumor cells to radiation: Molecular mechanisms and relevance to cancer radiotherapy. Int. J. Radiat. Biol. 2020, 96, 360–369. [Google Scholar] [CrossRef] [PubMed]
- Shibata, N.; Rouhana, L.; Agata, K. Cellular and molecular dissection of pluripotent adult somatic stem cells in planarians. Dev. Growth Differ. 2010, 52, 27–41. [Google Scholar] [CrossRef] [Green Version]



Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Ermakov, A.M.; Kamenskikh, K.A.; Ermakova, O.N.; Blagodatsky, A.S.; Popov, A.L.; Ivanov, V.K. Planarians as an In Vivo Experimental Model for the Study of New Radioprotective Substances. Antioxidants 2021, 10, 1763. https://doi.org/10.3390/antiox10111763
Ermakov AM, Kamenskikh KA, Ermakova ON, Blagodatsky AS, Popov AL, Ivanov VK. Planarians as an In Vivo Experimental Model for the Study of New Radioprotective Substances. Antioxidants. 2021; 10(11):1763. https://doi.org/10.3390/antiox10111763
Chicago/Turabian StyleErmakov, Artem M., Kristina A. Kamenskikh, Olga N. Ermakova, Artem S. Blagodatsky, Anton L. Popov, and Vladimir K. Ivanov. 2021. "Planarians as an In Vivo Experimental Model for the Study of New Radioprotective Substances" Antioxidants 10, no. 11: 1763. https://doi.org/10.3390/antiox10111763
APA StyleErmakov, A. M., Kamenskikh, K. A., Ermakova, O. N., Blagodatsky, A. S., Popov, A. L., & Ivanov, V. K. (2021). Planarians as an In Vivo Experimental Model for the Study of New Radioprotective Substances. Antioxidants, 10(11), 1763. https://doi.org/10.3390/antiox10111763