The Counteracting Effects of Exercise on High-Fat Diet-Induced Memory Impairment: A Systematic Review


Abstract
1. Introduction
2. Methods
2.1. Computerized Searches
2.2. Study Selection
2.3. Inclusionary Criteria
2.4. Data Extraction of Included Studies
3. Results
3.1. Retrieved Articles
3.2. Article Synthesis
4. Discussion
Funding
Conflicts of Interest
References
- DeSalvo, K.B.; Olson, R.; Casavale, K.O. Dietary Guidelines for Americans. JAMA 2016, 315, 1. [Google Scholar] [CrossRef] [PubMed]
- Webster, C.C.; Swart, J.; Noakes, T.D.; Smith, J.A. A Carbohydrate Ingestion Intervention in an Elite Athlete Who Follows a Low-Carbohydrate High-Fat Diet. Int. J. Sports Physiol. Perform. 2018, 13, 957–960. [Google Scholar] [CrossRef] [PubMed]
- Noakes, T.D.; Windt, J. Evidence that supports the prescription of low-carbohydrate high-fat diets: A narrative review. Br. J. Sports Med. 2017, 51, 133–139. [Google Scholar] [CrossRef]
- Molteni, R.; Wu, A.; Vaynman, S.; Ying, Z.; Barnard, R.; Gómez-Pinilla, F. Exercise reverses the harmful effects of consumption of a high-fat diet on synaptic and behavioral plasticity associated to the action of brain-derived neurotrophic factor. Neuroscience 2004, 123, 429–440. [Google Scholar] [CrossRef]
- Maesako, M.; Uemura, K.; Kubota, M.; Kuzuya, A.; Sasaki, K.; Asada, M.; Watanabe, K.; Hayashida, N.; Ihara, M.; Ito, H.; et al. Environmental enrichment ameliorated high-fat diet-induced Aβ deposition and memory deficit in APP transgenic mice. Neurobiol. Aging 2012, 33, 1011.e11–1011.e23. [Google Scholar] [CrossRef]
- Tulving, E. Elements of Episodic Memory; Oxford University Press: New York, NY, USA, 1983. [Google Scholar]
- Mishkin, M.; Suzuki, W.A.; Gadian, D.G.; Vargha-Khadem, F.; Price, T. Hierarchical organization of cognitive memory. Philos. Trans. R. Soc. B Boil. Sci. 1997, 352, 1461–1467. [Google Scholar] [CrossRef]
- Poo, M.-M.; Pignatelli, M.; Ryan, T.J.; Tonegawa, S.; Bonhoeffer, T.; Martin, K.C.; Rudenko, A.; Tsai, L.-H.; Tsien, R.W.; Fishell, G.; et al. What is memory? The present state of the engram. BMC Boil. 2016, 14, 1133. [Google Scholar] [CrossRef] [PubMed]
- Bliss, T.V.P.; Lømo, T. Long-lasting potentiation of synaptic transmission in the dentate area of the anaesthetized rabbit following stimulation of the perforant path. J. Physiol. 1973, 232, 331–356. [Google Scholar] [CrossRef]
- Bol’shakov, V. Mechanisms of long-term synaptic depression in the hippocampus. Rossiiskii Fiziologicheskii Zhurnal Imeni I.M. Sechenova 2001, 87, 441–447. [Google Scholar]
- Doyère, V.; Debiec, J.; Monfils, M.-H.; E Schafe, G.; E LeDoux, J. Synapse-specific reconsolidation of distinct fear memories in the lateral amygdala. Nat. Neurosci. 2007, 10, 414–416. [Google Scholar] [CrossRef]
- Whitlock, J.R.; Heynen, A.J.; Shuler, M.G.; Bear, M.F. Learning induces long-term potentiation in the hippocampus. Science 2006, 313, 1093–1097. [Google Scholar] [CrossRef]
- Gruart, A.; Muñoz, M.D.; Delgado-García, J.M. Involvement of the CA3-CA1 Synapse in the Acquisition of Associative Learning in Behaving Mice. J. Neurosci. 2006, 26, 1077–1087. [Google Scholar] [CrossRef] [PubMed]
- Hao, S.; Dey, A.; Yu, X.; Stranahan, A.M. Dietary obesity reversibly induces synaptic stripping by microglia and impairs hippocampal plasticity. Brain Behav. Immun. 2016, 51, 230–239. [Google Scholar] [CrossRef] [PubMed]
- Valladolid-Acebes, I.; Merino, B.; Principato, A.; Fole, A.; Barbas, C.; Lorenzo, M.P.; Del Olmo, N.; Ruiz-Gayo, M.; Cano, V.; Garcia, A. High-fat diets induce changes in hippocampal glutamate metabolism and neurotransmission. Am. J. Physiol. Metab. 2012, 302, 396–402. [Google Scholar] [CrossRef] [PubMed]
- Karimi, S.A.; Salehi, I.; Komaki, A.; Sarihi, A.; Zarei, M.; Shahidi, S. Effect of high-fat diet and antioxidants on hippocampal long-term potentiation in rats: An in vivo study. Brain Res. 2013, 1539, 1–6. [Google Scholar] [CrossRef] [PubMed]
- Wang, Z.; Fan, J.; Wang, J.; Li, Y.; Xiao, L.; Duan, D.; Wang, Q. Protective effect of lycopene on high-fat diet-induced cognitive impairment in rats. Neurosci. Lett. 2016, 627, 185–191. [Google Scholar] [CrossRef]
- Cano, V.; Valladolid-Acebes, I.; Hernández-Nuño, F.; Merino, B.; Del Olmo, N.; Chowen, J.A.; Ruiz-Gayo, M. Morphological changes in glial fibrillary acidic protein immunopositive astrocytes in the hippocampus of dietary-induced obese mice. NeuroReport 2014, 25, 819–822. [Google Scholar] [CrossRef] [PubMed]
- Page, K.C.; Jones, E.K.; Anday, E.K. Maternal and postweaning high-fat diets disturb hippocampal gene expression, learning, and memory function. Am. J. Physiol. Integr. Comp. Physiol. 2014, 306, 527–537. [Google Scholar] [CrossRef] [PubMed]
- Molteni, R.; Barnard, R.; Ying, Z.; Roberts, C.; Gómez-Pinilla, F. A high-fat, refined sugar diet reduces hippocampal brain-derived neurotrophic factor, neuronal plasticity, and learning. Neuroscience 2002, 112, 803–814. [Google Scholar] [CrossRef]
- Tozuka, Y.; Kumon, M.; Wada, E.; Onodera, M.; Mochizuki, H.; Wada, K. Maternal obesity impairs hippocampal BDNF production and spatial learning performance in young mouse offspring. Neurochem. Int. 2010, 57, 235–247. [Google Scholar] [CrossRef]
- Cassilhas, R.; Lee, K.; Fernandes, J.; Oliveira, M.; Tufik, S.; Meeusen, R.; De Mello, M. Spatial memory is improved by aerobic and resistance exercise through divergent molecular mechanisms. Neuroscience 2012, 202, 309–317. [Google Scholar] [CrossRef] [PubMed]
- Van Praag, H.; Christie, B.R.; Sejnowski, T.J.; Gage, F.H. Running enhances neurogenesis, learning, and long-term potentiation in mice. Proc. Natl. Acad. Sci. USA 1999, 96, 13427–13431. [Google Scholar] [CrossRef] [PubMed]
- Huang, Y.-Q.; Wu, C.; He, X.-F.; Wu, D.; He, X.; Liang, F.-Y.; Dai, G.-Y.; Pei, Z.; Xu, G.-Q.; Lan, Y. Effects of Voluntary Wheel-Running Types on Hippocampal Neurogenesis and Spatial Cognition in Middle-Aged Mice. Front. Cell. Neurosci. 2018, 12, 177. [Google Scholar] [CrossRef] [PubMed]
- Molteni, R.; Ying, Z.; Gomez-Pinilla, F.; Gómez-Pinilla, F. Differential effects of acute and chronic exercise on plasticity-related genes in the rat hippocampus revealed by microarray. Eur. J. Neurosci. 2002, 16, 1107–1116. [Google Scholar] [CrossRef] [PubMed]
- Dietrich, M.O.; Mantese, C.E.; Porciúncula, L.O.; Ghisleni, G.; Vinade, L.; Souza, D.O.; Portela, L.V. Exercise affects glutamate receptors in postsynaptic densities from cortical mice brain. Brain Res. 2005, 1065, 20–25. [Google Scholar] [CrossRef] [PubMed]
- Bramer, W.M.; Rethlefsen, M.L.; Kleijnen, J.; Franco, O.H. Optimal database combinations for literature searches in systematic reviews: A prospective exploratory study. Syst. Rev. 2017, 6, 245. [Google Scholar] [CrossRef] [PubMed]
- Klein, C.; Jonas, W.; Iggena, D.; Empl, L.; Rivalan, M.; Wiedmer, P.; Spranger, J.; Hellweg, R.; Winter, Y.; Steiner, B. Exercise prevents high-fat diet-induced impairment of flexible memory expression in the water maze and modulates adult hippocampal neurogenesis in mice. Neurobiol. Learn. Mem. 2016, 131, 26–35. [Google Scholar] [CrossRef]
- Maesako, M.; Uemura, K.; Iwata, A.; Kubota, M.; Watanabe, K.; Uemura, M.; Noda, Y.; Asada-Utsugi, M.; Kihara, T.; Takahashi, R.; et al. Continuation of Exercise Is Necessary to Inhibit High Fat Diet-Induced β-Amyloid Deposition and Memory Deficit in Amyloid Precursor Protein Transgenic Mice. PLoS ONE 2013, 8, e72796. [Google Scholar] [CrossRef]
- Maesako, M.; Uemura, K.; Kubota, M.; Kuzuya, A.; Sasaki, K.; Hayashida, N.; Asada-Utsugi, M.; Watanabe, K.; Uemura, M.; Kihara, T.; et al. Exercise Is More Effective than Diet Control in Preventing High Fat Diet-induced β-Amyloid Deposition and Memory Deficit in Amyloid Precursor Protein Transgenic Mice. J. Boil. Chem. 2012, 287, 23024–23033. [Google Scholar] [CrossRef]
- Woo, J.; Shin, K.O.; Park, S.Y.; Jang, K.S.; Kang, S. Effects of exercise and diet change on cognition function and synaptic plasticity in high fat diet induced obese rats. Lipids Heal. Dis. 2013, 12, 144. [Google Scholar] [CrossRef]
- Noble, E.E.; Mavanji, V.; Little, M.R.; Billington, C.J.; Kotz, C.M.; Wang, C. Exercise reduces diet-induced cognitive decline and increases hippocampal brain-derived neurotrophic factor in CA3 neurons. Neurobiol. Learn. Mem. 2014, 114, 40–50. [Google Scholar] [CrossRef] [PubMed]
- Cheng, J.; Chen, L.; Han, S.; Qin, L.; Chen, N.; Wan, Z. Treadmill Running and Rutin Reverse High Fat Diet Induced Cognitive Impairment in Diet Induced Obese Mice. J. Nutr. Health Aging 2016, 20, 503–508. [Google Scholar] [CrossRef] [PubMed]
- Kang, E.; Koo, J.; Jang, Y.; Yang, C.; Lee, Y.; Cosio-Lima, L.M.; Cho, J. Neuroprotective Effects of Endurance Exercise against High Fat Diet-Induced Hippocampal Neuroinflammation. J. Neuroendocr. 2016, 28, 28. [Google Scholar] [CrossRef] [PubMed]
- Kim, T.-W.; Choi, H.-H.; Chung, Y.-R. Treadmill exercise alleviates impairment of cognitive function by enhancing hippocampal neuroplasticity in the high-fat diet-induced obese mice. J. Exerc. Rehabil. 2016, 12, 156–162. [Google Scholar] [CrossRef] [PubMed]
- Park, H.S.; Cho, H.S.; Kim, T.W. Physical exercise promotes memory capability by enhancing hippocampal mitochondrial functions and inhibiting apoptosis in obesity-induced insulin resistance by high fat diet. Metab. Brain Dis. 2018, 33, 283–292. [Google Scholar] [CrossRef] [PubMed]
- Cheng, M.; Cong, J.; Wu, Y.; Xie, J.; Wang, S.; Zhao, Y.; Zang, X. Chronic Swimming Exercise Ameliorates Low-Soybean-Oil Diet-Induced Spatial Memory Impairment by Enhancing BDNF-Mediated Synaptic Potentiation in Developing Spontaneously Hypertensive Rats. Neurochem. Res. 2018, 43, 1047–1057. [Google Scholar] [CrossRef] [PubMed]
- Jeong, J.-H.; Koo, J.-H.; Cho, J.-Y.; Kang, E.-B. Neuroprotective effect of treadmill exercise against blunted brain insulin signaling, NADPH oxidase, and Tau hyperphosphorylation in rats fed a high-fat diet. Brain Res. Bull. 2018, 142, 374–383. [Google Scholar] [CrossRef]
- Jeong, J.-H.; Kang, E.-B. Effects of treadmill exercise on PI3K/AKT/GSK-3β pathway and tau protein in high-fat diet-fed rats. J. Exerc. Nutr. Biochem. 2018, 22, 9–14. [Google Scholar] [CrossRef] [PubMed]
- Shi, Z.; Li, C.; Yin, Y.; Yang, Z.; Xue, H.; Mu, N.; Wang, Y.; Liu, M.; Ma, H. Aerobic Interval Training Regulated SIRT3 Attenuates High-Fat-Diet-Associated Cognitive Dysfunction. BioMed Res. Int. 2018, 2018, 1–8. [Google Scholar] [CrossRef]
- Han, T.-K.; Leem, Y.-H.; Kim, H.-S. Treadmill exercise restores high fat diet-induced disturbance of hippocampal neurogenesis through β2-adrenergic receptor-dependent induction of thioredoxin-1 and brain-derived neurotrophic factor. Brain Res. 2019, 1707, 154–163. [Google Scholar] [CrossRef]
- Mehta, B.K.; Singh, K.K.; Banerjee, S. Effect of exercise on type 2 diabetes-associated cognitive impairment in rats. Int. J. Neurosci. 2019, 129, 252–263. [Google Scholar] [CrossRef] [PubMed]
- Blitzer, R.D. Teaching resources. Long-term potentiation: Mechanisms of induction and maintenance. Sci. STKE 2005, 2005, tr26. [Google Scholar] [CrossRef] [PubMed]
- Sweatt, J.D. Toward a Molecular Explanation for Long-Term Potentiation. Learn. Mem. 1999, 6, 399–416. [Google Scholar] [CrossRef] [PubMed]
- Buonarati, O.R.; Hammes, E.A.; Watson, J.F.; Greger, I.H.; Hell, J.W. Mechanisms of postsynaptic localization of AMPA-type glutamate receptors and their regulation during long-term potentiation. Sci. Signal. 2019, 12, eaar6889. [Google Scholar] [CrossRef] [PubMed]
- Sacktor, T.C.; Fenton, A.A. What does LTP tell us about the roles of CaMKII and PKMzeta in memory? Mol. Brain 2018, 11, 77. [Google Scholar] [CrossRef] [PubMed]
- Baltaci, S.B.; Mogulkoc, R.; Baltacim, A.K. Molecular Mechanisms of Early and Late LTP. Neurochem. Res. 2019, 44, 281–296. [Google Scholar] [CrossRef] [PubMed]
- Park, M. AMPA Receptor Trafficking for Postsynaptic Potentiation. Front. Cell. Neurosci. 2018, 12, 361. [Google Scholar] [CrossRef] [PubMed]
- Mizuno, M.; Yamada, K.; He, J.; Nakajima, A.; Nabeshima, T. Involvement of BDNF Receptor TrkB in Spatial Memory Formation. Learn. Mem. 2003, 10, 108–115. [Google Scholar] [CrossRef] [PubMed]
- Mizuno, M.; Yamada, K.; Olariu, A.; Nawa, H.; Nabeshima, T. Involvement of Brain-Derived Neurotrophic Factor in Spatial Memory Formation and Maintenance in a Radial Arm Maze Test in Rats. J. Neurosci. 2000, 20, 7116–7121. [Google Scholar] [CrossRef] [PubMed]
- Sleiman, S.F.; Henry, J.; Al-Haddad, R.; El Hayek, L.; Haidar, E.A.; Stringer, T.; Ulja, D.; Karuppagounder, S.S.; Holson, E.B.; Ratan, R.R.; et al. Exercise promotes the expression of brain derived neurotrophic factor (BDNF) through the action of the ketone body β-hydroxybutyrate. eLife 2016, 5, 5. [Google Scholar] [CrossRef] [PubMed]
- Qiao, S.; Peng, R.; Yan, H.; Gao, Y.; Wang, C.; Wang, S.; Zou, Y.; Xu, X.; Zhao, L.; Dong, J.; et al. Reduction of Phosphorylated Synapsin I (Ser-553) Leads to Spatial Memory Impairment by Attenuating GABA Release after Microwave Exposure in Wistar Rats. PLoS ONE 2014, 9, e95503. [Google Scholar] [CrossRef] [PubMed]
- John, J.P.P.; Sunyer, B.; Höger, H.; Pollak, A.; Lubec, G. Hippocampal synapsin isoform levels are linked to spatial memory enhancement by SGS742. Hippocampus 2009, 19, 731–738. [Google Scholar] [CrossRef] [PubMed]
- Vaynman, S.; Ying, Z.; Gómez-Pinilla, F.; Gómez-Pinilla, F. Exercise induces BDNF and synapsin I to specific hippocampal subfields. J. Neurosci. Res. 2004, 76, 356–362. [Google Scholar] [CrossRef]
- Jovanovic, J.N.; Czernik, A.J.; Fienberg, A.A.; Greengard, P.; Sihra, T.S. Synapsins as mediators of BDNF-enhanced neurotransmitter release. Nat. Neurosci. 2000, 3, 323–329. [Google Scholar] [CrossRef] [PubMed]
- Rossi, C.; Angelucci, A.; Costantin, L.; Braschi, C.; Mazzantini, M.; Babbini, F.; Fabbri, M.E.; Tessarollo, L.; Maffei, L.; Berardi, N.; et al. Brain-derived neurotrophic factor (BDNF) is required for the enhancement of hippocampal neurogenesis following environmental enrichment. Eur. J. Neurosci. 2006, 24, 1850–1856. [Google Scholar] [CrossRef] [PubMed]
- Dupret, D.; Revest, J.-M.; Koehl, M.; Ichas, F.; De Giorgi, F.; Costet, P.; Abrous, D.N.; Piazza, P.V. Spatial Relational Memory Requires Hippocampal Adult Neurogenesis. PLoS ONE 2008, 3, e1959. [Google Scholar] [CrossRef]
- França, T.F.A.; Bitencourt, A.M.; Maximilla, N.R.; Barros, D.M.; Monserrat, J.M. Hippocampal neurogenesis and pattern separation: A meta-analysis of behavioral data. Hippocampus 2017, 27, 937–950. [Google Scholar] [CrossRef]
- Lindqvist, A.; Mohapel, P.; Bouter, B.; Frielingsdorf, H.; Pizzo, D.; Brundin, P.; Erlanson-Albertsson, C.; Erlanson-Albertsson, C.; Erlanson-Albertsson, C. High-fat diet impairs hippocampal neurogenesis in male rats. Eur. J. Neurol. 2006, 13, 1385–1388. [Google Scholar] [CrossRef]
- Lee, J.; Duan, W.; Mattson, M.P. Evidence that brain-derived neurotrophic factor is required for basal neurogenesis and mediates, in part, the enhancement of neurogenesis by dietary restriction in the hippocampus of adult mice. J. Neurochem. 2002, 82, 1367–1375. [Google Scholar] [CrossRef]


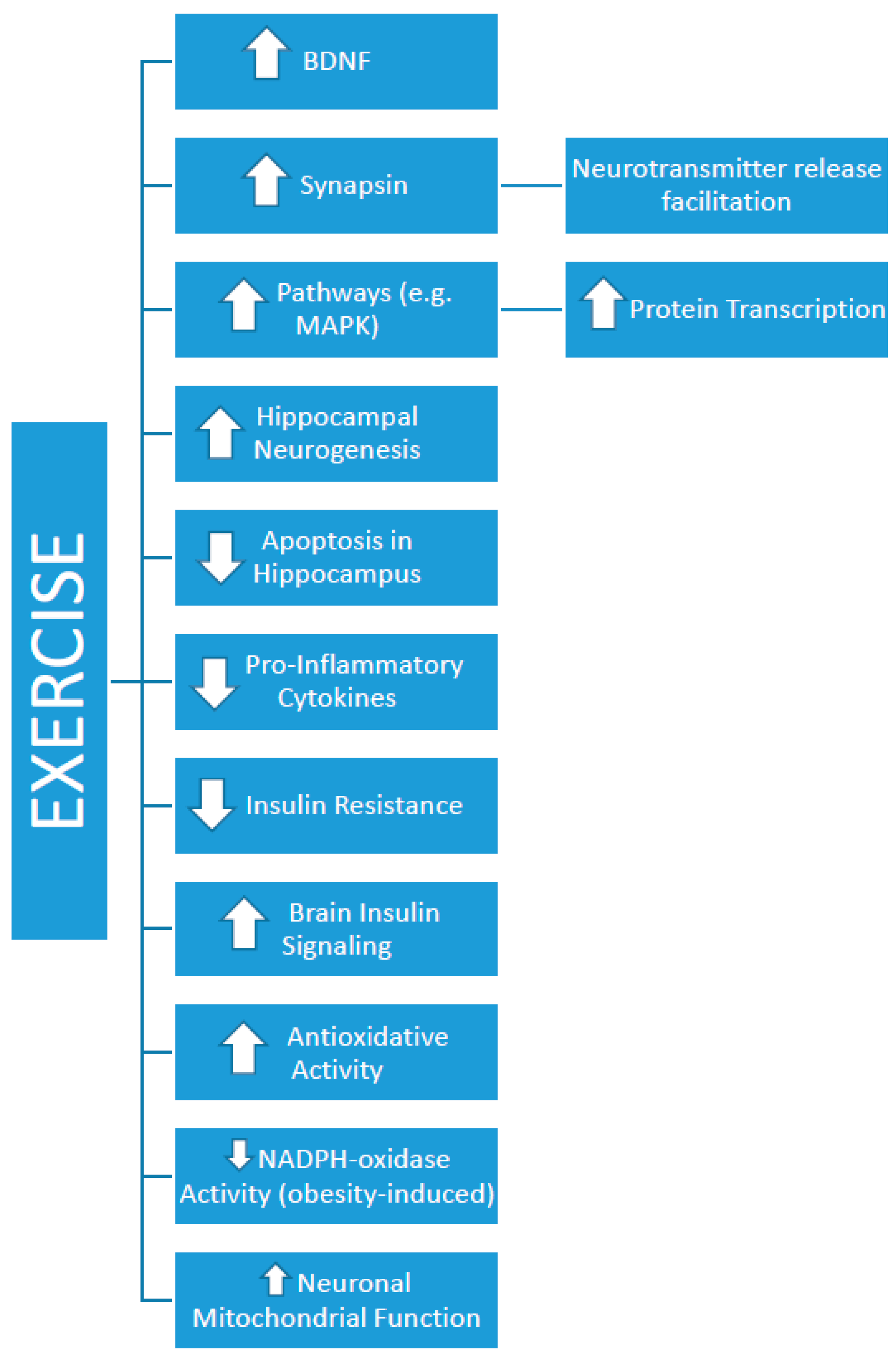{kind=link}
| Study | Subjects | Exercise Protocol | Diet Protocol | Temporal Assessment of Exercise and Diet | Memory Assessment | Did High-Fat Diet Impair Memory? | Did Exercise Counteract Diet-Induced Memory Impairment? | Mechanisms |
|---|---|---|---|---|---|---|---|---|
| Molteni et al. (2004) [4] | Fisher 344 rats, 2 months old | Free access to running wheel for 2 months. | 2 months on high in saturated and monounsaturated fat (primarily from lard plus a small amount of corn oil, approx. 39% energy) | Concurrent | Morris water maze | Yes | Yes | Exercise reversed the decrease in BDNF and its downstream effector, synapsin I (involved in BDNF release). Exercise also increase CREB transcription. |
| Maesako et al. (2012) [5] | APP transgenic mice overexpressing the familial AD-linked mutation | Enriched environment with access to running wheel; this occurred from weeks 10–20 (i.e., 10 weeks into the high-fat diet). | 20 weeks of high-fat diet, involving 60% fat, 20% CHO, and 20% protein | Concurrent | Morris water maze | Yes | Yes | Enriched environment attenuated diet-induced Aβ deposition. |
| Maesako et al. (2012) [30] | APP transgenic mice overexpressing the familial AD-linked mutation | Voluntary access to running wheel. | 20 weeks of high-fat diet, involving 60% fat, 20% CHO, and 20% protein | Concurrent | Morris water maze | Yes | Yes | Exercise attenuated diet-induced Aβ deposition and strengthened the activity of neprilysin, the Aβ-degrading enzyme. |
| Maesako et al. (2013) [29] | APP transgenic mice overexpressing the familial AD-linked mutation | Voluntary access to running wheel. | 20 weeks of high-fat diet, involving 60% fat, 20% CHO, and 20% protein | Concurrent | Morris water maze | Yes | Yes, but only if the exercise occurred throughout the majority of the diet protocol | |
| Woo et al. (2013) [31] | 4-week-old Sprague–Dawley rats | Treadmill exercise for the first 8 weeks, involving a progressive exercise program, ranging from 40 to 60 min/day of exercise. | 13-weeks of high-fat diet, involving 45% fat | Concurrent | Morris water maze | Yes | Yes | Upregulation of BDNF and MAPK. |
| Noble et al. (2014) [32] | 7-month-old Naïve rats | Forced treadmill or voluntary wheel access for 7 weeks | 16 weeks of high-fat diet | Exercise occurring after high-fat diet (treatment) | Two-way active avoidance test | Yes | Yes | Increased BDNF in CA3. |
| Cheng et al. (2016) [33] | Twelve-week-old C57BL/6J mice | Treadmill running, 60 min/day, 5 times/week, 15 m/min, for 16 weeks. | 16 weeks of high-fat diet ad libitum, involving 60% fat, 20% CHO, and 20% protein | Concurrent | Morris water maze | Yes | Yes | p-CREB, BACE1, IDE, and PSD95 were likely mediators of this effect. |
| Kang et al. (2016) [34] | Sprague–Dawley rats aged 8 weeks | Treadmill running, 30 min/day, 5 days/week, for 8 weeks. | High fat diet (60% fat) for 20 weeks | Exercise occurring after high-fat diet (treatment) | Passive avoidance task | Yes | Yes | Reduction in pro-inflammatory cytokines (TNF-α, interleukin-1β). |
| Kim et al. (2016) [35] | Male C57BL/6 mice, 4-weeks old | Treadmill exercise, ranging from 30 to 50 min/day; progressive over a 20-week period. | High-fat diet (60% fat) for 20 weeks ad libitum | Exercise occurring after high-fat diet (treatment) | Y-maze test and radial-8-arm maze test | Yes | Yes | Increased expression of BDNF and TrkB and enhanced cell proliferation. |
| Klein et al. (2016) [28] | Six-week-old female C57Bl/6N mice | Free access to running wheel. | 12 weeks of high-fat diet, involving 60% fat, 20% CHO, and 20% protein | Concurrent | Morris water maze | Yes, but only in adolescent | Yes | Modulation of hippocampal neurogenesis. |
| Park et al. (2017) [36] | Male 4-week-old C57BL/6 mice | Treadmill exercise, 6 days/week, approx. 40 min/day, for 12 weeks. | 20 weeks of high-fat diet, involving 60% fat ad libitum | Exercise occurring after high-fat diet (treatment) | Step-down avoidance task | Yes | Yes | Reduced insulin resistance, improved mitochondrial function, and reduced apoptosis in the hippocampus. |
| Cheng et al. (2018) [37] | Male 3-week-old SHR and normotensive Wistar–Kyoto rats | Swimming exercise for 6 weeks. | 6 weeks of low-soybean oil diet | Concurrently | Morris water maze | Yes | Yes | Up-regulation of BDNF and NMDA-r expression. |
| Jeong et al. (2018) [38] | Sprague–Dawley rats aged 20 weeks | Treadmill exercise for 8 weeks, 30 min/day, 8 m/min, 5 days/week. | High-fat diet for 20 weeks | Exercise occurring after high-fat diet (treatment) | Water maze and passive avoidance tasks | Yes | Yes | Improved brain insulin signaling, inhibition of obesity-induced NADPH-oxidase activity. |
| Jeong et al. (2018) [39] | Sprague–Dawley rats aged 8 weeks | Treadmill exercise for 8 weeks, 30 min/day, 5 days/week, progressive intensity. | High fat diet for 20 weeks, including 20% CHO, 60% fat, and 20% protein | Exercise occurring after high-fat diet (treatment) | Passive avoidance task | Yes | Yes | Improved brain insulin signaling (PI3K/AKT/GSK-3β), reduced tau hyperphosphorylation. |
| Shi et al. (2018) [40] | Male C57BL/6 mice and SIRT3 mice (2-months old) | Exercise started at week 6 and continued for the remaining 6 weeks. Engaged in aerobic intermittent training, 30 min/day, 5 days/week. Intermittent exercise involved 4-min bursts at 80–85% of VO2max, with 2 min active recovery periods. | High-fat diet of 45% kcal fat, 20% kcal protein, and 35% kcal CHO for 12 weeks | Concurrent | Morris water maze | Yes | Yes | SIRT3 upregulation and improvement in antioxidative activity |
| Han et al. (2019) [41] | Six-week-old C57BL/6 mice | 23 weeks of treadmill running, 30 min/day, 5 days/week, at 8 m/min. | 23 weeks of high-fat diet ad libitum, involving 60% fat | Concurrent | Morris water maze | Yes | Yes | Reduced number of apoptotic cells and increased BDNF. |
| Mehta et al. (2019) [42] | Sprague–Dawley male rats | Running wheel access for 6 weeks, 25–30 min/day, 5 days/week. | 15 days of high-fat diet (310 gm/kg Lard) | Exercise occurring after high-fat diet (treatment) | Passive avoidance and elevated plus maze | Yes | Yes | Reduction in neuroinflammatory markers (e.g., IL-1β, TNF-α). |
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Loprinzi, P.D.; Ponce, P.; Zou, L.; Li, H. The Counteracting Effects of Exercise on High-Fat Diet-Induced Memory Impairment: A Systematic Review. Brain Sci. 2019, 9, 145. https://doi.org/10.3390/brainsci9060145
Loprinzi PD, Ponce P, Zou L, Li H. The Counteracting Effects of Exercise on High-Fat Diet-Induced Memory Impairment: A Systematic Review. Brain Sciences. 2019; 9(6):145. https://doi.org/10.3390/brainsci9060145
Chicago/Turabian StyleLoprinzi, Paul D., Pamela Ponce, Liye Zou, and Hong Li. 2019. "The Counteracting Effects of Exercise on High-Fat Diet-Induced Memory Impairment: A Systematic Review" Brain Sciences 9, no. 6: 145. https://doi.org/10.3390/brainsci9060145
APA StyleLoprinzi, P. D., Ponce, P., Zou, L., & Li, H. (2019). The Counteracting Effects of Exercise on High-Fat Diet-Induced Memory Impairment: A Systematic Review. Brain Sciences, 9(6), 145. https://doi.org/10.3390/brainsci9060145