Pallidal Stimulation Modulates Pedunculopontine Nuclei in Parkinson’s Disease

{kind=link}
{kind=link}
{kind=link}
{kind=link}
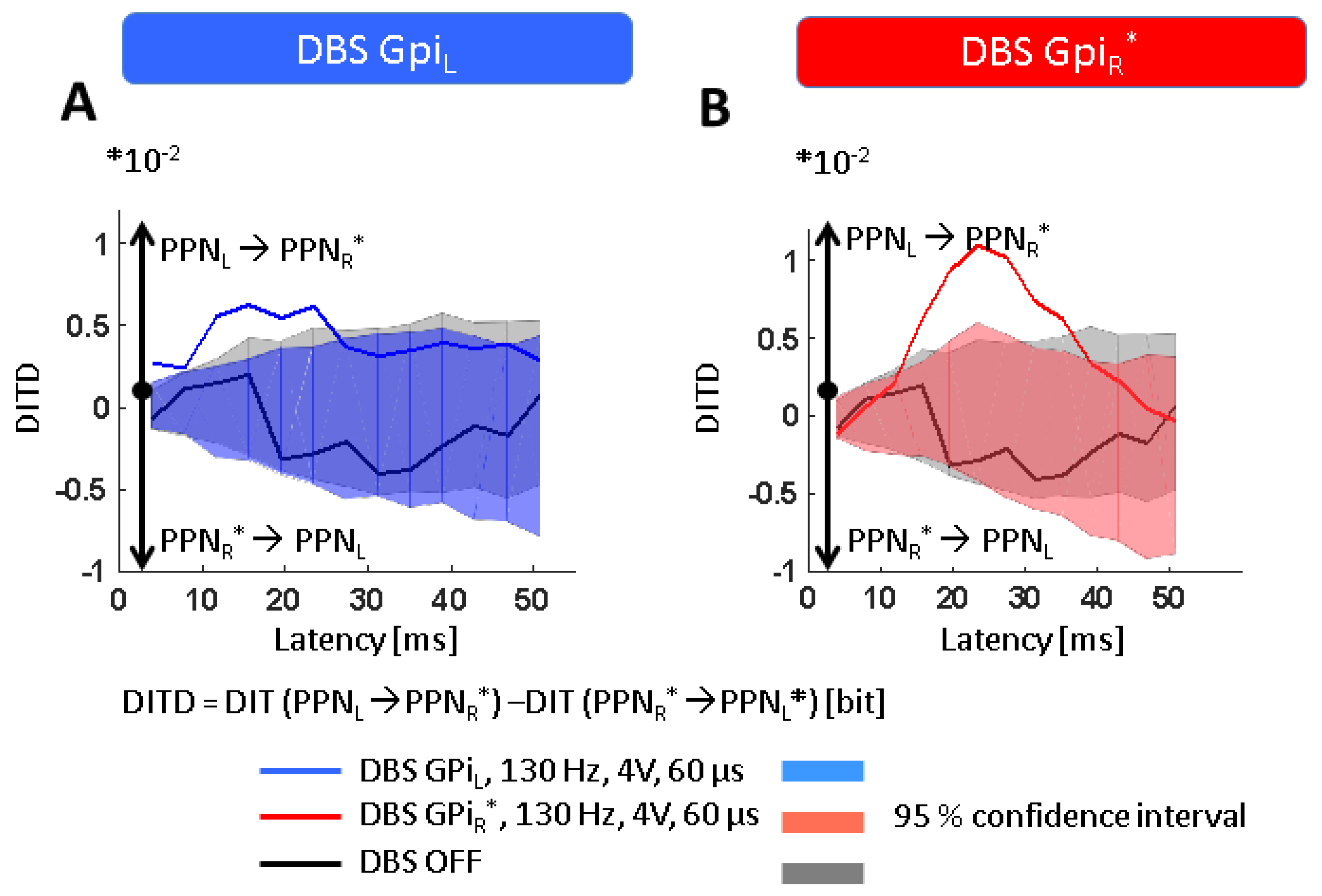{kind=link}
Abstract
1. Introduction
2. Methods
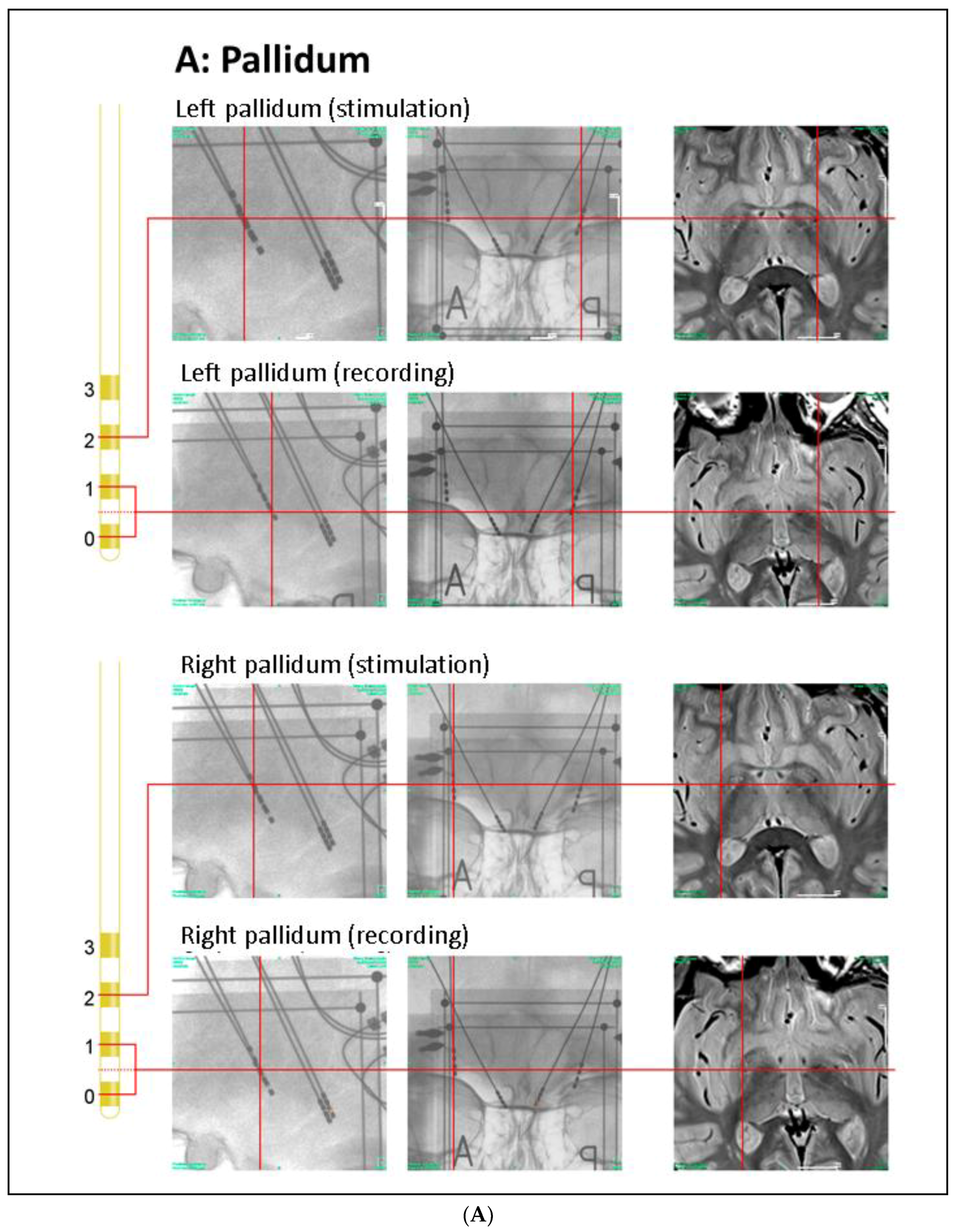
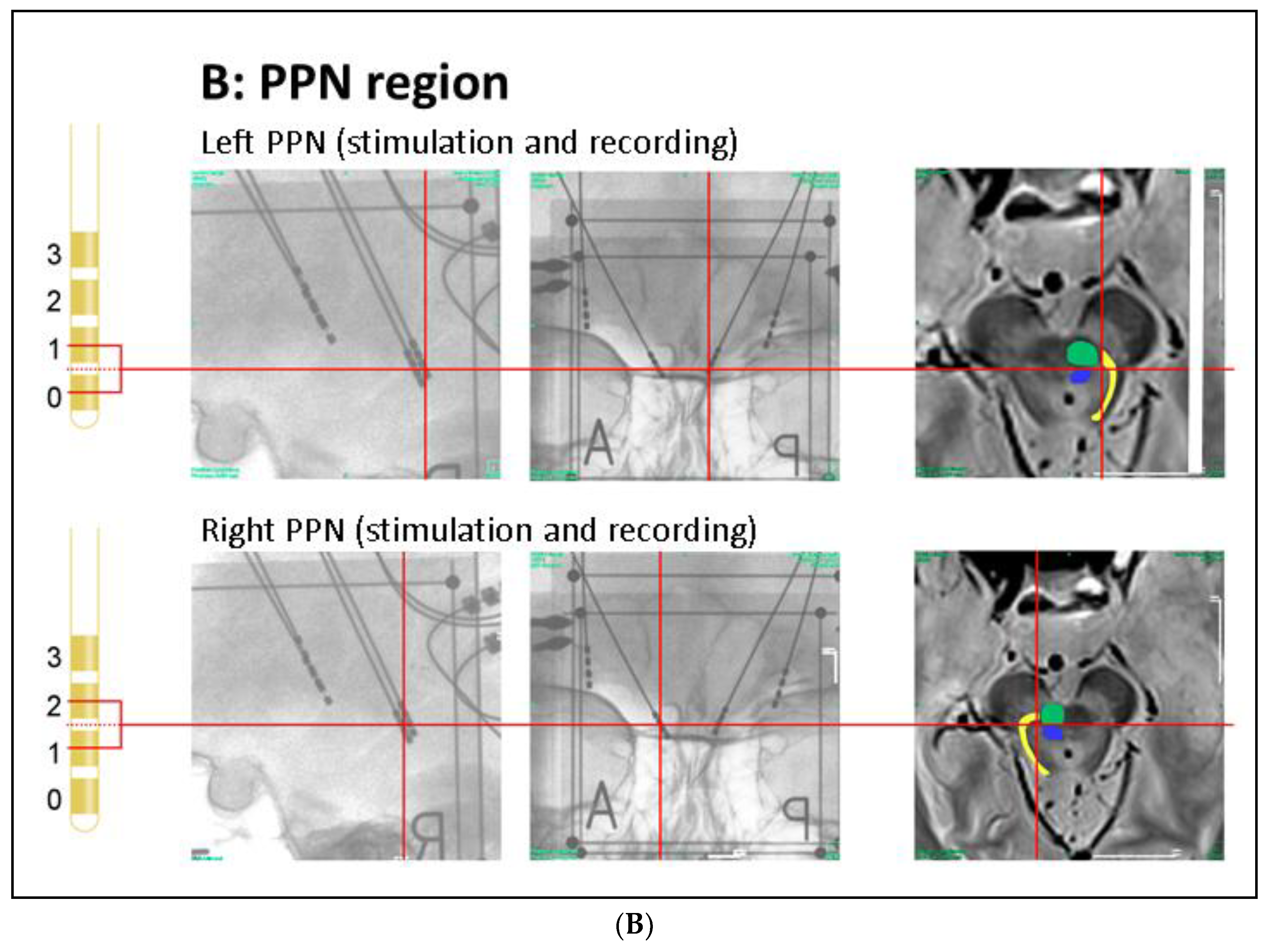
Patient’s Clinical Data and Surgery
3. Clinical Assessments
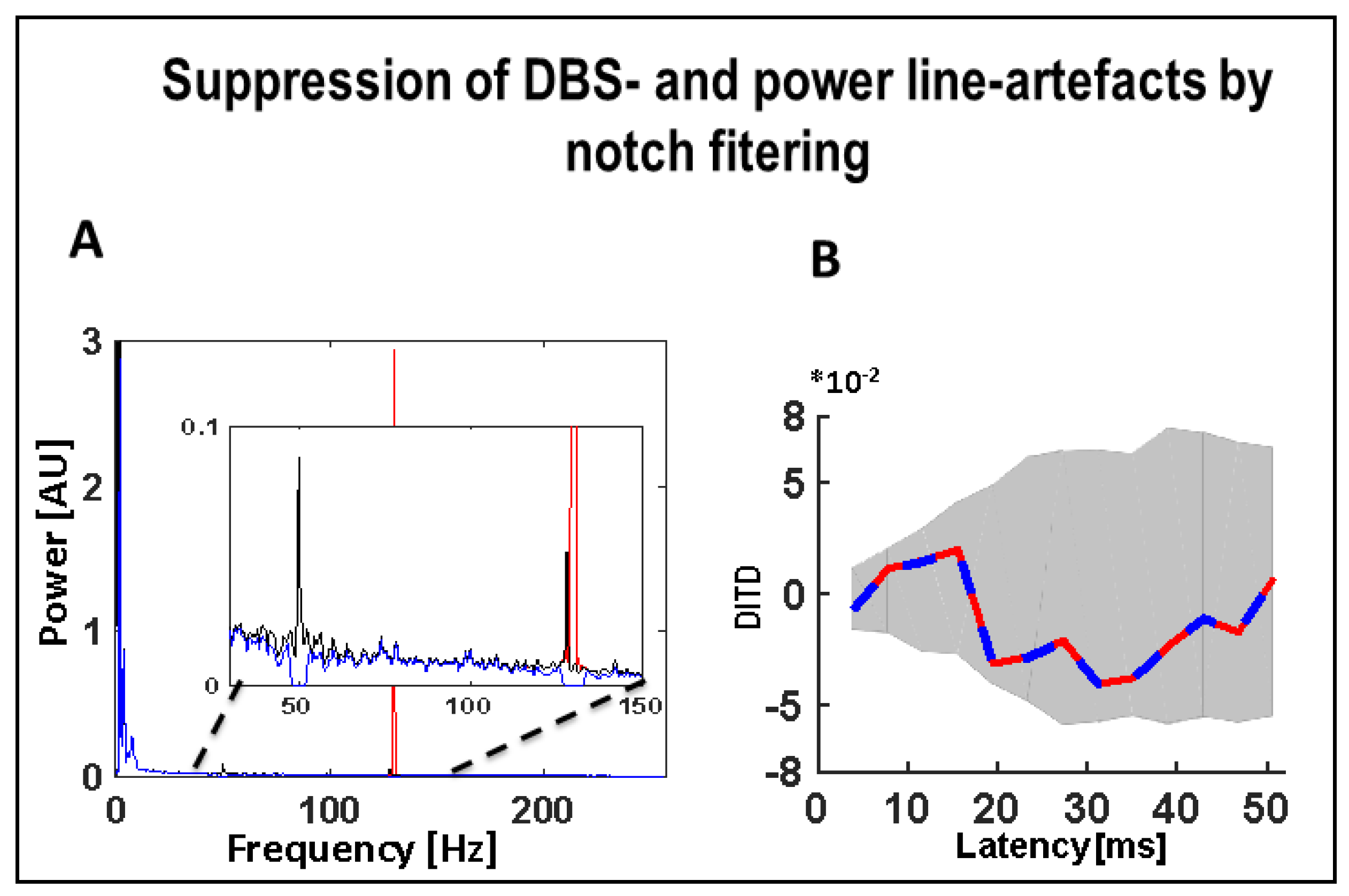
4. Recordings, Analysis, and Statistics
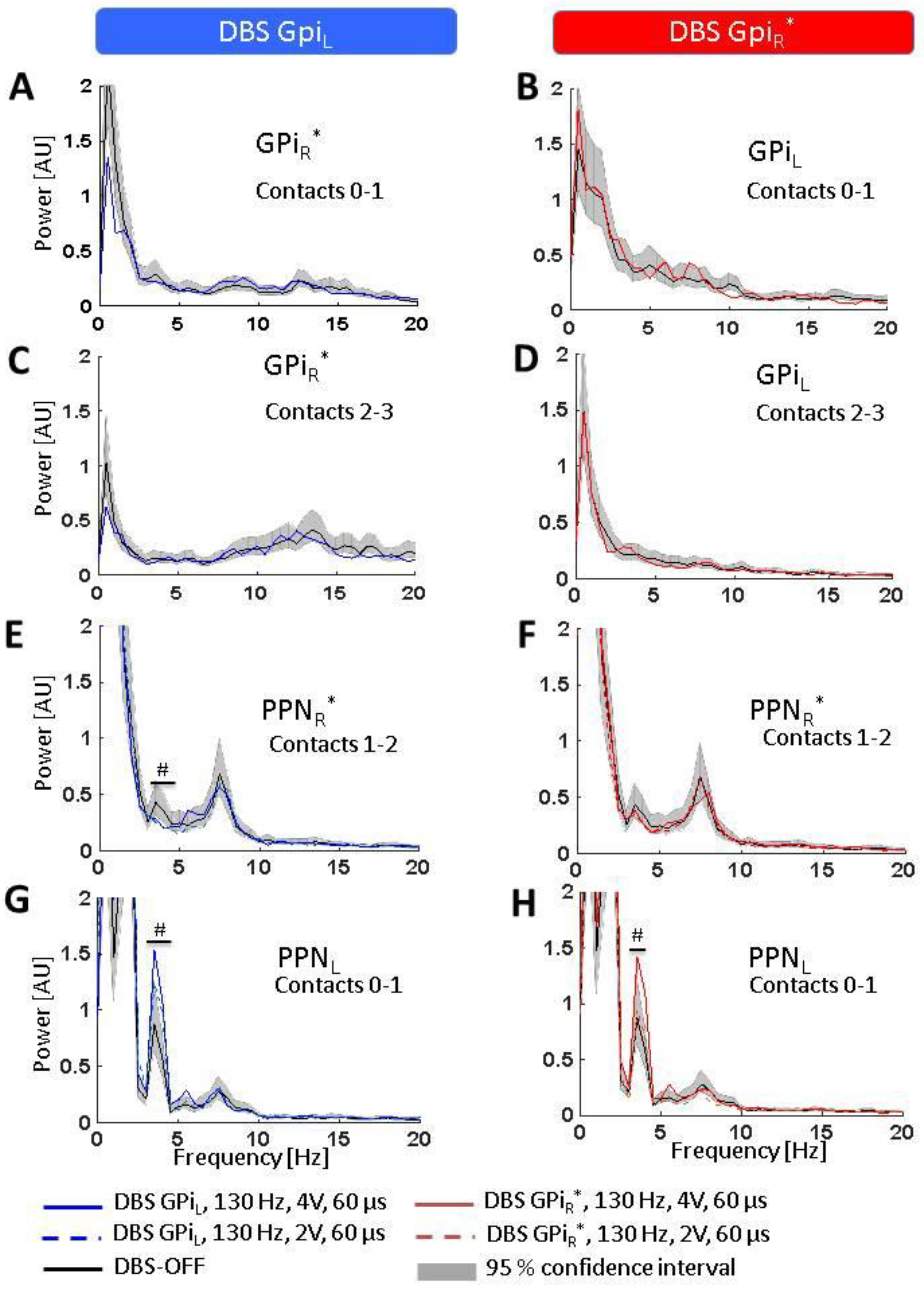
5. Results
6. Discussion
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Rahman, S.; Griffin, H.J.; Quinn, N.P.; Jahanshahi, M. Quality of life in Parkinson’s disease: The relative importance of the symptoms. Mov. Disord. 2008, 23, 1428–1434. [Google Scholar] [CrossRef] [PubMed]
- St George, R.J.; Nutt, J.G.; Burchiel, K.J.; Horak, F.B. A meta-regression of the long-term effects of deep brain stimulation on balance and gait in PD. Neurology 2010, 75, 1292–1299. [Google Scholar] [CrossRef] [PubMed]
- Benarroch, E.E. Pedunculopontine nucleus: Functional organization and clinical implications. Neurology 2013, 80, 1148–1155. [Google Scholar] [CrossRef] [PubMed]
- Stefani, A.; Lozano, A.M.; Peppe, A.; Stanzione, P.; Galati, S.; Tropepi, D.; Pierantozzi, M.; Brusa, L.; Scarnati, E.; Mazzone, P. Bilateral deep brain stimulation of the pedunculopontine and subthalamic nuclei in severe Parkinson’s disease. Brain J. Neurol. 2007, 130, 1596–1607. [Google Scholar] [CrossRef] [PubMed]
- Mazzone, P.; Insola, A.; Sposato, S.; Scarnati, E. The deep brain stimulation of the pedunculopontine tegmental nucleus. Neuromodul. J. Int. Neuromodul. Soc. 2009, 12, 191–204. [Google Scholar] [CrossRef] [PubMed]
- Moro, E.; Hamani, C.; Poon, Y.-Y.; Al-Khairallah, T.; Dostrovsky, J.O.; Hutchison, W.D.; Lozano, A.M. Unilateral pedunculopontine stimulation improves falls in Parkinson’s disease. Brain J. Neurol. 2010, 133, 215–224. [Google Scholar] [CrossRef] [PubMed]
- Ferraye, M.U.; Debu, B.; Fraix, V.; Goetz, L.; Ardouin, C.; Yelnik, J.; Henry-Lagrange, C.; Seigneuret, E.; Piallat, B.; Krack, P.; et al. Effects of pedunculopontine nucleus area stimulation on gait disorders in Parkinson’s disease. Brain J. Neurol. 2010, 133, 205–214. [Google Scholar] [CrossRef] [PubMed]
- Mazzone, P.; Scarnati, E.; Garcia-Rill, E. The Pedunculopontine Tegmental Nucleus: From Basic Neuroscience to Neurosurgical Applications Commentary: The Pedunculopontine Nucleus: Clinical Experience, Basic Questions and Future Directions. J. Neural Transm. 2011, 118, 1391–1396. [Google Scholar] [CrossRef] [PubMed]
- Hirsch, E.C.; Graybiel, A.M.; Duyckaerts, C.; Javoy-Agid, F. Neuronal loss in the pedunculopontine tegmental nucleus in Parkinson disease and in progressive supranuclear palsy. Proc. Natl. Acad. Sci. USA 1987, 84, 5976–5980. [Google Scholar] [CrossRef] [PubMed]
- Pereira, E.A.C.; Nandi, D.; Jenkinson, N.; Stein, J.F.; Green, A.L.; Aziz, T.Z. Pedunculopontine stimulation from primate to patient. J. Neural Transm. 2011, 118, 1453–1460. [Google Scholar] [CrossRef] [PubMed]
- Kuo, S.-H.; Kenney, C.; Jankovic, J. Bilateral pedunculopontine nuclei strokes presenting as freezing of gait. Mov. Disord. 2008, 23, 616–619. [Google Scholar] [CrossRef] [PubMed]
- Crossman, A.R.; Mitchell, I.J.; Sambrook, M.A. Regional brain uptake of 2-deoxyglucose in N-methyl-4-phenyl-1,2,3,6-tetrahydropyridine (MPTP)-induced parkinsonism in the macaque monkey. Neuropharmacology 1985, 24, 587–591. [Google Scholar] [CrossRef]
- Munro-Davies, L.E.; Winter, J.; Aziz, T.Z.; Stein, J.F. The role of the pedunculopontine region in basal-ganglia mechanisms of akinesia. Exp. Brain Res. 1999, 129, 511–517. [Google Scholar] [CrossRef] [PubMed]
- Stein, J.F.; Aziz, T.Z. Basal ganglia output to the PPN, a commentary. Exp. Neurol. 2012, 233, 745–746. [Google Scholar] [CrossRef] [PubMed]
- Zhang, J.; Wang, Z.I.; Baker, K.B.; Vitek, J.L. Effect of globus pallidus internus stimulation on neuronal activity in the pedunculopontine tegmental nucleus in the primate model of Parkinson’s disease. Exp. Neurol. 2012, 23, 575–580. [Google Scholar] [CrossRef] [PubMed]
- Hughes, A.J.; Daniel, S.E.; Kilford, L.; Lees, A.J. Accuracy of clinical diagnosis of idiopathic Parkinson’s disease: A clinico-pathological study of 100 cases. J. Neurol. Neurosurg. Psychiatry 1992, 55, 181–184. [Google Scholar] [CrossRef] [PubMed]
- Movement Disorder Society Task Force on Rating Scales for Parkinson’s Disease. The Unified Parkinson’s Disease Rating Scale (UPDRS): Status and recommendations. Mov. Disord. 2003, 18, 738–750. [Google Scholar]
- NICE Guidelines—National Collaborating Centre for Chronic Conditions (UK). Parkinson’s Disease: National Clinical Guideline for Diagnosis and Management in Primary and Secondary Care; Royal College of Physicians: London, UK, 2006. [Google Scholar]
- Voges, J.; Volkmann, J.; Allert, N.; Lehrke, R.; Koulousakis, A.; Freund, H.J.; Sturm, V. Bilateral high-frequency stimulation in the subthalamic nucleus for the treatment of Parkinson disease: Correlation of therapeutic effect with anatomical electrode position. J. Neurosurg. 2002, 96, 269–279. [Google Scholar] [CrossRef] [PubMed]
- Galazky, I.; Kaufmann, J.; Lorenzl, S.; Ebersbach, G.; Gandor, F.; Zaehle, T.; Specht, S.; Stallforth, S.; Sobieray, U.; Wirkus, E.; et al. Deep brain stimulation of the pedunculopontine nucleus for treatment of gait and balance disorder in progressive supranuclear palsy: Effects of frequency modulations and clinical outcome. Parkinsonism Relat. Disord. 2018, 50, 81–86. [Google Scholar] [CrossRef] [PubMed]
- Treuer, H.; Hunsche, S.; Hoevels, M.; Luyken, K.; Maarouf, M.; Voges, J.; Sturm, V. The influence of head frame distortions on stereotactic localization and targeting. Phys. Med. Biol. 2004, 49, 3877–3887. [Google Scholar] [CrossRef] [PubMed]
- Treuer, H.; Klein, D.; Maarouf, M.; Lehrke, R.; Voges, J.; Sturm, V. Accuracy and conformity of stereotactically guided interstitial brain tumour therapy using I-125 seeds. Radiother. Oncol. 2005, 77, 202–209. [Google Scholar] [CrossRef] [PubMed]
- Hunsche, S.; Sauner, D.; El Majdoub, F.; Neudorfer, C.; Poggenborg, J.; Goßmann, A.; Maarouf, M. Intensity-based 2D 3D registration for lead localization in robot guided deep brain stimulation. Phys. Med. Biol. 2017, 62, 2417–2426. [Google Scholar] [CrossRef] [PubMed]
- Defer, G.L.; Widner, H.; Marié, R.M.; Rémy, P.; Levivier, M. Core assessment program for surgical interventional therapies in Parkinson’s disease (CAPSIT-PD). Mov. Disord. 1999, 14, 572–584. [Google Scholar] [CrossRef]
- Welch, P. The use of fast Fourier transform for the estimation of power spectra: A method based on time averaging over short, modified periodograms. IEEE Trans. Audio Electroacoust. 1967, 15, 70–73. [Google Scholar] [CrossRef]
- Benjamini, Y.; Hochberg, Y. Controlling the false discovery rate: A practical and powerful approach to multiple testing. J. R. Stat. Soc. Ser. B Methodol. 1995, 57, 289–300. [Google Scholar]
- Kamitake, T.; Harashima, H.; Miyakawa, H. A time-series analysis method based on the directed transinformation. Electron. Commun. Jpn. Part I Commun. 1984, 67, 1–9. [Google Scholar] [CrossRef]
- Schreiber, T. Measuring information transfer. Phys. Rev. Lett. 2000, 85, 461–464. [Google Scholar] [CrossRef] [PubMed]
- Cao, L. Practical method for determining the minimum embedding dimension of a scalar time series. Phys. D Nonlinear Phenom. 1997, 110, 43–50. [Google Scholar] [CrossRef]
- Chen, Y.; Rangarajan, G.; Feng, J.; Ding, M. Analyzing multiple nonlinear time series with extended Granger causality. Phys. Lett. A 2004, 324, 26–35. [Google Scholar] [CrossRef]
- Jones, D.L.; Mogenson, G.J. Nucleus accumbens to globus pallidus GABA projections ubserving ambulatory activity. Am. J. Physiol. 1980, 238, R65–R69. [Google Scholar] [PubMed]
- Mogenson, G.J.; Nielsen, M. A study of the contribution of hippocampal- accumbens-subpallidal projections to locomotor activity. Behav. Neural Biol. 1984, 42, 38–51. [Google Scholar] [CrossRef]
- Swerdlow, N.R.; Koob, G.F. The neural substrates of apomorphine-stimulated locomotor activity following denervation of the nucleus accumbens. Life Sci. 1984, 35, 2537–2544. [Google Scholar] [CrossRef]
- Karachi, C.; Grabli, D.; Bernard, F.A.; Tandé, D.; Wattiez, N.; Belaid, H.; Bardinet, E.; Prigent, A.; Nothacker, H.P.; Hunot, S.; et al. Cholinergic mesencephalic neurons are involved in gait and postural disorders in Parkinson disease. J. Clin. Investig. 2010, 120, 2745–2754. [Google Scholar] [CrossRef] [PubMed]
- Nakao, N.; Ogura, M.; Nakai, K.; Itakura, T. Intrastriatal mesencephalic grafts affect neuronal activity in basal ganglia nuclei and their target structures in a rat model of Parkinson’s disease. J. Neurosci. 1998, 18, 1806–1817. [Google Scholar] [CrossRef] [PubMed]
- Thevathasan, W.; Pogosyan, A.; Hyam, J.A.; Jenkinson, N.; Foltynie, T.; Limousin, P.; Bogdanovic, M.; Zrinzo, L.; Green, A.L.; Aziz, T.Z.; et al. Alpha oscillations in the pedunculopontine nucleus correlate with gait performance in parkinsonism. Brain 2012, 135, 148–160. [Google Scholar] [CrossRef] [PubMed]
- Androulidakis, A.G.; Mazzone, P.; Litvak, V.; Penny, W.; Dileone, M.; Gaynor, L.M.; Tisch, S.; Di Lazzaro, V.; Brown, P. Oscillatory activity in the pedunculopontine area of patients with Parkinson’s disease. Exp. Neurol. 2008, 211, 59–66. [Google Scholar] [CrossRef] [PubMed]
- Tattersall, T.L.; Stratton, P.G.; Coyne, T.J.; Cook, R.; Silberstein, P.; Silburn, P.A.; Windels, F.; Sah, P. Imagined gait modulates neuronal network dynamics in the human pedunculopontine nucleus. Nat. Neurosci. 2014, 17, 449–454. [Google Scholar] [CrossRef] [PubMed]
- Pienaar, I.S.; Vernon, A.; Winn, P. The Cellular Diversity of the Pedunculopontine Nucleus: Relevance to Behavior in Health and Aspects of Parkinson’s Disease. Neuroscientist 2017, 23, 415–431. [Google Scholar] [CrossRef] [PubMed]
- Pienaar, I.S.; van de Berg, W. A non-cholinergic neuronal loss in the pedunculopontine nucleus of toxin-evoked parkinsonian rats. Exp. Neurol. 2013, 248, 213–223. [Google Scholar] [CrossRef] [PubMed]
- Zweig, R.M.; Jankel, W.R.; Hedreen, J.C.; Mayeux, R.; Price, D.L. The pedunculopontine nucleus in Parkinson’s disease. Ann. Neurol. 1989, 26, 41–46. [Google Scholar] [CrossRef] [PubMed]
- Hazrati, L.N.; Parent, A. Contralateral pallidothalamic and pallidotegmental projections in primates: An anterograde and retrograde labeling study. Brain Res. 1991, 567, 212–223. [Google Scholar] [CrossRef]
- Mena-Segovia, J.; Bolam, J.P.; Magill, P.J. Pedunculopontine nucleus and basal ganglia: Distant relatives or part of the same family? Trends Neurosci. 2004, 27, 585–588. [Google Scholar] [CrossRef] [PubMed]
- Peterson, D.S.; Fling, B.W.; Mancini, M.; Cohen, R.G.; Nutt, J.G.; Horak, F.B. Dual-task interference and brain structural connectivity in people with Parkinson’s disease who freeze. J. Neurol. Neurosurg. Psychiatry 2015, 86, 786–792. [Google Scholar] [CrossRef] [PubMed]
- Horn, A.; Reich, M.; Vorwerk, J.; Li, N.; Wenzel, G.; Fang, Q.; Schmitz-Hübsch, T.; Nickl, R.; Kupsch, A.; Volkmann, J.; et al. Connectivity predicts deep brain stimulation outcome in Parkinson disease. Ann. Neurol. 2017, 82, 67–78. [Google Scholar] [CrossRef] [PubMed]
- Hariz, M.I.; Rehncrona, S.; Quinn, N.P.; Speelman, J.D.; Wensing, C. Multicentre Advanced Parkinson’s Disease Deep Brain Stimulation Group. Multicenter study on deep brain stimulation in Parkinson’s disease: An independent assessment of reported adverse events at 4 years. Mov. Disord. 2008, 23, 416–421. [Google Scholar] [CrossRef] [PubMed]
- Schrader, C.; Capelle, H.H.; Kinfe, T.M.; Blahak, C.; Bäzner, H.; Lütjens, G.; Dressler, D.; Krauss, J.K. GPi-DBS may induce a hypokinetic gait disorder with freezing of gait in patients with dystonia. Neurology 2011, 77, 483–488. [Google Scholar] [CrossRef] [PubMed]
- Morita, H.; Hass, C.J.; Moro, E.; Sudhyadhom, A.; Kumar, R.; Okun, M.S. Pedunculopontine Nucleus Stimulation: Where are We Now and What Needs to be Done to Move the Field Forward? Front. Neurol. 2014, 5, 243. [Google Scholar] [CrossRef] [PubMed]
- Lam, S.; Moro, E.; Poon, Y.Y.; Lozano, A.M.; Fasano, A. Does dominant pedunculopontine nucleus exist? Brain 2015, 138, e323. [Google Scholar] [CrossRef] [PubMed]





© 2018 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Galazky, I.; Kluge, C.; Schmitt, F.C.; Kopitzki, K.; Zaehle, T.; Voges, J.; Büntjen, L.; Kupsch, A.; Hinrichs, H. Pallidal Stimulation Modulates Pedunculopontine Nuclei in Parkinson’s Disease. Brain Sci. 2018, 8, 117. https://doi.org/10.3390/brainsci8070117
Galazky I, Kluge C, Schmitt FC, Kopitzki K, Zaehle T, Voges J, Büntjen L, Kupsch A, Hinrichs H. Pallidal Stimulation Modulates Pedunculopontine Nuclei in Parkinson’s Disease. Brain Sciences. 2018; 8(7):117. https://doi.org/10.3390/brainsci8070117
Chicago/Turabian StyleGalazky, Imke, Christian Kluge, Friedhelm C. Schmitt, Klaus Kopitzki, Tino Zaehle, Jürgen Voges, Lars Büntjen, Andreas Kupsch, and Hermann Hinrichs. 2018. "Pallidal Stimulation Modulates Pedunculopontine Nuclei in Parkinson’s Disease" Brain Sciences 8, no. 7: 117. https://doi.org/10.3390/brainsci8070117
APA StyleGalazky, I., Kluge, C., Schmitt, F. C., Kopitzki, K., Zaehle, T., Voges, J., Büntjen, L., Kupsch, A., & Hinrichs, H. (2018). Pallidal Stimulation Modulates Pedunculopontine Nuclei in Parkinson’s Disease. Brain Sciences, 8(7), 117. https://doi.org/10.3390/brainsci8070117