Enhanced Sensitivity to Subphonemic Segments in Dyslexia: A New Instance of Allophonic Perception


Abstract
1. Introduction
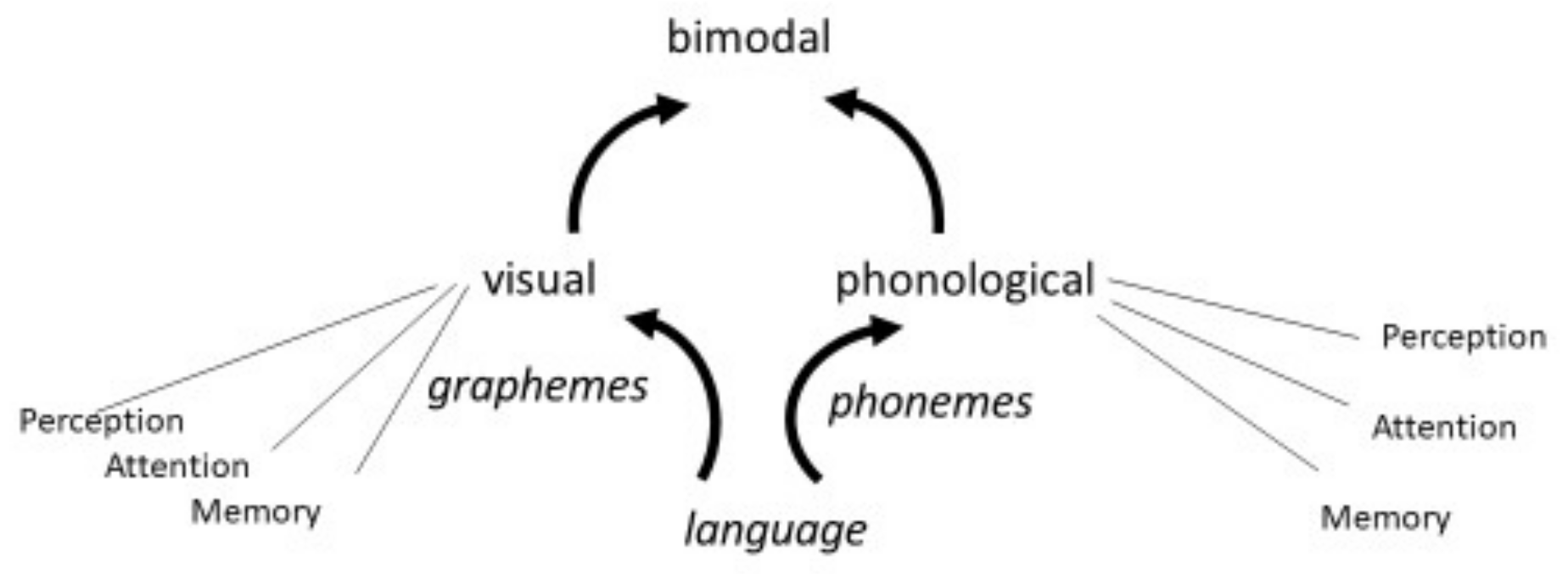
1.1. Allophonic Theory
1.2. The Present Study
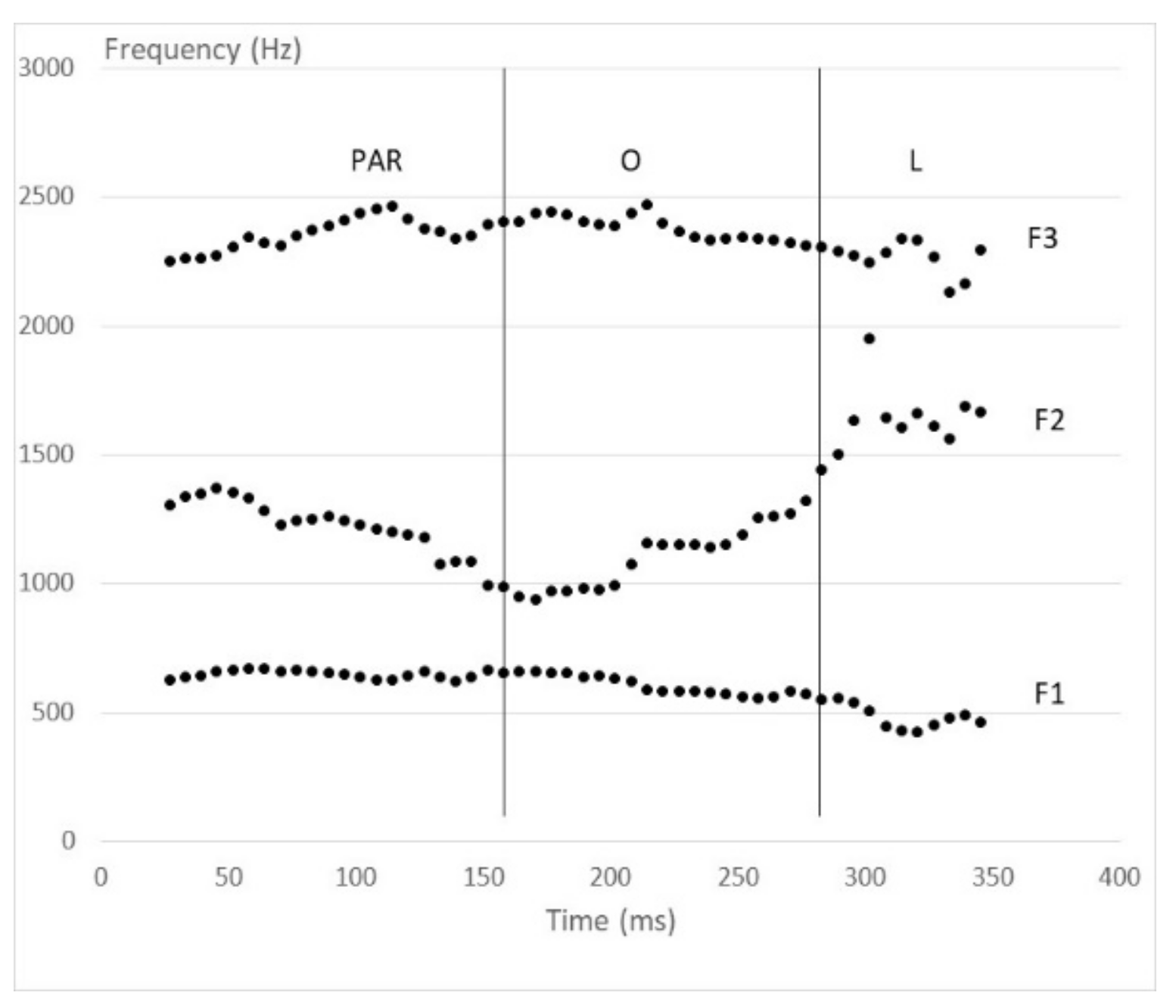
2. Materials and Methods
3. Results
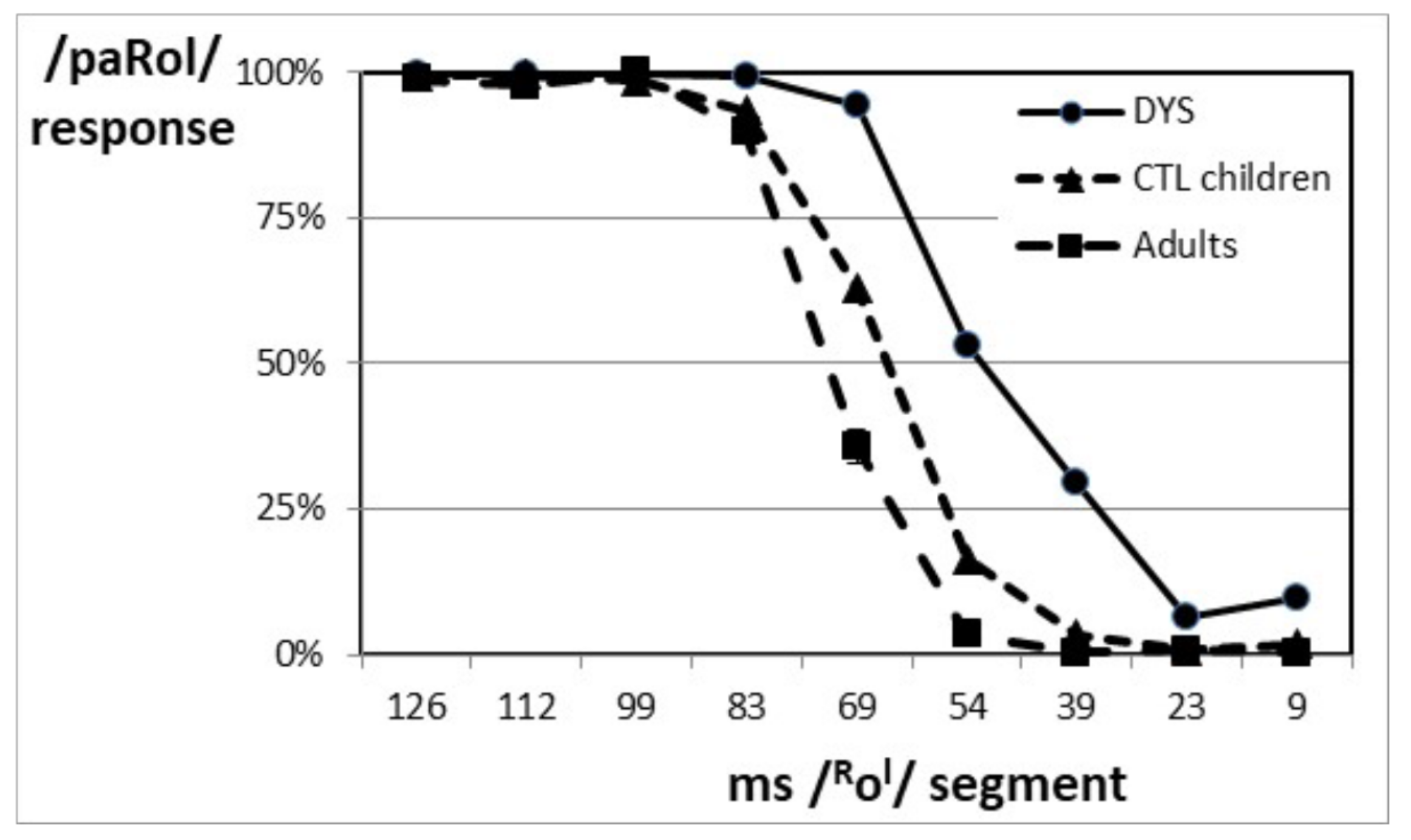
3.1. Whole Samples
3.2. Controlling the Effect of Chronological Age
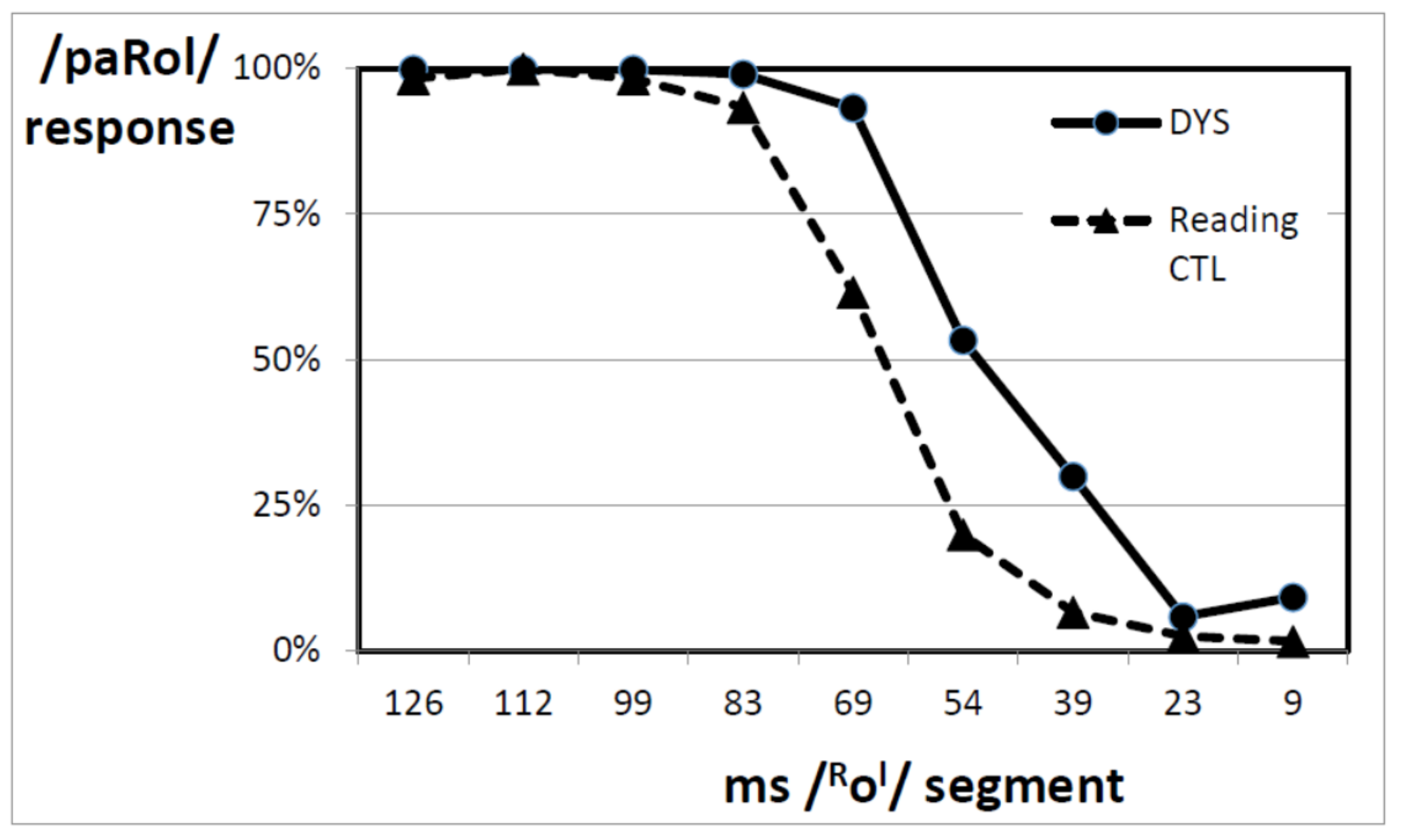
3.3. Controlling the Effect of Reading Age
4. Discussion
Theoretical Implications
5. Conclusions
Acknowledgments
Author Contributions
Conflicts of Interest
References
- Serniclaes, W.; Sprenger-Charolles, L. Reading impairment: From behavior to brain. In Handbook of Communication Disorders; Bahr, R., Silliman, E., Eds.; Routledge: London, UK, 2015; Chapter 3; pp. 34–45. [Google Scholar] [CrossRef]
- Jones, M.R. Time, our lost dimension: Toward a new theory of perception, attention, and memory. Psychol. Rev. 1976, 83, 323–355. [Google Scholar] [CrossRef] [PubMed]
- Liberman, I.; Shankweiler, D.; Fischer, F.; Carter, B. Explicit syllable and phoneme segmentation in the young child. J. Exp. Child Psychol. 1974, 18, 201–212. [Google Scholar] [CrossRef]
- Morais, J.; Bertelson, P.; Cary, L.; Alegria, J. Literacy training and speech segmentation. Cognition 1986, 24, 45–64. [Google Scholar] [CrossRef]
- Melby-Lervåg, M.; Lyster, S.A.H.; Hulme, C. Phonological skills and their role in learning to read: A meta-analytic review. Psychol. Bull. 2012, 138, 322–352. [Google Scholar] [CrossRef] [PubMed]
- Medina, V.; Hoonhorst, I.; Bogliotti, C.; Serniclaes, W. Development of voicing perception in French: Comparing adults, adolescents and children. J. Phon. 2010, 36, 493–503. [Google Scholar] [CrossRef]
- Schmidt, R. Attention, awareness, and individual differences in language learning. In Proceedings of the CLaSIC 2010, Singapore, 2–4 December 2010; Chan, W.M., Chi, S., Cin, K.N., Istanto, J., Nagami, M., Sew, J.W., Suthiwan, T., Walker, I., Eds.; National University of Singapore, Centre for Language Studies: Singapore, 2012; pp. 721–737. [Google Scholar]
- Serniclaes, W.; Ventura, P.; Morais, J.; Kolinsky, R. Categorical perception of speech sounds in illiterate adults. Cognition 2005, 98, B35–B44. [Google Scholar] [CrossRef] [PubMed]
- Boets, B.; Op de Beeck, H.P.; Vandermosten, M.; Scott, S.K.; Gillebert, C.R.; Mantini, D.; Bulthé, J.; Sunaert, S.; Wouters, J.; Ghesquière, P. Intact but less Accessible Phonetic Representations in Adults with Dyslexia. Science 2013, 6, 1251–1254. [Google Scholar] [CrossRef] [PubMed]
- Serniclaes, W.; Van Heghe, S.; Mousty, P.; Carré, R.; Sprenger-Charolles, L. Allophonic mode of speech perception in dyslexia. J. Exp. Child Psychol. 2004, 87, 336–361. [Google Scholar] [CrossRef] [PubMed]
- Werker, J.F.; Tees, R.C. Cross-language speech perception: Evidence for perceptual reorganization during the first year of life. Infant Behav. Dev. 1984, 7, 49–63. [Google Scholar] [CrossRef]
- Serniclaes, W. Features are phonological transforms of natural boundaries. In Where Do Phonological Features Come from; Clements, G.N., Ridouane, R., Eds.; John Benjamins: Amsterdam, The Netherlands, 2011; pp. 237–257. [Google Scholar]
- Hoonhorst, I.; Colin, C.; Markessis, E.; Radeau, M.; Deltenre, P.; Serniclaes, W. French native speakers in the making: From language-general to language-specific voicing boundaries. J. Exp. Child Psychol. 2009, 104, 353–366. [Google Scholar] [CrossRef] [PubMed]
- Hoonhorst, I.; Medina, V.; Colin, C.; Markessis, E.; Radeau, M.; Deltenre, P.; Serniclaes, W. The development of categorical perception: Comparisons between voicing, colors and facial expressions. Speech Commun. 2011, 53, 417–430. [Google Scholar] [CrossRef]
- Fant, G. Speech Sounds and Features; MIT Press: Cambridge, MA, USA, 1973. [Google Scholar]
- Trask, R.L. A Dictionary of Phonetics and Phonology; Routledge: London, UK, 1996. [Google Scholar]
- Boersma, P.; Weenink, D. Praat©: Doing Phonetics by Computer [Computer Program]. Available online: http://www.praat.org/ (accessed on 26 February 2018).
- Lisker, L.; Abramson, A.S. A cross-language study of voicing in initial stops: acoustical measurements. Word 1964, 20, 384–422. [Google Scholar] [CrossRef]
- Bogliotti, C.; Serniclaes, W.; Messaoud-Galusi, S.; Sprenger-Charolles, L. Discrimination of speech sounds by dyslexic children: Comparisons with chronological age and reading level controls. J. Exp. Child Psychol. 2008, 101, 137–175. [Google Scholar] [CrossRef] [PubMed]
- Noordenbos, M.W.; Segers, E.; Serniclaes, W.; Mitterer, H.; Verhoeven, L. Allophonic Mode of Speech Perception in Dutch Children at risk for Dyslexia: A Longitudinal Study. Res. Dev. Dis. 2012, 33, 1469–1483. [Google Scholar] [CrossRef] [PubMed]
- Noordenbos, M.W.; Segers, E.; Serniclaes, W.; Mitterer, H.; Verhoeven, L. Neural evidence of allophonic perception in children at risk for dyslexia. Neuropsychologia 2012, 50, 2010–2017. [Google Scholar] [CrossRef] [PubMed]
- Noordenbos, M.W.; Segers, E.; Serniclaes, W.; Verhoeven, L. Neural evidence of the allophonic mode of speech perception in adults with dyslexia. Clin. Neurophysiol. 2009, 124, 1151–1162. [Google Scholar] [CrossRef] [PubMed]
- Messaoud-Galusi, S.; Hazan, V.; Rosen, S. Investigating speech perception in children with dyslexia: Is there evidence of a consistent deficit in individuals? J. Speech Lang. Hear. Res. 2011, 54, 1682–1701. [Google Scholar] [CrossRef]
- Blomert, L. The neural signature of orthographic–phonological binding in successful and failing reading development. NeuroImage 2011, 57, 695–703. [Google Scholar] [CrossRef] [PubMed]
- Liberman, A.M.; Mattingly, I.G. The motor theory of speech perception. Cognition 1985, 21, 1–36. [Google Scholar] [CrossRef]
- Soundforge©. Sonic Foundry Inc. 2011. Available online: http://www.sonicfoundry.com (accessed on 26 February 2018).
- Matlab©. Version 6.1. 2011. Available online: https://fr.mathworks.com/products/matlab.html (accessed on 26 February 2018).
- Lefavrais, P. Test de l’Alouette (’Alouette’ Reading Test); Editions du Centre de Psychologie Appliquée: Paris, France, 1965. [Google Scholar]
- IBM SPSS© Statistics for Windows, version 24.0. 2017. Available online: https://www.ibm.com/analytics/fr/fr/technology/spss/ (accessed on 26 February 2018).
- Burnham, D. Language specific speech perception and the onset of reading. Read. Writ. 2003, 16, 573–609. [Google Scholar] [CrossRef]
- Goswami, U. A temporal sampling framework for developmental dyslexia. Trends Cogn. Sci. 2011, 15, 1–10. [Google Scholar] [CrossRef] [PubMed]
- Goswami, U. Sensory theories of developmental dyslexia: Three challenges for research. Nat. Rev. Neurosci. 2015, 16, 43–54. [Google Scholar] [CrossRef] [PubMed]
- Giraud, A.L.; Ramus, F. Neurogenetics and auditory pocessing in developmental dyslexia. Cur. Opin. Neurobiol. 2013, 23, 37–42. [Google Scholar] [CrossRef] [PubMed]
- Hämäläinen, J.; Landi, N.; Loberg, O.; Lohvansuu, K.; Pugh, K.; Leppänen, P.H.T. Brain event-related potentials to phoneme contrasts and their correlation to reading skills in school-age children. Int. J. Behav. Dev. 2017. [Google Scholar] [CrossRef]
- Noordenbos, M.; Serniclaes, W. Categorical perception in dyslexia: A meta-analysis. Sci. Stud. Read. 2015, 19, 340–359. [Google Scholar] [CrossRef]
- Zoubrinetzky, R.; Collet, G.; Serniclaes, W.; Nguyen-Morel, M.-A.; Valdois, S. Relationships between categorical perception of phonemes, phoneme awareness, and visual attention span in developmental dyslexia. PLoS ONE 2016, 11. [Google Scholar] [CrossRef] [PubMed]
- Hakvoort, B.; de Bree, E.; van der Leij, A.; Maassen, B.; van Setten, E.; Maurits, N.; van Zuijen, T.L. The Role of Categorical Speech Perception and Phonological Processing in Familial Risk Children With and Without Dyslexia. J. Speech Lang. Hear. Res. 2016, 59, 1448–1460. [Google Scholar] [CrossRef] [PubMed]
- Ziegler, J.C.; Bertrand, D.; Toth, D.; Csepe, V.; Reis, A.; Faisca, L.; Saine, N.; Lyytinen, H.; Vaessen, A.; Blomert, L. Orthographic Depth and Its Impact on Universal Predictors of Reading: A Cross-Language Investigation. Psychol. Sci. 2010, 21, 551–559. [Google Scholar] [CrossRef] [PubMed]
- Landerl, K.; Wimmer, H.; Frith, U. The impact of orthographic consistency on dyslexia: A German-English comparison. Cognition 1997, 63, 315–334. [Google Scholar] [CrossRef]
- Dufor, O.; Serniclaes, W.; Sprenger-Charolles, L.; Démonet, J.-F. Left pre-motor cortex and allophonic speech perception in dyslexia: A PET study. NeuroImage 2009, 46, 241–248. [Google Scholar] [CrossRef] [PubMed][Green Version]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Full Samples | Chronological Age Mean (SD) | Reading Age Mean (SD) | Reading Delay Mean (SD) |
|---|---|---|---|
| CTL (N = 52) | 7.60 years (0.36) | 8.65 years (1.23) | +1.06 years (1.18) |
| DYS (N = 19) | 9.55 years (1.51) | 6.68 years (0.61) | −2.87 years (1.15) |
| Difference | F(1,69) = 77.4 (p < 0.001) | F(1,69) = 43.9 (p < 0.001) | F(1,69) = 155 (p < 0.001) |
| Matched Samples | Chronological Age Mean (SD) | Reading Age Mean (SD) | Reading Delay Mean (SD) |
|---|---|---|---|
| Reading Age CTL (N = 12) | 7.42 years (0.29) | 7.20 years (0.29) | −0.22 years (0.39) |
| DYS (N = 12) | 10.3 years (1.11) | 7.02 years (0.51) | −3.31 years (0.96) |
| Difference | F(1,22) = 77.0 (p < 0.001) | F(1,22) = 1.12 (p = 0.30) | F(1,22) = 107 (p < 0.001) |
© 2018 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Serniclaes, W.; Seck, M. Enhanced Sensitivity to Subphonemic Segments in Dyslexia: A New Instance of Allophonic Perception. Brain Sci. 2018, 8, 54. https://doi.org/10.3390/brainsci8040054
Serniclaes W, Seck M. Enhanced Sensitivity to Subphonemic Segments in Dyslexia: A New Instance of Allophonic Perception. Brain Sciences. 2018; 8(4):54. https://doi.org/10.3390/brainsci8040054
Chicago/Turabian StyleSerniclaes, Willy, and M’ballo Seck. 2018. "Enhanced Sensitivity to Subphonemic Segments in Dyslexia: A New Instance of Allophonic Perception" Brain Sciences 8, no. 4: 54. https://doi.org/10.3390/brainsci8040054
APA StyleSerniclaes, W., & Seck, M. (2018). Enhanced Sensitivity to Subphonemic Segments in Dyslexia: A New Instance of Allophonic Perception. Brain Sciences, 8(4), 54. https://doi.org/10.3390/brainsci8040054