Deep Brain Stimulation—Possible Treatment Strategy for Pathologically Altered Body Weight?


Abstract
:1. Introduction
2. Effects of DBS on Food Intake and Body Weight Gain
3. Potential Application of DBS in Obesity
4. Potential Application of DBS in Binge-Eating Disorder
5. Potential Application of DBS in Anorexia Nervosa
6. Challenges of DBS in the Treatment of Eating Disorders
7. Conclusions
Acknowledgments
Author Contributions
Conflicts of Interest
References
- Ng, M.; Fleming, T.; Robinson, M.; Thomson, B.; Graetz, N.; Margono, C.; Mullany, E.C.; Biryukov, S.; Abbafati, C.; Abera, S.F.; et al. Global, regional, and national prevalence of overweight and obesity in children and adults during 1980–2013: A systematic analysis for the global burden of disease study 2013. Lancet 2014, 384, 766–781. [Google Scholar] [CrossRef]
- James, P.T.; Leach, R.; Kalamara, E.; Shayeghi, M. The worldwide obesity epidemic. Obes. Res. 2001, 9 (Suppl. 4), 228S–233S. [Google Scholar] [CrossRef] [PubMed]
- Lopresti, A.L.; Drummond, P.D. Obesity and psychiatric disorders: Commonalities in dysregulated biological pathways and their implications for treatment. Prog. Neuropsychopharmacol. Biol. Psychiatry 2013, 45, 92–99. [Google Scholar] [CrossRef] [PubMed]
- Yoshimoto, S.; Loo, T.M.; Atarashi, K.; Kanda, H.; Sato, S.; Oyadomari, S.; Iwakura, Y.; Oshima, K.; Morita, H.; Hattori, M.; et al. Obesity-induced gut microbial metabolite promotes liver cancer through senescence secretome. Nature 2013, 499, 97–101. [Google Scholar] [CrossRef] [PubMed]
- Wyatt, H.R. Update on treatment strategies for obesity. J. Clin. Endocrinol. Metab. 2013, 98, 1299–1306. [Google Scholar] [CrossRef] [PubMed]
- Valsamakis, G.; Konstantakou, P.; Mastorakos, G. New targets for drug treatment of obesity. Annu. Rev. Pharmacol. Toxicol. 2017, 57, 585–605. [Google Scholar] [CrossRef] [PubMed]
- Schauer, P.R.; Bhatt, D.L.; Kirwan, J.P.; Wolski, K.; Aminian, A.; Brethauer, S.A.; Navaneethan, S.D.; Singh, R.P.; Pothier, C.E.; Nissen, S.E.; et al. Bariatric surgery versus intensive medical therapy for diabetes—5-year outcomes. N. Engl. J. Med. 2017, 376, 641–651. [Google Scholar] [CrossRef] [PubMed]
- Mustelin, L.; Bulik, C.M.; Kaprio, J.; Keski-Rahkonen, A. Prevalence and correlates of binge eating disorder related features in the community. Appetite 2017, 109, 165–171. [Google Scholar] [CrossRef] [PubMed]
- Hutson, P.H.; Balodis, I.M.; Potenza, M.N. Binge-eating disorder: Clinical and therapeutic advances. Pharmacol. Ther. 2017. [CrossRef] [PubMed]
- Treasure, J.; Zipfel, S.; Micali, N.; Wade, T.; Stice, E.; Claudino, A.; Schmidt, U.; Frank, G.K.; Bulik, C.M.; Wentz, E. Anorexia nervosa. Nat. Rev. Dis. Prim. 2015, 1, 15074. [Google Scholar] [CrossRef] [PubMed]
- American Psychiatric Association. Diagnostic and Statistical Manual of Mental Disorders, 5th ed.; American Psychiatric Association Publishing: Arlington, VA, USA, 2013. [Google Scholar]
- Zipfel, S.; Wild, B.; Gross, G.; Friederich, H.C.; Teufel, M.; Schellberg, D.; Giel, K.E.; de Zwaan, M.; Dinkel, A.; Herpertz, S.; et al. Focal psychodynamic therapy, cognitive behaviour therapy, and optimised treatment as usual in outpatients with anorexia nervosa (ANTOP study): Randomised controlled trial. Lancet 2014, 383, 127–137. [Google Scholar] [CrossRef]
- Nagl, M.; Jacobi, C.; Paul, M.; Beesdo-Baum, K.; Hofler, M.; Lieb, R.; Wittchen, H.U. Prevalence, incidence, and natural course of anorexia and bulimia nervosa among adolescents and young adults. Eur. Child Adolesc. Psychiatry 2016, 25, 903–918. [Google Scholar] [CrossRef] [PubMed]
- Zipfel, S.; Giel, K.E.; Bulik, C.M.; Hay, P.; Schmidt, U. Anorexia nervosa: Aetiology, assessment, and treatment, Lancet. Psychiatry 2015, 2, 1099–1111. [Google Scholar] [CrossRef]
- Delgado, J.M.; Hamlin, H.; Chapman, W.P. Technique of intracranial electrode implacement for recording and stimulation and its possible therapeutic value in psychotic patients. Confin. Neurol. 1952, 12, 315–319. [Google Scholar] [CrossRef] [PubMed]
- Bekhtereva, N.P.; Grachev, K.V.; Orlova, A.N.; Iatsuksl. Utilization of multiple electrodes implanted in the subcortical structure of the human brain for the treatment of hyperkinesis. Zh. Nevropatol. Psikhiatr. Im. S. S. Korsakova 1963, 63, 3–8. [Google Scholar] [PubMed]
- Sem-Jacobsen, C.W. Depth electrographic stimulation and treatment of patients with Parkinson’s disease including neurosurgical technique. Acta Neurol. Scand. Suppl. 1965, 1 Pt 13, 365–377. [Google Scholar] [CrossRef]
- Hosobuchi, Y.; Adams, J.E.; Rutkin, B. Chronic thalamic stimulation for the control of facial anesthesia dolorosa. Arch Neurol. 1973, 29, 158–161. [Google Scholar] [CrossRef] [PubMed]
- Hassler, R.; Ore, G.D.; Bricolo, A.; Dieckmann, G.; Dolce, G. EEG and clinical arousal induced by bilateral long-term stimulation of pallidal systems in traumatic vigil coma. Electroencephalogr. Clin. Neurophysiol. 1969, 27, 689–690. [Google Scholar] [PubMed]
- Riklan, M.; Cullinan, T.; Shulman, M.; Cooper, I.S. A psychometric study of chronic cerebellar stimulation in man. Biol. Psychiatry 1976, 11, 543–574. [Google Scholar] [PubMed]
- Martinez-Ramirez, D.; Hu, W.; Bona, A.R.; Okun, M.S.; Shukla, A.W. Update on deep brain stimulation in Parkinson’s disease. Transl. Neurodegener. 2015, 4, 12. [Google Scholar] [CrossRef] [PubMed]
- Benabid, A.L.; Chabardes, S.; Mitrofanis, J.; Pollak, P. Deep brain stimulation of the subthalamic nucleus for the treatment of Parkinson’s disease. Lancet Neurol. 2009, 8, 67–81. [Google Scholar] [CrossRef]
- Odekerken, V.J.; Boel, J.A.; Schmand, B.A.; de Haan, R.J.; Figee, M.; van den Munckhof, P.; Schuurman, P.R.; de Bie, R.M.; NSTAPS Study Group. GPi vs. STN deep brain stimulation for Parkinson disease: Three-year follow-up. Neurology 2016, 86, 755–761. [Google Scholar] [CrossRef] [PubMed]
- Lizarraga, K.J.; Jagid, J.R.; Luca, C.C. Comparative effects of unilateral and bilateral subthalamic nucleus deep brain stimulation on gait kinematics in Parkinson’s disease: A randomized, blinded study. J. Neurol. 2016, 263, 1652–1656. [Google Scholar] [CrossRef] [PubMed]
- Martinez-Fernandez, R.; Zrinzo, L.; Aviles-Olmos, I.; Hariz, M.; Martinez-Torres, I.; Joyce, E.; Jahanshahi, M.; Limousin, P.; Foltynie, T. Deep brain stimulation for Gilles de la Tourette syndrome: A case series targeting subregions of the globus pallidus internus. Mov. Disord. 2011, 26, 1922–1930. [Google Scholar] [CrossRef] [PubMed]
- Huys, D.; Bartsch, C.; Koester, P.; Lenartz, D.; Maarouf, M.; Daumann, J.; Mai, J.K.; Klosterkotter, J.; Hunsche, S.; Visser-Vandewalle, V.; et al. Motor improvement and emotional stabilization in patients with Tourette syndrome after deep brain stimulation of the ventral anterior and ventrolateral motor part of the thalamus. Biol. Psychiatry 2016, 79, 392–401. [Google Scholar] [CrossRef] [PubMed]
- Lega, B.C.; Halpern, C.H.; Jaggi, J.L.; Baltuch, G.H. Deep brain stimulation in the treatment of refractory epilepsy: Update on current data and future directions. Neurobiol. Dis. 2010, 38, 354–360. [Google Scholar] [CrossRef] [PubMed]
- Salanova, V.; Witt, T.; Worth, R.; Henry, T.R.; Gross, R.E.; Nazzaro, J.M.; Labar, D.; Sperling, M.R.; Sharan, A.; Sandok, E.; et al. Long-term efficacy and safety of thalamic stimulation for drug-resistant partial epilepsy. Neurology 2015, 84, 1017–1025. [Google Scholar] [CrossRef] [PubMed]
- Schlaepfer, T.E.; Cohen, M.X.; Frick, C.; Kosel, M.; Brodesser, D.; Axmacher, N.; Joe, A.Y.; Kreft, M.; Lenartz, D.; Sturm, V. Deep brain stimulation to reward circuitry alleviates anhedonia in refractory major depression. Neuropsychopharmacology 2008, 33, 368–377. [Google Scholar] [CrossRef] [PubMed]
- Bergfeld, I.O.; Mantione, M.; Hoogendoorn, M.L.; Ruhe, H.G.; Notten, P.; van Laarhoven, J.; Visser, I.; Figee, M.; de Kwaasteniet, B.P.; Horst, F.; et al. Deep brain stimulation of the ventral anterior limb of the internal capsule for treatment-resistant depression: A randomized clinical trial. JAMA Psychiatry 2016, 73, 456–464. [Google Scholar] [CrossRef] [PubMed]
- Whiting, D.M.; Tomycz, N.D.; Bailes, J.; de Jonge, L.; Lecoultre, V.; Wilent, B.; Alcindor, D.; Prostko, E.R.; Cheng, B.C.; Angle, C.; et al. Lateral hypothalamic area deep brain stimulation for refractory obesity: A pilot study with preliminary data on safety, body weight, and energy metabolism. J. Neurosurg. 2013, 119, 56–63. [Google Scholar] [CrossRef] [PubMed]
- Hamani, C.; McAndrews, M.P.; Cohn, M.; Oh, M.; Zumsteg, D.; Shapiro, C.M.; Wennberg, R.A.; Lozano, A.M. Memory enhancement induced by hypothalamic/fornix deep brain stimulation. Ann. Neurol. 2008, 63, 119–123. [Google Scholar] [CrossRef] [PubMed]
- Lipsman, N.; Lam, E.; Volpini, M.; Sutandar, K.; Twose, R.; Giacobbe, P.; Sodums, D.J.; Smith, G.S.; Woodside, D.B.; Lozano, A.M. Deep brain stimulation of the subcallosal cingulate for treatment-refractory anorexia nervosa: 1 year follow-up of an open-label trial. Lancet Psychiatry 2017, 4, 285–294. [Google Scholar] [CrossRef]
- Prinz, P.; Kobelt, P.; Scharner, S.; Goebel-Stengel, M.; Harnack, D.; Faust, K.; Winter, Y.; Rose, M.; Stengel, A. Deep brain stimulation alters light phase food intake microstructure in rats. J. Physiol. Pharmacol. 2017, 68, 345–354. [Google Scholar] [PubMed]
- Zhang, C.; Wei, N.L.; Wang, Y.; Wang, X.; Zhang, J.G.; Zhang, K. Deep brain stimulation of the nucleus accumbens shell induces anti-obesity effects in obese rats with alteration of dopamine neurotransmission. Neurosci. Lett. 2015, 589, 1–6. [Google Scholar] [CrossRef] [PubMed]
- van der Plasse, G.; Schrama, R.; van Seters, S.P.; Vanderschuren, L.J.; Westenberg, H.G. Deep brain stimulation reveals a dissociation of consummatory and motivated behaviour in the medial and lateral nucleus accumbens shell of the rat. PLoS ONE 2012, 7, e33455. [Google Scholar] [CrossRef] [PubMed]
- Soto-Montenegro, M.L.; Pascau, J.; Desco, M. Response to deep brain stimulation in the lateral hypothalamic area in a rat model of obesity: In vivo assessment of brain glucose metabolism. Mol. Imaging Biol. 2014, 16, 830–837. [Google Scholar] [CrossRef] [PubMed]
- Lehmkuhle, M.J.; Mayes, S.M.; Kipke, D.R. Unilateral neuromodulation of the ventromedial hypothalamus of the rat through deep brain stimulation. J. Neural Eng. 2010, 7, 036006. [Google Scholar] [CrossRef] [PubMed]
- Melega, W.P.; Lacan, G.; Gorgulho, A.A.; Behnke, E.J.; de Salles, A.A. Hypothalamic deep brain stimulation reduces weight gain in an obesity-animal model. PLoS ONE 2012, 7, e30672. [Google Scholar] [CrossRef] [PubMed]
- Lacan, G.; de Salles, A.A.; Gorgulho, A.A.; Krahl, S.E.; Frighetto, L.; Behnke, E.J.; Melega, W.P. Modulation of food intake following deep brain stimulation of the ventromedial hypothalamus in the vervet monkey. Laboratory investigation. J. Neurosurg. 2008, 108, 336–342. [Google Scholar] [PubMed]
- Lipsman, N.; Woodside, D.B.; Giacobbe, P.; Hamani, C.; Carter, J.C.; Norwood, S.J.; Sutandar, K.; Staab, R.; Elias, G.; Lyman, C.H.; et al. Subcallosal cingulate deep brain stimulation for treatment-refractory anorexia nervosa: A phase 1 pilot trial. Lancet 2013, 381, 1361–1370. [Google Scholar] [CrossRef]
- Prinz, P.; Stengel, A. Control of food intake by gastrointestinal peptides: Mechanisms of action and possible modulation in the treatment of obesity. J. Neurogastroenterol. Motil. 2017, 23, 180–196. [Google Scholar] [CrossRef] [PubMed]
- Sauleau, P.; Drapier, S.; Duprez, J.; Houvenaghel, J.F.; Dondaine, T.; Haegelen, C.; Drapier, D.; Jannin, P.; Robert, G.; Le Jeune, F.; et al. Weight gain following pallidal deep brain stimulation: A PET study. PLoS ONE 2016, 11, e0153438. [Google Scholar] [CrossRef] [PubMed]
- Aiello, M.; Eleopra, R.; Foroni, F.; Rinaldo, S.; Rumiati, R.I. Weight gain after STN-DBS: The role of reward sensitivity and impulsivity. Cortex 2017, 92, 150–161. [Google Scholar] [CrossRef] [PubMed]
- Millan, S.H.; Hacker, M.L.; Turchan, M.; Molinari, A.L.; Currie, A.D.; Charles, D. Subthalamic nucleus deep brain stimulation in early stage Parkinson's disease is not associated with increased body mass index. Parkinsons Dis. 2017, 2017, 7163801. [Google Scholar] [CrossRef] [PubMed]
- Melse, M.; Temel, Y.; Tan, S.K.; Jahanshahi, A. Deep brain stimulation of the rostromedial tegmental nucleus: An unanticipated, selective effect on food intake. Brain Res. Bull. 2016, 127, 23–28. [Google Scholar] [CrossRef] [PubMed]
- Kelley, A.E.; Swanson, C.J. Feeding induced by blockade of AMPA and kainate receptors within the ventral striatum: A microinfusion mapping study. Behav. Brain Res. 1997, 89, 107–113. [Google Scholar] [CrossRef]
- Rosenwasser, A.M.; Boulos, Z.; Terman, M. Circadian organization of food intake and meal patterns in the rat. Physiol. Behav. 1981, 27, 33–39. [Google Scholar] [CrossRef]
- Lean, M.E.J.; Malkova, D. Altered gut and adipose tissue hormones in overweight and obese individuals: Cause or consequence? Int. J. Obes. 2016, 40, 622–632. [Google Scholar] [CrossRef] [PubMed]
- Halpern, C.H.; Tekriwal, A.; Santollo, J.; Keating, J.G.; Wolf, J.A.; Daniels, D.; Bale, T.L. Amelioration of binge eating by nucleus accumbens shell deep brain stimulation in mice involves D2 receptor modulation. J. Neurosci. 2013, 33, 7122–7129. [Google Scholar] [CrossRef] [PubMed]
- Dunn, J.P.; Cowan, R.L.; Volkow, N.D.; Feurer, I.D.; Li, R.; Williams, D.B.; Kessler, R.M.; Abumrad, N.N. Decreased dopamine type 2 receptor availability after bariatric surgery: Preliminary findings. Brain Res. 2010, 1350, 123–130. [Google Scholar] [CrossRef] [PubMed]
- Doucette, W.T.; Khokhar, J.Y.; Green, A.I. Nucleus accumbens deep brain stimulation in a rat model of binge eating. Transl. Psychiatry 2015, 5, e695. [Google Scholar] [CrossRef] [PubMed]
- Sani, S.; Jobe, K.; Smith, A.; Kordower, J.H.; Bakay, R.A. Deep brain stimulation for treatment of obesity in rats. J. Neurosurg. 2007, 107, 809–813. [Google Scholar] [CrossRef] [PubMed]
- Torres, N.; Chabardes, S.; Piallat, B.; Devergnas, A.; Benabid, A.L. Body fat and body weight reduction following hypothalamic deep brain stimulation in monkeys: An intraventricular approach. Int. J. Obes. 2012, 36, 1537–1544. [Google Scholar] [CrossRef] [PubMed]
- Brown, F.D.; Fessler, R.G.; Rachlin, J.R.; Mullan, S. Changes in food intake with electrical stimulation of the ventromedial hypothalamus in dogs. J. Neurosurg. 1984, 60, 1253–1257. [Google Scholar] [CrossRef] [PubMed]
- Ross, S.E.; Levin, E.L.; Itoga, C.A.; Schoen, C.B.; Selmane, R.; Aldridge, J.W. Deep brain stimulation in the central nucleus of the amygdala decreases ‘wanting’ and ‘liking’ of food rewards. Eur. J. Neurosci. 2016, 44, 2431–2445. [Google Scholar] [CrossRef] [PubMed]
- Mills, K.A.; Scherzer, R.; Starr, P.A.; Ostrem, J.L. Weight change after globus pallidus internus or subthalamic nucleus deep brain stimulation in Parkinson’s disease and dystonia. Stereotact. Funct. Neurosurg. 2012, 90, 386–393. [Google Scholar] [CrossRef] [PubMed]
- Barichella, M.; Marczewska, A.M.; Mariani, C.; Landi, A.; Vairo, A.; Pezzoli, G. Body weight gain rate in patients with Parkinson's disease and deep brain stimulation. Mov. Disord. 2003, 18, 1337–1340. [Google Scholar] [CrossRef] [PubMed]
- Novakova, L.; Ruzicka, E.; Jech, R.; Serranova, T.; Dusek, P.; Urgosik, D. Increase in body weight is a non-motor side effect of deep brain stimulation of the subthalamic nucleus in Parkinson’s disease. Neuro Endocrinol. Lett. 2007, 28, 21–25. [Google Scholar] [PubMed]
- Mantione, M.; van de Brink, W.; Schuurman, P.R.; Denys, D. Smoking cessation and weight loss after chronic deep brain stimulation of the nucleus accumbens: Therapeutic and research implications: Case report. Neurosurgery 2010, 66, E218. [Google Scholar] [CrossRef] [PubMed]
- Harat, M.; Rudas, M.; Zielinski, P.; Birska, J.; Sokal, P. Nucleus accumbens stimulation in pathological obesity. Neurol. Neurochir. Pol. 2016, 50, 207–210. [Google Scholar] [CrossRef] [PubMed]
- Israël, M.; Steiger, H.; Kolivakis, T.; McGregor, L.; Sadikot, A.F. Deep brain stimulation in the subgenual cingulate cortex for an intractable eating disorder. Biol. Psychiatry 2010, 67, e53–e54. [Google Scholar] [CrossRef] [PubMed]
- McLaughlin, N.C.; Didie, E.R.; Machado, A.G.; Haber, S.N.; Eskandar, E.N.; Greenberg, B.D. Improvements in anorexia symptoms after deep brain stimulation for intractable obsessive-compulsive disorder. Biol. Psychiatry 2013, 73, e29–e31. [Google Scholar] [CrossRef] [PubMed]
- Wu, H.; van Dyck-Lippens, P.J.; Santegoeds, R.; van Kuyck, K.; Gabriels, L.; Lin, G.; Pan, G.; Li, Y.; Li, D.; Zhan, S.; et al. Deep-brain stimulation for anorexia nervosa. World Neurosurg. 2013, 80, S29.e1–S29.e10. [Google Scholar] [CrossRef] [PubMed]
- Wang, J.; Chang, C.; Geng, N.; Wang, X.; Gao, G. Treatment of intractable anorexia nervosa with inactivation of the nucleus accumbens using stereotactic surgery. Stereotact. Funct. Neurosurg. 2013, 91, 364–372. [Google Scholar] [CrossRef] [PubMed]
- Blomstedt, P.; Naesström, M.; Bodlund, O. Deep brain stimulation in the bed nucleus of the stria terminalis and medial forebrain bundle in a patient with major depressive disorder and anorexia nervosa. Clin. Case Rep. 2017, 5, 679–684. [Google Scholar] [CrossRef] [PubMed]
- Moro, E.; Scerrati, M.; Romito, L.M.; Roselli, R.; Tonali, P.; Albanese, A. Chronic subthalamic nucleus stimulation reduces medication requirements in Parkinson’s disease. Neurology 1999, 53, 85–90. [Google Scholar] [CrossRef] [PubMed]
- Routtenberg, A.; Kuznesof, A.W. Self-starvation of rats living in activity wheels on a restricted feeding schedule. J. Comp. Physiol. Psychol. 1967, 64, 414–421. [Google Scholar] [CrossRef] [PubMed]
- Dietz, J.; Noecker, A.M.; McIntyre, C.C.; Mikos, A.; Bowers, D.; Foote, K.D.; Okun, M.S. Stimulation region within the globus pallidus does not affect verbal fluency performance. Brain Stimul. 2013, 6, 248–253. [Google Scholar] [CrossRef] [PubMed]
- Liu, Y.; Li, W.; Tan, C.; Liu, X.; Wang, X.; Gui, Y.; Qin, L.; Deng, F.; Hu, C.; Chen, L. Meta-analysis comparing deep brain stimulation of the globus pallidus and subthalamic nucleus to treat advanced Parkinson disease. J. Neurosurg. 2014, 121, 709–718. [Google Scholar] [CrossRef] [PubMed]
- Combs, H.L.; Folley, B.S.; Berry, D.T.; Segerstrom, S.C.; Han, D.Y.; Anderson-Mooney, A.J.; Walls, B.D.; van Horne, C. Cognition and depression following deep brain stimulation of the subthalamic nucleus and globus pallidus pars internus in Parkinson’s disease: A meta-analysis. Neuropsychol. Rev. 2015, 25, 439–454. [Google Scholar] [CrossRef] [PubMed]
- Okun, M.S.; Fernandez, H.H.; Wu, S.S.; Kirsch-Darrow, L.; Bowers, D.; Bova, F.; Suelter, M.; Jacobson, C.E.T.; Wang, X.; Gordon, C.W., Jr.; et al. Cognition and mood in Parkinson’s disease in subthalamic nucleus versus globus pallidus interna deep brain stimulation: The COMPARE trial. Ann. Neurol. 2009, 65, 586–595. [Google Scholar] [CrossRef] [PubMed]
- Groiss, S.J.; Wojtecki, L.; Sudmeyer, M.; Schnitzler, A. Deep brain stimulation in Parkinson’s disease. Ther. Adv. Neurol. Disord. 2009, 2, 20–28. [Google Scholar] [CrossRef] [PubMed]
- Péron, J.; Fruhholz, S.; Verin, M.; Grandjean, D. Subthalamic nucleus: A key structure for emotional component synchronization in humans. Neurosci. Biobehav. Rev. 2013, 37, 358–373. [Google Scholar] [CrossRef] [PubMed]
- Smeding, H.M.; Speelman, J.D.; Koning-Haanstra, M.; Schuurman, P.R.; Nijssen, P.; van Laar, T.; Schmand, B. Neuropsychological effects of bilateral STN stimulation in Parkinson disease: A controlled study. Neurology 2006, 66, 1830–1836. [Google Scholar] [CrossRef] [PubMed]
- Voon, V.; Krack, P.; Lang, A.E.; Lozano, A.M.; Dujardin, K.; Schupbach, M.; D’Ambrosia, J.; Thobois, S.; Tamma, F.; Herzog, J.; et al. A multicentre study on suicide outcomes following subthalamic stimulation for Parkinson’s disease. Brain 2008, 131 Pt 10, 2720–2728. [Google Scholar] [CrossRef]
- Julien, C.L.; Rimes, K.A.; Brown, R.G. Rumination and behavioural factors in Parkinson’s disease depression. J. Psychosom. Res. 2016, 82, 48–53. [Google Scholar] [CrossRef] [PubMed]
- Volkow, N.D.; Wang, G.J.; Baler, R.D. Reward, dopamine and the control of food intake: Implications for obesity. Trends Cogn. Sci. 2011, 15, 37–46. [Google Scholar] [CrossRef] [PubMed]
- Russo, S.J.; Nestler, E.J. The brain reward circuitry in mood disorders. Nat. Rev. Neurosci. 2013, 14, 609–625. [Google Scholar] [CrossRef] [PubMed]
- Ho, A.L.; Sussman, E.S.; Zhang, M.; Pendharkar, A.V.; Azagury, D.E.; Bohon, C.; Halpern, C.H. Deep brain stimulation for obesity. Cureus 2015, 7, e259. [Google Scholar] [CrossRef] [PubMed]
- Park, R.J.; Singh, I.; Pike, A.C.; Tan, J.O. Deep brain stimulation in anorexia nervosa: Hope for the hopeless or exploitation of the vulnerable? The Oxford neuroethics gold standard framework. Front. Psychiatry 2017, 8, 44. [Google Scholar] [CrossRef] [PubMed]
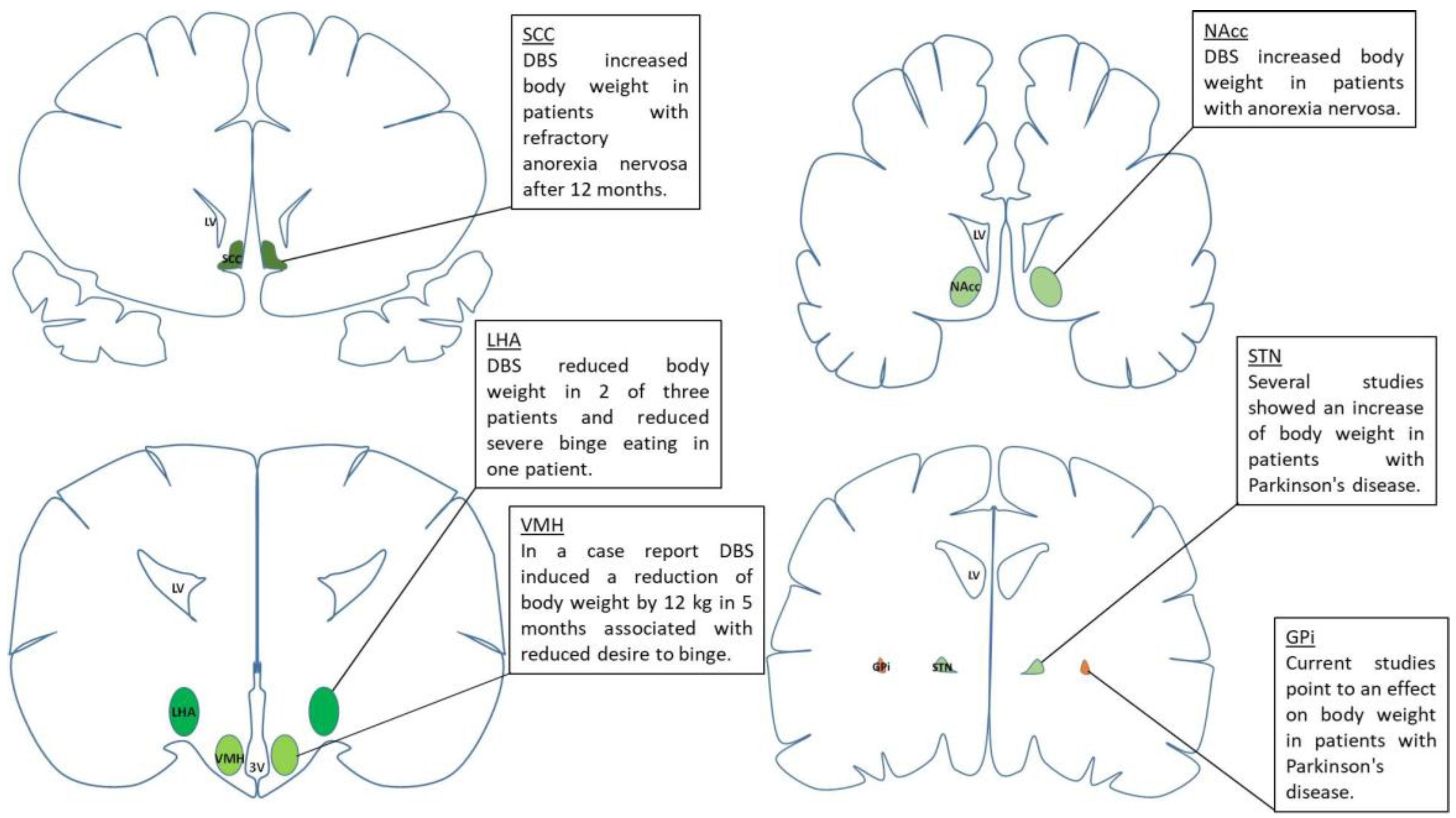

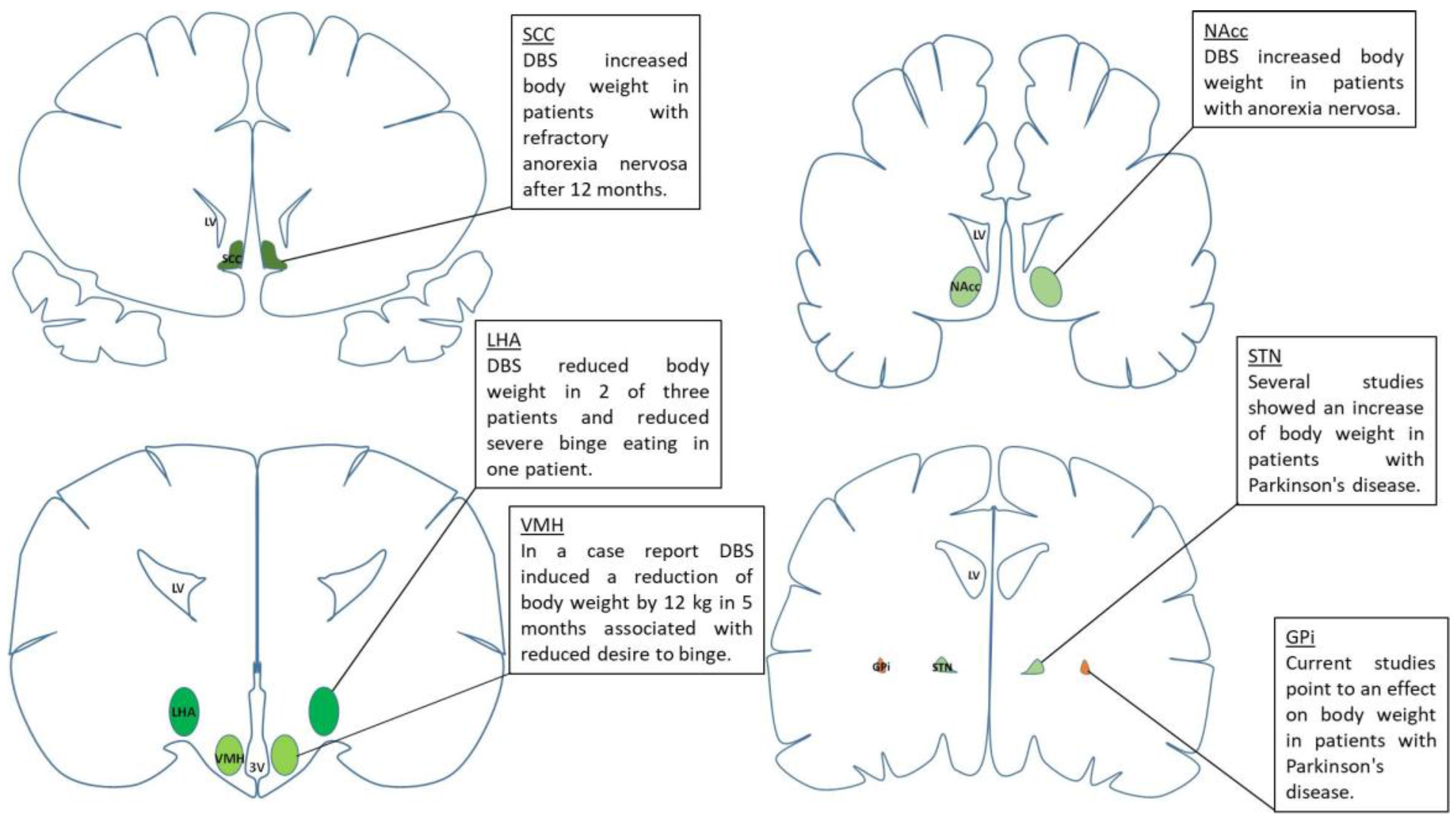{kind=link}
| Disease | Brain area | Mode | Species | Effect | Reference |
|---|---|---|---|---|---|
| Obesity | Nucleus accumbens shell | Unilateral, 160 Hz, 150 µA | Diet-induced obese mice | ↓ food intake; ↓ body weight | [50] |
| Nucleus accumbens shell | Unilateral, 130 Hz, 500 µA | Diet-induced obese rats | ↓ food intake; ↓ body weight | [35] | |
| Nucleus accumbens | Bilateral, 185 Hz, 3.5 V | Obese patient | ↓ body weight | [60] | |
| Nucleus accumbens | Bilateral, 130 Hz, 5 mA | Obese patient | ↓ body weight | [61] | |
| Lateral hypothalamus | Bilateral, 180-200 Hz, 2 V | Diet-induced obese rats | - food intake; ↓ body weight | [53] | |
| Lateral hypothalamus | Bilateral, 130 Hz, 300 µA | Obese Zucker rats | ↓ food intake; ↓ body weight | [37] | |
| Lateral hypothalamus | Bilateral, 185 Hz, 1–7 V | Obese patients | ↓ body weight | [31] | |
| Ventromedial hypothalamus | Unilateral, 150/500 Hz, 10 µA | Sprague-Dawley rats | ↓ body weight | [38] | |
| Ventromedial hypothalamus | Unilateral, 100 Hz | Dogs | ↓ food intake | [55] | |
| Ventromedial hypothalamus | Bilateral, 50 Hz, 0.5–1.5 mA | Göttingen minipigs | ↓ body weight | [39] | |
| Ventromedial hypothalamus | Unilateral, 80 Hz, 2V | Macaca fascicularis monkey | ↓ meal size; body weight; ↓ body fat | [54] | |
| Ventromedial hypothalamus | Bilateral, 50 Hz | Obese patient | - food intake; ↓ body weight | [32] | |
| Central nucleus of amygdala | Bilateral, 20/130 Hz, 125 µA | Sprague-Dawley rats | ↓ palatable food (sucrose) | [56] | |
| Rostromedial tegmental nucleus | Bilateral, 10 Hz, 20 µA | Sprague-Dawley rats | ↓ food intake | [46] | |
| Binge-eating disorder | Nucleus accumbens shell | Unilateral, 160 Hz, 150 µA | Mouse model of binge eating | ↓ binge eating | [50] |
| Nucleus accumbens core | Bilateral, 150 Hz, 150 µA | Rat model of binge eating | ↓ binge eating | [52] | |
| Lateral hypothalamus | Bilateral, 185 Hz, 1–7 V | Obese patients | ↓ severe binge eating | [31] | |
| Ventromedial hypothalamus | Bilateral, 50 Hz | Obese patient | ↓ tendency to binge | [32] | |
| Anorexia nervosa | Subcallosal cingulate | Bilateral, 130 Hz, 5–7 V | Anorexic patients | ↑ body weight | [33,41] |
| Subgenual cingulate cortex | Bilateral, 130 Hz, 5 mA | Anorexic patient | ↑ body weight | [62] | |
| Subgenual cingulate cortex | Bilateral, 120 Hz, 5 V | Underweight patient | ↑ body weight | [63] | |
| Nucleus accumbens | Bilateral, 180 Hz, 6 V | Anorexic patients | ↑ body weight | [64] | |
| Nucleus accumbens | Bilateral, 100 Hz, 6–8 V | Anorexic patients | ↑ body weight | [65] | |
| Bed nucleus of stria terminalis | Bilateral, 130 Hz, 4.3 V | Anorexic patient | - body weight, ↓ anxiety of food and eating | [66] | |
| Nucleus accumbens shell | Bilateral, 130 Hz, 100 µA | Sprague-Dawley rats | ↑ food intake (first hour) | [36] | |
| Nucleus accumbens shell | Bilateral, 130 Hz, 100 µA | Sprague-Dawley rats | - food intake; ↑body weight | [34] | |
| Internal globus pallidus | Bilateral, 130 Hz, 2.6 V | Patients with PD | ↑ body weight | [43] | |
| Ventromedial hypothalamus | Bilateral, 185 Hz, 2.5 -3.5 V | Vervet monkeys | ↑ food intake | [40] | |
| Subthalamic nucleus | Bilateral, 240 Hz, 2.4 mA | Patients with PD | ↑ high caloric and tasty food, ↑ body weight | [44] | |
| Subthalamic nucleus | Bilateral, 185 Hz, 2.9 V | Patients with PD | ↑ body weight, ↑ appetite | [67] | |
| Subthalamic nucleus | Not mentioned | Patients with PD | ↑ body weight | [57,58,59] |
© 2018 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Prinz, P.; Stengel, A. Deep Brain Stimulation—Possible Treatment Strategy for Pathologically Altered Body Weight? Brain Sci. 2018, 8, 19. https://doi.org/10.3390/brainsci8010019
Prinz P, Stengel A. Deep Brain Stimulation—Possible Treatment Strategy for Pathologically Altered Body Weight? Brain Sciences. 2018; 8(1):19. https://doi.org/10.3390/brainsci8010019
Chicago/Turabian StylePrinz, Philip, and Andreas Stengel. 2018. "Deep Brain Stimulation—Possible Treatment Strategy for Pathologically Altered Body Weight?" Brain Sciences 8, no. 1: 19. https://doi.org/10.3390/brainsci8010019
APA StylePrinz, P., & Stengel, A. (2018). Deep Brain Stimulation—Possible Treatment Strategy for Pathologically Altered Body Weight? Brain Sciences, 8(1), 19. https://doi.org/10.3390/brainsci8010019