
Contributions of Letter-Speech Sound Learning and Visual Print Tuning to Reading Improvement: Evidence from Brain Potential and Dyslexia Training Studies

Abstract
:1. Introduction
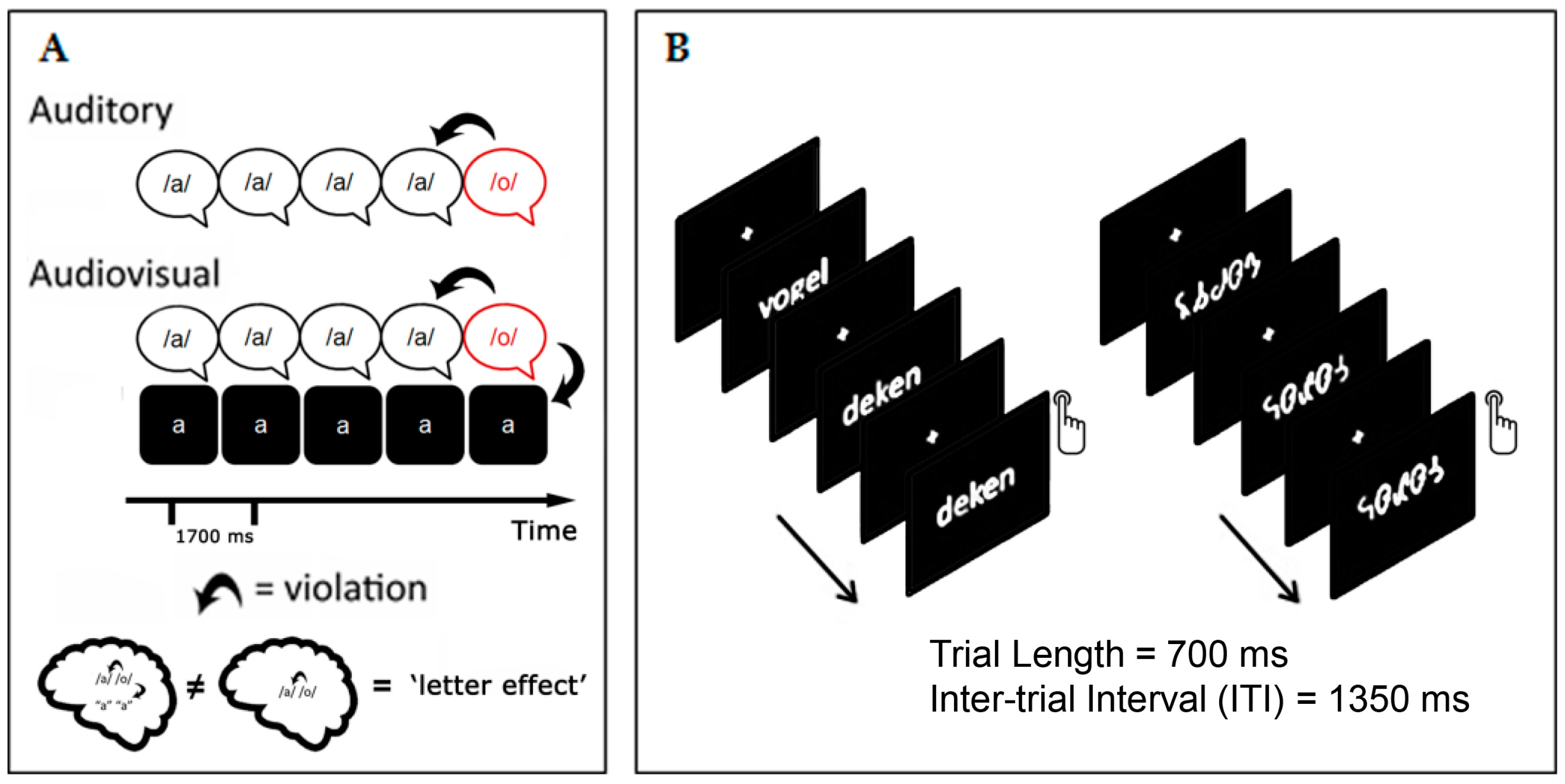
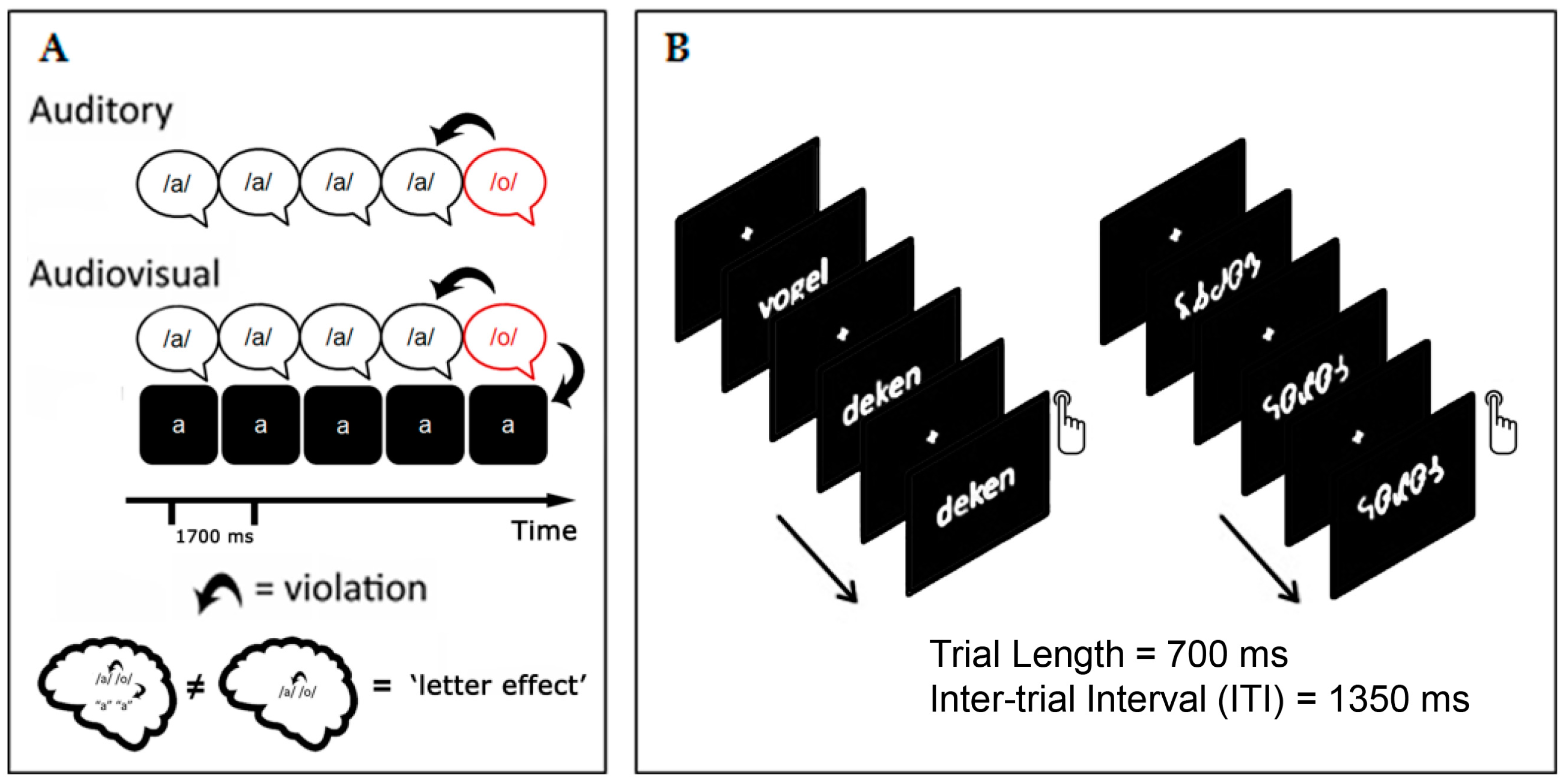
2. Electrophysiological Markers for Letter-Speech Sound Integration and Visual Specialization in Dyslexic Readers
2.1. Crossmodal MMN and Visual N170 Related to Reading Fluency in Dyslexic Children
2.2. Crossmodal MMN and Visual N170 as Predictors of Treatment Response
3. Current Results: Additional Analysis on MMN and N170
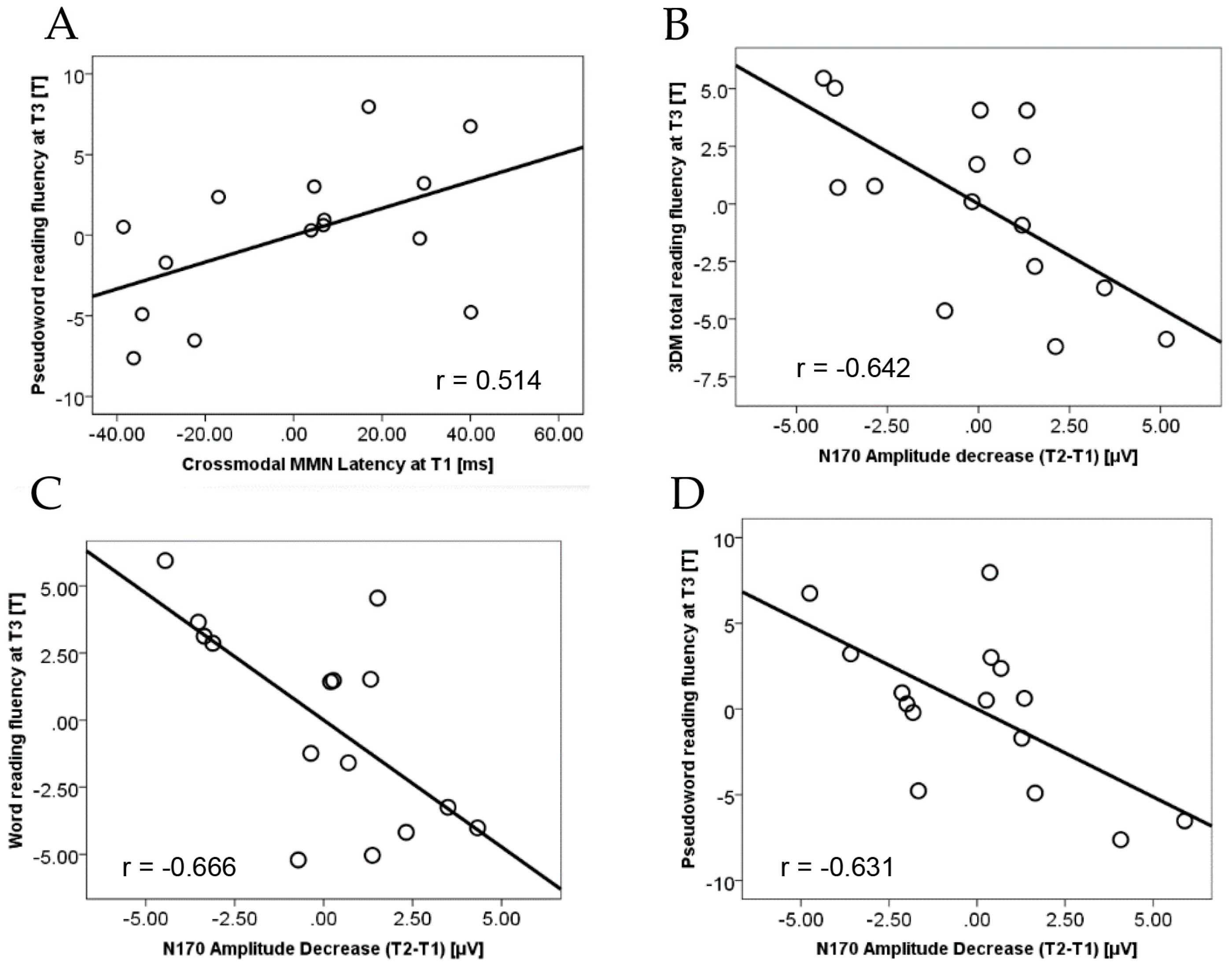
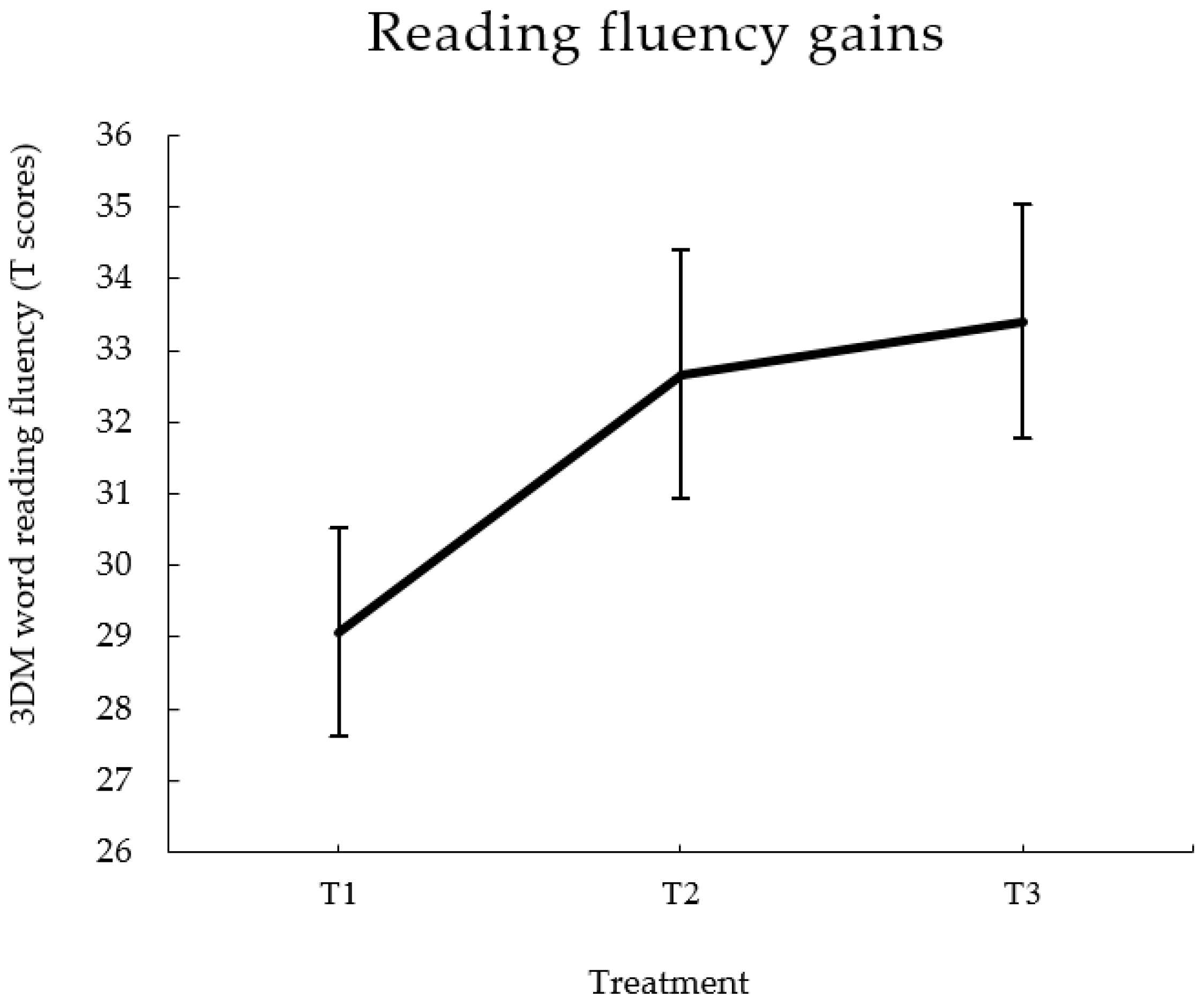
3.1. Prediction of Reading Gains after Extended Practice
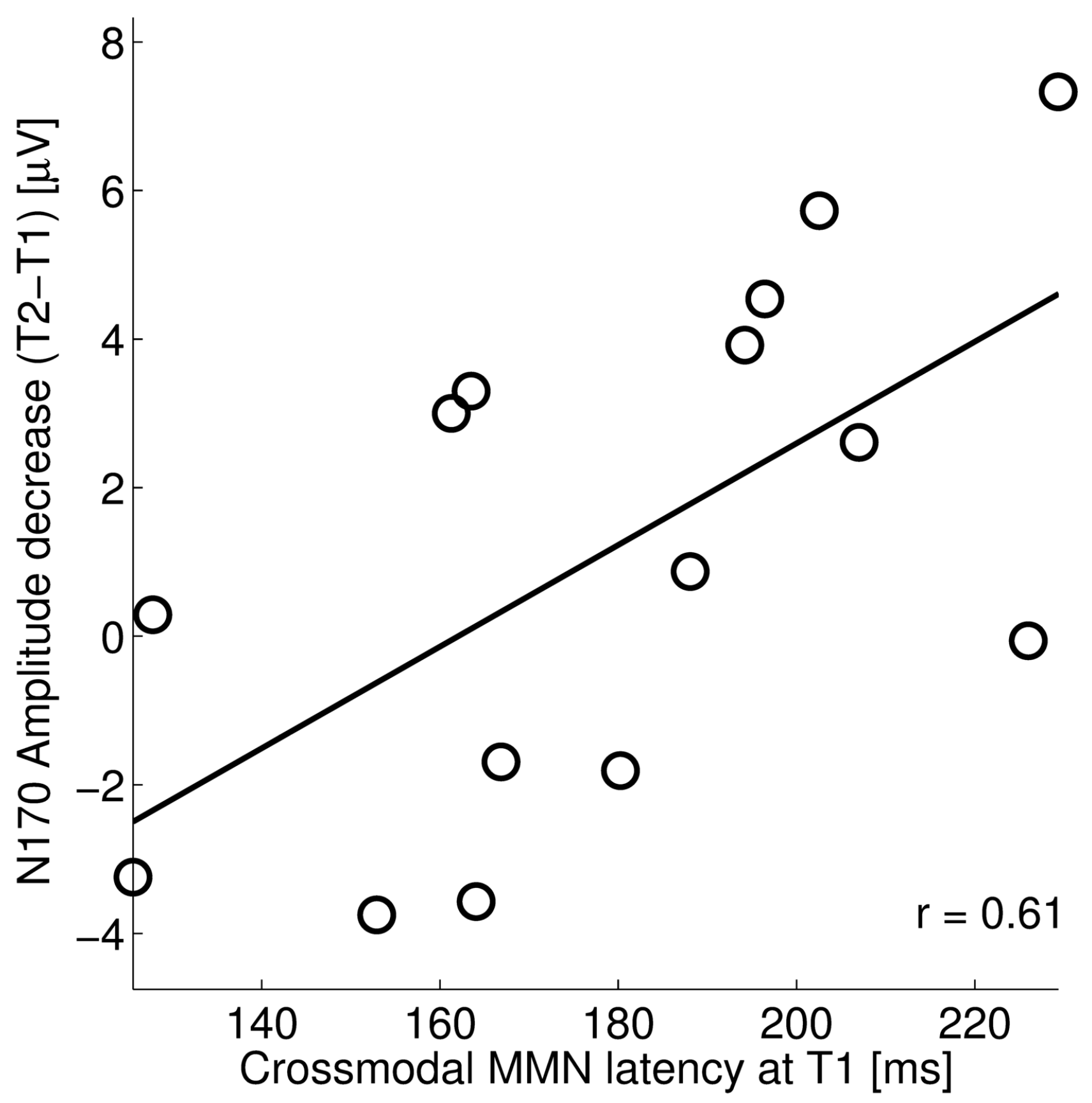
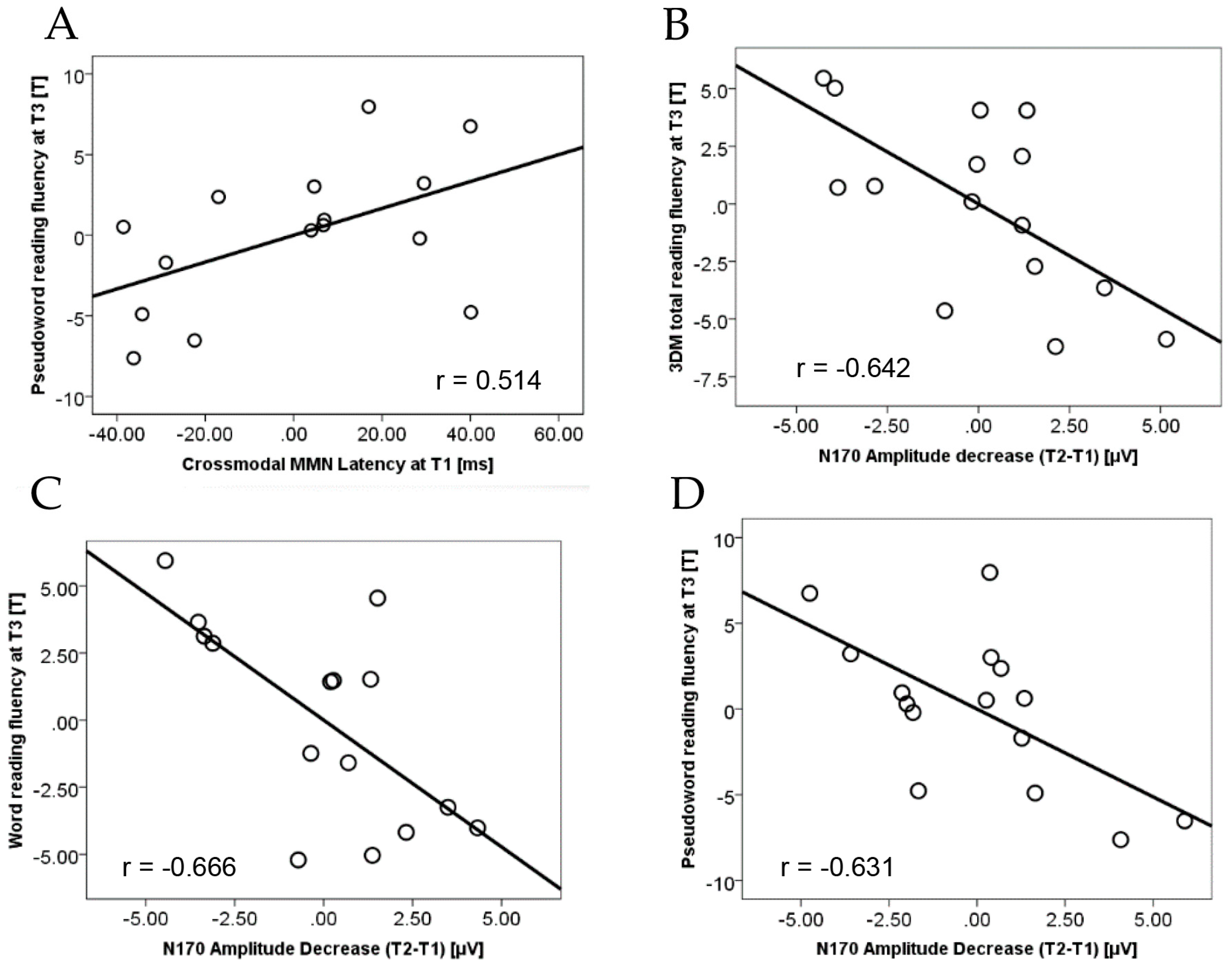
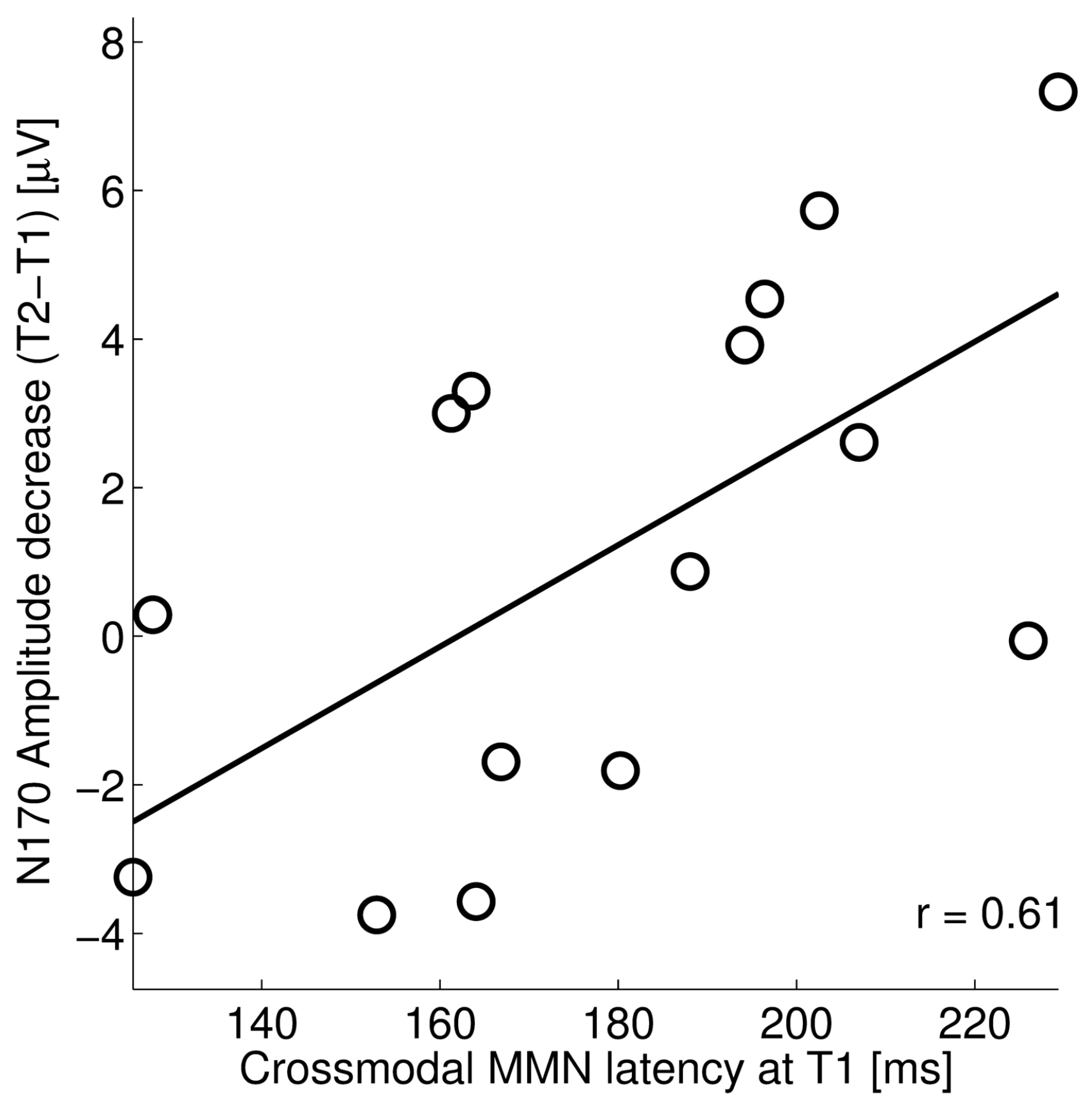
3.2. Relation between Crossmodal MMN and Visual N170
3.3. Summary of Results
4. Enhancing Letter-Speech Sound Binding by Associative, Implicit Learning
5. Associative, Implicit Learning and Other Cognitive Processes
5.1. Implicit Learning
5.2. Performance Monitoring
5.2.1. Error Processing
5.2.2. Feedback Learning
5.3. Conclusions from Associative Learning Studies
6. General Discussion and Future Research
Acknowledgments
Author Contributions
Conflicts of Interest
Appendix A

{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Dyslexics (N = 15) | Training Effects | |||||||||
|---|---|---|---|---|---|---|---|---|---|---|
| T1 | T2 | T3 | T2-T1 | T3-T2 | ||||||
| M | SD | M | SD | M | SD | t (14) | p-Value (One-Tailed) | t (14) | p-Value (One-Tailed) | |
| 3DM Word reading—accuracy (%) a | ||||||||||
| High Frequency | 91.98 | 7.48 | 96.25 | 7.86 | 97.43 | 1.74 | 2.17 | 0.024 | 0.60 | 0.280 |
| Low Frequency | 81.09 | 17.37 | 89.91 | 13.49 | 96.09 | 5.32 | 2.90 | 0.006 | 2.40 | 0.015 |
| Pseudowords | 69.00 | 17.09 | 76.44 | 20.21 | 80.99 | 11.16 | 1.34 | 0.100 | 0.96 | 0.176 |
| Total [T] b | 31.67 | 12.75 | 39.47 | 14.85 | 38.33 | 9.68 | 2.41 | 0.015 | −0.30 | 0.385 |
| 3DM Word reading—fluency [T] | ||||||||||
| High Frequency | 29.93 | 5.73 | 34.27 | 6.94 | 38.00 | 6.60 | 4.80 | 0.000 | 3.87 | 0.001 |
| Low Frequency | 29.93 | 6.39 | 33.93 | 6.72 | 34.07 | 6.83 | 4.37 | 0.000 | 0.12 | 0.451 |
| Pseudowords | 29.73 | 5.51 | 31.67 | 6.95 | 32.47 | 5.68 | 1.78 | 0.049 | 0.66 | 0.260 |
| Total | 29.07 | 5.62 | 32.67 | 6.72 | 33.40 | 6.32 | 4.41 | 0.000 | 0.94 | 0.182 |
References
- Zoccolotti, P.; De Luca, M.; Di Pace, E.; Gasperini, F.; Judica, A.; Spinelli, D. Word length effect in early reading and in developmental dyslexia. Brain Lang. 2005, 93, 369–373. [Google Scholar] [CrossRef] [PubMed]
- Seymour, P.H.K.; Aro, M.; Erskine, J.M. Foundation literacy acquisition in European orthographies. Br. J. Psychol. 2003, 94, 143–174. [Google Scholar] [CrossRef] [PubMed]
- Schmalz, X.; Marinus, E.; Coltheart, M.; Castles, A. Getting to the bottom of orthographic depth. Psychon. Bull. Rev. 2015, 22, 1614–1629. [Google Scholar] [CrossRef] [PubMed]
- Ehri, L.C. Learning to read words: Theory, findings, and issues. Sci. Stud. Read. 2005, 9, 167–188. [Google Scholar] [CrossRef]
- Snowling, M.J. Early identification and interventions for dyslexia: A contemporary view. J. Res. Spec. Educ. Needs 2013, 13, 7–14. [Google Scholar] [CrossRef] [PubMed]
- Blomert, L. Dyslexie in Nederland; Uitgeverij Nieuwezijds: Amsterdam, The Netherlands, 2005. [Google Scholar]
- Lyon, G.R.; Shaywitz, S.E.; Shaywitz, B.A. A definition of dyslexia. Ann. Dyslexia 2003, 53, 1–14. [Google Scholar] [CrossRef]
- Benjamin, C.F.A.; Gaab, N. What’s the story? The tale of reading fluency told at speed. Hum. Brain Mapp. 2012, 33, 2572–2585. [Google Scholar] [CrossRef] [PubMed]
- Shaywitz, S.E.S.; Shaywitz, B.B.A. Paying attention to reading: The neurobiology of reading and dyslexia. Dev. Psychopathol. 2008, 20, 1329–1349. [Google Scholar] [CrossRef] [PubMed]
- Tijms, J.; Hoeks, J.J.W.M.; Paulussen-Hoogeboom, M.C.; Smolenaars, A.J. Long-term effects of a psycholinguistic treatment for dyslexia. J. Res. Read. 2003, 26, 121–140. [Google Scholar] [CrossRef]
- Katzir, T.; Kim, Y.-S.; Wolf, M.; Morris, R.; Lovett, M.W. The Varieties of Pathways to Dysfluent Reading: Comparing Subtypes of Children With Dyslexia at Letter, Word, and Connected Text Levels of Reading. J. Learn. Disabil. 2008, 41, 47–66. [Google Scholar] [CrossRef] [PubMed]
- Leinonen, S.; Müller, K.; Leppänen, P.H.T.; Aro, M.; Ahonen, T.; Lyytinen, H. Heterogeneity in adult dyslexic readers: Relating processing skills to the speed and accuracy of oral text reading. Read. Writ. 2001, 14, 265–296. [Google Scholar] [CrossRef]
- Blomert, L.; Froyen, D. Multi-sensory learning and learning to read. Int. J. Psychophysiol. 2010, 77, 195–204. [Google Scholar] [CrossRef] [PubMed]
- Morais, J.; Alegria, J.; Content, A. The relationships between segmental analysis and alphabetic literacy: An interactive view. Cah. Psychol. Cogn. 1987, 7, 415–438. [Google Scholar]
- Wimmer, H.; Landerl, K.; Linortner, R.; Hummer, P. The relationship of phonemic awareness to reading acquisition: More consequence than precondition but still important. Cognition 1991, 40, 219–249. [Google Scholar] [CrossRef]
- Whitney, C.; Cornelissen, P. Letter-position encoding and dyslexia. J. Res. Read. 2005, 28, 274–301. [Google Scholar] [CrossRef]
- Nation, K. Form-meaning links in the development of visual word recognition. Philos. Trans. R. Soc. B Biol. Sci. 2009, 364, 3665–3674. [Google Scholar] [CrossRef] [PubMed]
- Snowling, M.J. The development of grapheme-phoneme correspondence in normal and dyslexic readers. J. Exp. Child Psychol. 1980, 29, 294–305. [Google Scholar] [CrossRef]
- Nation, K.; Cocksey, J. The relationship between knowing a word and reading it aloud in children’s word reading development. J. Exp. Child Psychol. 2009, 103, 296–308. [Google Scholar] [CrossRef] [PubMed]
- Nazir, T.A.; Jacobs, A.M.; O’Regan, J.K. Letter legibility and visual word recognition. Mem. Cognit. 1998, 26, 810–821. [Google Scholar] [CrossRef] [PubMed]
- New, B.; Ferrand, L.; Pallier, C.; Brysbaert, M. Reexamining the word length effect in visual word recognition: New evidence from the English Lexicon Project. Psychon. Bull. Rev. 2006, 13, 45–52. [Google Scholar] [CrossRef] [PubMed]
- Rayner, K.; Pollatsek, A.; Schotter, E.R. Reading: Word Identification and Eye Movements. In Handbook of Psychology, Volume 4: Experimental Psychology; Healy, A., Ed.; Wiley: Hoboken, NJ, USA, 2012; pp. 548–577. [Google Scholar]
- Reicher, G.M. Perceptual recognition as a function of meaninfulness of stimulus material. J. Exp. Psychol. 1969, 81, 275–280. [Google Scholar] [CrossRef] [PubMed]
- McCandliss, B.D.; Cohen, L.; Dehaene, S. The visual word form area: Expertise for reading in the fusiform gyrus. Trends Cogn. Sci. 2003, 7, 293–299. [Google Scholar] [CrossRef]
- Vaessen, A.; Blomert, L. Long-term cognitive dynamics of fluent reading development. J. Exp. Child Psychol. 2010, 105, 213–231. [Google Scholar] [CrossRef] [PubMed]
- Share, D.L. Phonological recoding and orthographic learning: A direct test of the self-teaching hypothesis. J. Exp. Child Psychol. 1999, 72, 95–129. [Google Scholar] [CrossRef] [PubMed]
- Share, D.L. Phonological recoding and self-teaching: Sine qua non of reading acquisition. Cognition 1995, 55, 151–218. [Google Scholar] [CrossRef]
- Ischebeck, A.; Indefrey, P.; Usui, N.; Nose, I.; Hellwig, F.; Taira, M. Reading in a regular orthography: An FMRI study investigating the role of visual familiarity. J. Cogn. Neurosci. 2004, 16, 727–741. [Google Scholar] [CrossRef] [PubMed]
- Carlisle, J.F.; Katz, L.A. Effects of word and morpheme familiarity on reading of derived words. Read. Writ. 2006, 19, 669–693. [Google Scholar] [CrossRef]
- Wimmer, H.; Goswami, U. The influence of orthographic consistency on reading development: Word recognition in English and German children. Cognition 1994, 51, 91–103. [Google Scholar] [CrossRef]
- Snowling, M.J. Dyslexia as a Phonological Deficit: Evidence and Implications. Child Psychol. Psychiatry Rev. 1998, 3, 4–11. [Google Scholar] [CrossRef]
- Ramus, F.; Szenkovits, G. What phonological deficit? Q. J. Exp. Psychol. 2008, 61, 129–141. [Google Scholar] [CrossRef] [PubMed]
- Blomert, L.; Willems, G. Is there a causal link from a phonological awareness deficit to reading failure in children at familial risk for dyslexia? Dyslexia 2010, 16, 300–317. [Google Scholar] [CrossRef] [PubMed]
- Aravena, S.; Snellings, P.; Tijms, J.; van der Molen, M.W. A lab-controlled simulation of a letter–speech sound binding deficit in dyslexia. J. Exp. Child Psychol. 2013, 115, 691–707. [Google Scholar] [CrossRef] [PubMed]
- Froyen, D.; Willems, G.; Blomert, L. Evidence for a specific cross-modal association deficit in dyslexia: An electrophysiological study of letter-speech sound processing. Dev. Sci. 2011, 14, 635–648. [Google Scholar] [CrossRef] [PubMed]
- Blau, V.; van Atteveldt, N.; Ekkebus, M.; Goebel, R.; Blomert, L. Reduced neural integration of letters and speech sounds links phonological and reading deficits in adult dyslexia. Curr. Biol. 2009, 19, 503–508. [Google Scholar] [CrossRef] [PubMed]
- Blau, V.; Reithler, J.; van Atteveldt, N.; Seitz, J.; Gerretsen, P.; Goebel, R.; Blomert, L. Deviant processing of letters and speech sounds as proximate cause of reading failure: A functional magnetic resonance imaging study of dyslexic children. Brain 2010, 133, 868–879. [Google Scholar] [CrossRef] [PubMed]
- Žarić, G.; Fraga González, G.; Tijms, J.; van der Molen, M.W.; Blomert, L.; Bonte, M. Reduced neural integration of letters and speech sounds in dyslexic children scales with individual differences in reading fluency. PLoS ONE 2014, 9, e110337. [Google Scholar] [CrossRef] [PubMed]
- Moll, K.; Hasko, S.; Groth, K.; Bartling, J.; Schulte-Körne, G. Letter-sound processing deficits in children with developmental dyslexia: An ERP study. Clin. Neurophysiol. 2016, 127, 1989–2000. [Google Scholar] [CrossRef] [PubMed]
- Žarić, G.; Fraga González, G.; Tijms, J.; van der Molen, M.W.; Blomert, L.; Bonte, M. Crossmodal deficit in dyslexic children: Practice affects the neural timing of letter-speech sound integration. Front. Hum. Neurosci. 2015, 9, 1–14. [Google Scholar] [CrossRef] [PubMed]
- Jones, M.W.; Kuipers, J.-R.; Thierry, G. ERPs Reveal the Time-Course of Aberrant Visual-Phonological Binding in Developmental Dyslexia. Front. Hum. Neurosci. 2016, 10, 1–13. [Google Scholar] [CrossRef] [PubMed]
- Karipidis, I.; Pleisch, G.; Röthlisberger, M.; Hofstetter, C.; Dornbierer, D.; Stämpfli, P.; Brem, S. Neural initialization of audiovisual integration in prereaders at varying risk for developmental dyslexia. Hum. Brain Mapp. 2016. [Google Scholar] [CrossRef] [PubMed]
- Seidenberg, M.S.; Tanenhaus, M.K. Orthographic effects on rhyme monitoring. J. Exp. Psychol. Hum. Learn. Mem. 1979, 5, 546–554. [Google Scholar] [CrossRef]
- Dijkstra, T.; Roelofs, A.; Fieuws, S. Orthographic effects on phoneme monitoring. Can. J. Exp. Psychol. 1995, 49, 264–271. [Google Scholar] [CrossRef] [PubMed]
- Ziegler, J.C.; Ferrand, L. Orthography shapes the perception of speech: The consistency effect in auditory word recognition. Psychon. Bull. Rev. 1998, 5, 683–689. [Google Scholar] [CrossRef]
- Cone, N.E.; Burman, D.D.; Bitan, T.; Bolger, D.J.; Booth, J.R. Developmental changes in brain regions involved in phonological and orthographic processing during spoken language processing. Neuroimage 2008, 41, 623–635. [Google Scholar] [CrossRef] [PubMed]
- Bishop, D.V.M. Dyslexia: What’s the problem? Dev. Sci. 2006, 9, 256–257. [Google Scholar] [CrossRef] [PubMed]
- Wimmer, H.; Schurz, M. Dyslexia in regular orthographies: Manifestation and causation. Dyslexia 2010, 16, 283–299. [Google Scholar] [CrossRef] [PubMed]
- Blomert, L. The neural signature of orthographic-phonological binding in successful and failing reading development. Neuroimage 2011, 57, 695–703. [Google Scholar] [CrossRef] [PubMed]
- Hahn, N.; Foxe, J.J.; Molholm, S. Impairments of multisensory integration and cross-sensory learning as pathways to dyslexia. Neurosci. Biobehav. Rev. 2014, 47, 384–392. [Google Scholar] [CrossRef] [PubMed]
- Mann, V.; Wimmer, H. Phoneme awareness and pathways into literacy: A comparison of German and American children. Read. Writ. 2002, 15, 653–682. [Google Scholar] [CrossRef]
- Schlaggar, B.L.; McCandliss, B.D. Development of neural systems for reading. Annu. Rev. Neurosci. 2007, 30, 475–503. [Google Scholar] [CrossRef] [PubMed]
- Liberman, I.Y.; Shankweiler, D.; Fischer, F.W.; Carter, B. Explicit syllable and phoneme segmentation in the young child. J. Exp. Child Psychol. 1974, 18, 201–212. [Google Scholar] [CrossRef]
- Pugh, K.R.; Mencl, W.E.; Jenner, A.R.; Lee, J.R.; Katz, L.; Frost, S.J.; Shaywitz, S.E.; Shaywitz, B.A. Neuroimaging studies of reading development and reading disability. Learn. Disabil. Res. Pract. 2001, 16, 240–249. [Google Scholar] [CrossRef]
- Dehaene, S.; Cohen, L.; Sigman, M.; Vinckier, F. The neural code for written words: A proposal. Trends Cogn. Sci. 2005, 9, 335–341. [Google Scholar] [CrossRef] [PubMed]
- Castro-Caldas, A.; Petersson, K.M.; Reis, A.; Stone-Elander, S.; Ingvar, M. The illiterate brain. Learning to read and write during childhood influences the functional organization of the adult brain. Brain 1998, 121, 1053–1063. [Google Scholar] [CrossRef] [PubMed]
- Brem, S.; Bach, S.; Kucian, K.; Guttorm, T.K.; Martin, E.; Lyytinen, H.; Brandeis, D.; Richardson, U. Brain sensitivity to print emerges when children learn letter-speech sound correspondences. Proc. Natl. Acad. Sci. USA 2010, 107, 7939–7944. [Google Scholar] [CrossRef] [PubMed]
- Dehaene, S.; Pegado, F.; Braga, L.W.; Ventura, P.; Nunes Filho, G.; Jobert, A.; Kolinsky, R.; Morais, J.; Cohen, L. How learning to read changes the cortical networks for vision and language. Science 2010, 330, 1359–1364. [Google Scholar] [CrossRef] [PubMed]
- Van Atteveldt, N.; Ansari, D. How symbols transform brain function: A review in memory of Leo Blomert. Trends Neurosci. Educ. 2014, 3, 44–49. [Google Scholar] [CrossRef]
- Simos, P.G.; Breier, J.I.; Fletcher, J.M.; Foorman, B.R.; Castillo, E.M.; Papanicolaou, A.C. Brain mechanisms for reading words and pseudowords: An integrated approach. Cereb. Cortex 2002, 12, 297–305. [Google Scholar] [CrossRef] [PubMed]
- Jobard, G.; Crivello, F.; Tzourio-Mazoyer, N. Evaluation of the dual route theory of reading: A metanalysis of 35 neuroimaging studies. Neuroimage 2003, 20, 693–712. [Google Scholar] [CrossRef]
- Brennan, C.; Cao, F.; Pedroarena-Leal, N.; McNorgan, C.; Booth, J.R. Reading acquisition reorganizes the phonological awareness network only in alphabetic writing systems. Hum. Brain Mapp. 2013, 34, 3354–3368. [Google Scholar] [CrossRef] [PubMed]
- Monzalvo, K.; Dehaene-Lambertz, G. How reading acquisition changes children’s spoken language network. Brain Lang. 2013, 127, 356–365. [Google Scholar] [CrossRef] [PubMed]
- Sandak, R.; Mencl, W.; Frost, S.J.; Pugh, K.R. The neurobiological basis of skilled and impaired reading: Recent findings and new directions. Sci. Stud. Read. 2004, 8, 273–292. [Google Scholar] [CrossRef]
- Brunswick, N.; McCrory, E.; Price, C.J.; Frith, C.D.; Frith, U. Explicit and implicit processing of words and pseudowords by adult developmental dyslexics A search for Wernicke’s Wortschatz? Brain 1999, 122, 1901–1917. [Google Scholar] [CrossRef] [PubMed]
- Paulesu, E.; Démonet, J.F.; Fazio, F.; McCrory, E.; Chanoine, V.; Brunswick, N.; Cappa, S.F.; Cossu, G.; Habib, M.; Frith, C.D.; et al. Dyslexia: Cultural diversity and biological unity. Science 2001, 291, 2165–2167. [Google Scholar] [CrossRef] [PubMed]
- Simos, P.G.; Breier, J.I.; Fletcher, J.M.; Bergman, E.; Papanicolaou, A.C. Cerebral mechanisms involved in word reading in dyslexic children: A magnetic source imaging approach. Cereb. Cortex 2000, 10, 809–816. [Google Scholar] [CrossRef] [PubMed]
- Fraga González, G.; Žarić, G.; Tijms, J.; Bonte, M.; Blomert, L.; van der Molen, M.W. Brain-potential analysis of visual word recognition in dyslexics and typically reading children. Front. Hum. Neurosci. 2014, 8, 474. [Google Scholar] [CrossRef] [PubMed]
- Fraga González, G.; Žarić, G.; Tijms, J.; Bonte, M.L.M.; Blomert, L.; Leppänen, P.H.T.; van der Molen, M.W. Responsivity to dyslexia training indexed by the N170 amplitude of the brain potential elicited by word reading. Brain Cogn. 2016, 106, 42–54. [Google Scholar] [CrossRef] [PubMed]
- Fraga González, G.; Žarić, G.; Tijms, J.; Bonte, M.; Blomert, L.; van der Molen, M.W. A Randomized Controlled Trial on The Beneficial Effects of Training Letter-Speech Sound Integration on Reading Fluency in Children with Dyslexia. PLoS ONE 2015, 10, e0143914. [Google Scholar]
- Tijms, J. Effectiveness of computer-based treatment for dyslexia in a clinical care setting: Outcomes and moderators. Educ. Psychol. 2011, 31, 873–896. [Google Scholar] [CrossRef]
- Fraga González, G.; van der Molen, M.J.W.; Žarić, G.; Bonte, M.M.; Tijms, J.; Blomert, L.; Stam, C.J. Graph analysis of EEG resting state functional networks in dyslexic readers. Clin. Neurophysiol. 2016, 127, 3165–3175. [Google Scholar] [CrossRef] [PubMed]
- Žarić, G.; Correia, J.M.; Fraga González, G.; Tijms, J.; van der Molen, M.W.; Blomert, L.; Bonte, M. Altered patterns of directed connectivity within the reading network of dyslexic children and their relation to reading dysfluency. Dev. Cogn. Neurosci. 2017, 23, 1–13. [Google Scholar] [CrossRef] [PubMed]
- Näätänen, R. The perception of speech sounds by the human brain as reflected by the mismtch negativity (MMN) and its magnetic equivalent (MMNm). Psychophysiology 2001, 38, 1–21. [Google Scholar] [CrossRef] [PubMed]
- Näätänen, R.; Paavilainen, P.; Rinne, T.; Alho, K. The mismatch negativity (MMN) in basic research of central auditory processing: A review. Clin. Neurophysiol. 2007, 118, 2544–2590. [Google Scholar] [CrossRef] [PubMed]
- Froyen, D.; Bonte, M.; van Atteveldt, N.; Blomert, L. The long road to automation: Neurocognitive development of letter-speech sound processing. J. Cogn. Neurosci. 2009, 21, 567–580. [Google Scholar] [CrossRef] [PubMed]
- Froyen, D.; Van Atteveldt, N.; Bonte, M.; Blomert, L. Cross-modal enhancement of the MMN to speech-sounds indicates early and automatic integration of letters and speech-sounds. Neurosci. Lett. 2008, 430, 23–28. [Google Scholar] [CrossRef] [PubMed]
- Blomert, L.; Vaessen, A.A. 3DM Differential Diagnostics for Dyslexia: Cognitive Analysis of Reading and Spelling; Boom Test: Amsterdam, The Netherlands, 2009. [Google Scholar]
- Bentin, S.; Mouchetant-Rostaing, Y.; Giard, M.H.; Echallier, J.F.; Pernier, J. ERP manifestations of processing printed words at different psycholinguistic levels: Time course and scalp distribution. J. Cogn. Neurosci. 1999, 11, 235–260. [Google Scholar] [CrossRef] [PubMed]
- Maurer, U.; Brem, S.; Bucher, K.; Brandeis, D. Emerging neurophysiological specialization for letter strings. J. Cogn. Neurosci. 2005, 17, 1532–1552. [Google Scholar] [CrossRef] [PubMed]
- Maurer, U.; Mccandliss, B.D. The Development of the Visual Expertise for Words: The Contribution of Electrophysiology; Grigorenko, E.L., Naples, A.J., Eds.; Lawrence Erlbaum Associates: Mahwah, NJ, USA, 2007. [Google Scholar]
- Maurer, U.; Schulz, E.; Brem, S.; Van der Mark, S.; Bucher, K.; Martin, E.; Brandeis, D. The development of print tuning in children with dyslexia: Evidence from longitudinal ERP data supported by fMRI. Neuroimage 2011, 57, 714–722. [Google Scholar] [CrossRef] [PubMed]
- Araújo, S.; Bramão, I.; Faísca, L.; Petersson, K.M.; Reis, A. Electrophysiological correlates of impaired reading in dyslexic pre-adolescent children. Brain Cogn. 2012, 79, 79–88. [Google Scholar] [CrossRef] [PubMed]
- Helenius, P.; Tarkiainen, A.; Cornelissen, P.L.; Hansen, P.C.; Salmelin, R. Dissociation of normal feature analysis and deficient processing of letter-strings in dyslexic adults. Cereb. Cortex 1999, 9, 476–483. [Google Scholar] [CrossRef] [PubMed]
- Mahé, G.; Bonnefond, A.; Gavens, N.; Dufour, A.; Doignon-Camus, N. Impaired visual expertise for print in French adults with dyslexia as shown by N170 tuning. Neuropsychologia 2012, 50, 3200–3206. [Google Scholar] [CrossRef] [PubMed]
- Korinth, S.P.; Sommer, W.; Breznitz, Z. Does silent reading speed in normal adult readers depend on early visual processes? evidence from event-related brain potentials. Brain Lang. 2012, 120, 15–26. [Google Scholar] [CrossRef] [PubMed]
- Maurer, U.; Brem, S.; Kranz, F.; Bucher, K.; Benz, R.; Halder, P.; Steinhausen, H.-C.; Brandeis, D. Coarse neural tuning for print peaks when children learn to read. Neuroimage 2006, 33, 749–758. [Google Scholar] [CrossRef] [PubMed]
- Maurer, U.; Brem, S.; Bucher, K.; Kranz, F.; Benz, R.; Steinhausen, H.-C.; Brandeis, D. Impaired tuning of a fast occipito-temporal response for print in dyslexic children learning to read. Brain 2007, 130, 3200–3210. [Google Scholar] [CrossRef] [PubMed]
- Bonte, M.L.M.; Blomert, L. Developmental dyslexia: ERP correlates of anomalous phonological processing during spoken word recognition. Brain Res. Cogn. Brain Res. 2004, 21, 360–376. [Google Scholar] [CrossRef] [PubMed]
- Savill, N.J.; Thierry, G. Reading for sound with dyslexia: Evidence for early orthographic and late phonological integration deficits. Brain Res. 2011, 1385, 192–205. [Google Scholar] [CrossRef] [PubMed]
- Ramus, F.; Ahissar, M. Developmental dyslexia: The difficulties of interpreting poor performance, and the importance of normal performance. Cogn. Neuropsychol. 2012, 29, 104–122. [Google Scholar] [CrossRef] [PubMed]
- Neuhoff, N.; Bruder, J.; Bartling, J.; Warnke, A.; Remschmidt, H.; Müller-Myhsok, B.; Schulte-Körne, G. Evidence for the late MMN as a neurophysiological endophenotype for dyslexia. PLoS ONE 2012, 7, e34909. [Google Scholar] [CrossRef] [PubMed]
- Shestakova, A.; Huotilainen, M.; Čeponienė, R.; Cheour, M. Event-related potentials associated with second language learning in children. Clin. Neurophysiol. 2003, 114, 1507–1512. [Google Scholar] [CrossRef]
- Nash, H.M.; Gooch, D.; Hulme, C.; Mahajan, Y.; McArthur, G.; Steinmetzger, K.; Snowling, M.J. Are the literacy difficulties that characterize developmental dyslexia associated with a failure to integrate letters and speech sounds? Dev. Sci. 2016. [Google Scholar] [CrossRef] [PubMed]
- Dujardin, T.; Etienne, Y.; Contentin, C.; Bernard, C.; Largy, P.; Mellier, D.; Lalonde, R.; Rebaï, M. Behavioral performances in participants with phonological dyslexia and different patterns on the N170 component. Brain Cogn. 2011, 75, 91–100. [Google Scholar] [CrossRef] [PubMed]
- Alexander, A.W.; Slinger-Constant, A.-M. Current status of treatments for dyslexia: Critical review. J. Child Neurol. 2004, 19, 744–758. [Google Scholar] [CrossRef] [PubMed]
- Gabrieli, J.D.E. Dyslexia: A new synergy between education and cognitive neuroscience. Science 2009, 325, 280–283. [Google Scholar] [CrossRef] [PubMed]
- Wolff, U. Effects of a randomised reading intervention study: An application of structural equation modelling. Dyslexia 2011, 17, 295–311. [Google Scholar] [CrossRef] [PubMed]
- Ehri, L.C. Phases of acquisition in learning to read words and implications for teaching. In Word Recognition in Beginning Literacy; Stainthorp, R., Tomlinson, P., Eds.; Erlbaum: Mahwah, NJ, USA, 2002; pp. 7–28. [Google Scholar]
- Tijms, J. A Process-Oriented Evaluation of a Computerised Treatment for Dyslexia. Educ. Psychol. 2004, 24, 767–791. [Google Scholar] [CrossRef]
- McCandliss, B.D.; Noble, K.G. The development of reading impairment: A cognitive neuroscience model. Ment. Retard. Dev. Disabil. Res. Rev. 2003, 9, 196–204. [Google Scholar] [CrossRef] [PubMed]
- Baron, J.; Thurston, I. An analysis of the world-superiority effect. Cogn. Psychol. 1973, 4, 207–228. [Google Scholar] [CrossRef]
- Davydov, V.V. The state of research on learning activity. J. Russ. East Eur. Psychol. 1995, 33, 55–70. [Google Scholar] [CrossRef]
- Schneider, W. Controlled & automatic processing: Behavior, theory, and biological mechanisms. Cogn. Sci. 2003, 27, 525–559. [Google Scholar]
- Folia, V.; Uddén, J.; Forkstam, C.; Ingvar, M.; Hagoort, P.; Petersson, K.M. Implicit learning and dyslexia. Ann. N. Y. Acad. Sci. 2008, 1145, 132–150. [Google Scholar] [CrossRef] [PubMed]
- Gombert, J. Implicit and explicit learning to read: Implication as for subtypes of dyslexia. Curr. Psychol. Lett. 2003, 10, 1–7. [Google Scholar]
- Aravena, S.; Tijms, J. Reading Fluency and dyslexia: Innovative developments in the role of associative learning and repetitive exposure in skill acquisition. In Educational Psychology: Cognition and Learning, Individual Differences, and Motivation; Larson, J.E., Ed.; Nova Science: Hauppage, NY, USA, 2009; pp. 113–141. [Google Scholar]
- Tijms, J. The Development of Reading Accuracy and Reading Rate during Treatment of Dyslexia. Educ. Psychol. 2007, 27, 273–294. [Google Scholar] [CrossRef]
- Aravena, S.; Tijms, J.; Snellings, P.; van der Molen, M.W. Predicting responsiveness to intervention in dyslexia using dynamic assessment. Learn. Individ. Differ. 2016, 49, 209–215. [Google Scholar] [CrossRef]
- Aravena, S.; Tijms, J.; Snellings, P.; van der Molen, M.W. Predicting individual differences in reading and spelling skill with an artificial script-based letter-speech sound training. J. Learn. Disabil. 2017, in press. [Google Scholar]
- Hedenius, M.; Persson, J.; Alm, P.A.; Ullman, M.T.; Howard, J.H.; Howard, D.V.; Jennische, M. Impaired implicit sequence learning in children with developmental dyslexia. Res. Dev. Disabil. 2013, 34, 3924–3935. [Google Scholar] [CrossRef] [PubMed]
- Harm, M.W.; Seidenberg, M.S. Phonology, reading acquisition, and dyslexia: Insights from connectionist models. Psychol. Rev. 1999, 106, 491–528. [Google Scholar] [CrossRef] [PubMed]
- Frost, R.; Siegelman, N.; Narkiss, A.; Afek, L. What predicts successful literacy acquisition in a second language? Psychol. Sci. 2013, 24, 1243–1252. [Google Scholar] [CrossRef] [PubMed]
- Nissen, M.J.; Bullemer, P. Attentional requirements of learning: Evidence from performance measures. Cogn. Psychol. 1987, 19, 1–32. [Google Scholar] [CrossRef]
- Howard, J.H.; Howard, D.V.; Japikse, K.C.; Eden, G.F. Dyslexics are impaired on implicit higher-order sequence learning, but not on implicit spatial context learning. Neuropsychologia 2006, 44, 1131–1144. [Google Scholar] [CrossRef] [PubMed]
- Gabay, Y.; Thiessen, E.D.; Holt, L.L. Impaired Statistical Learning in Developmental Dyslexia. J. Speech Lang. Hear. Res. 2015, 58, 934–945. [Google Scholar] [CrossRef] [PubMed]
- Gabay, Y.; Schiff, R.; Vakil, E. Dissociation between online and offline learning in developmental dyslexia. J. Clin. Exp. Neuropsychol. 2012, 34, 279–288. [Google Scholar] [CrossRef] [PubMed]
- Hedenius, M.; Persson, J.; Tremblay, A.; Adi-Japha, E.; Veríssimo, J.; Dye, C.D.; Alm, P.; Jennische, M.; Bruce Tomblin, J.; Ullman, M.T. Grammar predicts procedural learning and consolidation deficits in children with Specific Language Impairment. Res. Dev. Disabil. 2011, 32, 2362–2375. [Google Scholar] [CrossRef] [PubMed]
- Vakil, E.; Lowe, M.; Goldfus, C. Performance of Children With Developmental Dyslexia on Two Skill Learning Tasks—Serial Reaction Time and Tower of Hanoi Puzzle A Test of the Specific Procedural Learning Difficulties Theory. J. Learn. Disabil. 2015, 48, 471–481. [Google Scholar] [CrossRef] [PubMed]
- Kelly, S.W.; Griffiths, S.; Frith, U. Evidence for implicit sequence learning in dyslexia. Dyslexia 2002, 8, 43–52. [Google Scholar] [CrossRef] [PubMed]
- Rüsseler, J.; Gerth, I.; Münte, T.F. Implicit learning is intact in adult developmental dyslexic readers: Evidence from the serial reaction time task and artificial grammar learning. J. Clin. Exp. Neuropsychol. 2006, 28, 808–827. [Google Scholar] [CrossRef] [PubMed]
- Lum, J.A.G.; Ullman, M.T.; Conti-Ramsden, G. Procedural learning is impaired in dyslexia: Evidence from a meta-analysis of serial reaction time studies. Res. Dev. Disabil. 2013, 34, 3460–3476. [Google Scholar] [CrossRef] [PubMed]
- Frost, R.; Armstrong, B.C.; Siegelman, N.; Christiansen, M.H. Domain generality versus modality specificity: The paradox of statistical learning. Trends Cogn. Sci. 2015, 19, 117–125. [Google Scholar] [CrossRef] [PubMed]
- Pothos, E.M.; Kirk, J. Investigating learning deficits associated with dyslexia. Dyslexia 2004, 10, 61–76. [Google Scholar] [CrossRef] [PubMed]
- Pavlidou, E.V.; Williams, J.M. Implicit learning and reading: Insights from typical children and children with developmental dyslexia using the artificial grammar learning (AGL) paradigm. Res. Dev. Disabil. 2014, 35, 1457–1472. [Google Scholar] [CrossRef] [PubMed]
- Pavlidou, E.V.; Kelly, M.L.; Williams, J.M. Do children with developmental dyslexia have impairments in implicit learning? Dyslexia 2010, 16, 143–161. [Google Scholar] [CrossRef] [PubMed]
- Pavlidou, E.V.; Williams, J.M.; Kelly, L.M. Artificial grammar learning in primary school children with and without developmental dyslexia. Ann. Dyslexia 2009, 59, 55–77. [Google Scholar] [CrossRef] [PubMed]
- Kahta, S.; Schiff, R. Implicit learning deficits among adults with developmental dyslexia. Ann. Dyslexia 2016, 66, 235–250. [Google Scholar] [CrossRef] [PubMed]
- Katan, P.; Kahta, S.; Sasson, A.; Schiff, R. Performance of children with developmental dyslexia on high and low topological entropy artificial grammar learning task. Ann. Dyslexia 2016. [Google Scholar] [CrossRef] [PubMed]
- Bonte, M.L.M.; Poelmans, H.; Blomert, L. Deviant neurophysiological responses to phonological regularities in speech in dyslexic children. Neuropsychologia 2007, 45, 1427–1437. [Google Scholar] [CrossRef] [PubMed]
- Noordenbos, M.W.; Segers, E.; Mitterer, H.; Serniclaes, W.; Verhoeven, L. Deviant neural processing of phonotactic probabilities in adults with dyslexia. Neuroreport 2013, 24, 746–750. [Google Scholar] [CrossRef] [PubMed]
- Bonte, M.L.; Mitterer, H.; Zellagui, N.; Poelmans, H.; Blomert, L. Auditory cortical tuning to statistical regularities in phonology. Clin. Neurophysiol. 2005, 116, 2765–2774. [Google Scholar] [CrossRef] [PubMed]
- Norris, D.; McQueen, J.M.; Cutler, A. Perceptual learning in speech. Cogn. Psychol. 2003, 47, 204–238. [Google Scholar] [CrossRef]
- Keetels, M.; Schakel, L.; Bonte, M.; Vroomen, J. Phonetic recalibration of speech by text. Attent. Percept. Psychophys. 2016, 78, 938–945. [Google Scholar] [CrossRef] [PubMed]
- Keetels, M.; Tilburg University, Tilburg, The Netherlands. Unpublished work. 2017.
- Ridderinkhof, K.R.; van den Wildenberg, W.P.M.; Segalowitz, S.J.; Carter, C.S. Neurocognitive mechanisms of cognitive control: The role of prefrontal cortex in action selection, response inhibition, performance monitoring, and reward-based learning. Brain Cogn. 2004, 56, 129–140. [Google Scholar] [CrossRef] [PubMed]
- Yeung, N.; Botvinick, M.M.; Cohen, J.D. The Neural Basis of Error Detection: Conflict Monitoring and the Error-Related Negativity. Psychol. Rev. 2004, 111, 931–959. [Google Scholar] [CrossRef] [PubMed]
- Nieuwenhuis, S.; Ridderinkhof, K.R.; Talsma, D.; Coles, M.G.H.; Holroyd, C.B.; Kok, A.; van der Molen, M.W. A computational account of altered error processing in older age: Dopamine and the error-related negativity. Cogn. Affect. Behav. Neurosci. 2002, 2, 19–36. [Google Scholar] [CrossRef] [PubMed]
- Falkenstein, M.; Hoormann, J.; Christ, S.; Hohnsbein, J. ERP components on reaction errors and their functional significance: A tutorial. Biol. Psychol. 2000, 51, 87–107. [Google Scholar] [CrossRef]
- Coles, M.G.; Scheffers, M.K.; Holroyd, C.B. Why is there an ERN/Ne on correct trials? Response representations, stimulus-related components, and the theory of error-processing. Biol. Psychol. 2001, 56, 173–189. [Google Scholar] [CrossRef]
- Holroyd, C.B.; Coles, M.G. The neural basis of human error processing: Reinforcement learning, dopamine, and the error-related negativity. Psychol Rev 2002, 109, 679–709. [Google Scholar] [CrossRef] [PubMed]
- Horowitz-Kraus, T. Improvement of the Error-detection Mechanism in Adults with Dyslexia Following Reading Acceleration Training. Dyslexia 2016, 22, 173–189. [Google Scholar] [CrossRef] [PubMed]
- Horowitz-Kraus, T. Does Development Affect the Error-Related Negativity of Impaired and Skilled Readers?: An ERP Study. Dev. Neuropsychol. 2011, 36, 914–932. [Google Scholar] [CrossRef] [PubMed]
- Horowitz-Kraus, T.; Breznitz, Z. Can the Error Detection Mechanism Benefit from Training the Working Memory? A Comparison between Dyslexics and Controls—An ERP Study. PLoS ONE 2009, 4, e7141. [Google Scholar]
- Horowitz-Kraus, T.; Breznitz, Z. An error-detection mechanism in reading among dyslexic and regular readers—An ERP study. Clin. Neurophysiol. 2008, 119, 2238–2246. [Google Scholar] [CrossRef] [PubMed]
- Horowitz-Kraus, T.; Breznitz, Z. Compensated Dyslexics Have a More Efficient Error Detection System Than Noncompensated Dyslexics. J. Child Neurol. 2012, 28, 1266–1276. [Google Scholar] [CrossRef] [PubMed]
- Sailer, U.; Fischmeister, F.P.S.; Bauer, H. Effects of learning on feedback-related brain potentials in a decision-making task. Brain Res. 2010, 1342, 85–93. [Google Scholar] [CrossRef] [PubMed]
- Polich, J. Updating P300: An integrative theory of P3a and P3b. Clin. Neurophysiol. 2007, 118, 2128–2148. [Google Scholar] [CrossRef] [PubMed]
- Bellebaum, C.; Daum, I. Learning-related changes in reward expectancy are reflected in the feedback-related negativity. Eur. J. Neurosci. 2008, 27, 1823–1835. [Google Scholar] [CrossRef] [PubMed]
- Groen, Y.; Wijers, A.A.; Mulder, L.J.M.; Minderaa, R.B.; Althaus, M. Physiological correlates of learning by performance feedback in children: A study of EEG event-related potentials and evoked heart rate. Biol. Psychol. 2007, 76, 174–187. [Google Scholar] [CrossRef] [PubMed]
- Kraus, D.; Horowitz-Kraus, T. The effect of learning on feedback-related potentials in adolescents with dyslexia: An EEG-ERP study. PLoS ONE 2014, 9, e100486. [Google Scholar] [CrossRef] [PubMed]
- Kruschke, J.K. Toward a Unified Model of Attention in Associative Learning. J. Math. Psychol. 2001, 45, 812–863. [Google Scholar] [CrossRef]
- Le Pelley, M.E. The role of associative history in models of associative learning: A selective review and a hybrid model. Q. J. Exp. Psychol. Sect. B 2010, 57, 193–243. [Google Scholar] [CrossRef] [PubMed]
- Facoetti, A.; Paganoni, P.; Turatto, M.; Marzola, V.; Mascetti, G.G. Visual-spatial attention in developmental dyslexia. Cortex 2000, 36, 109–123. [Google Scholar] [CrossRef]
- Valdois, S.; Bosse, M.-L.; Tainturier, M.-J. The cognitive deficits responsible for developmental dyslexia: Review of evidence for a selective visual attentional disorder. Dyslexia 2004, 10, 339–363. [Google Scholar] [CrossRef] [PubMed]
- Germanò, E.; Gagliano, A.; Curatolo, P. Comorbidity of ADHD and dyslexia. Dev. Neuropsychol. 2010, 35, 475–493. [Google Scholar] [CrossRef] [PubMed]
- Willcutt, E.G.; Pennington, B.F. Comorbidity of Reading Disability and Attention-Deficit/Hyperactivity Disorder: Differences by Gender and Subtype. J. Learn. Disabil. 2000, 33, 179–191. [Google Scholar] [CrossRef] [PubMed]
- Monzalvo, K.; Fluss, J.; Billard, C.; Dehaene, S.; Dehaene-Lambertz, G. Cortical networks for vision and language in dyslexic and normal children of variable socio-economic status. Neuroimage 2012, 61, 258–274. [Google Scholar] [CrossRef] [PubMed]
- Hashimoto, R.; Sakai, K.L. Learning letters in adulthood: Direct visualization of cortical plasticity for forming a new link between orthography and phonology. Neuron 2004, 42, 311–322. [Google Scholar] [CrossRef]
- Perrone-Bertolotti, M.; Vidal, J.R.; de Palma, L.; Hamamé, C.M.; Ossandon, T.; Kahane, P.; Minotti, L.; Bertrand, O.; Lachaux, J.P. Turning visual shapes into sounds: Early stages of reading acquisition revealed in the ventral occipitotemporal cortex. Neuroimage 2014, 90, 298–307. [Google Scholar] [CrossRef] [PubMed]
- Xue, G.; Chen, C.; Jin, Z.; Dong, Q. Language experience shapes fusiform activation when processing a logographic artificial language: An fMRI training study. Neuroimage 2006, 31, 1315–1326. [Google Scholar] [CrossRef] [PubMed]
- Mei, L.; Xue, G.; Lu, Z.L.; He, Q.; Zhang, M.; Xue, F.; Chen, C.; Dong, Q. Orthographic transparency modulates the functional asymmetry in the fusiform cortex: An artificial language training study. Brain Lang. 2013, 125, 165–172. [Google Scholar] [CrossRef] [PubMed]
- Price, C.J.; Devlin, J.T. The interactive account of ventral occipitotemporal contributions to reading. Trends Cogn. Sci. 2011, 15, 246–253. [Google Scholar] [CrossRef] [PubMed]
- Maurer, U.; Zevin, J.D.; McCandliss, B.D. Left-lateralized N170 effects of visual expertise in reading: Evidence from Japanese syllabic and logographic scripts. J. Cogn. Neurosci. 2008, 20, 1878–1891. [Google Scholar] [CrossRef] [PubMed]
- Richlan, F.; Kronbichler, M.; Wimmer, H. Meta-analyzing brain dysfunctions in dyslexic children and adults. Neuroimage 2011, 56, 1735–1742. [Google Scholar] [CrossRef] [PubMed]
- Kronschnabel, J.; Schmid, R.; Maurer, U.; Brandeis, D. Visual print tuning deficits in dyslexic adolescents under minimized phonological demands. Neuroimage 2013, 74, 58–69. [Google Scholar] [CrossRef] [PubMed]
- Crone, E.A.; Jennings, J.R.; van der Molen, M.W. Developmental change in feedback processing as reflected by phasic heart rate changes. Dev. Psychol. 2004, 40, 1228–1238. [Google Scholar] [CrossRef] [PubMed]
- Klimesch, W.; Doppelmayr, M.; Wimmer, H.; Gruber, W.; Röhm, D.; Schwaiger, J.; Hutzler, F. Alpha and beta band power changes in normal and dyslexic children. Clin. Neurophysiol. 2001, 112, 1186–1195. [Google Scholar] [CrossRef]
- Vourkas, M.; Micheloyannis, S.; Simos, P.G.; Rezaie, R.; Fletcher, J.M.; Cirino, P.T.; Papanicolaou, A.C. Dynamic task-specific brain network connectivity in children with severe reading difficulties. Neurosci. Lett. 2011, 488, 123–128. [Google Scholar] [CrossRef] [PubMed]
- Pennington, B.F.; Santerre–Lemmon, L.; Rosenberg, J.; MacDonald, B.; Boada, R.; Friend, A.; Leopold, D.R.; Samuelsson, S.; Byrne, B.; Willcutt, E.G.; et al. Individual prediction of dyslexia by single versus multiple deficit models. J. Abnorm. Psychol. 2012, 121, 212–214. [Google Scholar]
© 2017 by the authors; licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC-BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Fraga González, G.; Žarić, G.; Tijms, J.; Bonte, M.; Van der Molen, M.W. Contributions of Letter-Speech Sound Learning and Visual Print Tuning to Reading Improvement: Evidence from Brain Potential and Dyslexia Training Studies. Brain Sci. 2017, 7, 10. https://doi.org/10.3390/brainsci7010010
Fraga González G, Žarić G, Tijms J, Bonte M, Van der Molen MW. Contributions of Letter-Speech Sound Learning and Visual Print Tuning to Reading Improvement: Evidence from Brain Potential and Dyslexia Training Studies. Brain Sciences. 2017; 7(1):10. https://doi.org/10.3390/brainsci7010010
Chicago/Turabian StyleFraga González, Gorka, Gojko Žarić, Jurgen Tijms, Milene Bonte, and Maurits W. Van der Molen. 2017. "Contributions of Letter-Speech Sound Learning and Visual Print Tuning to Reading Improvement: Evidence from Brain Potential and Dyslexia Training Studies" Brain Sciences 7, no. 1: 10. https://doi.org/10.3390/brainsci7010010
APA StyleFraga González, G., Žarić, G., Tijms, J., Bonte, M., & Van der Molen, M. W. (2017). Contributions of Letter-Speech Sound Learning and Visual Print Tuning to Reading Improvement: Evidence from Brain Potential and Dyslexia Training Studies. Brain Sciences, 7(1), 10. https://doi.org/10.3390/brainsci7010010