
Emotion, Motivation, Reasoning, and How Their Brain Systems Are Related


Abstract
1. Introduction and Aims
2. A Theory of Emotion
2.1. A Definition and Theory of Emotion
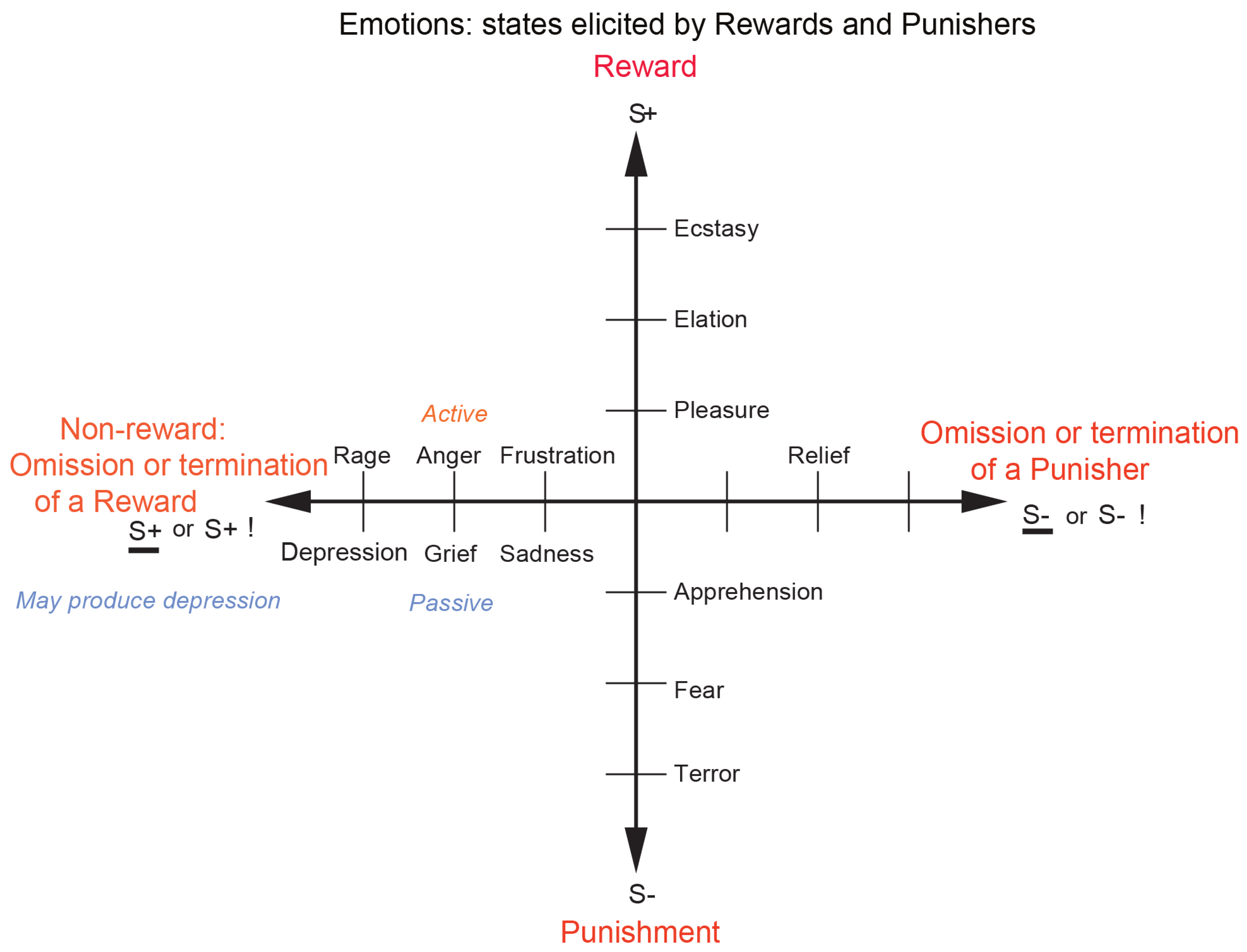
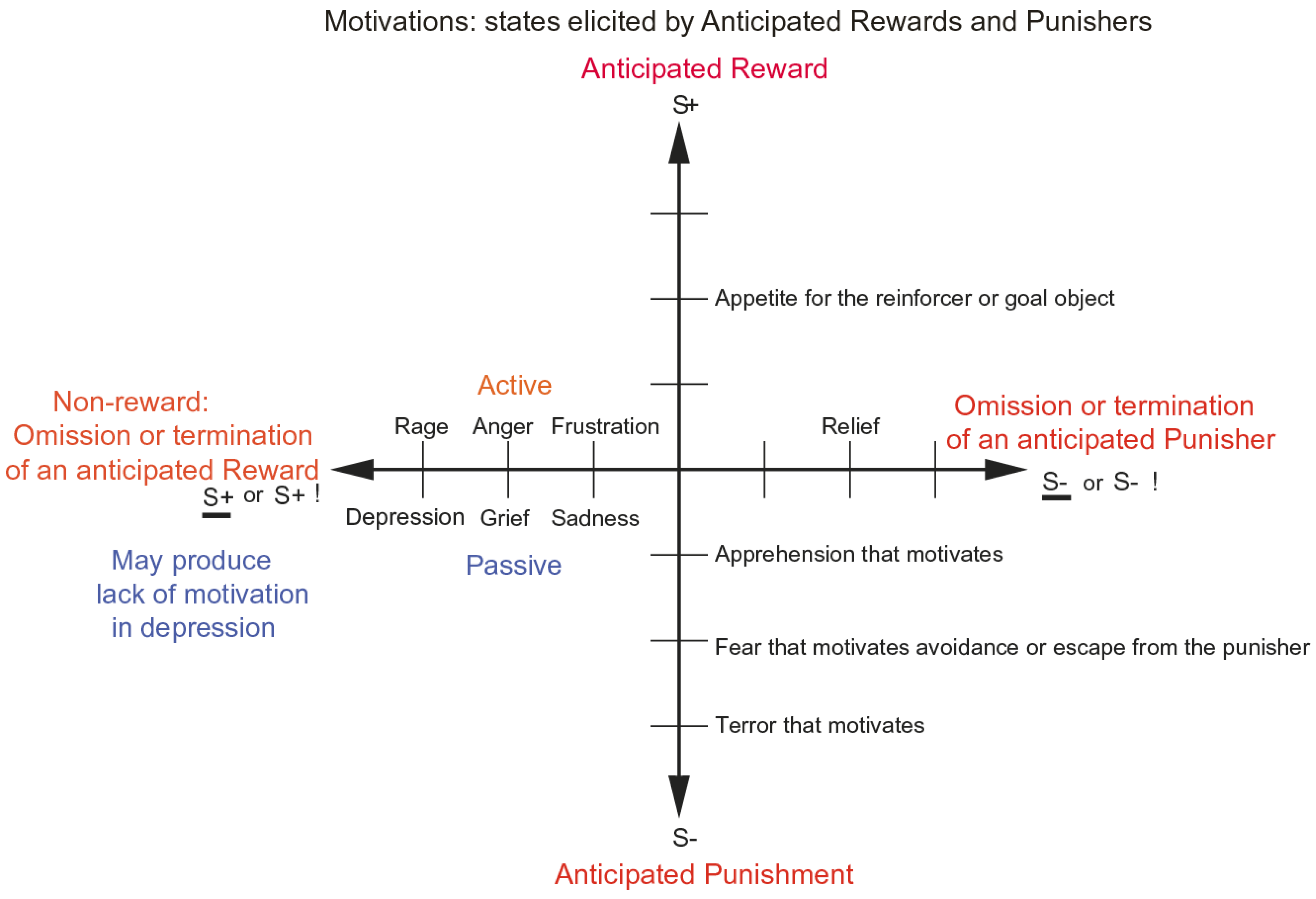
- The reinforcement contingency (e.g., whether a reward or punishment is given or withheld) (see Figure 1).
- The intensity of the reinforcer (see Figure 1).
- Any environmental stimulus might have a number of different reinforcement associations. (For example, a stimulus might be associated both with the presentation of a reward and a punisher, allowing states such as conflict and guilt to arise.)
- Emotions elicited by stimuli associated with different primary reinforcers will be different, as described above, for different primary reinforcers, each of which will produce different affective states, as shown in Table 1.
- Emotions elicited by different secondary reinforcing stimuli will be different from each other (even if the primary reinforcer is similar). For example, the same touch to the arm but by different people might give rise to very different emotions. Cognitive states and semantic knowledge can contribute to emotion in these ways, as well as in other ways that might arise because, for example, of reasoning in the rational brain system.
- The emotion elicited can depend on whether an active or passive behavioural response is possible. (For example, if an active behavioural response can occur due to the omission of a positive reinforcer, then anger might be produced, but if only passive behaviour is possible, then sadness, depression, or grief might occur (see Figure 1)).
2.2. The Functions of Emotions
3. A Framework for Understanding the Neuroscience of Emotion in Humans and Other Primates
3.1. A Framework for the Neuroscience of Emotion
3.2. The Human Medial Orbitofrontal Cortex Represents Reward Value
3.3. The Human Lateral Orbitofrontal Cortex Represents Punishers and Non-Rewards and Is Involved in Changing Emotional Behaviour
3.4. The Ventromedial Prefrontal Cortex and Reward-Related Decision-Making
3.5. The Amygdala
4. A Theory of Motivation and Brain Systems That Implement Motivation
4.1. The Outline of a Theory of Motivation
4.2. Motivational States Are States That Modulate the Reward or Punishment Value of Instrumental Reinforcers and Are Different from Taxes, Approach Responses, Classical Conditioning, and Stimulus–Response Habits
4.2.1. Taxes
4.2.2. Habit or Stimulus–Response Learning
4.2.3. Rewards and Punishers: Instrumental Goals for Action Towards Which Motivation Is Directed
4.2.4. Motivation, Instrumental, Action–Outcome, and Goal-Directed Learning
4.2.5. Gene-Specified Rewards and the Mechanisms of Evolution
4.2.6. Biological Economy in the Specification of Rewards and Punishers, for They Can Be Used to Implement Both Motivation and Emotion
4.2.7. Wanting vs. Liking and Goal-Directed Motivational Behaviour
5. Some Implications and Extensions of the Understanding of Emotion, Motivation, and Their Brain Mechanisms
5.1. Top-Down Cognitive Effects on Reward Value and Affective Responses, for Example, on the Reward Value and Pleasantness of Taste, Olfactory, and Flavour Stimuli
5.2. Effects of Top-Down Selective Attention to Affective Value Versus Intensity on Representations of Stimuli Including Those Involved in Taste, Olfactory, and Flavour Processing
5.3. Individual Differences in the Reward Systems, Evolution, and Personality
5.4. A Reasoning, Rational, Route to Action
5.4.1. Decisions Between the Emotional and Reasoning Systems
5.4.2. The Selfish Gene vs. the Selfish Phenotype and Evolution
5.4.3. A Higher-Order Syntactic Thought (HOST) Approach to Consciousness Including Emotional Feelings
5.5. The Neurology of Human Emotion: The Orbitofrontal Cortex Compared to the Amygdala
5.6. A Psychiatric Disorder of Emotion: Depression
5.7. Role of Reward and Emotion in Episodic and Semantic Memory and in Memory Consolidation
6. Comparison with Other Theories of Emotion
6.1. The James–Lange and Other Bodily Theories of Emotion Including Damasio’s Theory
6.2. Appraisal Theory
- (a)
- Relevance: How relevant is this event for me? Does it directly affect me or my social reference group?
- (b)
- Implications: What are the implications or consequences of this event, and how do they affect my well-being and my immediate or long-term goals?
- (c)
- Coping Potential: How well can I cope with or adjust to these consequences?
- (d)
- Normative Significance: What is the significance of this event for my self-concept and for social norms and values?
6.3. Panksepp’s Theory of Emotion
6.4. Dimensional, Categorical, and Other Theories of Emotion
7. Comparison with Other Theories of Motivation
7.1. Bindra’s Approach to Emotion and Motivation
7.2. The Development of Rolls’ Theory of Motivation
7.2.1. Internal Motivational States
7.2.2. Motivational States Elicited by Emotional States That Are Elicited by External Stimuli
7.3. Wanting vs. Liking
7.4. Emotion as Wanting
7.5. Evaluation
8. Conclusions and Highlights
- A new approach is further developed here to produce a unified understanding of emotion and motivation, with the underlying brain mechanisms considered in more detail elsewhere [6,7,8,12] but summarized next. In this unified theory of emotion and motivation, motivational states are states in which instrumental goal-directed actions are performed to obtain rewards or avoid punishers, and emotional states are states that are elicited when the reward or punisher is or is not received. This greatly simplifies our understanding of emotion and motivation, for the same set of genes and associated brain systems can define the primary or unlearned rewards and punishers such as a sweet taste or pain that can be used for both emotion and motivation. A very wide range of emotions can be understood by taking into account the reinforcement contingency, the particular primary reinforcer, the particular secondary reinforcer, the particular combination of reinforcers, the intensity of each reinforcer, etc., as specified above in Section 2.1 [6,7,8].
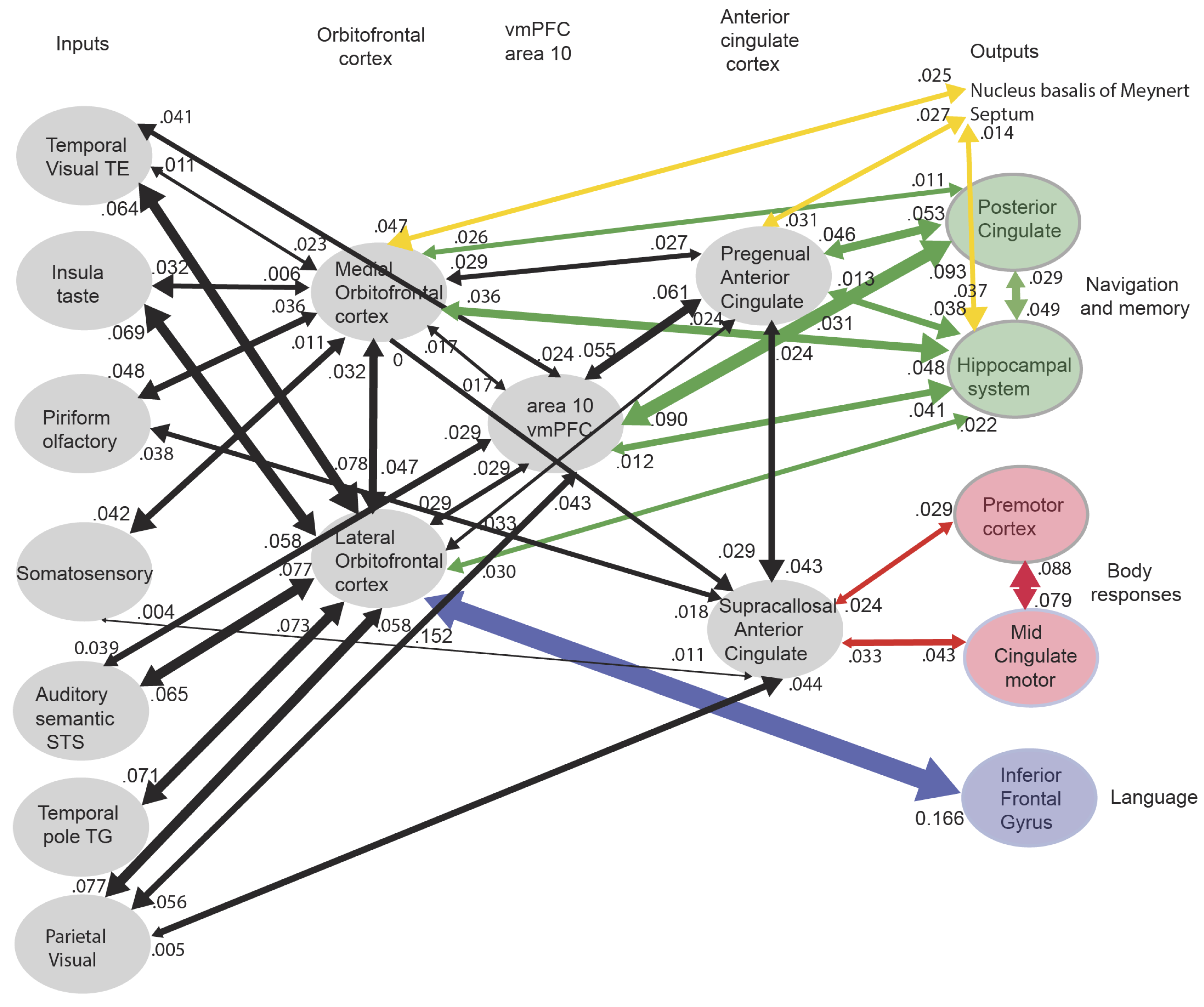
- New evidence on the connectivity in humans of brain systems involved in emotion and motivation is available from measures of the effective connectivity between 360 cortical regions in the Human Connectome Project MultiModal Parcellation atlas (HCP-MMP) [65] and is complemented by the addition of 66 subcortical regions [66]. The cortical regions in this atlas are defined by anatomical characteristics (cortical myelin content and cortical thickness), functional connectivity, and task-related fMRI and provide a useful basis for understanding brain regions with different connectivity and potentially different computational functions. Some of the following points reflect advances in our understanding of brain systems involved in emotion by taking into account the effective connectivity of the human brain, complemented by functional connectivity and diffusion tractography [20,61,67,68,184,213,270,271].
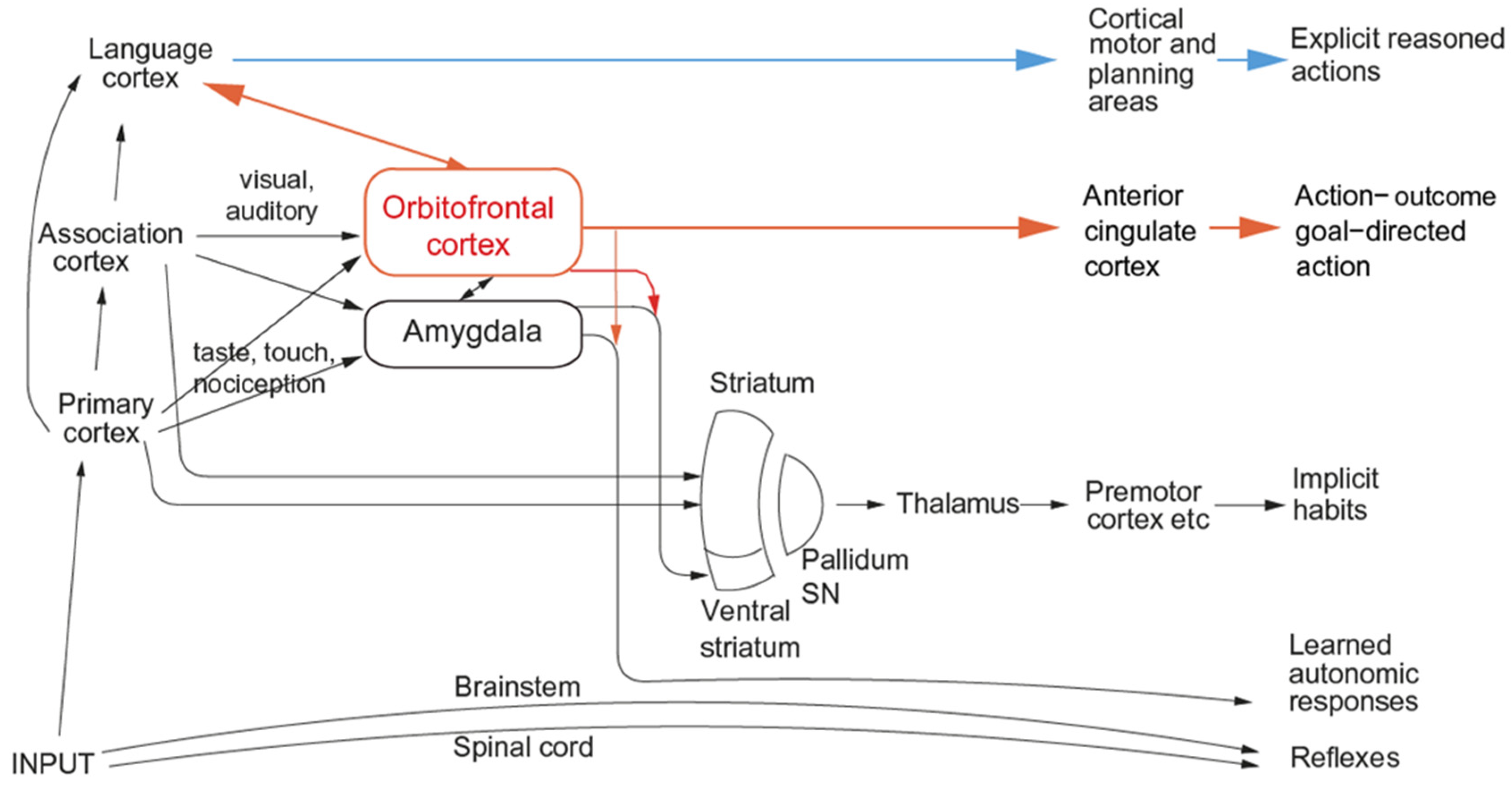
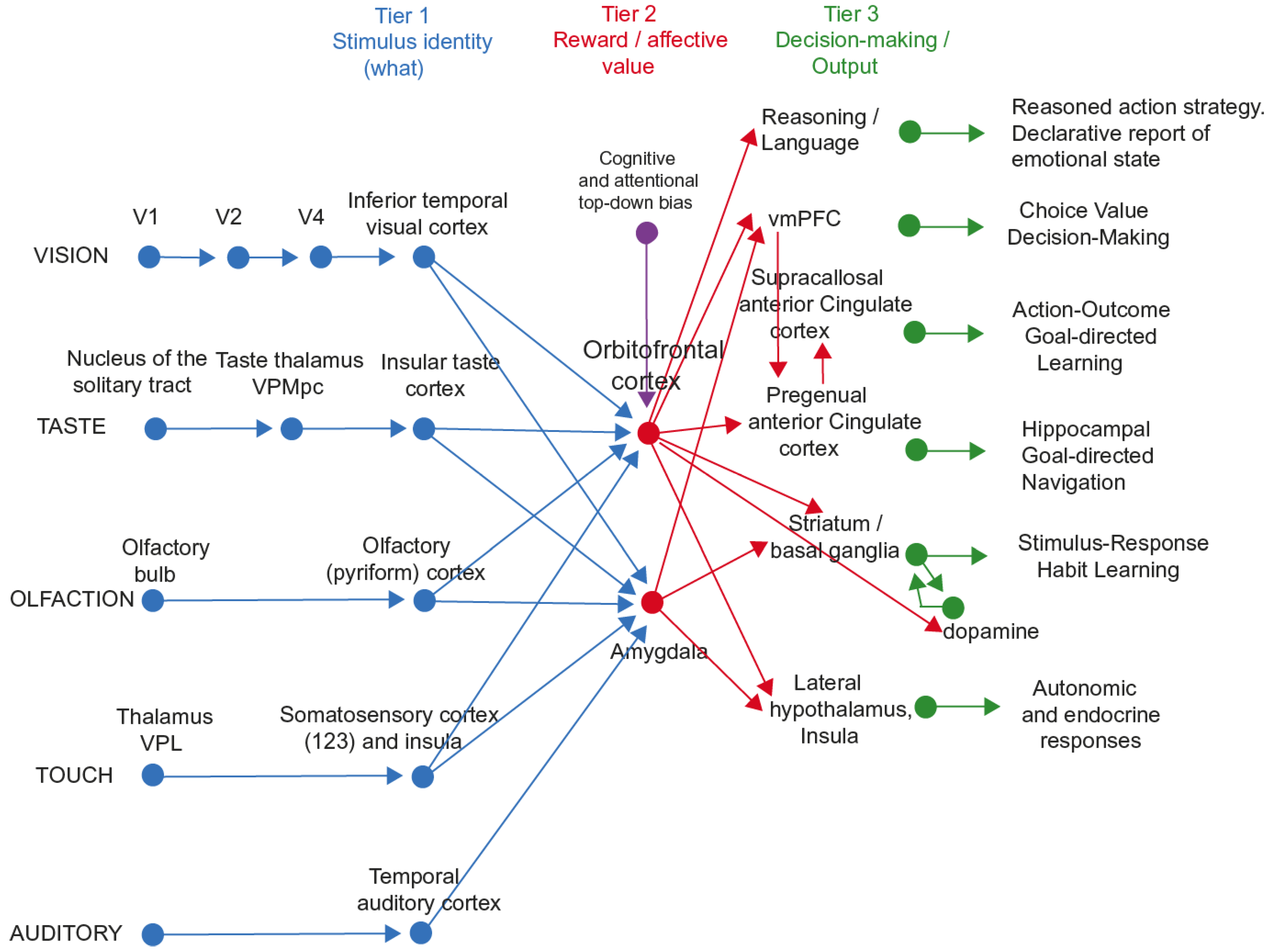
- It is shown that the primate including the human orbitofrontal cortex represents primary reinforcers such as taste, pain, and pleasant touch, with this information reaching the orbitofrontal cortex from the primary taste cortex in the anterior insula and from somatosensory cortical regions. It is shown that the primate including the human orbitofrontal cortex learns associations between these primary reinforcers and secondary reinforcers such as the sight of food or of an aversive stimulus in one trial and can reverse these associations in one trial using a rule- or model-based computation model. These stimulus–stimulus learned representations are of expected value (and can be thought of as predictions). The representations in the orbitofrontal cortex are value based and are appropriate for being the goals for motivated behaviour and for eliciting emotional states. Actions are not represented in the primate orbitofrontal cortex. Other inputs to the orbitofrontal cortex are about socially relevant stimuli such as face expression and face identity and relate to inputs from the cortex in the superior temporal sulcus. Rewards tend to be represented in the human medial orbitofrontal cortex and punishers and non-reward in the lateral orbitofrontal cortex. This evidence is complemented by the effects of damage to the orbitofrontal cortex in humans, which impairs reward-related reversal learning, emotional responses, and subjective emotional feelings. In primates including humans, reward and punisher value (the ‘valence’ of stimuli) is not represented in cortical stages of sensory processing prior to the orbitofrontal cortex, such as the insular primary taste cortex and inferior temporal visual cortex.
- The human medial and lateral orbitofrontal cortex and the vmPFC, have connectivity to the pregenual anterior cingulate cortex, which is strongly activated by rewards and which projects to the hippocampal system, both directly and via the posterior cingulate cortex (Figure 4) [20]. It is proposed that this provides the route for rewards and emotional states to become part of episodic memory. It is further proposed that the reward/emotional value of recalled episodic memories is important in influencing which memories are further processed and become incorporated into long-term semantic memory. It is further proposed that this route enables goals for navigation to enter the human hippocampal system, and indeed, navigation is almost always to obtain goals, which are reflected in hippocampal neuronal activity [272,273].
- The human pregenual anterior cingulate cortex has effective connectivity to the septum, from which cholinergic neurons important in memory consolidation project to the hippocampus (Figure 4) [20]. The human medial orbitofrontal cortex (region pOFC) has effective connectivity to the basal forebrain magnocellular nucleus of Meynert, from which cholinergic neurons important in memory consolidation project to the hippocampus (Figure 4) [20]. It is proposed that through these routes, the value system can influence memory consolidation. Consistent with this, damage to the vmPFC/anterior cingulate cortex in humans impairs memory. It is argued that the human orbitofrontal cortex/vmPFC/pregenual anterior cingulate cortex is not a memory system, but a value system, and that this value/emotion system influences memory and memory consolidation through these connectivities [64].
- The orbitofrontal cortex and pregenual anterior cingulate cortex have connectivity in humans to the supracallosal anterior cingulate cortex, which in turn has connectivity to premotor cortical regions including the midcingulate premotor cortex. It is proposed that these routes provide for action–outcome learning in the supracallosal anterior cingulate cortex, where the outcome is the reward or punisher received from the orbitofrontal cortex and pregenual anterior cingulate cortex [8,12].
- With this foundation, it is proposed that the function of the primate orbitofrontal cortex in emotion is to represent rewards and punishers and to implement stimulus–reward/punisher association learning and reversal (i.e., stimulus–stimulus learning). It is argued that in contrast, the role of the supracallosal anterior cingulate cortex is to learn associations between actions and the rewards/punishers that follow the actions, and with this action–outcome learning to influence the future choice of actions when reward/aversive expected value stimuli are received from the orbitofrontal cortex.
- It is shown that the human amygdala has effective connectivity from relatively few cortical regions, primarily those in the anterior temporal lobe, and even less effective connectivity back to the neocortex. The outputs of the human amygdala are directed primarily to brainstem regions involved in autonomic responses, cortical arousal, and some behavioural responses. In line with this, there is evidence that the human amygdala is much less involved in reported, experienced, declarative, emotion than the orbitofrontal cortex. This is a key re-evaluation of the functions of the human amygdala in emotion [61].
- It is shown that in addition to these emotion-related outputs to behaviour, in humans and perhaps in other animals, there is a rational, reasoning route to action that may override the genes selected during evolution to specify the rewards and punishers important in the control of goal-directed behaviour. The reasoning route to action may make choices in the interests of the individual and the phenotype, not in the interests of the gene-specified rewards, but it could also optimize the actions taken to obtain gene-specified rewards.
- Damage to the orbitofrontal cortex in humans can produce neurological changes such as reduced ability to respond correctly to emotion-relevant stimuli such as face and voice expression and to learn and change behaviour in response to reinforcement contingencies. It is shown that altered connectivity of the orbitofrontal cortex with other brain regions, and the sensitivity of the medial orbitofrontal cortex to rewards and of the lateral orbitofrontal cortex to punishers, is involved in human depression. Damage to the human orbitofrontal cortex, and much less to the amygdala, impairs human subjective emotional experiences and feelings.
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Rolls, E.T. The Brain and Emotion; Oxford University Press: Oxford, UK, 1999. [Google Scholar]
- Rolls, E.T. Precis of The Brain and Emotion. Behav. Brain Sci. 2000, 23, 177–191; discussion 192–233. [Google Scholar] [CrossRef]
- Weiskrantz, L. Emotion. In Analysis of Behavioural Change; Weiskrantz, L., Ed.; Harper and Row: New York, NY, USA; London, UK, 1968; pp. 50–90. [Google Scholar]
- Gray, J.A. Elements of a Two-Process Theory of Learning; Academic Press: London, UK, 1975. [Google Scholar]
- Rolls, E.T. Emotion Explained; Oxford University Press: Oxford, UK, 2005. [Google Scholar]
- Rolls, E.T. Emotion and Decision-Making Explained; Oxford University Press: Oxford, UK, 2014. [Google Scholar]
- Rolls, E.T. The Brain, Emotion, and Depression; Oxford University Press: Oxford, UK, 2018. [Google Scholar]
- Rolls, E.T. Emotion, motivation, decision-making, the orbitofrontal cortex, anterior cingulate cortex, and the amygdala. Brain Struct. Funct. 2023, 228, 1201–1257. [Google Scholar] [CrossRef]
- Zhang, B.; Rolls, E.T.; Wang, X.; Xie, C.; Cheng, W.; Feng, J. Roles of the medial and lateral orbitofrontal cortex in major depression and its treatment. Mol. Psychiatry 2024, 29, 914–928. [Google Scholar] [CrossRef] [PubMed]
- Rolls, E.T.; Cheng, W.; Feng, J. The orbitofrontal cortex: Reward, emotion, and depression. Brain Commun. 2020, 2, fcaa196. [Google Scholar] [CrossRef] [PubMed]
- Rolls, E.T. The orbitofrontal cortex and emotion in health and disease, including depression. Neuropsychologia 2019, 128, 14–43. [Google Scholar] [CrossRef] [PubMed]
- Rolls, E.T. Brain Computations and Connectivity; Oxford University Press: Oxford, UK, 2023. [Google Scholar]
- Rolls, E.T. Cerebral Cortex: Principles of Operation; Oxford University Press: Oxford, UK, 2016. [Google Scholar]
- Rolls, E.T. Brain Computations: What and How; Oxford University Press: Oxford, UK, 2021. [Google Scholar]
- Rolls, E.T. What are emotional states, and why do we have them? Emot. Rev. 2013, 5, 241–247. [Google Scholar] [CrossRef]
- Cardinal, N.; Parkinson, J.A.; Hall, J.; Everitt, B.J. Emotion and motivation: The role of the amygdala, ventral striatum, and prefrontal cortex. Neurosci. Biobehav. Rev. 2002, 26, 321–352. [Google Scholar] [CrossRef]
- Critchley, H.D.; Rolls, E.T. Responses of primate taste cortex neurons to the astringent tastant tannic acid. Chem. Senses 1996, 21, 135–145. [Google Scholar] [CrossRef]
- Adolphs, R.; Anderson, D.J. The Neuroscience of Emotion; Princeton University Press: Princeton, NJ, USA, 2018. [Google Scholar]
- Rolls, E.T. The cingulate cortex and limbic systems for emotion, action, and memory. Brain Struct. Funct. 2019, 224, 3001–3018. [Google Scholar] [CrossRef]
- Rolls, E.T.; Deco, G.; Huang, C.C.; Feng, J. The human orbitofrontal cortex, vmPFC, and anterior cingulate cortex effective connectome: Emotion, memory, and action. Cereb. Cortex 2022, 33, 330–356. [Google Scholar] [CrossRef]
- Dickinson, A. Actions and habits: The development of behavioural autonomy. Philos. Trans. R. Soc. Lond. B Biol. Sci. 1985, 308, 67–78. [Google Scholar]
- Dickinson, A. Instrumental conditioning revisited: Updating dual-process theory. Assoc. Learn. Cogn. 2016, 51, 177–195. [Google Scholar]
- LeDoux, J.E. Emotion circuits in the brain. Annu. Rev. Neurosci. 2000, 23, 155–184. [Google Scholar] [CrossRef]
- Rogan, M.T.; Staubli, U.V.; LeDoux, J.E. Fear conditioning induces associative long-term potentiation in the amygdala. Nature 1997, 390, 604–607. [Google Scholar] [CrossRef] [PubMed]
- Quirk, G.J.; Armony, J.L.; Repa, J.C.; Li, X.F.; LeDoux, J.E. Emotional memory: A search for sites of plasticity. Cold Spring Harb. Symp. Quant. Biol. 1996, 61, 247–257. [Google Scholar]
- LeDoux, J.E. Emotion: Clues from the brain. Annu. Rev. Psychol. 1995, 46, 209–235. [Google Scholar] [CrossRef]
- Davis, M. NMDA receptors and fear extinction: Implications for cognitive behavioral therapy. Dialogues Clin. Neurosci. 2011, 13, 463–474. [Google Scholar] [CrossRef] [PubMed]
- Antoniadis, E.A.; Winslow, J.T.; Davis, M.; Amaral, D.G. The nonhuman primate amygdala is necessary for the acquisition but not the retention of fear-potentiated startle. Biol. Psychiatry 2009, 65, 241–248. [Google Scholar] [CrossRef]
- Davis, M.; Antoniadis, E.A.; Amaral, D.G.; Winslow, J.T. Acoustic startle reflex in rhesus monkeys: A review. Rev. Neurosci. 2008, 19, 171–185. [Google Scholar] [CrossRef]
- Davis, M. Neural systems involved in fear and anxiety measured with fear-potentiated startle. Am. Psychol. 2006, 61, 741–756. [Google Scholar] [CrossRef]
- Davis, M.; Campeau, S.; Kim, M.; Falls, W.A. Neural systems and emotion: The amygdala’s role in fear and anxiety. In Brain and Memory: Modulation and Mediation of Neuroplasticity; McGaugh, J.L., Weinberger, N.M., Lynch, G., Eds.; Oxford University Press: New York, NY, USA, 1995. [Google Scholar]
- Davis, M. The role of the amygdala in emotional learning. Int. Rev. Neurobiol. 1994, 36, 225–266. [Google Scholar] [PubMed]
- Taschereau-Dumouchel, V.; Michel, M.; Lau, H.; Hofmann, S.G.; LeDoux, J.E. Putting the “mental” back in “mental disorders”: A perspective from research on fear and anxiety. Mol. Psychiatry 2022, 27, 1322–1330. [Google Scholar] [CrossRef] [PubMed]
- Mobbs, D.; Adolphs, R.; Fanselow, M.S.; Barrett, L.F.; LeDoux, J.E.; Ressler, K.; Tye, K.M. Viewpoints: Approaches to defining and investigating fear. Nat. Neurosci. 2019, 22, 1205–1216. [Google Scholar] [CrossRef] [PubMed]
- LeDoux, J.; Brown, R.; Pine, D.; Hofmann, S. Know thyself: Well-being and subjective experience. Cerebrum 2018, 2018, cer-01-18. Available online: https://www.ncbi.nlm.nih.gov/pmc/articles/PMC6353121/ (accessed on 12 March 2025).
- LeDoux, J.E.; Pine, D.S. Using neuroscience to help understand fear and anxiety: A two-system framework. Am. J. Psychiatry 2016, 173, 1083–1093. [Google Scholar] [CrossRef]
- LeDoux, J. Low roads and higher order thoughts in emotion. Cortex 2014, 59, 214–215. [Google Scholar] [CrossRef]
- LeDoux, J. Rethinking the emotional brain. Neuron 2012, 73, 653–676. [Google Scholar] [CrossRef]
- Rolls, E.T. Neural computations underlying phenomenal consciousness: A Higher Order Syntactic Thought theory. Front. Psychol. (Conscious. Res.) 2020, 11, 655. [Google Scholar] [CrossRef]
- Brown, R.; Lau, H.; LeDoux, J.E. Understanding the Higher-Order Approach to Consciousness. Trends Cogn. Sci. 2019, 23, 754–768. [Google Scholar] [CrossRef]
- Rosenthal, D.M. Varieties of Higher-Order Theory. In Higher Order Theories of Consciousness; Gennaro, R.J., Ed.; John Benjamins: Amsterdam, The Netherlands, 2004; pp. 17–44. [Google Scholar]
- Rolls, E.T. Neuroculture: On the Implications of Brain Science; Oxford University Press: Oxford, UK, 2012. [Google Scholar]
- Rolls, E.T. A computational neuroscience approach to consciousness. Neural Netw. 2007, 20, 962–982. [Google Scholar] [CrossRef]
- Grabenhorst, F.; Rolls, E.T. Value, pleasure, and choice in the ventral prefrontal cortex. Trends Cogn. Sci. 2011, 15, 56–67. [Google Scholar] [CrossRef]
- Rolls, E.T.; Grabenhorst, F. The orbitofrontal cortex and beyond: From affect to decision-making. Prog. Neurobiol. 2008, 86, 216–244. [Google Scholar] [CrossRef] [PubMed]
- Rushworth, M.F.; Noonan, M.P.; Boorman, E.D.; Walton, M.E.; Behrens, T.E. Frontal cortex and reward-guided learning and decision-making. Neuron 2011, 70, 1054–1069. [Google Scholar] [CrossRef] [PubMed]
- Rushworth, M.F.; Kolling, N.; Sallet, J.; Mars, R.B. Valuation and decision-making in frontal cortex: One or many serial or parallel systems? Curr. Opin. Neurobiol. 2012, 22, 946–955. [Google Scholar] [CrossRef]
- Schultz, W. Reward prediction error. Curr. Biol. 2017, 27, R369–R371. [Google Scholar] [CrossRef]
- Schultz, W. Reward functions of the basal ganglia. J. Neural Transm. 2016, 123, 679–693. [Google Scholar] [CrossRef]
- Schultz, W. Dopamine reward prediction-error signalling: A two-component response. Nat. Rev. Neurosci. 2016, 17, 183–195. [Google Scholar] [CrossRef] [PubMed]
- Rolls, E.T. The roles of the orbitofrontal cortex via the habenula in non-reward and depression, and in the responses of serotonin and dopamine neurons. Neurosci. Biobehav. Rev. 2017, 75, 331–334. [Google Scholar] [CrossRef]
- Rolls, E.T. The Orbitofrontal Cortex; Oxford University Press: Oxford, UK, 2019. [Google Scholar]
- Critchley, H.D.; Harrison, N.A. Visceral influences on brain and behavior. Neuron 2013, 77, 624–638. [Google Scholar] [CrossRef]
- Quadt, L.; Critchley, H.; Nagai, Y. Cognition, emotion, and the central autonomic network. Auton. Neurosci. 2022, 238, 102948. [Google Scholar] [CrossRef]
- Johansen, J.P.; Tarpley, J.W.; LeDoux, J.E.; Blair, H.T. Neural substrates for expectation-modulated fear learning in the amygdala and periaqueductal gray. Nat. Neurosci. 2010, 13, 979–986. [Google Scholar] [CrossRef]
- LeDoux, J.E.; Iwata, J.; Cicchetti, P.; Reis, D.J. Different projections of the central amygdaloid nucleus mediate autonomic and behavioral correlates of conditioned fear. J. Neurosci. 1988, 8, 2517–2529. [Google Scholar] [CrossRef]
- Delgado, M.R.; Jou, R.L.; Phelps, E.A. Neural systems underlying aversive conditioning in humans with primary and secondary reinforcers. Front. Neurosci. 2011, 5, 71. [Google Scholar] [CrossRef] [PubMed]
- Phelps, E.A. Emotion and cognition: Insights from studies of the human amygdala. Annu. Rev. Psychol. 2006, 57, 27–53. [Google Scholar] [CrossRef]
- Delgado, M.R.; Olsson, A.; Phelps, E.A. Extending animal models of fear conditioning to humans. Biol. Psychol. 2006, 73, 39–48. [Google Scholar] [CrossRef] [PubMed]
- Phelps, E.A.; LeDoux, J.E. Contributions of the amygdala to emotion processing: From animal models to human behavior. Neuron 2005, 48, 175–187. [Google Scholar] [CrossRef]
- Rolls, E.T.; Deco, G.; Huang, C.-C.; Feng, J. Human amygdala compared to orbitofrontal cortex connectivity, and emotion. Prog. Neurobiol. 2023, 220, 102385. [Google Scholar] [CrossRef] [PubMed]
- Luo, Q.; Ge, T.; Grabenhorst, F.; Feng, J.; Rolls, E.T. Attention-dependent modulation of cortical taste circuits revealed by Granger causality with signal-dependent noise. PLoS Comput. Biol. 2013, 9, e1003265. [Google Scholar]
- Rolls, E.T. Emotion, higher order syntactic thoughts, and consciousness. In Frontiers of Consciousness; Weiskrantz, L., Davies, M.K., Eds.; Oxford University Press: Oxford, UK, 2008; pp. 131–167. [Google Scholar]
- Rolls, E.T. The hippocampus, ventromedial prefrontal cortex, and episodic and semantic memory. Prog. Neurobiol. 2022, 217, 102334. [Google Scholar] [CrossRef]
- Glasser, M.F.; Coalson, T.S.; Robinson, E.C.; Hacker, C.D.; Harwell, J.; Yacoub, E.; Ugurbil, K.; Andersson, J.; Beckmann, C.F.; Jenkinson, M.; et al. A multi-modal parcellation of human cerebral cortex. Nature 2016, 536, 171–178. [Google Scholar] [CrossRef]
- Huang, C.C.; Rolls, E.T.; Feng, J.; Lin, C.P. An extended Human Connectome Project multimodal parcellation atlas of the human cortex and subcortical areas. Brain Struct. Funct. 2022, 227, 763–778. [Google Scholar] [CrossRef]
- Rolls, E.T.; Deco, G.; Huang, C.-C.; Feng, J. The human language effective connectome. Neuroimage 2022, 258, 119352. [Google Scholar] [CrossRef] [PubMed]
- Rolls, E.T.; Deco, G.; Huang, C.C.; Feng, J. The effective connectivity of the human hippocampal memory system. Cereb. Cortex 2022, 32, 3706–3725. [Google Scholar] [CrossRef]
- Rolls, E.T. Emotion and Decision-Making Explained: Response to commentators. Cortex 2015, 62, 203–210. [Google Scholar] [CrossRef] [PubMed]
- Rolls, E.T. Emotion and Decision-Making Explained: Précis. Cortex 2014, 59, 185–193. [Google Scholar] [CrossRef]
- Rolls, E.T. The Orbitofrontal Cortex; Oxford Research Encyclopedia of Neuroscience; ORE-NEU_00560; Oxford University Press: Oxford, UK, 2025. [Google Scholar]
- Rolls, E.T. The neuroscience of emotional disorders. In Handbook of Clinical Neurology: Disorders of Emotion in Neurologic Disease; Heilman, K.M., Nadeau, S.E., Eds.; Elsevier: Oxford, UK, 2021; Volume 183, pp. 1–26. [Google Scholar]
- Passingham, R.E. Understanding the Prefrontal Cortex: Selective Advantage, Connectivity and Neural Operations; Oxford University Press: Oxford, UK, 2021. [Google Scholar]
- Rolls, E.T.; Zhang, C.; Feng, J. Reward-specific satiety and reward-specific motivation: Neural bases. 2025. [Google Scholar]
- Rolls, E.T. Neuroscience Discoveries; Oxford University Press: Oxford, UK, 2025. [Google Scholar]
- Rolls, E.T.; Murzi, E.; Yaxley, S.; Thorpe, S.J.; Simpson, S.J. Sensory-specific satiety: Food-specific reduction in responsiveness of ventral forebrain neurons after feeding in the monkey. Brain Res. 1986, 368, 79–86. [Google Scholar] [CrossRef] [PubMed]
- Critchley, H.D.; Rolls, E.T. Hunger and satiety modify the responses of olfactory and visual neurons in the primate orbitofrontal cortex. J. Neurophysiol. 1996, 75, 1673–1686. [Google Scholar] [CrossRef]
- Rolls, E.T. Reward systems in the brain and nutrition. Annu. Rev. Nutr. 2016, 36, 435–470. [Google Scholar] [CrossRef]
- Thorpe, S.J.; Rolls, E.T.; Maddison, S. The orbitofrontal cortex: Neuronal activity in the behaving monkey. Exp. Brain Res. 1983, 49, 93–115. [Google Scholar] [CrossRef]
- Rolls, E.T.; Critchley, H.D.; Mason, R.; Wakeman, E.A. Orbitofrontal cortex neurons: Role in olfactory and visual association learning. J. Neurophysiol. 1996, 75, 1970–1981. [Google Scholar] [CrossRef]
- Rolls, E.T.; Deco, G. Non-reward neural mechanisms in the orbitofrontal cortex. Cortex 2016, 83, 27–38. [Google Scholar] [CrossRef]
- Deco, G.; Rolls, E.T. Synaptic and spiking dynamics underlying reward reversal in orbitofrontal cortex. Cereb. Cortex 2005, 15, 15–30. [Google Scholar] [CrossRef] [PubMed]
- Hervig, M.E.; Fiddian, L.; Piilgaard, L.; Bozic, T.; Blanco-Pozo, M.; Knudsen, C.; Olesen, S.F.; Alsio, J.; Robbins, T.W. Dissociable and paradoxical roles of rat medial and lateral orbitofrontal cortex in visual serial reversal learning. Cereb. Cortex 2020, 30, 1016–1029. [Google Scholar] [CrossRef] [PubMed]
- Rolls, E.T.; Critchley, H.D.; Browning, A.S.; Inoue, K. Face-selective and auditory neurons in the primate orbitofrontal cortex. Exp. Brain Res. 2006, 170, 74–87. [Google Scholar] [CrossRef] [PubMed]
- Hasselmo, M.E.; Rolls, E.T.; Baylis, G.C.; Nalwa, V. Object-centred encoding by face-selective neurons in the cortex in the superior temporal sulcus of the monkey. Exp. Brain Res. 1989, 75, 417–429. [Google Scholar] [CrossRef]
- Hasselmo, M.E.; Rolls, E.T.; Baylis, G.C. The role of expression and identity in the face-selective responses of neurons in the temporal visual cortex of the monkey. Behav. Brain Res. 1989, 32, 203–218. [Google Scholar] [CrossRef]
- Pitcher, D.; Ungerleider, L.G. Evidence for a third visual pathway specialized for social perception. Trends Cogn. Sci. 2021, 25, 100–110. [Google Scholar] [CrossRef]
- Pitcher, D.; Ianni, G.; Ungerleider, L.G. A functional dissociation of face-, body- and scene-selective brain areas based on their response to moving and static stimuli. Sci. Rep. 2019, 9, 8242. [Google Scholar] [CrossRef]
- Kringelbach, M.L.; O’Doherty, J.; Rolls, E.T.; Andrews, C. Activation of the human orbitofrontal cortex to a liquid food stimulus is correlated with its subjective pleasantness. Cereb. Cortex 2003, 13, 1064–1071. [Google Scholar] [CrossRef]
- De Araujo, I.E.; Rolls, E.T.; Kringelbach, M.L.; McGlone, F.; Phillips, N. Taste-olfactory convergence, and the representation of the pleasantness of flavour, in the human brain. Eur. J. Neurosci. 2003, 18, 2059–2068. [Google Scholar] [CrossRef]
- Grabenhorst, F.; Rolls, E.T.; Bilderbeck, A. How cognition modulates affective responses to taste and flavor: Top down influences on the orbitofrontal and pregenual cingulate cortices. Cereb. Cortex 2008, 18, 1549–1559. [Google Scholar] [CrossRef] [PubMed]
- Grabenhorst, F.; Rolls, E.T. Selective attention to affective value alters how the brain processes taste stimuli. Eur. J. Neurosci. 2008, 27, 723–729. [Google Scholar] [CrossRef]
- Kringelbach, M.L.; Rolls, E.T. Neural correlates of rapid reversal learning in a simple model of human social interaction. Neuroimage 2003, 20, 1371–1383. [Google Scholar] [CrossRef] [PubMed]
- O’Doherty, J.; Kringelbach, M.L.; Rolls, E.T.; Hornak, J.; Andrews, C. Abstract reward and punishment representations in the human orbitofrontal cortex. Nat. Neurosci. 2001, 4, 95–102. [Google Scholar] [CrossRef] [PubMed]
- Xie, C.; Jia, T.; Rolls, E.T.; Robbins, T.W.; Sahakian, B.J.; Zhang, J.; Liu, Z.; Cheng, W.; Luo, Q.; Zac Lo, C.Y.; et al. Reward versus nonreward sensitivity of the medial versus lateral orbitofrontal cortex relates to the severity of depressive symptoms. Biol. Psychiatry Cogn. Neurosci. Neuroimaging 2021, 6, 259–269. [Google Scholar] [CrossRef]
- Rolls, E.T.; Vatansever, D.; Li, Y.; Cheng, W.; Feng, J. Rapid rule-based reward reversal and the lateral orbitofrontal cortex. Cereb. Cortex Commun. 2020, 1, tgaa087. [Google Scholar] [CrossRef]
- O’Doherty, J.; Winston, J.; Critchley, H.; Perrett, D.; Burt, D.M.; Dolan, R.J. Beauty in a smile: The role of medial orbitofrontal cortex in facial attractiveness. Neuropsychologia 2003, 41, 147–155. [Google Scholar] [CrossRef]
- Rolls, E.T.; Kringelbach, M.L.; De Araujo, I.E.T. Different representations of pleasant and unpleasant odors in the human brain. Eur. J. Neurosci. 2003, 18, 695–703. [Google Scholar] [CrossRef]
- Rolls, E.T.; O’Doherty, J.; Kringelbach, M.L.; Francis, S.; Bowtell, R.; McGlone, F. Representations of pleasant and painful touch in the human orbitofrontal and cingulate cortices. Cereb. Cortex 2003, 13, 308–317. [Google Scholar] [CrossRef]
- McCabe, C.; Rolls, E.T.; Bilderbeck, A.; McGlone, F. Cognitive influences on the affective representation of touch and the sight of touch in the human brain. Soc. Cogn. Affect. Neurosci. 2008, 3, 97–108. [Google Scholar] [CrossRef]
- Völlm, B.A.; de Araujo, I.E.T.; Cowen, P.J.; Rolls, E.T.; Kringelbach, M.L.; Smith, K.A.; Jezzard, P.; Heal, R.J.; Matthews, P.M. Methamphetamine activates reward circuitry in drug naïve human subjects. Neuropsychopharmacology 2004, 29, 1715–1722. [Google Scholar] [CrossRef]
- Hornak, J.; Bramham, J.; Rolls, E.T.; Morris, R.G.; O’Doherty, J.; Bullock, P.R.; Polkey, C.E. Changes in emotion after circumscribed surgical lesions of the orbitofrontal and cingulate cortices. Brain 2003, 126, 1691–1712. [Google Scholar] [CrossRef] [PubMed]
- Rolls, E.T.; Hornak, J.; Wade, D.; McGrath, J. Emotion-related learning in patients with social and emotional changes associated with frontal lobe damage. J. Neurol. Neurosurg. Psychiatry 1994, 57, 1518–1524. [Google Scholar] [CrossRef]
- Hornak, J.; Rolls, E.T.; Wade, D. Face and voice expression identification in patients with emotional and behavioural changes following ventral frontal lobe damage. Neuropsychologia 1996, 34, 247–261. [Google Scholar] [CrossRef]
- Hornak, J.; O’Doherty, J.; Bramham, J.; Rolls, E.T.; Morris, R.G.; Bullock, P.R.; Polkey, C.E. Reward-related reversal learning after surgical excisions in orbitofrontal and dorsolateral prefrontal cortex in humans. J. Cogn. Neurosci. 2004, 16, 463–478. [Google Scholar] [CrossRef] [PubMed]
- Berlin, H.; Rolls, E.T.; Kischka, U. Impulsivity, time perception, emotion, and reinforcement sensitivity in patients with orbitofrontal cortex lesions. Brain 2004, 127, 1108–1126. [Google Scholar] [CrossRef]
- Berlin, H.; Rolls, E.T. Time perception, impulsivity, emotionality, and personality in self-harming Borderline Personality Disorder Patients. J. Personal. Disord. 2004, 18, 358–378. [Google Scholar] [CrossRef]
- Du, J.; Rolls, E.T.; Cheng, W.; Li, Y.; Gong, W.; Qiu, J.; Feng, J. Functional connectivity of the orbitofrontal cortex, anterior cingulate cortex, and inferior frontal gyrus in humans. Cortex 2020, 123, 185–199. [Google Scholar] [CrossRef]
- Bechara, A.; Damasio, H.; Tranel, D.; Damasio, A.R. The Iowa Gambling Task and the somatic marker hypothesis: Some questions and answers. Trends Cogn. Sci. 2005, 9, 159–162; discussion 162–164. [Google Scholar] [CrossRef]
- Bechara, A.; Damasio, H.; Tranel, D.; Damasio, A.R. Deciding advantageously before knowing the advantageous strategy. Science 1997, 275, 1293–1295. [Google Scholar] [CrossRef]
- Glascher, J.; Adolphs, R.; Damasio, H.; Bechara, A.; Rudrauf, D.; Calamia, M.; Paul, L.K.; Tranel, D. Lesion mapping of cognitive control and value-based decision making in the prefrontal cortex. Proc. Natl. Acad. Sci. USA 2012, 109, 14681–14686. [Google Scholar] [CrossRef]
- Grabenhorst, F.; Rolls, E.T.; Parris, B.A. From affective value to decision-making in the prefrontal cortex. Eur. J. Neurosci. 2008, 28, 1930–1939. [Google Scholar] [CrossRef]
- Rolls, E.T.; Grabenhorst, F.; Deco, G. Decision-making, errors, and confidence in the brain. J. Neurophysiol. 2010, 104, 2359–2374. [Google Scholar] [CrossRef] [PubMed]
- Rolls, E.T.; Grabenhorst, F.; Deco, G. Choice, difficulty, and confidence in the brain. Neuroimage 2010, 53, 694–706. [Google Scholar] [CrossRef]
- Passingham, R.E.P.; Wise, S.P. The Neurobiology of the Prefrontal Cortex; Oxford University Press: Oxford, UK, 2012. [Google Scholar]
- LeDoux, J.E. The Emotional Brain; Simon and Schuster: New York, NY, USA, 1996. [Google Scholar]
- LeDoux, J.E. The amygdala, fear conditioning and emotion. In The Amygdala: A Functional Analysis; Aggleton, J.P., Ed.; Oxford University Press: Oxford, UK, 2000. [Google Scholar]
- Davis, M. The role of the amygdala in conditioned fear. In The Amygdala; Aggleton, J.P., Ed.; Wiley-Liss: New York, NY, USA, 1992; pp. 255–305. [Google Scholar]
- Whalen, P.J.; Phelps, E.A. (Eds.) The Human Amygdala; Guilford: New York, NY, USA, 2009. [Google Scholar]
- Feinstein, J.S.; Adolphs, R.; Damasio, A.; Tranel, D. The human amygdala and the induction and experience of fear. Curr. Biol. 2011, 21, 34–38. [Google Scholar] [CrossRef] [PubMed]
- Damasio, A.; Damasio, H.; Tranel, D. Persistence of feelings and sentience after bilateral damage of the insula. Cereb. Cortex 2013, 23, 833–846. [Google Scholar] [CrossRef]
- Adolphs, R.; Gosselin, F.; Buchanan, T.W.; Tranel, D.; Schyns, P.; Damasio, A.R. A mechanism for impaired fear recognition after amygdala damage. Nature 2005, 433, 68–72. [Google Scholar] [CrossRef] [PubMed]
- Kennedy, D.P.; Adolphs, R. Reprint of: Impaired fixation to eyes following amygdala damage arises from abnormal bottom-up attention. Neuropsychologia 2011, 49, 589–595. [Google Scholar] [CrossRef]
- LeDoux, J.E.; Brown, R. A higher-order theory of emotional consciousness. Proc. Natl. Acad. Sci. USA 2017, 114, E2016–E2025. [Google Scholar] [CrossRef]
- LeDoux, J.E. Thoughtful feelings. Curr. Biol. 2020, 30, R619–R623. [Google Scholar] [CrossRef]
- Fellows, L.K. Orbitofrontal contributions to value-based decision making: Evidence from humans with frontal lobe damage. Ann. N. Y. Acad. Sci. 2011, 1239, 51–58. [Google Scholar] [CrossRef]
- Camille, N.; Tsuchida, A.; Fellows, L.K. Double dissociation of stimulus-value and action-value learning in humans with orbitofrontal or anterior cingulate cortex damage. J. Neurosci. 2011, 31, 15048–15052. [Google Scholar] [CrossRef] [PubMed]
- Klein-Flugge, M.C.; Jensen, D.E.A.; Takagi, Y.; Priestley, L.; Verhagen, L.; Smith, S.M.; Rushworth, M.F.S. Relationship between nuclei-specific amygdala connectivity and mental health dimensions in humans. Nat. Hum. Behav. 2022, 6, 1705–1722. [Google Scholar] [CrossRef]
- Damasio, A.R. The somatic marker hypothesis and the possible functions of the prefrontal cortex. Philos. Trans. R. Soc. Lond. B Biol. Sci. 1996, 351, 1413–1420. [Google Scholar] [CrossRef]
- Damasio, A.R. Descartes’ Error; Putnam: New York, NY, USA, 1994. [Google Scholar]
- Rolls, E.T. Motivation Explained: Ultimate and proximate accounts of hunger and appetite. Adv. Motiv. Sci. 2016, 3, 187–249. [Google Scholar]
- Rolls, B.J.; Rolls, E.T. The control of drinking. Br. Med. Bull. 1981, 37, 127–130. [Google Scholar] [CrossRef] [PubMed]
- Rolls, B.J.; Wood, R.J.; Rolls, E.T.; Lind, H.; Lind, R.; Ledingham, J. Thirst following water deprivation in humans. Am. J. Physiol. 1980, 239, R476–R482. [Google Scholar] [CrossRef]
- Rolls, B.J.; Wood, R.J.; Rolls, E.T. Thirst: The initiation, maintenance, and termination of drinking. Prog. Psychobiol. Physiol. Psychol. 1980, 9, 263–321. [Google Scholar]
- Schultz, W. Dopamine reward prediction error coding. Dialogues Clin. Neurosci. 2016, 18, 23–32. [Google Scholar] [CrossRef]
- Teitelbaum, P. The biology of drive. In Readings in Behavior; Ven der Kloot, W., Walcott, C., Dabe, B., Eds.; Holt, Rinehart, Winston: New York, NY, USA, 1974; p. 199. [Google Scholar]
- Grabenhorst, F.; D’Souza, A.A.; Parris, B.A.; Rolls, E.T.; Passingham, R.E. A common neural scale for the subjective pleasantness of different primary rewards. Neuroimage 2010, 51, 1265–1274. [Google Scholar] [CrossRef]
- Rolls, E.T.; Browning, A.S.; Inoue, K.; Hernadi, S. Novel visual stimuli activate a population of neurons in the primate orbitofrontal cortex. Neurobiol. Learn. Mem. 2005, 84, 111–123. [Google Scholar] [CrossRef]
- Kehoe, P.; Blass, E.M. Gustatory determinants of suckling in albino rats 5-20 days of age. Dev. Psychobiol. 1985, 18, 67–82. [Google Scholar] [CrossRef]
- Rozin, P.; Kalat, J.W. Specific hungers and poison avoidance as adaptive specializations of learning. Psychol. Rev. 1971, 78, 459–486. [Google Scholar] [CrossRef]
- Rolls, E.T. The orbitofrontal cortex, food reward, body weight and obesity. Soc. Cogn. Affect. Neurosci. 2023, 18, nsab044. [Google Scholar] [CrossRef] [PubMed]
- Berthoud, H.R.; Morrison, C.D.; Ackroff, K.; Sclafani, A. Learning of food preferences: Mechanisms and implications for obesity and metabolic diseases. Int. J. Obes. 2021, 45, 2156–2168. [Google Scholar] [CrossRef] [PubMed]
- Deutsch, J.A. The Structural Basis of Behavior; University of Chicago Press: Chicago, IL, USA, 1960. [Google Scholar]
- Sternson, S.M. Hypothalamic survival circuits: Blueprints for purposive behaviors. Neuron 2013, 77, 810–824. [Google Scholar] [CrossRef] [PubMed]
- Rolls, B.J.; Rolls, E.T. Thirst; Cambridge University Press: Cambridge, UK, 1982. [Google Scholar]
- de Araujo, I.E.T.; Kringelbach, M.L.; Rolls, E.T.; McGlone, F. Human cortical responses to water in the mouth, and the effects of thirst. J. Neurophysiol. 2003, 90, 1865–1876. [Google Scholar] [CrossRef]
- Buss, D.M. Sex differences in human mate preferences: Evoluntionary hypotheses tested in 37 cultures. Behav. Brain Sci. 1989, 12, 1–14. [Google Scholar] [CrossRef]
- Buss, D.M. Evolutionary Psychology: The New Science of the Mind, 5th ed.; Pearson: New York, NY, USA, 2015. [Google Scholar]
- Dawkins, R. The Blind Watchmaker; Longman: Harlow, UK, 1986. [Google Scholar]
- Dawkins, R. The Extended Phenotype: The Gene as the Unit of Selection; W.H. Freeman: San Francisco, CA, USA, 1982. [Google Scholar]
- Dawkins, R. The Selfish Gene; Oxford University Press: Oxford, UK, 1976. [Google Scholar]
- Berridge, K.C.; Robinson, T.E.; Aldridge, J.W. Dissecting components of reward: ‘liking’, ‘wanting’, and learning. Curr. Opin. Pharmacol. 2009, 9, 65–73. [Google Scholar] [CrossRef]
- Berridge, K.C.; Robinson, T.E. What is the role of dopamine in reward: Hedonic impact, reward learning, or incentive salience? Brain Res. Brain Res. Rev. 1998, 28, 309–369. [Google Scholar] [CrossRef]
- Berridge, K.C. Food reward: Brain substrates of wanting and liking. Neurosci. Biobehav. Rev. 1996, 20, 1–25. [Google Scholar] [CrossRef] [PubMed]
- Berridge, K.C.; Robinson, T.E. Parsing reward. Trends Neurosci. 2003, 26, 507–513. [Google Scholar] [CrossRef]
- Robinson, T.E.; Berridge, K.C. Addiction. Annu. Rev. Psychol. 2003, 54, 25–53. [Google Scholar] [CrossRef]
- Stice, E.; Figlewicz, D.P.; Gosnell, B.A.; Levine, A.S.; Pratt, W.E. The contribution of brain reward circuits to the obesity epidemic. Neurosci. Biobehav. Rev. 2013, 37, 2047–2058. [Google Scholar] [CrossRef]
- Gosnell, B.A.; Levine, A.S. Reward systems and food intake: Role of opioids. Int. J. Obes. 2009, 33 (Suppl. 2), S54–S58. [Google Scholar] [CrossRef] [PubMed]
- Grill, H.J.; Norgren, R. Chronically decerebrate rats demonstrate satiation but not bait shyness. Science 1978, 201, 267–269. [Google Scholar] [CrossRef]
- Steiner, J.E.; Glaser, D.; Hawilo, M.E.; Berridge, K.C. Comparative expression of hedonic impact: Affective reactions to taste by human infants and other primates. Neurosci. Biobehav. Rev. 2001, 25, 53–74. [Google Scholar] [CrossRef] [PubMed]
- Balleine, B.W. The meaning of behavior: Discriminating reflex and volition in the brain. Neuron 2019, 104, 47–62. [Google Scholar] [CrossRef]
- Rolls, B.J.; Rowe, E.A.; Rolls, E.T.; Kingston, B.; Megson, A.; Gunary, R. Variety in a meal enhances food intake in man. Physiol. Behav. 1981, 26, 215–221. [Google Scholar] [CrossRef]
- Warlow, S.M.; Berridge, K.C. Incentive motivation: ‘wanting’ roles of central amygdala circuitry. Behav. Brain Res. 2021, 411, 113376. [Google Scholar] [CrossRef]
- Nguyen, D.; Naffziger, E.E.; Berridge, K.C. Positive Affect: Nature and brain bases of liking and wanting. Curr. Opin. Behav. Sci. 2021, 39, 72–78. [Google Scholar] [CrossRef] [PubMed]
- Berridge, K.C.; Dayan, P. Liking. Curr. Biol. 2021, 31, R1555–R1557. [Google Scholar] [CrossRef] [PubMed]
- De Araujo, I.E.; Rolls, E.T.; Velazco, M.I.; Margot, C.; Cayeux, I. Cognitive modulation of olfactory processing. Neuron 2005, 46, 671–679. [Google Scholar] [CrossRef]
- Rolls, E.T.; Deco, G. The Noisy Brain: Stochastic Dynamics as a Principle of Brain Function; Oxford University Press: Oxford, UK, 2010. [Google Scholar]
- Rolls, E.T.; Deco, G. Attention in natural scenes: Neurophysiological and computational bases. Neural Netw. 2006, 19, 1383–1394. [Google Scholar] [CrossRef] [PubMed]
- Deco, G.; Rolls, E.T.; Zihl, J. A neurodynamical model of visual attention. In Neurobiology of Attention; Itti, L., Rees, G., Tsotos, J., Eds.; Elsevier: San Diego, CA, USA, 2005; Chapter 597; pp. 593–599. [Google Scholar]
- Deco, G.; Rolls, E.T. Neurodynamics of biased competition and co-operation for attention: A model with spiking neurons. J. Neurophysiol. 2005, 94, 295–313. [Google Scholar] [CrossRef]
- Deco, G.; Rolls, E.T. Attention, short-term memory, and action selection: A unifying theory. Prog. Neurobiol. 2005, 76, 236–256. [Google Scholar] [CrossRef]
- Deco, G.; Rolls, E.T. A neurodynamical cortical model of visual attention and invariant object recognition. Vision Res. 2004, 44, 621–642. [Google Scholar] [CrossRef]
- Deco, G.; Rolls, E.T. Attention and working memory: A dynamical model of neuronal activity in the prefrontal cortex. Eur. J. Neurosci. 2003, 18, 2374–2390. [Google Scholar] [CrossRef]
- Rolls, E.T. A biased activation theory of the cognitive and attentional modulation of emotion. Front. Hum. Neurosci. 2013, 7, 74. [Google Scholar] [CrossRef]
- Rolls, E.T.; Grabenhorst, F.; Margot, C.; da Silva, M.A.A.P.; Velazco, M.I. Selective attention to affective value alters how the brain processes olfactory stimuli. J. Cogn. Neurosci. 2008, 20, 1815–1826. [Google Scholar] [CrossRef]
- Grabenhorst, F.; Rolls, E.T. Attentional modulation of affective vs sensory processing: Functional connectivity and a top-down biased activation theory of selective attention. J. Neurophysiol. 2010, 104, 1649–1660. [Google Scholar] [CrossRef] [PubMed]
- Ge, T.; Feng, J.; Grabenhorst, F.; Rolls, E.T. Componential Granger causality, and its application to identifying the source and mechanisms of the top-down biased activation that controls attention to affective vs sensory processing. Neuroimage 2012, 59, 1846–1858. [Google Scholar] [CrossRef] [PubMed]
- Rolls, E.T. Taste, olfactory, and food texture reward processing in the brain and the control of appetite. Proc. Nutr. Soc. 2012, 71, 488–501. [Google Scholar] [CrossRef] [PubMed]
- Rolls, E.T. A non-reward attractor theory of depression. Neurosci. Biobehav. Rev. 2016, 68, 47–58. [Google Scholar] [CrossRef]
- Rolls, E.T.; McCabe, C. Enhanced affective brain representations of chocolate in cravers vs non-cravers. Eur. J. Neurosci. 2007, 26, 1067–1076. [Google Scholar] [CrossRef]
- Beaver, J.D.; Lawrence, A.D.; Ditzhuijzen, J.v.; Davis, M.H.; Woods, A.; Calder, A.J. Individual differences in reward drive predict neural responses to images of food. J. Neurosci. 2006, 26, 5160–5166. [Google Scholar] [CrossRef]
- Goldman-Rakic, P.S. The prefrontal landscape: Implications of functional architecture for understanding human mentation and the central executive. Philos. Trans. R. Soc. B 1996, 351, 1445–1453. [Google Scholar]
- Kolb, B.; Whishaw, I.Q. Fundamentals of Human Neuropsychology, 8th ed.; MacMillan: New York, NY, USA, 2021. [Google Scholar]
- Rolls, E.T.; Deco, G.; Huang, C.C.; Feng, J. Prefrontal and somatosensory-motor cortex effective connectivity in humans. Cereb. Cortex 2023, 33, 4939–4963. [Google Scholar] [CrossRef]
- Fuster, J.M. The Prefrontal Cortex, 5th ed.; Academic Press: London, UK, 2015. [Google Scholar]
- Fuster, J.M. The prefrontal cortex makes the brain a preadaptive system. Proc. IEEE 2014, 102, 417–426. [Google Scholar] [CrossRef]
- Miller, E.K.; Lundqvist, M.; Bastos, A.M. Working Memory 2.0. Neuron 2018, 100, 463–475. [Google Scholar] [CrossRef]
- Shallice, T.; Cipolotti, L. The prefrontal cortex and neurological impairments of active thought. Annu. Rev. Psychol. 2018, 69, 157–180. [Google Scholar] [CrossRef] [PubMed]
- Rolls, E.T. Consciousness, decision-making, and neural computation. In Perception-Action Cycle: Models, Algorithms and Systems; Cutsuridis, V., Hussain, A., Taylor, J.G., Eds.; Springer: Berlin/Heidelberg, Germany, 2011; pp. 287–333. [Google Scholar]
- Rolls, E.T. From brain mechanisms of emotion and decision-making to neuroeconomics. In The State of Mind in Economics; Teschl, M., Kirman, A., Eds.; Cambridge University Press: Cambridge, UK, 2025. [Google Scholar]
- Rolls, E.T. Emotion and reasoning in human decision-making. Econ. Open-Access Open-Assess. E-J. 2019, 13. [Google Scholar] [CrossRef]
- Dawkins, R. The Selfish Gene, 2nd ed.; Oxford University Press: Oxford, UK, 1989. [Google Scholar]
- Hamilton, W.D. The genetical evolution of social behaviour. J. Theor. Biol. 1964, 7, 1–52. [Google Scholar] [CrossRef] [PubMed]
- Hamilton, W.D. Narrow Roads of Gene Land; W. H. Freeman: New York, NY, USA, 1996. [Google Scholar]
- Ridley, M. The Red Queen: Sex and the Evolution of Human Nature; Penguin: London, UK, 1993. [Google Scholar]
- Maia, T.V.; McClelland, J.L. A reexamination of the evidence for the somatic marker hypothesis: What participants really know in the Iowa gambling task. Proc. Natl. Acad. Sci. USA 2004, 101, 16075–16080. [Google Scholar] [CrossRef] [PubMed]
- Rolls, E.T. The affective neuroscience of consciousness: Higher order linguistic thoughts, dual routes to emotion and action, and consciousness. In Cambridge Handbook of Consciousness; Zelazo, P., Moscovitch, M., Thompson, E., Eds.; Cambridge University Press: Cambridge, UK, 2007; pp. 831–859. [Google Scholar]
- Grabenhorst, F.; Rolls, E.T.; Parris, B.A.; D’Souza, A. How the brain represents the reward value of fat in the mouth. Cereb. Cortex 2010, 20, 1082–1091. [Google Scholar] [CrossRef]
- Rolls, E.T.; Grabenhorst, F.; Franco, L. Prediction of subjective affective state from brain activations. J. Neurophysiol. 2009, 101, 1294–1308. [Google Scholar] [CrossRef]
- Rolls, E.T.; Grabenhorst, F.; Parris, B.A. Warm pleasant feelings in the brain. Neuroimage 2008, 41, 1504–1513. [Google Scholar] [CrossRef]
- Dawkins, M.S. The Science of Animal Welfare: Understanding What Animals Want; Oxford University Press: Oxford, UK, 2021. [Google Scholar]
- Dawkins, M.S. Who Is Conscious? A Guide to the Minds of Animals; Oxford University Press: Oxford, UK, 2025. [Google Scholar]
- Descartes, R. The Philosophical Writings of Descartes (3 Volumes, 1984-1991); Cambridge University Press: Cambridge, UK, 1644. [Google Scholar]
- Dennett, D.C. Consciousness Explained; Penguin: London, UK, 1991. [Google Scholar]
- Block, N. Two neural correlates of consciousness. Trends Cogn. Sci. 2005, 9, 46–52. [Google Scholar] [CrossRef]
- Carruthers, P. Human and Animal Minds; Oxford University Press: Oxford, UK, 2019. [Google Scholar]
- Rolls, E.T. A neuroscience levels of explanation approach to the mind and the brain. Front. Comput. Neurosci. 2021, 15, 649679. [Google Scholar] [CrossRef]
- Rolls, E.T. Mind Causality: A computational neuroscience approach. Front. Comput. Neurosci. 2021, 15, 706505. [Google Scholar] [CrossRef]
- Rolls, E.T.; Cheng, W.; Du, J.; Wei, D.; Qiu, J.; Dai, D.; Zhou, Q.; Xie, P.; Feng, J. Functional connectivity of the right inferior frontal gyrus and orbitofrontal cortex in depression. Soc. Cogn. Affect. Neurosci. 2020, 15, 75–86. [Google Scholar] [CrossRef] [PubMed]
- Cheng, W.; Rolls, E.T.; Qiu, J.; Yang, D.; Ruan, H.; Wei, D.; Zhao, L.; Meng, J.; Xie, P.; Feng, J. Functional connectivity of the precuneus in unmedicated patients with depression. Biol. Psychiatry Cogn. Neurosci. Neuroimaging 2018, 3, 1040–1049. [Google Scholar] [CrossRef] [PubMed]
- Cheng, W.; Rolls, E.T.; Qiu, J.; Xie, X.; Wei, D.; Huang, C.C.; Yang, A.C.; Tsai, S.J.; Li, Q.; Meng, J.; et al. Increased functional connectivity of the posterior cingulate cortex with the lateral orbitofrontal cortex in depression. Transl. Psychiatry 2018, 8, 90. [Google Scholar] [CrossRef]
- Cheng, W.; Rolls, E.T.; Qiu, J.; Liu, W.; Tang, Y.; Huang, C.C.; Wang, X.; Zhang, J.; Lin, W.; Zheng, L.; et al. Medial reward and lateral non-reward orbitofrontal cortex circuits change in opposite directions in depression. Brain 2016, 139, 3296–3309. [Google Scholar] [CrossRef]
- Rolls, E.T.; Deco, G.; Huang, C.-C.; Feng, J. Multiple cortical visual streams in humans. Cereb. Cortex 2023, 33, 3319–3349. [Google Scholar] [CrossRef] [PubMed]
- Rolls, E.T.; Xiang, J.-Z. Reward-spatial view representations and learning in the hippocampus. J. Neurosci. 2005, 25, 6167–6174. [Google Scholar] [CrossRef]
- Strongman, K.T. The Psychology of Emotion, 5th ed.; Wiley: London, UK, 2003. [Google Scholar]
- Keltner, D.; Oatley, K.; Jenkins, J.M. Understanding Emotions; Wiley: Hoboken, NJ, USA, 2018. [Google Scholar]
- James, W. What is an emotion? Mind 1884, 9, 188–205. [Google Scholar] [CrossRef]
- Lange, C. The emotions. In The Emotions; Dunlap, E., Ed.; Williams and Wilkins: Baltimore, MD, USA, 1885. [Google Scholar]
- Quadt, L.; Critchley, H.D.; Garfinkel, S.N. The neurobiology of interoception in health and disease. Ann. N. Y. Acad. Sci. 2018, 1428, 112–128. [Google Scholar] [CrossRef]
- Levenson, R.W.; Ekman, P.; Friesen, W.V. Voluntary facial action generates emotion-specific autonomic nervous system activity. Psychophysiology 1990, 27, 363–384. [Google Scholar] [CrossRef]
- Ekman, P.; Levenson, R.W.; Friesen, W.V. Autonomic nervous system activity distinguishes between the emotions. Science 1983, 221, 1208–1210. [Google Scholar] [CrossRef]
- Cannon, W.B. Again the James-Lange theory of emotion: A critical examination and an alternative theory. Psychol. Rev. 1931, 38, 281–295. [Google Scholar] [CrossRef]
- Cannon, W.B. The James-Lange theory of emotion: A critical examination and an alternative theory. Am. J. Psychol. 1927, 39, 106–124. [Google Scholar] [CrossRef]
- Hohmann, G.W. Some effects of spinal cord lesions on experienced emotional feelings. Psychophysiology 1966, 3, 143–156. [Google Scholar] [CrossRef] [PubMed]
- Bermond, B.; Fasotti, L.; Niewenhuyse, B.; Shuerman, J. Spinal cord lesions, peripheral feedback and intensities of emotional feelings. Cogn. Emot. 1991, 5, 201–220. [Google Scholar] [CrossRef]
- Schachter, S.; Singer, J. Cognitive, social and physiological determinants of emotional state. Psychol. Rev. 1962, 69, 387–399. [Google Scholar] [CrossRef] [PubMed]
- Reisenzein, R. The Schachter theory of emotion: Two decades later. Psychol. Bull. 1983, 94, 239–264. [Google Scholar] [CrossRef]
- Kraut, R.E.; Johnson, R.E. Social and emotional messages of smiling: An ethological approach. J. Pers. Soc. Psychol. 1979, 37, 1539–1553. [Google Scholar] [CrossRef]
- Heims, H.C.; Critchley, H.D.; Dolan, R.; Mathias, C.J.; Cipolotti, L. Social and motivational functioning is not critically dependent on feedback of autonomic responses: Neuropsychological evidence from patients with pure autonomic failure. Neuropsychologia 2004, 42, 1979–1988. [Google Scholar] [CrossRef] [PubMed]
- Maia, T.V.; McClelland, J.L. The somatic marker hypothesis: Still many questions but no answers: Response to Bechara et al. Trends Cogn. Sci. 2005, 9, 162–164. [Google Scholar] [CrossRef]
- Adolphs, R.; Damasio, H.; Tranel, D.; Cooper, G.; Damasio, A.R. A role for somatosensory cortices in the visual recognition of emotion as revealed by three-dimensional lesion mapping. J. Neurosci. 2000, 20, 2683–2690. [Google Scholar] [CrossRef]
- Rolls, E.T. The orbitofrontal cortex and reward. Cereb. Cortex 2000, 10, 284–294. [Google Scholar] [CrossRef]
- Rolls, E.T. A theory of emotion, and its application to understanding the neural basis of emotion. Cogn. Emot. 1990, 4, 161–190. [Google Scholar] [CrossRef]
- Frijda, N.H. The Emotions; Cambridge University Press: Cambridge, UK, 1986. [Google Scholar]
- Moors, A.; Ellsworth, P.C.; Scherer, K.R.; Frijda, N.H. Appraisal theories of emotion: State of the art and future development. Emot. Rev. 2013, 5, 119–124. [Google Scholar] [CrossRef]
- Oatley, K.; Jenkins, J.M. Understanding Emotions; Blackwell: Oxford, UK, 1996. [Google Scholar]
- Scherer, K.S. The dynamic architecture of emotion: Evidence for the component process model. Cogn. Emot. 2009, 23, 1307–1351. [Google Scholar] [CrossRef]
- Panksepp, J. Affective Neuroscience: The Foundations of Human and Animal Emotions; Oxford University Press: New York, NY, USA, 1998. [Google Scholar]
- Panksepp, J. The basic emotional circuits of mammalian brains: Do animals have affective lives? Neurosci. Biobehav. Rev. 2011, 35, 1791–1804. [Google Scholar] [CrossRef] [PubMed]
- Ekman, P. Emotions Revealed: Understanding Thoughts and Feelings; Weidenfeld and Nicolson: London, UK, 2003. [Google Scholar]
- Seligman, M.E. On the generality of the laws of learning. Psychol. Rev. 1970, 77, 406–418. [Google Scholar] [CrossRef]
- Darwin, C. The Expression of the Emotions in Man and Animals, 3rd ed.; University of Chicago Press: Chicago, IL, USA, 1872. [Google Scholar]
- Ekman, P. An argument for basic emotions. Cogn. Emot. 1992, 6, 169–200. [Google Scholar] [CrossRef]
- Scherer, K.R. Psychological models of emotion. In The Neuropsychology of Emotion; Borod, J.C., Ed.; Oxford University Press: Oxford, UK, 2000; pp. 137–162. [Google Scholar]
- Harmon-Jones, E.; Harmon-Jones, C.; Summerell, E. On the importance of both dimensional and discrete models of emotion. Behav. Sci. 2017, 7, 66. [Google Scholar] [CrossRef] [PubMed]
- Barrett, L.F. Discrete emotions or dimensions? The role of valence focus and arousal focus. Cogn. Emot. 1998, 12, 579–599. [Google Scholar] [CrossRef]
- Miller, G.A. The magic number seven, plus or minus two: Some limits on our capacity for the processing of information. Psychol. Rev. 1956, 63, 81–93. [Google Scholar] [CrossRef]
- Ben-Ze’ev, A. The Subtlety of Emotions; MIT press: Cambridge, MA, USA, 2001. [Google Scholar]
- LeDoux, J.E. Emotion and the amygdala. In The Amygdala; Aggleton, J.P., Ed.; Wiley-Liss: New York, NY, USA, 1992; pp. 339–351. [Google Scholar]
- LeDoux, J.E.; Daw, N.D. Surviving threats: Neural circuit and computational implications of a new taxonomy of defensive behaviour. Nat. Rev. Neurosci. 2018, 19, 269–282. [Google Scholar] [CrossRef] [PubMed]
- Barrett, L.F. The theory of constructed emotion: An active inference account of interoception and categorization. Soc. Cogn. Affect. Neurosci. 2017, 12, 1833. [Google Scholar] [CrossRef] [PubMed]
- Kleckner, I.R.; Zhang, J.; Touroutoglou, A.; Chanes, L.; Xia, C.; Simmons, W.K.; Quigley, K.S.; Dickerson, B.C.; Barrett, L.F. Evidence for a large-scale brain system supporting allostasis and interoception in humans. Nat. Hum. Behav. 2017, 1, 0069. [Google Scholar] [CrossRef]
- Bindra, D. A unified interpretation of emotion and motivation. Ann. N. Y. Acad. Sci. 1969, 159, 1071–1083. [Google Scholar] [CrossRef]
- Rolls, E.T. The neural basis of brain-stimulation reward. Prog. Neurobiol. 1974, 3, 71–160. [Google Scholar] [CrossRef]
- Mora, F.; Avrith, D.B.; Phillips, A.G.; Rolls, E.T. Effects of satiety on self-stimulation of the orbitofrontal cortex in the monkey. Neurosci. Lett. 1979, 13, 141–145. [Google Scholar] [CrossRef]
- Rolls, E.T.; Burton, M.J.; Mora, F. Neurophysiological analysis of brain-stimulation reward in the monkey. Brain Res. 1980, 194, 339–357. [Google Scholar] [CrossRef]
- Burton, M.J.; Rolls, E.T.; Mora, F. Effects of hunger on the responses of neurones in the lateral hypothalamus to the sight and taste of food. Exp. Neurol. 1976, 51, 668–677. [Google Scholar] [CrossRef] [PubMed]
- Rolls, E.T.; Sienkiewicz, Z.J.; Yaxley, S. Hunger modulates the responses to gustatory stimuli of single neurons in the caudolateral orbitofrontal cortex of the macaque monkey. Eur. J. Neurosci. 1989, 1, 53–60. [Google Scholar] [CrossRef]
- Rolls, B.J.; Rolls, E.T.; Rowe, E.A.; Sweeney, K. Sensory specific satiety in man. Physiol. Behav. 1981, 27, 137–142. [Google Scholar] [CrossRef]
- Rolls, B.J.; Rowe, E.A.; Rolls, E.T. How sensory properties of foods affect human feeding behaviour. Physiol. Behav. 1982, 29, 409–417. [Google Scholar] [CrossRef]
- Nicolaidis, S.; Rowland, N. Metering of intravenous versus oral nutrients and regulation of energy balance. Am. J. Physiol. 1976, 231, 661–668. [Google Scholar] [CrossRef] [PubMed]
- Nicolaidis, S.; Rowland, N. Systemic vs. oral and gasto-intestinal metering of fluid intake. In Control Mechanisms of Drinking; Peters, G., Fitzsimons, J.T., Eds.; Springer: Berlin/Heidelberg, Germany, 1975. [Google Scholar]
- Monteiro, M.P.; Batterham, R.L. The Importance of the Gastrointestinal Tract in Controlling Food Intake and Regulating Energy Balance. Gastroenterology 2017, 152, 1707–1717.e2. [Google Scholar] [CrossRef] [PubMed]
- Phillips, P.; Rolls, B.J.; Ledingham, J.; Morton, J. Body fluid changes, thirst and drinking in man during free access to water. Physiol. Behav. 1984, 33, 357–363. [Google Scholar] [CrossRef] [PubMed]
- Rolls, E.T.; Rolls, B.J.; Rowe, E.A. Sensory-specific and motivation-specific satiety for the sight and taste of food and water in man. Physiol. Behav. 1983, 30, 185–192. [Google Scholar] [CrossRef]
- Morales, I.; Berridge, K.C. ‘Liking’ and ‘wanting’ in eating and food reward: Brain mechanisms and clinical implications. Physiol. Behav. 2020, 227, 113152. [Google Scholar] [CrossRef]
- Berridge, K.C. Evolving Concepts of Emotion and Motivation. Front. Psychol. 2018, 9, 1647. [Google Scholar] [CrossRef] [PubMed]
- Robinson, T.E.; Berridge, K.C. The Incentive-Sensitization Theory of Addiction 30 Years On. Annu. Rev. Psychol. 2025, 76, 29–58. [Google Scholar] [CrossRef]
- Reeve, J.M. Understanding Motivation and Emotion, 6th ed.; John Wiley & Sons: Hoboken, NJ, USA, 2024. [Google Scholar]
- Rolls, E.T.; Wirth, S.; Deco, G.; Huang, C.-C.; Feng, J. The human posterior cingulate, retrosplenial and medial parietal cortex effective connectome, and implications for memory and navigation. Hum. Brain Mapp. 2023, 44, 629–655. [Google Scholar] [CrossRef]
- Rolls, E.T.; Deco, G.; Huang, C.C.; Feng, J. The human posterior parietal cortex: Effective connectome, and its relation to function. Cereb. Cortex 2023, 33, 3142–3170. [Google Scholar] [CrossRef]
- Rolls, E.T. Hippocampal spatial view cells, place cells, and concept cells: View representations. Hippocampus 2023, 33, 667–687. [Google Scholar] [CrossRef] [PubMed]
- Rolls, E.T. Hippocampal spatial view cells for memory and navigation, and their underlying connectivity in humans. Hippocampus 2023, 33, 533–572. [Google Scholar] [CrossRef] [PubMed]
- Huang, C.-C.; Rolls, E.T.; Hsu, C.-C.H.; Feng, J.; Lin, C.-P. Extensive cortical connectivity of the human hippocampal memory system: Beyond the “what” and “where” dual-stream model. Cereb. Cortex 2021, 31, 4652–4669. [Google Scholar] [CrossRef] [PubMed]
- Ma, Q.; Rolls, E.T.; Huang, C.-C.; Cheng, W.; Feng, J. Extensive cortical functional connectivity of the human hippocampal memory system. Cortex 2022, 147, 83–101. [Google Scholar] [CrossRef]
- Rolls, E.T.; Rauschecker, J.P.; Deco, G.; Huang, C.C.; Feng, J. Auditory cortical connectivity in humans. Cereb. Cortex 2023, 33, 6207–6227. [Google Scholar] [CrossRef]
- Glasser, M.F.; Smith, S.M.; Marcus, D.S.; Andersson, J.L.; Auerbach, E.J.; Behrens, T.E.; Coalson, T.S.; Harms, M.P.; Jenkinson, M.; Moeller, S.; et al. The Human Connectome Project’s neuroimaging approach. Nat. Neurosci. 2016, 19, 1175–1187. [Google Scholar] [CrossRef]

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Taste | |
| Salt taste | reward in salt deficiency |
| Sweet | reward in energy deficiency |
| Bitter | punisher, an indicator of possible poison |
| Sour | punisher |
| Umami | reward, an indicator of protein; produced by monosodium glutamate and inosine monophosphate |
| Tannic acid | punisher; it prevents absorption of protein; found in old leaves; probably somatosensory not gustatory (Critchley and Rolls 1996 [17]) |
| Odor | |
| Putrefying odor | Punisher, a hazard to health |
| Pheromones | reward (depending on the hormonal state) |
| Somatosensory | |
| Pain | punisher |
| Touch | reward |
| Grooming | reward; to give grooming may also be a primary reinforcer |
| Washing | reward |
| Temperature | reward if it tends to help maintain normal body temperature; otherwise, a punisher |
| Visual | |
| Snakes, etc. | punisher for, e.g., primates |
| Youthfulness | reward, associated with mate choice |
| Beauty, e.g., symmetry | reward |
| Secondary sexual characteristics | rewards |
| Face expression | reward (e.g., smile) or punisher (e.g., threat) |
| Blue sky, cover, open space | reward, indicator of safety |
| Flowers | reward (indicator of fruit later in the season?) |
| Auditory | |
| Warning call | punisher |
| Aggressive vocalization | punisher |
| Soothing vocalization | reward (part of the evolutionary history of music, which at least in its origins taps into the channels used for the communication of emotions) |
| Reproduction | |
| Courtship | reward |
| Sexual behavior | reward (different reinforcers, including a low waist-to-hip ratio, and attractiveness influenced by symmetry and being found attractive by members of the other sex). |
| Mate guarding | reward for a male to protect his parental investment. |
| Jealousy results if his mate is courted by another male, | |
| because this may ruin his parental investment | |
| Nest building | reward (when expecting young) |
| Parental attachment (love) | reward (good for the parent’s genes |
| both when the attachment is to the other parent or an infant) | |
| Infant attachment to parents (love) Crying of infant | reward (good for the infant’s genes) punisher to parents; produced to promote successful development |
| Power, status, wealth, resources | Attractive to females, who may benefit from resources for their offspring. |
| Attractive to males as they make males attractive to females. | |
| Body size | Large in males may be attractive to females as a signal |
| for the provision of protection | |
| and of the ability of her male offspring to compete for a mate. | |
| Small in females may be attractive to males as a neotenous sign | |
| of youth, and therefore fertility | |
| Other | |
| Novel stimuli | rewards (encourage animals to investigate the full possibilities |
| of the multidimensional space in which their genes are operating) | |
| Sleep | reward; minimizes nutritional requirements and protects from danger |
| Altruism to genetic kin | reward (kin altruism) |
| Altruism to other individuals | reward while the altruism is reciprocated |
| in a ‘tit-for-tat’ reciprocation (reciprocal altruism). | |
| Forgiveness, honesty, and altruistic punishment | |
| are some associated heuristics. | |
| May provide underpinning for some aspects of what is felt to be moral. | |
| punisher when the altruism is not reciprocated | |
| Group acceptance, reputation Control over actions | reward (social greeting might indicate this). These goals can account for why some cultural goals are pursued reward |
| Play | reward |
| Danger, stimulation, excitement | reward if not too extreme (adaptive because of practice?) |
| Exercise | reward (keeps the body fit for action) |
| Mind reading | reward; practice in reading others’ minds, which might be adaptive |
| Solving an intellectual problem | reward (practice in which might be adaptive) |
| Storing, collecting | reward (e.g., food) |
| Habitat preference, home, territory | reward |
| Some responses | reward (e.g., pecking in chickens, pigeons; |
| adaptive because it is a simple way in which eating grain | |
| can be programmed for a relatively fixed type of environmental stimulus) |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2025 by the author. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Rolls, E.T. Emotion, Motivation, Reasoning, and How Their Brain Systems Are Related. Brain Sci. 2025, 15, 507. https://doi.org/10.3390/brainsci15050507
Rolls ET. Emotion, Motivation, Reasoning, and How Their Brain Systems Are Related. Brain Sciences. 2025; 15(5):507. https://doi.org/10.3390/brainsci15050507
Chicago/Turabian StyleRolls, Edmund T. 2025. "Emotion, Motivation, Reasoning, and How Their Brain Systems Are Related" Brain Sciences 15, no. 5: 507. https://doi.org/10.3390/brainsci15050507
APA StyleRolls, E. T. (2025). Emotion, Motivation, Reasoning, and How Their Brain Systems Are Related. Brain Sciences, 15(5), 507. https://doi.org/10.3390/brainsci15050507