Abstract
Acetylcholine is a critical modulatory neurotransmitter for cognitive function. Cholinergic drugs improve cognitive performance and enhance neuronal activity in the sensory and association cortices. An alternative means of improving cognitive function is through the use of deep brain stimulation. Prior animal studies have demonstrated that stimulation of the nucleus basalis of Meynert through DBS improves cognitive performance on a visual working memory task to the same degree as cholinesterase inhibitors. Additionally, unlike current pharmacological treatments for neurocognitive disorders, DBS does not lose efficacy over time and adverse effects are rare. These findings suggest that DBS may be a promising alternative for treating cognitive impairments in neurodegenerative disorders such as Alzheimer’s disease. Thus, further research and human trials should be considered to assess the potential of DBS as a therapeutic treatment for these disorders.
1. Introduction
Working memory is a core cognitive function that allows us to maintain information in our mind over a period of seconds and underpins more complex abilities such as planning, language, and fluid intelligence [1]. Due to this central role in cognition, improving working memory has been a goal of interventions aiming to restore cognitive function impairments due to a multitude of pathological conditions such as traumatic brain injury, stroke, schizophrenia, and Alzheimer’s disease [2,3,4]. Ascending cholinergic input to the neocortex is a critical neuromodulator in working memory [5]. Cholinergic drugs are frontline medications for many conditions that compromise cognitive function [6]. Visual working memory performance is enhanced through such cholinergic modulation [7]. In recent years, deep brain stimulation (DBS) targeting the cholinergic basal forebrain has emerged as an alternative to pharmacologic approaches to modulate acetylcholine release and improve cognitive function [8]. To understand the promises and challenges involving this approach, here we will review the actions of neuromodulation systems and acetylcholine in particular, and we will discuss the role of cholinergic innervation with respect to visual working memory, the mechanisms of action of deep brain stimulation targeting the cholinergic system that support its viability as an intervention, and the experimental evidence of its effectiveness. As this field is still relatively new, most of the latter evidence comes from animal studies, particularly in non-human primates; this will be the emphasis of our review.
Monoamines and acetylcholine are important neuromodulatory systems that govern the cognitive functions mediated by the prefrontal cortex (PFC). The interplay between these systems is crucial for the execution of cognitive functions including arousal systems [9]. Fluctuations in dopamine, serotonin, and norepinephrine levels affect the performance of PFC cognitive processes such as working memory, attention, and decision-making.
In the dorsolateral PFC (dlPFC), neuromodulation of the monoamines and acetylcholine systems are especially critical [9]. Dopamine primarily acts on D1 receptors to increase cAMP production in the primate dlPFC. However, this action is dose dependent and there is a range of optimal stimulation, as too little or too much has been shown to impair working memory function [10]. The administration of D1R agonists has been shown to reduce noise during working memory representations [11]. The increased cAMP production opens K+ channels maintaining delay-related firing of relevant inputs while suppressing the response to distractor inputs [11].
Misfiring and loss of regulation of these neuromodulatory systems in the dlPFC leads to cognitive impairments and is generally seen with aging [9]. While the exact functions of these neuromodulators may not be completely understood, we do know that acetylcholine and dopamine are essential for proper cognitive processing, specifically working memory [12]. Studies focused on monoaminergic and cholinergic innervation and activation will be the key to understanding how we can retain and improve working memory function.
2. Cholinergic System
The primate neocortex receives its acetylcholine innervation from the nucleus basalis of Meynert (NB) [13,14]. Nucleus basalis activity levels during wakefulness reflect alertness levels [15] and are systematically related to executive function [16,17,18,19]. Ascending cholinergic projections spread across the neocortex, allowing the system to control the excitability of most cortical areas; however, the modulation can still be specific for particular areas and stimuli [20].
Endogenously released acetylcholine acts on cholinergic receptors, both nicotinic and muscarinic. Nicotinic receptors are ion channels that allow for the passage of calcium and sodium ions into the postsynaptic neuron when acetylcholine molecules are bound to them. This regulation of calcium signaling increases synaptic transmission and plasticity and enhances the release of dopamine. Muscarinic receptors are G-protein coupled receptors that activate the phosphoinositide and cAMP cascading pathways. These activated pathways result in changes to the postsynaptic neuron’s excitability. Specifically, the activation of M1 receptors within the PFC has been shown to increase the signal-to-noise ratio and filter out irrelevant information [9]. Activation of nicotinic α7 receptors similarly facilitates spatial working memory. When acetylcholine binds to the α7 receptors, it allows for the activation of the NMDA receptors, which is crucial for maintaining persistent activity during spatial working memory tasks [12]. The human PFC releases acetylcholine in response to a target in an inverted U-shaped relationship with task performance [21]. In monkey dlPFC neurons, cholinergic agents have reduced the effect of distractors [22], which is consistent with cholinesterase inhibitors enhancing target detection amongst distractors [23]. These results suggest acetylcholine may be essential for perceptual representations in natural environments, among competing stimuli.
Cholinergic action is essential for cognitive functions and behavioral outputs; conditions that cause cholinergic deficits result in cognitive function decline [24,25]. Basal forebrain lesions in non-human primates cause deficits in cognitive domains including spatial attention, working memory, and task acquisition [26,27,28]. Deafferentation of cholinergic projections to the prefrontal cortex impairs monkeys’ selectively in spatial working memory tasks, while leaving performance in tests of decision-making and episodic memory unaffected [29]. Decrease in the action of acetylcholine, for example by administration of muscarinic receptor antagonists, impairs cognitive abilities in both humans and animals [30,31,32,33,34]. Similarly, atrophy of the NB has been observed in cognitive disorders and has been linked to the decline of cholinergic neurotransmission and cognitive impairment [35]. Dysfunction of acetylcholine-releasing neurons in the basal forebrain contributes to the cognitive declines observed in Alzheimer’s disease [17,18,19,36]. Reduced acetylcholine release, alterations in choline transport, and changes in nicotinic and muscarinic receptor expression have been observed [18,19,36].
Conversely, systemic administration of acetylcholinesterase inhibitors is widely used to treat executive function deficits in human patients [37,38,39,40,41]. Such drugs have also been shown to improve performance in working memory tasks in monkeys [18,42]. Acetylcholinesterase inhibitors that increase cholinergic neurotransmission in the brain by preventing the breakdown of acetylcholine can partially reverse cognitive deficits [43,44]. However, the value of cholinergic medications is limited; high doses result in cognitive improvements that peak in ~3 months [45], and typically attenuate after a year. Even moderate doses are discontinued in some patients due to peripheral side effects that arise due to off-target effects of the drugs [44].
3. Neural Activity Underlying Visual Working Memory
The neural mechanisms underlying visual working memory have been extensively studied in recent years and have been a matter of some debate [46,47]. Improved visual working memory performance involves activation in the dorsolateral prefrontal cortex and the inferior parietal lobule [48,49]. It is notable that similar effects are observed with the administration of a cholinesterase inhibitor during visual working memory tasks in Alzheimer’s patients, including heightened activation in the prefrontal and parietal cortex during the memory maintenance period [50].
Individual neurons exhibit persistent activity with selectivity for spatial locations [51,52]. Computational models typically simulate persistent activity by networks of neurons with recurrent connections between similarly tuned units, as illustrated in Figure 1A [53,54].
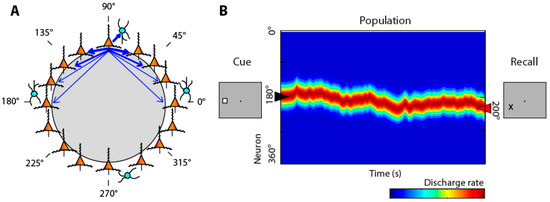
Figure 1.
(A) Schematic diagram of the bump attractor model. Neurons representing different stimulus location are arranged in a ring so as to indicate each neuron’s preferred location. Synaptic connections between neurons with similar preference are stronger. (B) Schematic of the pattern of activity in the network. Y axis represents neurons in the ring with different spatial preference. A stimulus appearing at the left creates a peak of activity in the network centered on the 180° location. This peak may drift in time during the delay period of the task. The location of the peak at the end of the delay period determines the location that the subject recalls. From Ref. [55].
Activation in the network behaves as a continuous attractor [56]. An initial stimulus appearance generates a bump (peak) of activity in the network, which is maintained in the delay period, but may drift randomly. The peak of the bump at the end of the delay period determines the location recalled by the subject (Figure 1B), hence the term bump attractor [57]. Drifts in neuronal activity account for deviations of behavior: persistent activity recorded from trials in which monkeys make eye movements deviating clockwise vs. counterclockwise relative to the true location of the stimulus yields slightly different tuning curves (Figure 1, bottom), implying that the peak of activity at the end of the delay period determines the recalled location [57].
PFC neurons also generate persistent discharges in memory tasks that require the maintenance of stimulus features such as shape, color, or direction of motion, and the identity of objects, faces, and abstract pictures [58].
4. Action of Acetylcholine on the Visual Cortex
Cholinergic release, during states of elevated alertness or focused attention, enhances the processing of visual information [59]. Acetylcholine controls the flow of sensory information in the cortex by increasing the gain of thalamic input and thus enhances contrast sensitivity, a mechanism mediated by nicotinic receptors [60]. Administration of acetylcholine or cholinergic agonists in the sensory cortices similarly enhances responses to stimuli. This is the case in monkey V1, where responses to stimulus presentations improve under acetylcholine iontophoresis [61]. The effect is blocked by muscarinic antagonists. Similarly, cholinergic agonists in the middle temporal area (MT), an area of the visual cortex implicated in motion perception, affect responses to visual stimuli, which are also blocked by muscarinic antagonists [62]. Importantly, neurons that are not driven by the sensory inputs are suppressed rather than facilitated by acetylcholine administration, thus increasing signal-to-noise ratio as well as reducing top-down influences on sensory responses, such as that related to expectation [21,63]. More direct evidence about the short-term action of acetylcholine has been obtained by optogenetic, phasic stimulation of cholinergic neurons in rodents, which has been shown to increase visual cortical firing rate during the period of visual stimulus presentations [64]. Substantial species differences between primates and rodents have been reported, however [65].
The effects of acetylcholine are not uniform between cortical areas. Whereas in area V1 acetylcholine enhances responses to attended over unattended stimuli [61], no such effect was observed in area MT [62]. The diversity of cholinergic effects can be attributed, at least in part, to different patterns of expression of muscarinic and nicotinic receptors in pyramidal neurons and interneurons in different cortical areas [12,66].
5. Effects of Acetylcholine on the Prefrontal Cortex
Cholinergic agonists generally increase the activity of prefrontal neurons [22,67,68]. Conversely, systemic administration of the muscarinic antagonist scopolamine reduces prefrontal activity [69], as does micro-iontophoresis of muscarinic and nicotinic-α7 inhibitors [67,70,71]. The increase in activity by agonists is selective for the preferred location of the neuron, so that tuning is enhanced (schematically shown in Figure 2A).
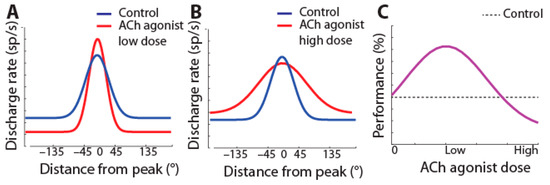
Figure 2.
(A) Schematic effect of iontophoresis of low doses of cholinergic agonists on tuning of prefrontal neurons: firing rate increases at the neuron’s preferred (peak) location, resulting in sharpened tuning. (B) At high doses, agonists broaden tuning. (C) Hypothesized effect of dosage of cholinergic agonists on performance: improvement of performance is expected at doses that sharpen tuning.
This sharpening of prefrontal neurons’ tuning functions takes place with low doses of cholinergic agonists. Broader tuning (as in Figure 2B) has been observed in pharmacological studies by cholinergic overstimulation with high doses of carbachol or M1R allosteric inhibitors administered with iontophoresis in working memory tasks [71,72,73].
The decrease in selectivity by cholinergic overstimulation [71,72,73] has been assumed to correspond to the descending section of an inverted-U function, representing a regime over which cholinergic agonists impair performance (as in Figure 2C) [74]. The effectiveness of drugs targeting other neurotransmitter systems, e.g., dopamine, is also often interpreted in terms of increased stimulus selectivity [75].
6. Deep Brain Stimulation
An alternative method of improving cholinergic function is to selectively activate the NB, the exclusive source of cholinergic innervation of the neocortex in primates, including humans [76]. Such a method has several advantages, including avoidance of side effects caused by peripheral cholinergic targets and control of the specific time intervals during which NB stimulation is applied, e.g., during waking hours, when acetylcholine is normally released [77]. Additionally, stimulation is likely to enable the activation of non-cholinergic projection neurons [78,79,80,81], and the co-transmission of GABA from cholinergic terminals [82,83], better mirroring the physiological conditions of natural acetylcholine release.
The biophysical mechanisms of DBS are the subject of some debate, but continuous high-frequency stimulation generally used to treat movement disorders appears to overall suppress the activity of the implanted area through overstimulation, by a rate of efferent targets [84,85,86]. This results in a reduced impact of the implanted region on its target areas. In most DBS applications, rectangular monophasic or biphasic voltage pulses are used to evoke the desired physiological effects [87]. Biphasic stimulation consisting of a cathodal stimulation phase followed by an anodic reversal phase is typically used in clinical applications [88,89]. Deep brain stimulation is not cell type specific, as electrical stimulation induces extracellular depolarization of all excitable structures in the target spatial region including fibers of passage irrespective of biochemical composition and morphology, but it is spatially selective around the electrode lead. The activating function, or ability to elicit an all-or-none action potential response, drops off sharply at approximately 2 mm from the electrode lead for a typical current near 3 mA. The square root of the current scales with the radius of the activating function [90,91,92]. The spatial specificity of these fields is quite difficult to match with non-invasive stimulation [93]. Increasingly sophisticated stimulation has been made possible in movement disorders, for example addressing different subtypes of Parkinson’s disease with precise targeting of the subthalamic nucleus or internal segment of the globus pallidus [94].
7. Cholinergic Deep Brain Stimulation in Visual Working Memory
A series of recent studies have investigated the use of cholinergic DBS for visual working memory and attention. In one study, monkeys were trained on a delayed-match-to-sample task in which they were required to remember an initial stimulus for a short delay period and then indicate, out of two options, which object was the match of the initial stimulus. Unexpectedly, applying continuous stimulation to the nucleus basalis of Meynert at high frequencies (e.g., 80 Hz or higher) similar to the high frequencies used in movement disorders did not improve but instead impaired behavioral performance in the cognitive task used [95]. After testing different stimulation parameters, an alternative stimulation regime involving intermittent stimulation was found to improve performance. This regime involved delivering 1200 pulses per minute for 15–20 s (i.e., with a frequency of stimulation of 80–60 Hz, respectively) alternating with intervals of no pulses lasting 40–45 s. Moreover, the intermittent stimulation parameters were tested in conjunction with Donepezil, a cholinesterase inhibitor. The combination of DBS with Donepezil did not increase performance any more than DBS alone or Donepezil alone, suggesting a ceiling effect. Further results from this study suggested intermittent stimulation could be blocked by both nicotinic and muscarinic antagonists [95]. The benefits of cholinergic deep brain stimulation were not limited to a single task. In a different study, monkeys receiving cholinergic stimulation performed better in a continuous visual attention task by increasing the hit rate and decreasing the false alarm rate [96]. More recently, cholinergic stimulation has shown promise in improving cognitive function in aged monkeys [97].
Another recent study also examined the effects of cholinergic deep brain stimulation on neuronal activity [98]. The pattern of intermittent stimulation allows for recordings to be interleaved with periods of stimulation (Figure 3).
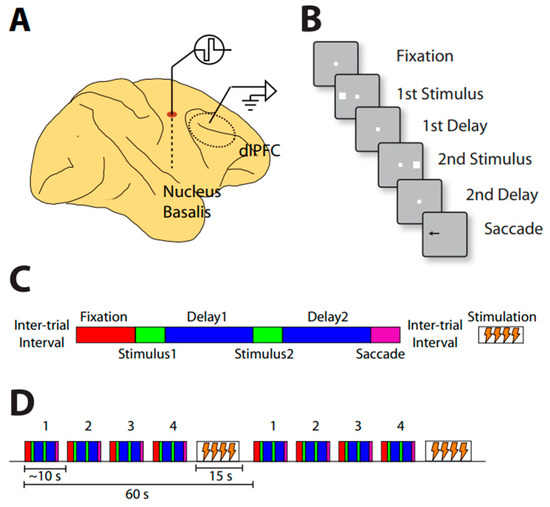
Figure 3.
(A) Illustration of simultaneous NB stimulation and neurophysiological recordings from the prefrontal cortex. (B) Schematic diagram of the behavioral task. Successive frames represent task events. (C) Diagram of the relative timing of task events and NB stimulation. Behavioral trials comprise the fixation period, visual stimulus presentation, delay period, and response periods, now represented by different color rectangles. Stimulation is delivered in between trials after the saccade. (D) Trial structure during neural recording. Behavioral trials lasting approximately 45 s are interleaved with 15 s periods of stimulation, when no behavior is performed. Comparisons can be made between blocks of trials involving stimulation during the 15 s pause period, and blocks of trials not delivering stimulation (sham).
Neural activity in the prefrontal cortex was recorded during a task in which two stimuli were presented in a sequence. Depending on the color of the fixation point, the monkeys had to make an eye movement to the remembered location of the first or second stimulus. Stimulation of 80 Hz frequency was applied between trials for 15 s, and this was repeated after the proceeding 45 s trial was completed. NB stimulation increased overall activity and behavioral performance during the working memory task (Figure 4).
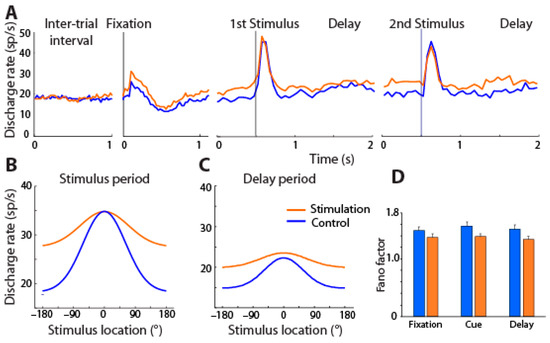
Figure 4.
(A) Mean firing rate across PFC neurons with (orange) and without NB stimulation (blue) is shown for different task intervals, involving presentation of the best visual stimulus of each neuron. (B,C) Average tuning function from all PFC neurons for cue and delay period with and without stimulation; best stimulus has been shifted to 0° location in each case. (D) Mean Fano factor of spike counts during the fixation, cue, and delay period of the task under control and NB stimulation. From Ref. [98].
Unexpectedly, and unlike the results of cholinergic agonists reviewed above, the tuning of prefrontal neurons generally broadened, i.e., neurons responded more strongly to stimuli that were less optimal. Sharper tuning is typically associated with increased performance for conditions in which decoding precise stimulus information is essential, as in discrimination tasks, after perceptual learning [99,100,101,102]. Theoretical studies have shown, however, that broader tuning curves can produce either worse or better performance depending on the task and noise conditions in the network [103,104,105,106]. For conditions in which stimuli are highly discriminable, and performance depends on the ability to filter distractors or other intervening noise and implement the correct rule, as was the case in the task we used, broader tuning may lead to more efficient coding [107]. Unlike the effects of perceptual learning mentioned above, learning to perform a working memory task also induces broadening of neural selectivity [108,109].
Cholinergic NB stimulation had a number of other effects on neural activity. It reduced the trial-to-trial variability of the neuronal firing rate, consistent with the effects of cholinergic drugs [110,111]. NB stimulation also produced changes in the rhythmicity of local field potentials, leading most significantly to a decrease in α power in the prefrontal cortex [112].
8. Potential for Human Intervention
In a few recent studies (Table 1), human patients were implanted with electrodes that allowed stimulation of the nucleus basalis of Meynert, among other targets [76,113,114,115,116,117,118,119]. Implantation and stimulation pulses in the range of 20 Hz (and higher) were safe and well tolerated, and adverse effects were comparable to those of movement disorders. Improvement in glucose utilization, and some cognitive benefits as captured by the neuropsychiatric inventory, were reported. However, compelling cognitive improvements were not observed for the stimulation group. All of these clinical studies relied on the application of continuous stimulation at 20 Hz or higher, leaving open the possibility that a protocol of intermittent stimulation might be more effective. Animal studies have shown the benefits and cognitive improvements of intermittent stimulation over continuous stimulation, which has not yet been explored in human trials [95]. Using intermittent stimulation allows for greater flexibility of stimulation parameters and reduces the risk of desensitization of the target areas. Furthermore, intermittent stimulation provides a more controlled approach that could be leveraged for more sustainable cognitive improvement.

Table 1.
Human clinical studies targeting the nucleus basalis of Meynert (NB) [76,113,114,115,116,117,118,119,120].
There are still challenges and limitations that need to be addressed before DBS can be considered as a therapy for memory deficits, including determination of optimal stimulation parameters. No studies have determined the optimal stimulation protocol for humans. At present, determination of optimal parameters is based on a trial-and-error method that will be a time-consuming process. Furthermore, we must examine the progression of these neurodegenerative diseases to determine the time frame in which intervention will be the most effective. As with any major surgery, implantation of DBS electrodes poses risk. Despite the lack of any reported major adverse effects, implantation surgery may lead to intracerebral hemorrhage, seizures, and other complications, including the risk of infection at the surgical site. There is also the matter of electrode placement. There needs to be a method to precisely determine and implant the electrodes in the most favorable location to ensure the validity of any observed effects and reduce the chances of lead migration and fracture. Although these limitations preclude the wide adoption of the method at present, ongoing research effects may make this a reality.
9. Conclusions
The cholinergic system is essential in regulating cognitive processes, and degeneration of the cholinergic system is a key component of neurocognitive disorders. Currently, there are no curative treatments for these neurocognitive disorders. However, cholinergic deep brain stimulation targeting the nucleus basalis of Meynert, a major source of cholinergic input to the cortex, has shown promise for improving cognitive function, specifically visual working memory and attention, in nonhuman primate models. Continuous stimulation at frequencies like those used in movement disorders impaired performance. However, intermittent stimulation in both adult and aged monkeys improved performance on delayed match-to-sample tasks. Additionally, these effects are accompanied by changes in neuronal activity in the prefrontal cortex, including increased firing rates, decreased trial-to-trial variability, broadened tuning, and changes in rhythmicity. Clinical studies in humans have demonstrated that DBS targeting the NB is well-tolerated and there is room for cognitive function improvement. Still, more research is needed to determine the optimal parameters of stimulation in humans and to fully elucidate the potential of cholinergic DBS as a therapeutic intervention for cognitive deficits. Overall, these findings highlight the importance of cholinergic system activation in cognitive function and suggest DBS of the NB may serve as an alternative therapy for treating neurocognitive disorders.
Author Contributions
Conceptualization, C.C. and J.M.B.; writing—original draft preparation, J.M.B. and C.C.; writing—review and editing, Z.W., S.K.B., and D.J.E.; supervision, C.C.; project administration, C.C.; funding acquisition, C.C. All authors have read and agreed to the published version of the manuscript.
Funding
The research reported in this paper was supported by NIH grants from the National Eye Institute under award number R01 EY017077 and the National Institute on Aging under award numbers RF1 AG060754 and P20 AG068082.
Data Availability Statement
No new data were created for this study. All data relevant to this research can be found in the original studies cited.
Acknowledgments
We wish to acknowledge Rye Jaffe and Junda Zhu for their helpful comments on the manuscript.
Conflicts of Interest
The authors declare no conflict of interest.
References
- Baddeley, A. Working memory: Theories, models, and controversies. Annu. Rev. Psychol. 2012, 63, 1–29. [Google Scholar] [CrossRef]
- Constantinidis, C.; Klingberg, T. The neuroscience of working memory capacity and training. Nat. Rev. Neurosci. 2016, 17, 438–449. [Google Scholar] [CrossRef]
- Subramaniam, K.; Luks, T.L.; Fisher, M.; Simpson, G.V.; Nagarajan, S.; Vinogradov, S. Computerized cognitive training restores neural activity within the reality monitoring network in schizophrenia. Neuron 2012, 73, 842–853. [Google Scholar] [CrossRef]
- Westerberg, H.; Jacobaeus, H.; Hirvikoski, T.; Clevberger, P.; Ostensson, M.L.; Bartfai, A.; Klingberg, T. Computerized working memory training after stroke—A pilot study. Brain Inj. 2007, 21, 21–29. [Google Scholar] [CrossRef]
- Hasselmo, M.E.; Sarter, M. Modes and models of forebrain cholinergic neuromodulation of cognition. Neuropsychopharmacology 2011, 36, 52–73. [Google Scholar] [CrossRef] [PubMed]
- Rountree, S.D.; Chan, W.; Pavlik, V.N.; Darby, E.J.; Siddiqui, S.; Doody, R.S. Persistent treatment with cholinesterase inhibitors and/or memantine slows clinical progression of Alzheimer disease. Alzheimers Res. Ther. 2009, 1, 7. [Google Scholar] [CrossRef] [PubMed]
- Ricciardi, E.; Pietrini, P.; Schapiro, M.B.; Rapoport, S.I.; Furey, M.L. Cholinergic modulation of visual working memory during aging: A parametric PET study. Brain Res. Bull. 2009, 79, 322–332. [Google Scholar] [CrossRef] [PubMed]
- Subramaniam, S.; Blake, D.T.; Constantinidis, C. Cholinergic Deep Brain Stimulation for Memory and Cognitive Disorders. J. Alzheimers Dis. 2021, 83, 491–503. [Google Scholar] [CrossRef] [PubMed]
- Cools, R.; Arnsten, A.F.T. Neuromodulation of prefrontal cortex cognitive function in primates: The powerful roles of monoamines and acetylcholine. Neuropsychopharmacology 2022, 47, 309–328. [Google Scholar] [CrossRef]
- Wang, M.; Datta, D.; Enwright, J.; Galvin, V.; Yang, S.T.; Paspalas, C.; Kozak, R.; Gray, D.L.; Lewis, D.A.; Arnsten, A.F.T. A novel dopamine D1 receptor agonist excites delay-dependent working memory-related neuronal firing in primate dorsolateral prefrontal cortex. Neuropharmacology 2019, 150, 46–58. [Google Scholar] [CrossRef] [PubMed]
- Vijayraghavan, S.; Wang, M.; Birnbaum, S.G.; Williams, G.V.; Arnsten, A.F. Inverted-U dopamine D1 receptor actions on prefrontal neurons engaged in working memory. Nat. Neurosci. 2007, 10, 376–384. [Google Scholar] [CrossRef] [PubMed]
- Galvin, V.C.; Arnsten, A.F.T.; Wang, M. Evolution in Neuromodulation—The Differential Roles of Acetylcholine in Higher Order Association vs. Primary Visual Cortices. Front. Neural Circuits 2018, 12, 67. [Google Scholar] [CrossRef] [PubMed]
- Hendry, S.H.; Jones, E.G.; Killackey, H.P.; Chalupa, L.M. Choline acetyltransferase-immunoreactive neurons in fetal monkey cerebral cortex. Brain Res. 1987, 465, 313–317. [Google Scholar] [CrossRef] [PubMed]
- Mesulam, M.M.; Mufson, E.J.; Levey, A.I.; Wainer, B.H. Atlas of cholinergic neurons in the forebrain and upper brainstem of the macaque based on monoclonal choline acetyltransferase immunohistochemistry and acetylcholinesterase histochemistry. Neuroscience 1984, 12, 669–686. [Google Scholar] [CrossRef]
- Saper, C.B.; Scammell, T.E.; Lu, J. Hypothalamic regulation of sleep and circadian rhythms. Nature 2005, 437, 1257–1263. [Google Scholar] [CrossRef]
- Ananth, M.R.; Rajebhosale, P.; Kim, R.; Talmage, D.A.; Role, L.W. Basal forebrain cholinergic signalling: Development, connectivity and roles in cognition. Nat. Rev. Neurosci. 2023, 24, 233–251. [Google Scholar] [CrossRef] [PubMed]
- Coyle, J.T.; Price, D.L.; DeLong, M.R. Alzheimer’s disease: A disorder of cortical cholinergic innervation. Science 1983, 219, 1184–1190. [Google Scholar] [CrossRef]
- Terry, A.V., Jr.; Buccafusco, J.J. The cholinergic hypothesis of age and Alzheimer’s disease-related cognitive deficits: Recent challenges and their implications for novel drug development. J. Pharmacol. Exp. Ther. 2003, 306, 821–827. [Google Scholar] [CrossRef]
- Sarter, M.; Bruno, J.P. Cognitive functions of cortical acetylcholine: Toward a unifying hypothesis. Brain Res. Rev. 1997, 23, 28–46. [Google Scholar] [CrossRef]
- Sarter, M.; Parikh, V. Choline transporters, cholinergic transmission and cognition. Nat. Rev. Neurosci. 2005, 6, 48–56. [Google Scholar] [CrossRef]
- Bentley, P.; Driver, J.; Dolan, R.J. Cholinergic modulation of cognition: Insights from human pharmacological functional neuroimaging. Prog. Neurobiol. 2011, 94, 360–388. [Google Scholar] [CrossRef] [PubMed]
- Sun, Y.; Yang, Y.; Galvin, V.C.; Yang, S.; Arnsten, A.F.; Wang, M. Nicotinic α4β2 Cholinergic Receptor Influences on Dorsolateral Prefrontal Cortical Neuronal Firing during a Working Memory Task. J. Neurosci. 2017, 37, 5366–5377. [Google Scholar] [CrossRef] [PubMed]
- Gratton, C.; Yousef, S.; Aarts, E.; Wallace, D.L.; D’Esposito, M.; Silver, M.A. Cholinergic, But Not Dopaminergic or Noradrenergic, Enhancement Sharpens Visual Spatial Perception in Humans. J. Neurosci. 2017, 37, 4405–4415. [Google Scholar] [CrossRef] [PubMed]
- Ballinger, E.C.; Ananth, M.; Talmage, D.A.; Role, L.W. Basal Forebrain Cholinergic Circuits and Signaling in Cognition and Cognitive Decline. Neuron 2016, 91, 1199–1218. [Google Scholar] [CrossRef] [PubMed]
- Blatt, J.; Vellage, A.; Baier, B.; Muller, N.G. The contribution of acetylcholine and dopamine to subprocesses of visual working memor—What patients with amnestic mild cognitive impairment and Parkinson’s disease can tell us. Neuropsychologia 2014, 61, 89–95. [Google Scholar] [CrossRef]
- Aigner, T.G.; Mitchell, S.J.; Aggleton, J.P.; DeLong, M.R.; Struble, R.G.; Price, D.L.; Wenk, G.L.; Pettigrew, K.D.; Mishkin, M. Transient impairment of recognition memory following ibotenic-acid lesions of the basal forebrain in macaques. Exp. Brain Res. 1991, 86, 18–26. [Google Scholar] [CrossRef] [PubMed]
- Ridley, R.M.; Bowes, P.M.; Baker, H.F.; Crow, T.J. An involvement of acetylcholine in object discrimination learning and memory in the marmoset. Neuropsychologia 1984, 22, 253–263. [Google Scholar] [CrossRef]
- Voytko, M.L.; Olton, D.S.; Richardson, R.T.; Gorman, L.K.; Tobin, J.R.; Price, D.L. Basal forebrain lesions in monkeys disrupt attention but not learning and memory. J. Neurosci. 1994, 14, 167–186. [Google Scholar] [CrossRef]
- Croxson, P.L.; Kyriazis, D.A.; Baxter, M.G. Cholinergic modulation of a specific memory function of prefrontal cortex. Nat. Neurosci. 2011, 14, 1510–1512. [Google Scholar] [CrossRef]
- Deutsch, J.A. The cholinergic synapse and the site of memory. Science 1971, 174, 788–794. [Google Scholar] [CrossRef]
- Drachman, D.A. Memory and cognitive function in man: Does the cholinergic system have a specific role? Neurology 1977, 27, 783–790. [Google Scholar] [CrossRef]
- Everitt, B.J.; Robbins, T.W. Central cholinergic systems and cognition. Annu. Rev. Psychol. 1997, 48, 649–684. [Google Scholar] [CrossRef] [PubMed]
- Bartus, R.T.; Johnson, H.R. Short-term memory in the rhesus monkey: Disruption from the anti-cholinergic scopolamine. Pharmacol. Biochem. Behav. 1976, 5, 39–46. [Google Scholar] [CrossRef] [PubMed]
- Buccafusco, J.J.; Terry, A.V., Jr.; Webster, S.J.; Martin, D.; Hohnadel, E.J.; Bouchard, K.A.; Warner, S.E. The scopolamine-reversal paradigm in rats and monkeys: The importance of computer-assisted operant-conditioning memory tasks for screening drug candidates. Psychopharmacology 2007, 199, 481–494. [Google Scholar] [CrossRef]
- Mesulam, M. The cholinergic lesion of Alzheimer’s disease: Pivotal factor or side show? Learn. Mem. 2004, 11, 43–49. [Google Scholar] [CrossRef] [PubMed]
- Bartus, R.T. On neurodegenerative diseases, models, and treatment strategies: Lessons learned and lessons forgotten a generation following the cholinergic hypothesis. Exp. Neurol. 2000, 163, 495–529. [Google Scholar] [CrossRef]
- Rogers, S.L.; Farlow, M.R.; Doody, R.S.; Mohs, R.; Friedhoff, L.T. A 24-week, double-blind, placebo-controlled trial of donepezil in patients with Alzheimer’s disease. Donepezil Study Group. Neurology 1998, 50, 136–145. [Google Scholar] [CrossRef]
- Courtney, C.; Farrell, D.; Gray, R.; Hills, R.; Lynch, L.; Sellwood, E.; Edwards, S.; Hardyman, W.; Raftery, J.; Crome, P.; et al. Long-term donepezil treatment in 565 patients with Alzheimer’s disease (AD2000): Randomised double-blind trial. Lancet 2004, 363, 2105–2115. [Google Scholar]
- Birks, J.S.; Harvey, R. Donepezil for dementia due to Alzheimer’s disease. Cochrane Database Syst. Rev. 2018, 6, CD001190. [Google Scholar] [CrossRef] [PubMed]
- Doody, R.S.; Geldmacher, D.S.; Gordon, B.; Perdomo, C.A.; Pratt, R.D. Open-label, multicenter, phase 3 extension study of the safety and efficacy of donepezil in patients with Alzheimer’s disease. Arch. Neurol. 2001, 58, 427–433. [Google Scholar] [CrossRef]
- Winblad, B.; Engedal, K.; Soininen, H.; Verhey, F.; Waldemar, G.; Wimo, A.; Wetterholm, A.L.; Zhang, R.; Haglund, A.; Subbiah, P. A 1-year, randomized, placebo-controlled study of donepezil in patients with mild to moderate AD. Neurology 2001, 57, 489–495. [Google Scholar] [CrossRef] [PubMed]
- Buccafusco, J.J.; Terry, A.V. Donepezil-induced improvement in delayed matching accuracy by young and old rhesus monkeys. J. Mol. Neurosci. 2004, 24, 85–91. [Google Scholar] [CrossRef] [PubMed]
- Francis, P.T.; Palmer, A.M.; Snape, M.; Wilcock, G.K. The cholinergic hypothesis of Alzheimer’s disease: A review of progress. J. Neurol. Neurosurg. Psychiatry 1999, 66, 137–147. [Google Scholar] [CrossRef] [PubMed]
- Farlow, M.R.; Salloway, S.; Tariot, P.N.; Yardley, J.; Moline, M.L.; Wang, Q.; Brand-Schieber, E.; Zou, H.; Hsu, T.; Satlin, A. Effectiveness and tolerability of high-dose (23 mg/d) versus standard-dose (10 mg/d) donepezil in moderate to severe Alzheimer’s disease: A 24-week, randomized, double-blind study. Clin. Ther. 2010, 32, 1234–1251. [Google Scholar] [CrossRef]
- Yoshida, T.; Ha-Kawa, S.; Yoshimura, M.; Nobuhara, K.; Kinoshita, T.; Sawada, S. Effectiveness of treatment with donepezil hydrochloride and changes in regional cerebral blood flow in patients with Alzheimer’s disease. Ann. Nucl. Med. 2007, 21, 257–265. [Google Scholar] [CrossRef]
- Riley, M.R.; Constantinidis, C. Role of prefrontal persistent activity in working memory. Front. Syst. Neurosci. 2016, 9, 181. [Google Scholar] [CrossRef]
- Lundqvist, M.; Herman, P.; Miller, E.K. Working Memory: Delay Activity, Yes! Persistent Activity? Maybe Not. J. Neurosci. 2018, 38, 7013–7019. [Google Scholar] [CrossRef]
- Olesen, P.J.; Westerberg, H.; Klingberg, T. Increased prefrontal and parietal activity after training of working memory. Nat. Neurosci. 2004, 7, 75–79. [Google Scholar] [CrossRef]
- Cai, Y.; Yang, C.; Wang, S.; Xue, G. The Neural Mechanism Underlying Visual Working Memory Training and Its Limited Transfer Effect. J. Cogn. Neurosci. 2022, 34, 2082–2099. [Google Scholar] [CrossRef]
- Bentley, P.; Driver, J.; Dolan, R.J. Cholinesterase inhibition modulates visual and attentional brain responses in Alzheimer’s disease and health. Brain 2008, 131, 409–424. [Google Scholar] [CrossRef]
- Funahashi, S.; Bruce, C.J.; Goldman-Rakic, P.S. Mnemonic coding of visual space in the monkey’s dorsolateral prefrontal cortex. J. Neurophysiol. 1989, 61, 331–349. [Google Scholar] [CrossRef] [PubMed]
- Meyer, T.; Qi, X.L.; Stanford, T.R.; Constantinidis, C. Stimulus selectivity in dorsal and ventral prefrontal cortex after training in working memory tasks. J. Neurosci. 2011, 31, 6266–6276. [Google Scholar] [CrossRef] [PubMed]
- Hart, E.; Huk, A.C. Recurrent circuit dynamics underlie persistent activity in the macaque frontoparietal network. eLife 2020, 9, e52460. [Google Scholar] [CrossRef] [PubMed]
- Compte, A.; Brunel, N.; Goldman-Rakic, P.S.; Wang, X.J. Synaptic mechanisms and network dynamics underlying spatial working memory in a cortical network model. Cereb. Cortex 2000, 10, 910–923. [Google Scholar] [CrossRef]
- Xie, Y.; Liu, Y.H.; Constantinidis, C.; Zhou, X. Neural Mechanisms of Working Memory Accuracy Revealed by Recurrent Neural Networks. Front. Syst. Neurosci. 2022, 16. [Google Scholar]
- Seung, H.S.; Lee, D.D.; Reis, B.Y.; Tank, D.W. Stability of the memory of eye position in a recurrent network of conductance-based model neurons. Neuron 2000, 26, 259–271. [Google Scholar] [CrossRef]
- Wimmer, K.; Nykamp, D.Q.; Constantinidis, C.; Compte, A. Bump attractor dynamics in prefrontal cortex explains behavioral precision in spatial working memory. Nat. Neurosci. 2014, 17, 431–439. [Google Scholar] [CrossRef]
- Jaffe, R.J.; Constantinidis, C. Working Memory: From Neural Activity to the Sentient Mind. Compr. Physiol. 2021, 11, 2547–2587. [Google Scholar] [CrossRef]
- Disney, A.A. Neuromodulatory Control of Early Visual Processing in Macaque. Annu. Rev. Vis. Sci. 2021, 7, 181–199. [Google Scholar] [CrossRef]
- Disney, A.A.; Aoki, C.; Hawken, M.J. Gain modulation by nicotine in macaque v1. Neuron 2007, 56, 701–713. [Google Scholar] [CrossRef]
- Herrero, J.L.; Roberts, M.J.; Delicato, L.S.; Gieselmann, M.A.; Dayan, P.; Thiele, A. Acetylcholine contributes through muscarinic receptors to attentional modulation in V1. Nature 2008, 454, 1110–1114. [Google Scholar] [CrossRef] [PubMed]
- Veith, V.K.; Quigley, C.; Treue, S. Cholinergic manipulations affect sensory responses but not attentional enhancement in macaque MT. BMC Biol. 2021, 19, 49. [Google Scholar] [CrossRef] [PubMed]
- Yu, A.J.; Dayan, P. Uncertainty, neuromodulation, and attention. Neuron 2005, 46, 681–692. [Google Scholar] [CrossRef] [PubMed]
- Pinto, L.; Goard, M.J.; Estandian, D.; Xu, M.; Kwan, A.C.; Lee, S.H.; Harrison, T.C.; Feng, G.; Dan, Y. Fast modulation of visual perception by basal forebrain cholinergic neurons. Nat. Neurosci. 2013, 16, 1857–1863. [Google Scholar] [CrossRef] [PubMed]
- Disney, A.A.; Robert, J.S. Translational implications of the anatomical nonequivalence of functionally equivalent cholinergic circuit motifs. Proc. Natl. Acad. Sci. USA 2019, 116, 26181–26186. [Google Scholar] [CrossRef]
- Thiele, A.; Bellgrove, M.A. Neuromodulation of Attention. Neuron 2018, 97, 769–785. [Google Scholar] [CrossRef]
- Yang, Y.; Paspalas, C.D.; Jin, L.E.; Picciotto, M.R.; Arnsten, A.F.; Wang, M. Nicotinic α7 receptors enhance NMDA cognitive circuits in dorsolateral prefrontal cortex. Proc. Natl. Acad. Sci. USA 2013, 110, 12078–12083. [Google Scholar] [CrossRef]
- Dasilva, M.; Brandt, C.; Gotthardt, S.; Gieselmann, M.A.; Distler, C.; Thiele, A. Cell class-specific modulation of attentional signals by acetylcholine in macaque frontal eye field. Proc. Natl. Acad. Sci. USA 2019, 116, 20180–20189. [Google Scholar] [CrossRef]
- Zhou, X.; Qi, X.L.; Douglas, K.; Palaninathan, K.; Kang, H.S.; Buccafusco, J.J.; Blake, D.T.; Constantinidis, C. Cholinergic modulation of working memory activity in primate prefrontal cortex. J. Neurophysiol. 2011, 106, 2180–2188. [Google Scholar] [CrossRef]
- Major, A.J.; Vijayraghavan, S.; Everling, S. Muscarinic Attenuation of Mnemonic Rule Representation in Macaque Dorsolateral Prefrontal Cortex during a Pro- and Anti-Saccade Task. J. Neurosci. 2015, 35, 16064–16076. [Google Scholar] [CrossRef]
- Galvin, V.C.; Yang, S.T.; Paspalas, C.D.; Yang, Y.; Jin, L.E.; Datta, D.; Morozov, Y.M.; Lightbourne, T.C.; Lowet, A.S.; Rakic, P.; et al. Muscarinic M1 Receptors Modulate Working Memory Performance and Activity via KCNQ Potassium Channels in the Primate Prefrontal Cortex. Neuron 2020, 106, 649–661.e4. [Google Scholar] [CrossRef]
- Major, A.J.; Vijayraghavan, S.; Everling, S. Cholinergic Overstimulation Attenuates Rule Selectivity in Macaque Prefrontal Cortex. J. Neurosci. 2018, 38, 1137–1150. [Google Scholar] [CrossRef] [PubMed]
- Vijayraghavan, S.; Major, A.J.; Everling, S. Muscarinic M1 Receptor Overstimulation Disrupts Working Memory Activity for Rules in Primate Prefrontal Cortex. Neuron 2018, 98, 1256–1268.e1254. [Google Scholar] [CrossRef] [PubMed]
- Galvin, V.C.; Arnsten, A.F.T.; Wang, M. Involvement of Nicotinic Receptors in Working Memory Function. Curr. Top. Behav. Neurosci. 2020, 45, 89–99. [Google Scholar] [CrossRef] [PubMed]
- Williams, G.V.; Goldman-Rakic, P.S. Modulation of memory fields by dopamine D1 receptors in prefrontal cortex. Nature 1995, 376, 572–575. [Google Scholar] [CrossRef]
- Freund, H.J.; Kuhn, J.; Lenartz, D.; Mai, J.K.; Schnell, T.; Klosterkoetter, J.; Sturm, V. Cognitive functions in a patient with Parkinson-dementia syndrome undergoing deep brain stimulation. Arch. Neurol. 2009, 66, 781–785. [Google Scholar] [CrossRef] [PubMed]
- Saper, C.B.; Fuller, P.M. Wake-sleep circuitry: An overview. Curr. Opin. Neurobiol. 2017, 44, 186–192. [Google Scholar] [CrossRef]
- Walker, L.C.; Price, D.L.; Young, W.S., 3rd. GABAergic neurons in the primate basal forebrain magnocellular complex. Brain Res. 1989, 499, 188–192. [Google Scholar] [CrossRef]
- Kim, T.; Thankachan, S.; McKenna, J.T.; McNally, J.M.; Yang, C.; Choi, J.H.; Chen, L.; Kocsis, B.; Deisseroth, K.; Strecker, R.E.; et al. Cortically projecting basal forebrain parvalbumin neurons regulate cortical gamma band oscillations. Proc. Natl. Acad. Sci. USA 2015, 112, 3535–3540. [Google Scholar] [CrossRef]
- Semba, K. Multiple output pathways of the basal forebrain: Organization, chemical heterogeneity, and roles in vigilance. Behav. Brain Res. 2000, 115, 117–141. [Google Scholar] [CrossRef]
- Detari, L.; Rasmusson, D.D.; Semba, K. The role of basal forebrain neurons in tonic and phasic activation of the cerebral cortex. Prog. Neurobiol. 1999, 58, 249–277. [Google Scholar] [CrossRef] [PubMed]
- Granger, A.J.; Wang, W.; Robertson, K.; El-Rifai, M.; Zanello, A.F.; Bistrong, K.; Saunders, A.; Chow, B.W.; Nunez, V.; Turrero Garcia, M.; et al. Cortical ChAT(+) neurons co-transmit acetylcholine and GABA in a target- and brain-region-specific manner. eLife 2020, 9, e57749. [Google Scholar] [CrossRef] [PubMed]
- Granger, A.J.; Mulder, N.; Saunders, A.; Sabatini, B.L. Cotransmission of acetylcholine and GABA. Neuropharmacology 2016, 100, 40–46. [Google Scholar] [CrossRef] [PubMed]
- Vitek, J.L. Mechanisms of deep brain stimulation: Excitation or inhibition. Mov. Disord. 2002, 17, S69–S72. [Google Scholar] [CrossRef]
- Mahlknecht, P.; Limousin, P.; Foltynie, T. Deep brain stimulation for movement disorders: Update on recent discoveries and outlook on future developments. J. Neurol. 2015, 262, 2583–2595. [Google Scholar] [CrossRef]
- Wichmann, T.; DeLong, M.R. Deep Brain Stimulation for Neurologic and Neuropsychiatric Disorders. Neuron 2006, 52, 197–204. [Google Scholar] [CrossRef]
- Montemurro, N.; Aliaga, N.; Graff, P.; Escribano, A.; Lizana, J. New Targets and New Technologies in the Treatment of Parkinson’s Disease: A Narrative Review. Int. J. Environ. Res. Public Health 2022, 19, 8799. [Google Scholar] [CrossRef]
- Popovych, V.O.; Lysyansky, B.; Tass, P.A. Closed-loop deep brain stimulation by pulsatile delayed feedback with increased gap between pulse phases. Sci. Rep. 2017, 7, 1033. [Google Scholar] [CrossRef]
- Kent, A.R.; Grill, W.M. Recording evoked potentials during deep brain stimulation: Development and validation of instrumentation to suppress the stimulus artefact. J. Neural Eng. 2012, 9, 036004. [Google Scholar] [CrossRef]
- Stoney, S.D.; Thompson, W.D.; Asanuma, H. Excitation of pyramidal tract cells by intracortical microstimulation: Effective extent of stimulating current. J. Neurophysiol. 1968, 31, 659–669. [Google Scholar] [CrossRef]
- Murasugi, C.M.; Salzman, C.D.; Newsome, W.T. Microstimulation in visual area MT: Effects of varying pulse amplitude and frequency. J. Neurosci. 1993, 13, 1719–1729. [Google Scholar] [CrossRef] [PubMed]
- McIntyre, C.C.; Mori, S.; Sherman, D.L.; Thakor, V.N.; Vitek, J.L. Electric field and stimulating influence generated by deep brain stimulation of the subthalamic nucleus. Clin. Neurophysiol. 2004, 115, 589–595. [Google Scholar] [CrossRef] [PubMed]
- Grossman, N.; Bono, D.; Dedic, N.; Kodandaramaiah, S.B.; Rudenko, A.; Suk, H.J.; Cassara, A.M.; Neufeld, E.; Kuster, N.; Tsai, L.H.; et al. Noninvasive Deep Brain Stimulation via Temporally Interfering Electric Fields. Cell 2017, 169, 1029–1041.e1016. [Google Scholar] [CrossRef] [PubMed]
- Fan, S.; Liu, D.; Shi, L.; Meng, F.; Fang, H.; Liu, H.; Zhang, H.; Yang, A.; Zhang, J. Differential Effects of Subthalamic Nucleus and Globus Pallidus Internus Deep Brain Stimulation on Motor Subtypes in Parkinson’s Disease. World Neurosurg. 2022, 164, e245–e255. [Google Scholar] [CrossRef] [PubMed]
- Liu, R.; Crawford, J.; Callahan, P.M.; Terry, V.A.; Constantinidis, C.; Blake, D.T. Intermittent Stimulation of the Nucleus Basalis of Meynert Improves Working Memory in Adult Monkeys. Curr. Biol. 2017, 27, 2640–2646. [Google Scholar] [CrossRef]
- Liu, R.; Crawford, J.; Callahan, P.M.; Terry, A.V.; Constantinidis, C.; Blake, D.T. Intermittent stimulation in the nucleus basalis of meynert improves sustained attention in rhesus monkeys. Neuropharmacology 2018, 137, 202–210. [Google Scholar] [CrossRef]
- Chung, S.; Bava, J.; Wang, Z.; Garin, C.M.; Clemencich, K.; Pennington, K.R.; Bick, S.K.; Englot, D.J.; Blake, D.T.; Constantinidis, C. Intermittent stimulation of the nucleus basalis improves working memory in aged monkeys. Soc. Neurosci. Abstr. 2022, 654, 623. [Google Scholar]
- Qi, X.L.; Liu, R.; Singh, B.; Bestue, D.; Compte, A.; Vazdarjanova, A.I.; Blake, D.T.; Constantinidis, C. Nucleus basalis stimulation enhances working memory by stabilizing stimulus representations in primate prefrontal cortical activity. Cell Rep. 2021, 36, 109469. [Google Scholar] [CrossRef]
- Li, W.; Piech, V.; Gilbert, C.D. Perceptual learning and top-down influences in primary visual cortex. Nat. Neurosci. 2004, 7, 651–657. [Google Scholar] [CrossRef]
- Yang, T.; Maunsell, J.H. The effect of perceptual learning on neuronal responses in monkey visual area V4. J. Neurosci. 2004, 24, 1617–1626. [Google Scholar] [CrossRef]
- Raiguel, S.; Vogels, R.; Mysore, S.G.; Orban, G.A. Learning to see the difference specifically alters the most informative V4 neurons. J. Neurosci. 2006, 26, 6589–6602. [Google Scholar] [CrossRef]
- Sanayei, M.; Chen, X.; Chicharro, D.; Distler, C.; Panzeri, S.; Thiele, A. Perceptual learning of fine contrast discrimination changes neuronal tuning and population coding in macaque V4. Nat. Commun. 2018, 9, 4238. [Google Scholar] [CrossRef] [PubMed]
- Ma, W.J.; Beck, J.M.; Latham, P.E.; Pouget, A. Bayesian inference with probabilistic population codes. Nat. Neurosci. 2006, 9, 1432–1438. [Google Scholar] [CrossRef]
- Zhang, K.; Sejnowski, T.J. Neuronal tuning: To sharpen or broaden? Neural Comput. 1999, 11, 75–84. [Google Scholar] [CrossRef] [PubMed]
- Butts, D.A.; Goldman, M.S. Tuning curves, neuronal variability, and sensory coding. PLoS Biol. 2006, 4, e92. [Google Scholar] [CrossRef]
- Pouget, A.; Deneve, S.; Ducom, J.C.; Latham, P.E. Narrow versus wide tuning curves: What’s best for a population code? Neural Comput. 1999, 11, 85–90. [Google Scholar] [CrossRef]
- Stein, H.; Barbosa, J.; Compte, A. Towards biologically constrained attractor models of schizophrenia. Curr. Opin. Neurobiol. 2021, 70, 171–181. [Google Scholar] [CrossRef]
- Qi, X.L.; Meyer, T.; Stanford, T.R.; Constantinidis, C. Changes in Prefrontal Neuronal Activity after Learning to Perform a Spatial Working Memory Task. Cereb. Cortex 2011, 21, 2722–2732. [Google Scholar] [CrossRef]
- Qi, X.L.; Constantinidis, C. Neural changes after training to perform cognitive tasks. Behav. Brain Res. 2013, 241, 235–243. [Google Scholar] [CrossRef] [PubMed]
- Thiele, A.; Herrero, J.L.; Distler, C.; Hoffmann, K.P. Contribution of cholinergic and GABAergic mechanisms to direction tuning, discriminability, response reliability, and neuronal rate correlations in macaque middle temporal area. J. Neurosci. 2012, 32, 16602–16615. [Google Scholar] [CrossRef]
- Minces, V.; Pinto, L.; Dan, Y.; Chiba, A.A. Cholinergic shaping of neural correlations. Proc. Natl. Acad. Sci. USA 2017, 114, 5725–5730. [Google Scholar] [CrossRef]
- Singh, B.; Qi, X.L.; Blake, D.T.; Constantinidis, C. Rhythmicity of Prefrontal Local Field Potentials after Nucleus Basalis Stimulation. eNeuro 2022, 9, ENEURO.0380-21.2022. [Google Scholar] [CrossRef]
- Kuhn, J.; Hardenacke, K.; Lenartz, D.; Gruendler, T.; Ullsperger, M.; Bartsch, C.; Mai, J.K.; Zilles, K.; Bauer, A.; Matusch, A.; et al. Deep brain stimulation of the nucleus basalis of Meynert in Alzheimer’s dementia. Mol. Psychiatry 2015, 20, 353–360. [Google Scholar] [CrossRef] [PubMed]
- Gratwicke, J.; Zrinzo, L.; Kahan, J.; Peters, A.; Beigi, M.; Akram, H.; Hyam, J.; Oswal, A.; Day, B.; Mancini, L.; et al. Bilateral Deep Brain Stimulation of the Nucleus Basalis of Meynert for Parkinson Disease Dementia: A Randomized Clinical Trial. JAMA Neurol. 2017, 75, 169–178. [Google Scholar] [CrossRef] [PubMed]
- Sasikumar, S.; Cohn, M.; Harmsen, I.E.; Loh, A.; Cho, S.S.; Saenz-Farret, M.; Maciel, R.; Soh, D.; Boutet, A.; Germann, J.; et al. Single-Trajectory Multiple-Target Deep Brain Stimulation for Parkinsonian Mobility and Cognition. Mov. Disord. 2022, 37, 635–640. [Google Scholar] [CrossRef]
- Nombela, C.; Lozano, A.; Villanueva, C.; Barcia, J.A. Simultaneous Stimulation of the Globus Pallidus Interna and the Nucleus Basalis of Meynert in the Parkinson-Dementia Syndrome. Dement. Geriatr. Cogn. Disord. 2019, 47, 19–28. [Google Scholar] [CrossRef]
- Maltete, D.; Wallon, D.; Bourilhon, J.; Lefaucheur, R.; Danaila, T.; Thobois, S.; Defebvre, L.; Dujardin, K.; Houeto, J.L.; Godefroy, O.; et al. Nucleus Basalis of Meynert Stimulation for Lewy Body Dementia: A Phase I Randomized Clinical Trial. Neurology 2021, 96, e684–e697. [Google Scholar] [CrossRef]
- Jiang, Y.; Yuan, T.S.; Chen, Y.C.; Guo, P.; Lian, T.H.; Liu, Y.Y.; Liu, W.; Bai, Y.T.; Zhang, Q.; Zhang, W.; et al. Deep brain stimulation of the nucleus basalis of Meynert modulates hippocampal-frontoparietal networks in patients with advanced Alzheimer’s disease. Transl. Neurodegener. 2022, 11, 51. [Google Scholar] [CrossRef]
- Bogdan, I.D.; Oterdoom, D.L.M.; van Laar, T.; Huitema, R.B.; Odekerken, V.J.; Boel, J.A.; de Bie, R.M.A.; van Dijk, J.M.C.; on behalf of the NSTAPS Study Group. Serendipitous Stimulation of Nucleus Basalis of Meynert—The Effect of Unintentional, Long-Term High-Frequency Stimulation on Cognition in Parkinson’s Disease. J. Clin. Med. 2022, 11, 337. [Google Scholar] [CrossRef]
- Gratwicke, J.; Oswal, A.; Akram, H.; Jahanshahi, M.; Hariz, M.; Zrinzo, L.; Foltynie, T.; Litvak, V. Resting state activity and connectivity of the nucleus basalis of Meynert and globus pallidus in Lewy body dementia and Parkinson’s disease dementia. NeuroImage 2020, 221, 117184. [Google Scholar] [CrossRef]




Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).