
Sex Differences in Cognitive-Motor Dual-Task Training Effects and in Brain Processing of Semi-Elite Basketball Players


Abstract
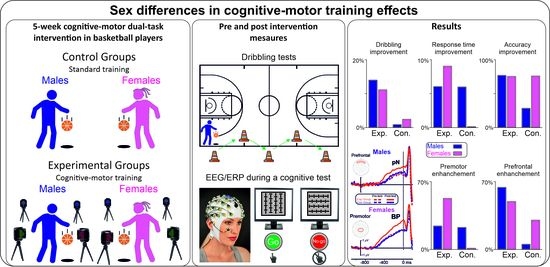
1. Introduction
2. Materials and Methods
2.1. Participants
2.2. Procedure
2.2.1. Motor Training
- The athlete was activated from a stationary position, in front of a cone, and alternated dribbles with the right hand and left hand, under the legs, behind the back, in/out front, and side wave with the right and left hand.
- The athlete uses the cone positioned at 5.5 m as a reference. He sprints by dribbling with his right hand up to the cone and then performs backward dribbling with his right hand until he reaches the starting position where he performs a front-hand change and restarts with his left hand. Each time he/she returns to the starting position, the athlete uses different hand changes to restart (e.g., frontal, under the legs, behind the back).
- The athlete uses two positioned hurdles (refer to Figure 1) and performed the exercise similar to the previous one but using different distances in retreat depending on the cone and the different hand changes.
2.2.2. Cognitive-Motor Dual-Task Training (CMDT)
- To activate, the athlete must close only the “Witty-SEM” with the “Blue 7” as quickly as possible by performing a different hand change each time (e.g., frontal, under the legs, behind the back).
- The athlete is positioned in front of two devices. “A” is green all the time, while “B” changes colour and configuration. The athlete must perform different hand changes depending on the colour of the “B” photocell (e.g., blue-under the legs; red-behind the back) and by dribbling close to the “A”.
- The athlete must be ready to discriminate the photocell with the different configurations within a few milliseconds while performing hand changes (e.g., frontal). Once he/she detects the different devices, he/she must sprint-dribble and close it and then perform backward dribbles to return to the starting position and continue with the exercise.
2.2.3. Basketball Performance Tests
2.2.4. Cognitive Test
Behavioral Data
EEG Recording
2.3. Statistical Analysis
3. Results
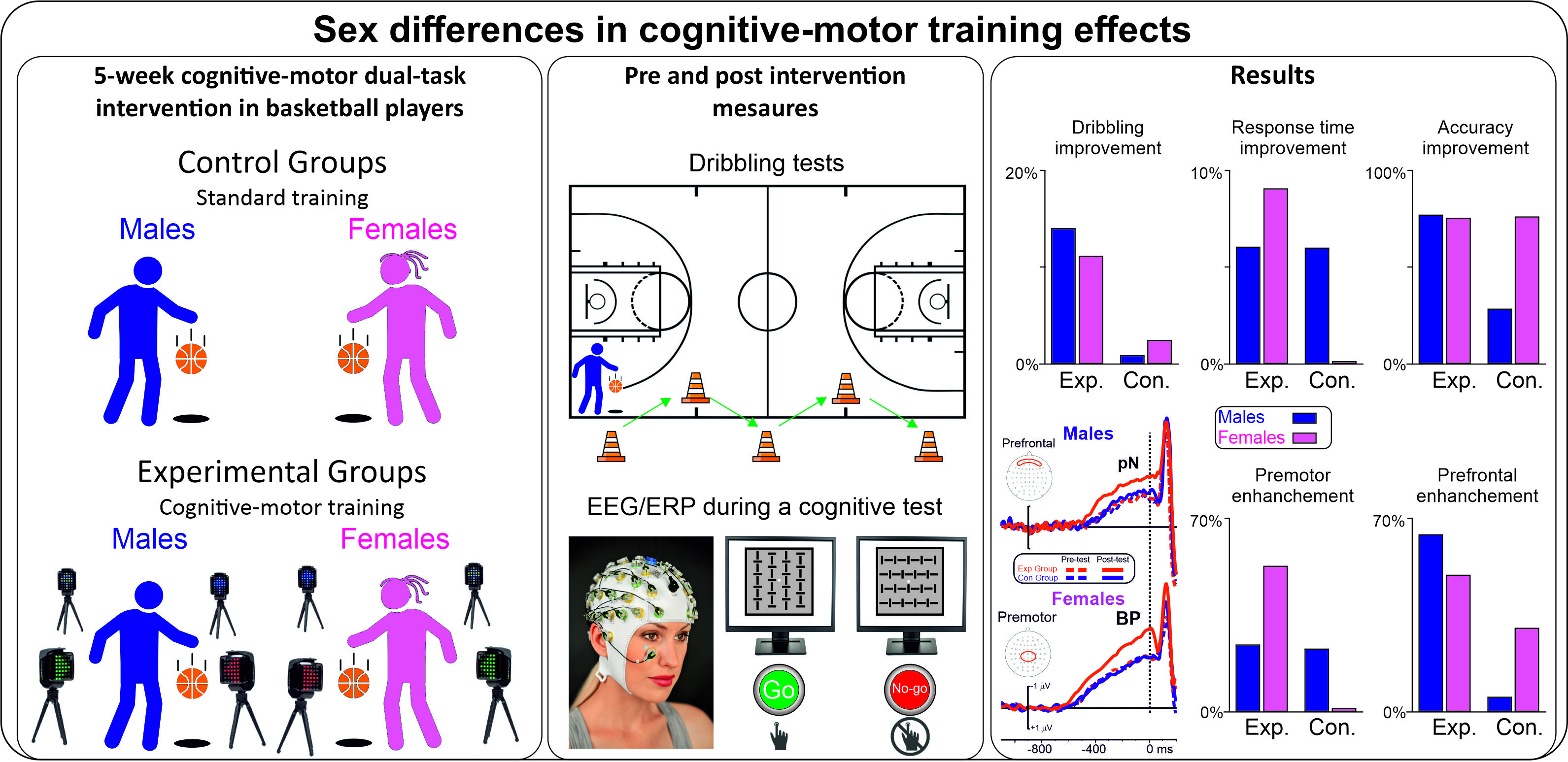
3.1. Basketball Performance Tests
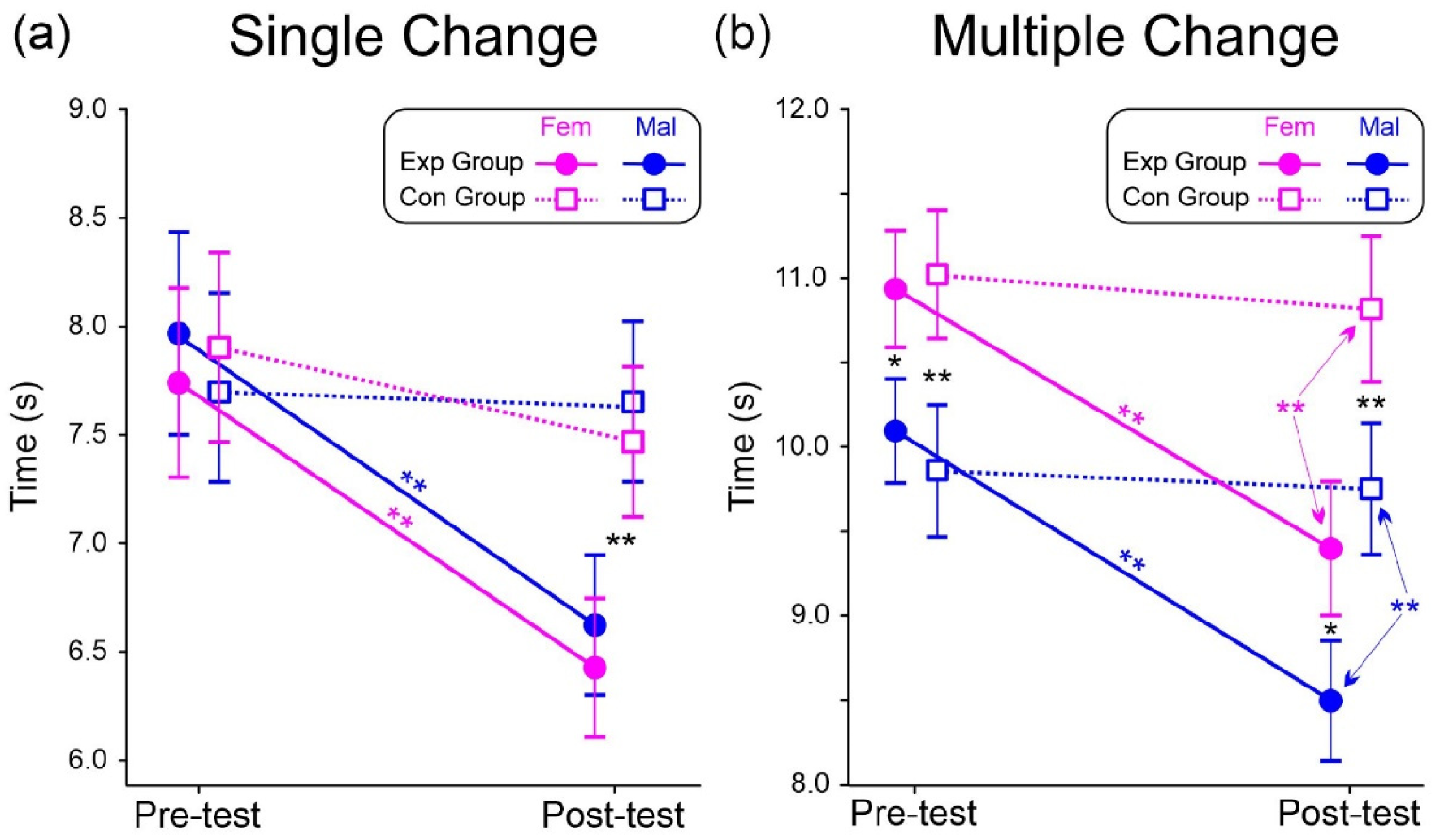
3.2. Cognitive Test: Behavioral Data
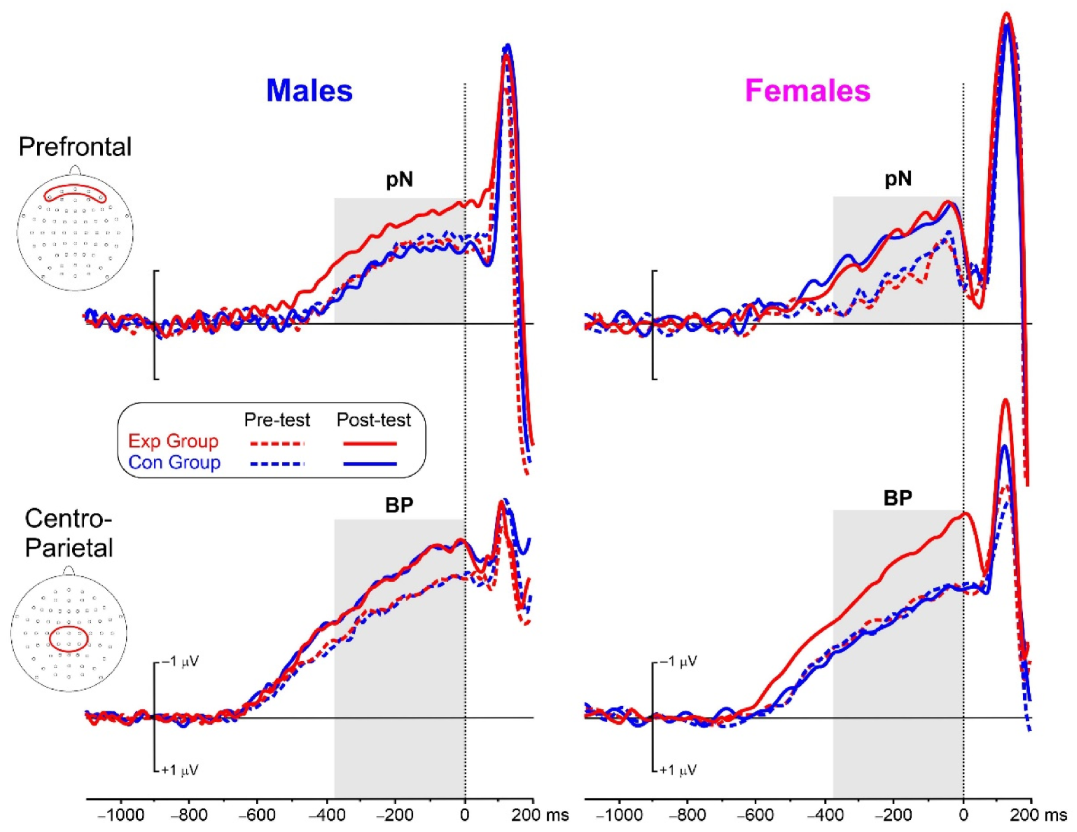
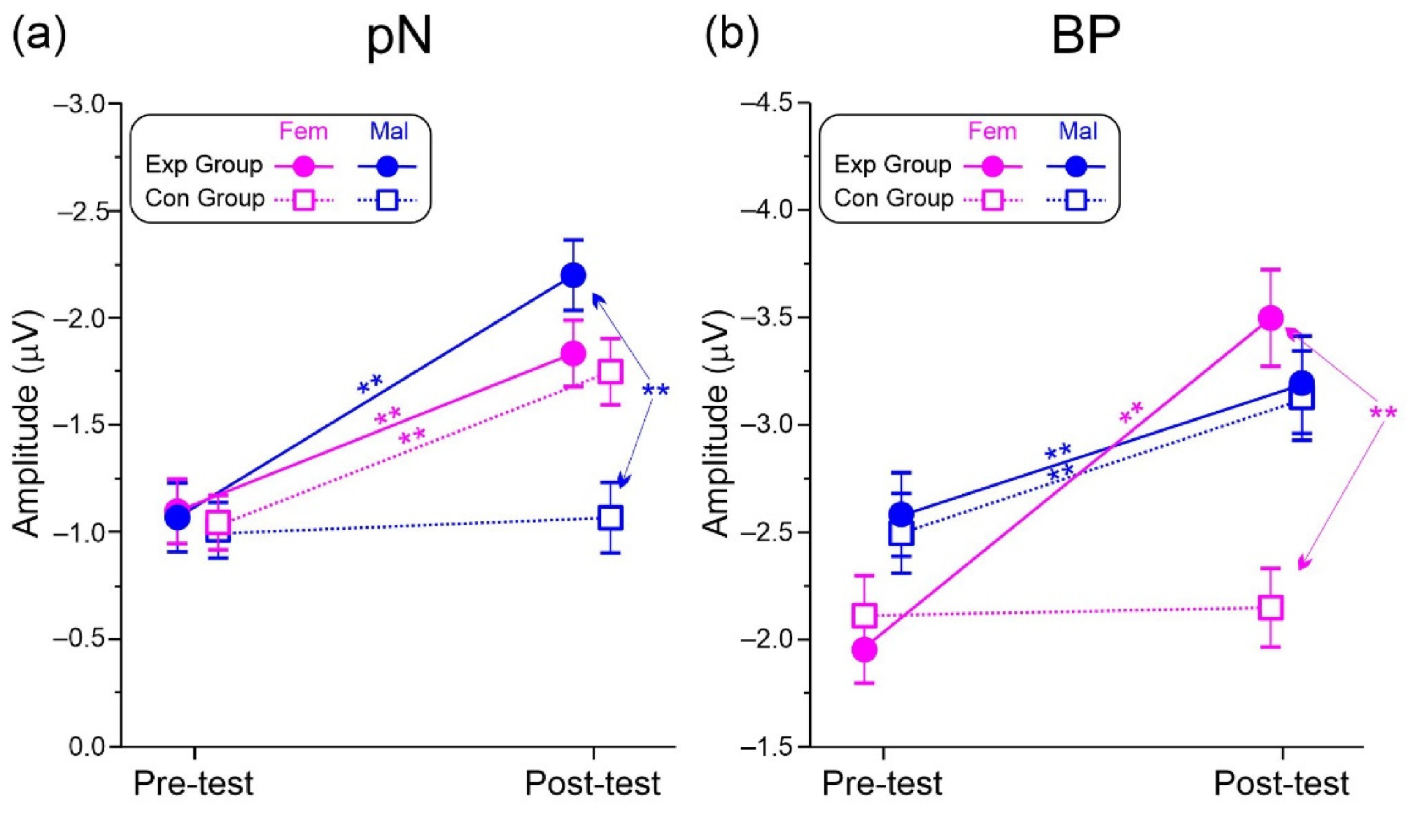
3.3. Cognitive Test: ERP Results
3.4. Correlation Analysis
3.5. Post-Hoc Power Analysis
4. Discussion
5. Limitations
6. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Wollesen, B.; Janssen, T.I.; Müller, H.; Voelcker-Rehage, C. Effects of cognitive-motor dual task training on cognitive and physical performance in healthy children and adolescents: A scoping review. Acta Psychol. 2022, 224, 103498. [Google Scholar] [CrossRef] [PubMed]
- Wollesen, B.; Wildbredt, A.; van Schooten, K.S.; Lim, M.L.; Delbaere, K. The effects of cognitive-motor training interventions on executive functions in older people: A systematic review and meta-analysis. Eur. Rev. Aging Phys. Act. 2020, 17, 1–22. [Google Scholar] [CrossRef] [PubMed]
- Lucia, S.; Bianco, V.; Boccacci, L.; Di Russo, F. Effects of a Cognitive-Motor Training on Anticipatory Brain Functions and Sport Performance in Semi-Elite Basketball Players. Brain Sci. 2022, 12, 68. [Google Scholar] [CrossRef] [PubMed]
- Lucia, S.; Bianco, V.; Di Russo, F. Specific effect of a cognitive-motor dual-task training on sport performance and brain processing associated with decision-making in semi-elite basketball players. Psychol. Sport Exerc. 2023, 64, 102302. [Google Scholar] [CrossRef]
- Halpern, D.F.; Collaer, M.L. Sex differences in visuospatial abilities: More than meets the eye. In The Cambridge Handbook of Visuospatial Thinking; Shah, P., Miyake, A., Eds.; Cambridge University Press: New York, NY, USA, 2005; pp. 170–212. [Google Scholar] [CrossRef]
- Millslagle, D.G. Recognition accuracy by experienced men and women players of basketball. Percept. Mot. Ski. 2002, 95, 163–172. [Google Scholar] [CrossRef]
- Weigelt, M.; Memmert, D. The mental rotation ability of expert basketball players: Identifying on-court plays. Res. Q. Exerc. Sport 2021, 92, 137–145. [Google Scholar] [CrossRef]
- Milley, K.R.; Ouellette, G.P. Putting attention on the spot in coaching: Shifting to an external focus of attention with imagery techniques to improve basketball free-throw shooting performance. Front. Psychol. 2021, 12, 645676. [Google Scholar] [CrossRef]
- El Moutaraji, I.; Lotfi, S.; Talbi, M. Cross Effect of Exercise, Sex and Level of Sport Expertise on Cognition: Visual Perception, Information Processing and Motor Response. J. Hum. Mov. Sport. Sci. 2021, 99, 412–420. [Google Scholar] [CrossRef]
- Kellis, S.E.; Tsitskaris, G.K.; Nikopoulou, M.D.; Mousikou, K.C. The evaluation of jumping ability of male and female basketball players according to their chronological age and major leagues. J. Strength Cond. Res. 1999, 13, 40–46. [Google Scholar]
- Gómez, M.A.; Lorenzo, A.; Sampaio, J. Men and women’s basketball teams use different game tactics to score points in fast-breaks, transition offenses and set-plays. Iber. Congr. Basketb. Res. 2007, 4, 1–4. [Google Scholar]
- Scanlan, A.T.; Dascombe, B.J.; Kidcaff, A.P.; Peucker, J.L.; Dalbo, V.J. Sex-specific activity demands experienced during semiprofessional basketball game play. Int. J. Sport. Physiol. Perform. 2015, 10, 618–625. [Google Scholar] [CrossRef]
- Bianco, V.; Berchicci, M.; Quinzi, F.; Perri, R.L.; Spinelli, D.; Di Russo, F. Females are more proactive, males are more reactive: Neural basis of the sex-related speed/accuracy trade-off in visuo-motor tasks. Brain Struct. Funct. 2019, 225, 187–201. [Google Scholar] [CrossRef]
- Silverman, I.W. Sex diferences in simple visual reaction time: A historical meta-analysis. Sex Roles 2006, 54, 57–68. [Google Scholar] [CrossRef]
- Fozard, J.L.; Vercruyssen, M.; Reynolds, S.L.; Hancock, P.A.; Quilter, R.E. Age diferences and changes in reaction time: The Baltimore Longitudinal Study of Aging. J. Gerontol. 1994, 49, P179. [Google Scholar] [CrossRef]
- Shibasaki, H.; Hallett, M. What is the Bereitschaftspotential? Clin. Neurophysiol. 2006, 117, 2341–2356. [Google Scholar] [CrossRef]
- Di Russo, F.; Berchicci, M.; Bianco, V.; Perri, R.L.; Pitzalis, S.; Quinzi, F.; Spinelli, D. Normative event-related potentials from sensory and cognitive tasks reveal occipital and frontal activities prior and following visual events. Neuroimage 2019, 196, 173–187. [Google Scholar] [CrossRef]
- Faul, F.; Erdfelder, E.; Buchner, A.; Lang, A.-G. Statistical power analyses using G*Power 3.1: Tests for correlation and regression analyses. Behav. Res. Methods 2009, 41, 1149–1160. [Google Scholar] [CrossRef]
- Oldfield, R.C. The assessment and analysis of handedness: The Edinburgh inventory. Neuropsychologia 1971, 9, 97–113. [Google Scholar] [CrossRef]
- Swann, C.; Moran, A.; Piggott, D. Defining elite athletes: Issues in the study of expert performance in sport psychology. Psychol. Sport Exerc. 2015, 16, 3–14, reprinted in Brain Sci. 2022, 12, 17–37. [Google Scholar] [CrossRef]
- Capobianco, A. Insegnare la Pallacanestro. Guida Didattica per il Corso Allievo Allenatore; Calzetti & Mariucci: Torgiano, Italy, 2014. [Google Scholar]
- Ranisavljev, I.; Mandic, R.; Cosic, M.; Blagojevic, P.; Dopsaj, M. NBA Pre-Draft Combine is the weak predictor of rookie basketball player’s performance. J. Hum. Sport Exerc. 2021, 16, 493–502. [Google Scholar] [CrossRef]
- Ramirez-Campillo, R.; Gentil, P.; Moran, J.; Dalbo, V.J.; Scanlan, A.T. Dribble deficit enables measurement of dribbling speed independent of sprinting speed in collegiate, male, basketball players. J. Strength Cond. Res. 2021, 35, 2040–2045. [Google Scholar] [CrossRef]
- Ivanović, J.; Kukić, F.; Greco, G.; Koropanovski, N.; Jakovljević, S.; Dopsaj, M. Specific Physical Ability Prediction in Youth Basketball Players According to Playing Position. Int. J. Environ. Res. Public Health 2022, 19, 977. [Google Scholar] [CrossRef] [PubMed]
- Luck, S.J.; Gaspelin, N. How to get statistically significant effects in any ERP experiment (and why you shouldn’t). Psychophysiology 2017, 54, 1–14. [Google Scholar] [CrossRef] [PubMed]
- Beckerman, H.; Roebroeck, M.E.; Lankhorst, G.J.; Becher, J.G.; Bezemer, P.D.; Verbeek, A.L.M. Smallest real difference: A link between reproducibility and responsiveness. Qual. Life Res. 2001, 10, 571–578. [Google Scholar] [CrossRef] [PubMed]
- Spierer, D.K.; Petersen, R.A.; Duffy, K.; Corcoran, B.M.; Rawls-Martin, T. Sex influence on response time to sensory stimuli. J. Strength Cond. Res. 2010, 24, 957–963. [Google Scholar] [CrossRef]
- O’Brien-Smith, J.; Bennett, K.J.; Fransen, J.; Smith, M.R. Same or different? A comparison of anthropometry, physical fitness and perceptual motor characteristics in male and female youth soccer players. Sci. Med. Footb. 2020, 4, 37–44. [Google Scholar] [CrossRef]
- Cheuvront, S.N.; Carter, R.; DeRuisseau, K.C.; Moffatt, R.J. Running performance differences between men and women. Sport. Med. 2005, 35, 1017–1024. [Google Scholar] [CrossRef]
- Fairweather, H.; Hutt, S.J. Sex differences in a perceptual-motor skill in children. In Gender differences: Their ontogeny and significance; Ounsted, C., Taylor, D.C., Eds.; Churchill Livingstone: London, UK, 1972; pp. 159–175. [Google Scholar]
- Perri, R.L.; Berchicci, M.; Spinelli, D.; Di Russo, F. Individual differences in response speed and accuracy are associated to specifc brain activities of two interacting systems. Front. Behav. Neurosci. 2014, 8, 251. [Google Scholar] [CrossRef]
- Garrett, D.D.; Samanez-Larkin, G.R.; MacDonald, S.W.; Lindenberger, U.; McIntosh, A.R.; Grady, C.L. Moment-to-moment brain signal variability: A next frontier in human brain mapping? Neurosci. Biobehav. Rev. 2013, 37, 610–624. [Google Scholar] [CrossRef]
- Arsalidou, M.; Pascual-Leone, J.; Johnson, J.; Morris, D.; Taylor, M.J. A balancing act of the brain: Activations and deactivations driven by cognitive load. Brain Behav. 2013, 3, 273–285. [Google Scholar] [CrossRef]
- Lövdén, M.; Bäckman, L.; Lindenberger, U.; Schaefer, S.; Schmiedek, F. A theoretical framework for the study of adult cognitive plasticity. Psychol. Bull. 2010, 136, 659. [Google Scholar] [CrossRef]
- Hansen, A. The Real Happy Pill: Power Up Your Brain by Moving Your Body; Simon and Schuster: New York, NY, USA, 2017. [Google Scholar]
- McNamee, D.; Wolpert, D.M. Internal models in biological control. Annu. Rev. Control Robot. Auton. Syst. 2019, 2, 339. [Google Scholar] [CrossRef]
- Wolpert, D.M.; Ghahramani, Z.; Jordan, M.I. An internal model for sensorimotor integration. Science 1995, 269, 1880–1882. [Google Scholar] [CrossRef]
- Heidarnia, E.; Letafatkar, A.; Khaleghi-Tazji, M.; Grooms, D.R. Comparing the effect of a simulated defender and dual-task on lower limb coordination and variability during a side-cut in basketball players with and without anterior cruciate ligament injury. J. Biomech. 2022, 133, 110965. [Google Scholar] [CrossRef]
- Fortes, L.S.; Ferreira, M.E.; Faro, H.; Penna, E.M.; Almeida, S.S. Brain Stimulation Over the Motion-Sensitive Midtemporal Area Reduces Deleterious Effects of Mental Fatigue on Perceptual–Cognitive Skills in Basketball Players. J. Sport Exerc. Psychol. 2022, 44, 272–285. [Google Scholar] [CrossRef]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Effects | F(1,48) | p | ηp2 | Mean (SD) | Mean (SD) | |
|---|---|---|---|---|---|---|
| Single Change tests | Group | 8.6 | 0.005 | 0.152 | Exp = 7.18 (0.71) | Con = 7.68 (0.74) |
| Test | 114.5 | <0.001 | 0.704 | Pre = 7.83 (0.81) | Post = 7.03 (0.70) | |
| Sex | 0.3 | 0.560 | 0.007 | Mal = 7.48 (0.75) | Fem = 7.38 (0.74) | |
| Group × Test | 55.2 | <0.001 | 0.535 | |||
| Group × Sex | 0.3 | 0.568 | 0.007 | |||
| Test × Sex | 1.3 | 0.253 | 0.027 | |||
| Group × Treat × Sex | 2.3 | 0.134 | 0.046 | |||
| Multiple Change tests | Group | 13.4 | <0.001 | 0.218 | Exp = 9.78 (0.86) | Con = 10.36 (0.88) |
| Test | 126.1 | <0.001 | 0.724 | Pre = 10.49 (0.89) | Post = 9.65 (0.81) | |
| Sex | 36.4 | <0.001 | 0.431 | Mal = 9.59 (0.83) | Fem = 10.55 (0.88) | |
| Group × Test | 82.1 | <0.001 | 0.631 | |||
| Group × Sex | 1.1 | 0.294 | 0.023 | |||
| Test × Sex | 2.7 | 0.106 | 0.054 | |||
| Group × Test × Sex | 5.0 | 0.029 | 0.095 |
| Effects | F(1,48) | p | ηp2 | Mean (SD) | Mean (SD) | |
|---|---|---|---|---|---|---|
| RT | Group | 1.4 | 0.240 | 0.029 | Exp = 465 (63) | Con = 449 (60) |
| Test | 29.5 | <0.001 | 0.381 | Pre = 471 (65) | Post = 444 (60) | |
| Sex | 4.2 | 0.047 | 0.080 | Mal = 470 (66) | Fem = 447 (61) | |
| Group × Test | 5.9 | 0.018 | 0.110 | |||
| Group × Sex | 0.5 | 0.462 | 0.011 | |||
| Test × Sex | 0.5 | 0.491 | 0.010 | |||
| Group × Test × Sex | 4.5 | 0.040 | 0.085 | |||
| FA | Group | 0.3 | 0.574 | 0.007 | Exp = 5.57% (1.34) | Con = 5.97% (1.36) |
| Test | 105.8 | <0.001 | 0.688 | Pre = 8.51% (1.48) | Post = 3.02% (0.91) | |
| Sex | 2.4 | 0.128 | 0.048 | Mal = 6.32% (1.13) | Fem = 5.21% (1.14) | |
| Group × Test | 5.8 | 0.020 | 0.107 | |||
| Group × Sex | 1.7 | 0.202 | 0.034 | |||
| Test × Sex | 2.6 | 0.113 | 0.051 | |||
| Group × Test × Sex | 4.4 | 0.041 | 0.083 |
| Effects | F(1,48) | p | ηp2 | Mean (SD) | Mean (SD) | |
|---|---|---|---|---|---|---|
| pN | Group | 1.6 | 0.208 | 0.033 | Exp = −1.48 (0.26) | Con = −1.26 (0.22) |
| Test | 92.9 | <0.001 | 0.659 | Pre = −1.18 (0.21) | Post = −1.66 (0.28) | |
| Sex | 0.4 | 0.515 | 0.008 | Mal = −1.32 (0.25) | Fem = −1.43 (0.23) | |
| Group × Test | 24.9 | <0.001 | 0.341 | |||
| Group × Sex | 2.3 | 0.134 | 0.046 | |||
| Test × Sex | 0.1 | 0.816 | 0.001 | |||
| Group × Test × Sex | 9.3 | 0.004 | 0.162 | |||
| BP | Group | 0.8 | 0.368 | 0.017 | Exp = −2.89 (0.46) | Con = −2.65 (0.44) |
| Test | 32.0 | <0.001 | 0.400 | Pre = −2.47 (0.44) | Post = −3.07 (0.55) | |
| Sex | 0.4 | 0.502 | 0.009 | Mal = −2.86 (0.48) | Fem = −2.68 (0.47) | |
| Group × Test | 8.3 | 0.005 | 0.147 | |||
| Group × Sex | 0.5 | 0.459 | 0.011 | |||
| Test × Sex | >0.1 | 0.968 | 0.001 | |||
| Group × Test × Sex | 8.2 | 0.006 | 0.146 |
| Pre Minus Post | r | p |
|---|---|---|
| BP vs. Basket | 0.412 | 0.008 |
| pN vs. Basket | 0.389 | 0.018 |
| RT vs. Basket | 0.332 | 0.023 |
| FA vs. Basket | 0.319 | 0.031 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Lucia, S.; Aydin, M.; Di Russo, F. Sex Differences in Cognitive-Motor Dual-Task Training Effects and in Brain Processing of Semi-Elite Basketball Players. Brain Sci. 2023, 13, 443. https://doi.org/10.3390/brainsci13030443
Lucia S, Aydin M, Di Russo F. Sex Differences in Cognitive-Motor Dual-Task Training Effects and in Brain Processing of Semi-Elite Basketball Players. Brain Sciences. 2023; 13(3):443. https://doi.org/10.3390/brainsci13030443
Chicago/Turabian StyleLucia, Stefania, Merve Aydin, and Francesco Di Russo. 2023. "Sex Differences in Cognitive-Motor Dual-Task Training Effects and in Brain Processing of Semi-Elite Basketball Players" Brain Sciences 13, no. 3: 443. https://doi.org/10.3390/brainsci13030443
APA StyleLucia, S., Aydin, M., & Di Russo, F. (2023). Sex Differences in Cognitive-Motor Dual-Task Training Effects and in Brain Processing of Semi-Elite Basketball Players. Brain Sciences, 13(3), 443. https://doi.org/10.3390/brainsci13030443