
Anhedonia in Semantic Dementia—Exploring Right Hemispheric Contributions to the Loss of Pleasure

 , , , and
, , , and 
Abstract
:1. Introduction
2. Materials and Methods
2.1. Ethics Statement
2.2. Clinical and Cognitive Assessment
2.3. Assessment of Anhedonia
2.4. Assessment of Related Mood and Motivational Disturbances
2.5. Statistical Analysis
2.6. Image Acquisition
2.7. Voxel-Based Morphometry of Grey Matter Group Differences
2.8. Individual Differences in Atrophy of Hedonic Hotspots and SD Disease Epicentres
2.9. Grey Matter Contributions to Anhedonia in Neurodegenerative Dementia Syndromes
2.10. Clarifying Unique Regional Contributors to Anhedonia in SD
3. Results
3.1. Demographic, Clinical and Cognitive Variables
3.2. Neuropsychiatric and Behavioural Changes
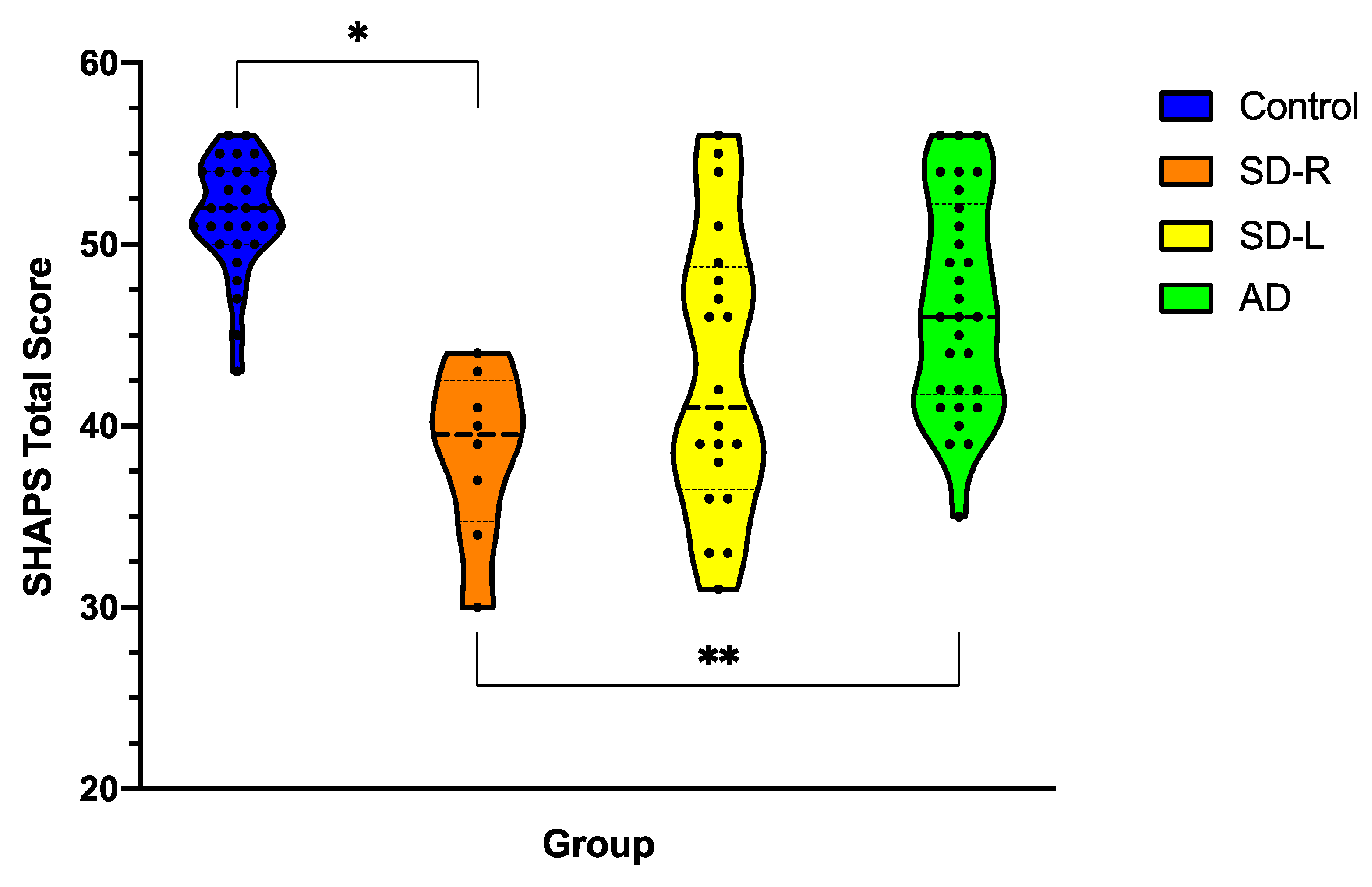
3.3. Anhedonia Severity across Dementia Syndromes
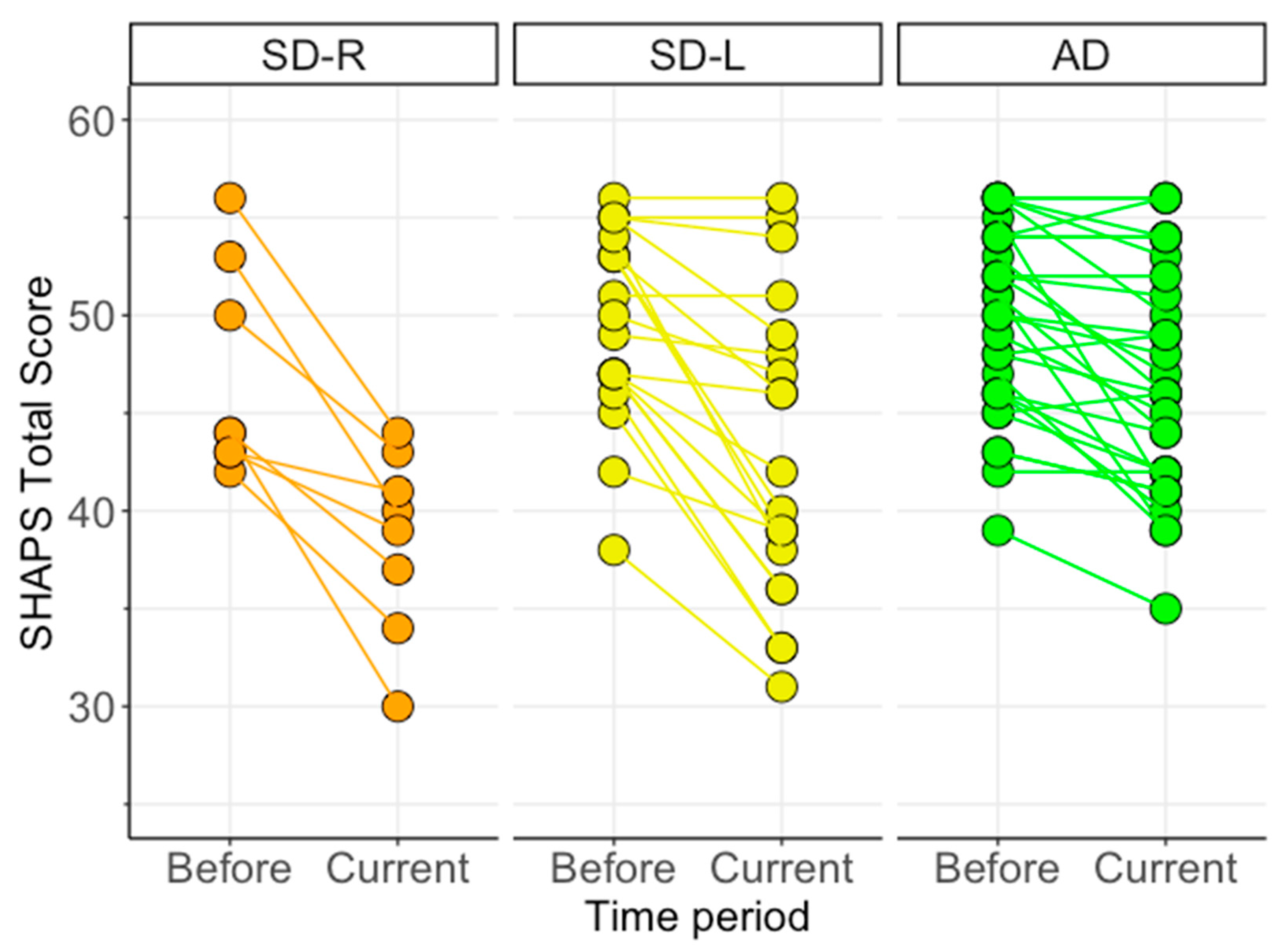
3.4. Changes in Anhedonia Following Dementia Onset
3.5. Associations between Anhedonia and Clinical Measures of Interest
3.6. Neuroimaging Results
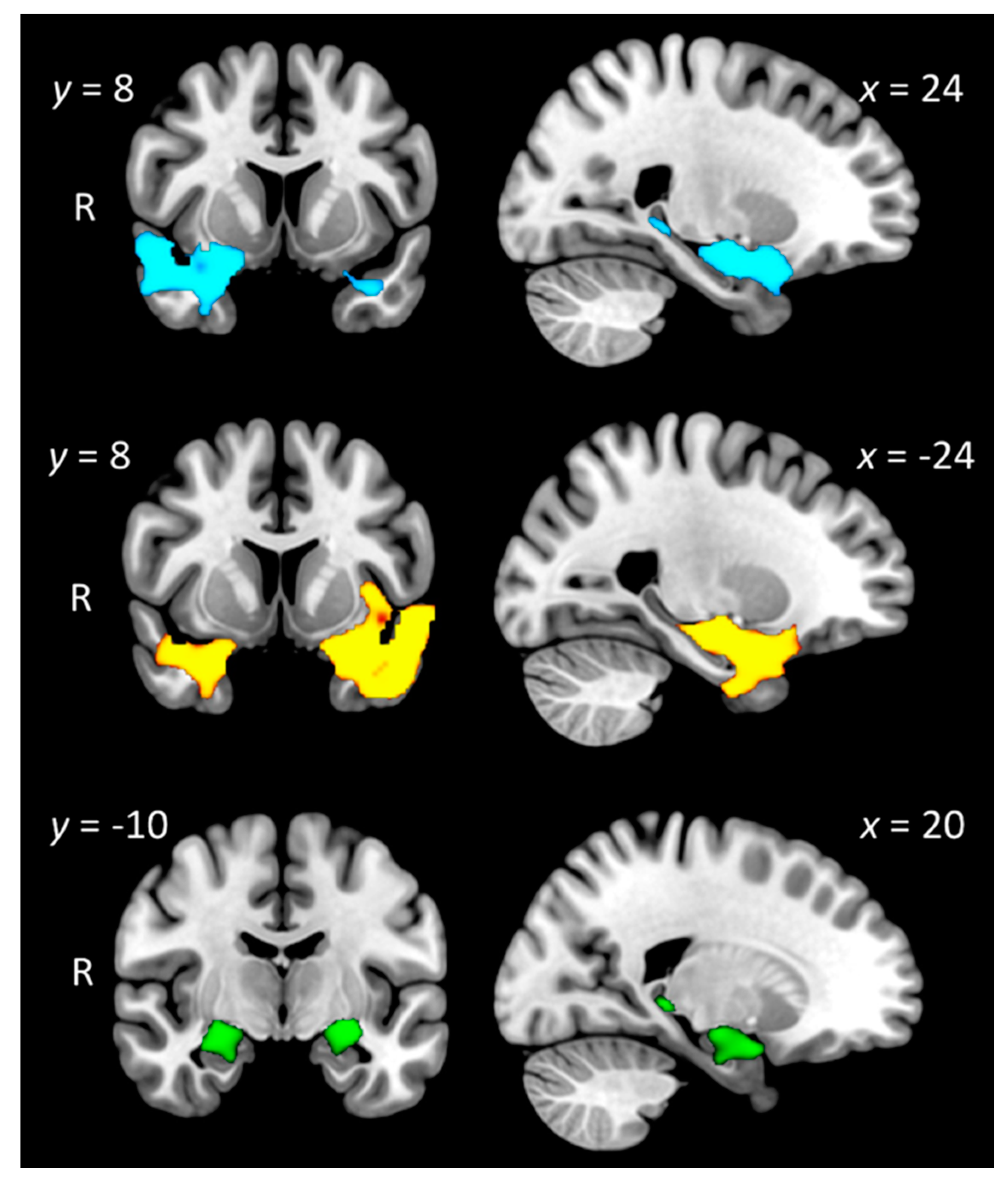
3.6.1. Group Differences in Grey Matter Intensity
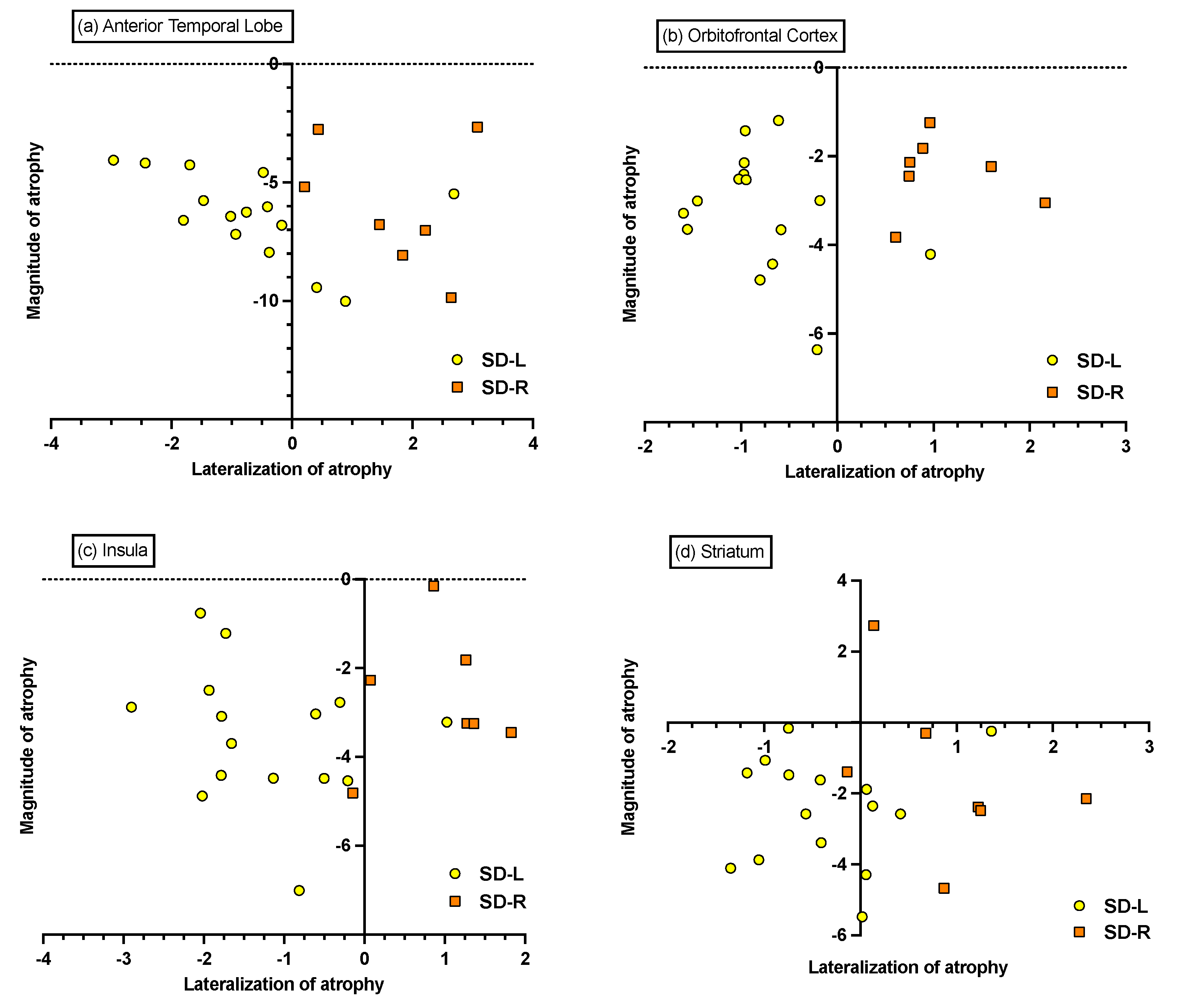
3.6.2. Individual Differences in Magnitude and Lateralisation of Atrophy
3.6.3. Associations between Grey Matter Intensity and Anhedonia on the SHAPS
3.6.4. A Unique Role for Right Temporal Regions in Anhedonia in SD
3.6.5. Mediation Analysis
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Ayano, G. Dopamine: Receptors, Functions, Synthesis, Pathways, Locations and Mental Disorders: Review of Literatures. J. Ment. Disord. Treat. 2016, 2, 1–4. [Google Scholar] [CrossRef]
- Nestler, E.J.; Carlezon, W.A. The Mesolimbic Dopamine Reward Circuit in Depression. Biol. Psychiatry 2006, 59, 1151–1159. [Google Scholar] [CrossRef]
- Hauser, T.U.; Eldar, E.; Dolan, R.J. Separate mesocortical and mesolimbic pathways encode effort and reward learning signals. Proc. Natl. Acad. Sci. USA 2017, 114, E7395–E7404. [Google Scholar] [CrossRef] [Green Version]
- Chau, D.T.; Roth, R.M.; Green, A.I. The neural circuitry of reward and its relevance to psychiatric disorders. Curr. Psychiatry Rep. 2004, 6, 391–399. [Google Scholar] [CrossRef] [PubMed]
- Bromberg-Martin, E.S.; Matsumoto, M.; Hikosaka, O. Dopamine in Motivational Control: Rewarding, Aversive, and Alerting. Neuron 2010, 68, 815–834. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Elliott, R.; Newman, J.L.; Longe, O.A.; Deakin, J. Instrumental responding for rewards is associated with enhanced neuronal response in subcortical reward systems. NeuroImage 2004, 21, 984–990. [Google Scholar] [CrossRef] [PubMed]
- Berridge, K.C.; Kringelbach, M. Pleasure Systems in the Brain. Neuron 2015, 86, 646–664. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Coenen, V.A.; Schlaepfer, T.E.; Maedler, B.; Panksepp, J. Cross-species affective functions of the medial forebrain bundle—Implications for the treatment of affective pain and depression in humans. Neurosci. Biobehav. Rev. 2011, 35, 1971–1981. [Google Scholar] [CrossRef] [PubMed]
- Husain, M.; Roiser, J.P. Neuroscience of apathy and anhedonia: A transdiagnostic approach. Nat. Rev. Neurosci. 2018, 19, 470–484. [Google Scholar] [CrossRef]
- Ahmed, R.M.; Devenney, E.M.; Irish, M.; Ittner, A.; Naismith, S.; Ittner, L.; Rohrer, J.; Halliday, G.; Eisen, A.; Hodges, J.R.; et al. Neuronal network disintegration: Common pathways linking neurodegenerative diseases. J. Neurol. Neurosurg. Psychiatry 2016, 87, 1234–1241. [Google Scholar] [CrossRef] [Green Version]
- Seeley, W.W.; Zhou, J.; Kim, E.J. Frontotemporal dementia: What can the behavioral variant teach us about human brain organization? Neuroscientist 2012, 18, 373–385. [Google Scholar] [CrossRef]
- Irish, M.; Piguet, O.; Hodges, J.R. Self-projection and the default network in frontotemporal dementia. Nat. Rev. Neurol. 2012, 8, 152–161. [Google Scholar] [CrossRef] [PubMed]
- Strikwerda-Brown, C.; Ramanan, S.; Irish, M. Neurocognitive mechanisms of theory of mind impairment in neurodegeneration: A transdiagnostic approach. Neuropsychiatr. Dis. Treat. 2019, 15, 557–573. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kumfor, F.; Irish, M.; Hodges, J.R.; Piguet, O. Discrete Neural Correlates for the Recognition of Negative Emotions: Insights from Frontotemporal Dementia. PLoS ONE 2013, 8, e67457. [Google Scholar] [CrossRef] [Green Version]
- Sturm, V.E.; Ascher, E.A.; Miller, B.L.; Levenson, R.W. Diminished self-conscious emotional responding in frontotemporal lobar degeneration patients. Emotion 2008, 8, 861–869. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dermody, N.; Wong, S.; Ahmed, R.; Piguet, O.; Hodges, J.R.; Irish, M. Uncovering the Neural Bases of Cognitive and Affective Empathy Deficits in Alzheimer’s Disease and the Behavioral-Variant of Frontotemporal Dementia. J. Alzheimers Dis. 2016, 53, 801–816. [Google Scholar] [CrossRef] [PubMed]
- Lough, S.; Kipps, C.; Treise, C.; Watson, P.; Blair, J.R.; Hodges, J.R. Social reasoning, emotion and empathy in frontotemporal dementia. Neuropsychology 2006, 44, 950–958. [Google Scholar] [CrossRef] [PubMed]
- Bora, E.M.; Walterfang, D.V. Theory of mind in behavioural-variant frontotemporal dementia and Alzheimer’s disease: A meta-analysis. J. Neurol. Neurosurg. Psychiatry 2015, 86, 714–719. [Google Scholar] [CrossRef]
- Synn, A.; Mothakunnel, A.; Kumfor, F.; Chen, Y.; Piguet, O.; Hodges, J.R.; Irish, M. Mental States in Moving Shapes: Distinct Cortical and Subcortical Contributions to Theory of Mind Impairments in Dementia. J. Alzheimers Dis. 2017, 61, 521–535. [Google Scholar] [CrossRef] [Green Version]
- Perry, D.C.; Sturm, V.E.; Seeley, W.W.; Miller, B.L.; Kramer, J.H.; Rosen, H.J. Anatomical correlates of reward-seeking behaviours in behavioural variant frontotemporal dementia. Brain 2014, 137, 1621–1626. [Google Scholar] [CrossRef] [Green Version]
- Mateus-Pinheiro, A.; Patrício, P.; Alves, N.D.; Machado-Santos, A.R.; Emorais, M.; Bessa, J.M.; Esousa, N.; Epinto, L. The Sweet Drive Test: Refining phenotypic characterization of anhedonic behavior in rodents. Front. Behav. Neurosci. 2014, 8, 74. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Shaw, S.R.; El-Omar, H.; Roquet, D.; Hodges, J.R.; Piguet, O.; Ahmed, R.M.; Whitton, A.E.; Irish, M. Uncovering the prevalence and neural substrates of anhedonia in frontotemporal dementia. Brain 2021, 144, 1551–1564. [Google Scholar] [CrossRef]
- Woolley, J.D.; Gorno-Tempini, M.-L.; Seeley, W.W.; Rankin, K.; Lee, S.S.; Matthews, B.R.; Miller, B.L. Binge eating is associated with right orbitofrontal-insular-striatal atrophy in frontotemporal dementia. Neurology 2007, 69, 1424–1433. [Google Scholar] [CrossRef] [PubMed]
- Whitwell, J.L.; Sampson, E.L.; Loy, C.T.; Warren, J.E.; Rossor, M.; Fox, N.; Warren, J. VBM signatures of abnormal eating behaviours in frontotemporal lobar degeneration. NeuroImage 2007, 35, 207–213. [Google Scholar] [CrossRef] [PubMed]
- Ahmed, R.M.; Irish, M.; Henning, E.; Dermody, N.; Bartley, L.; Kiernan, M.C.; Piguet, O.; Farooqi, S.; Hodges, J.R. Assessment of Eating Behavior Disturbance and Associated Neural Networks in Frontotemporal Dementia. JAMA Neurol. 2016, 73, 282–290. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ahmed, R.M.; Irish, M.; Piguet, O.; Halliday, G.; Ittner, L.; Farooqi, S.; Hodges, J.R.; Kiernan, M.C. Amyotrophic lateral sclerosis and frontotemporal dementia: Distinct and overlapping changes in eating behaviour and metabolism. Lancet Neurol. 2016, 15, 332–342. [Google Scholar] [CrossRef] [Green Version]
- Arsalidou, M.; Vijayarajah, S.; Sharaev, M. Basal ganglia lateralization in different types of reward. Brain Imaging Behav. 2020, 14, 1–29. [Google Scholar] [CrossRef]
- Zhang, S.; Hu, S.; Chao, H.H.; Li, C.-S.R. Hemispheric lateralization of resting-state functional connectivity of the ventral striatum: An exploratory study. Brain Struct. Funct. 2017, 222, 2573–2583. [Google Scholar] [CrossRef]
- Lopez-Persem, A.; Roumazeilles, L.; Folloni, D.; Marche, K.; Fouragnan, E.F.; Khalighinejad, N.; Rushworth, M.F.S.; Sallet, J. Differential functional connectivity underlying asymmetric reward-related activity in human and nonhuman primates. Proc. Natl. Acad. Sci. USA 2020, 117, 28452–28462. [Google Scholar] [CrossRef]
- Peet, B.T.; Castro-Suarez, S.; Miller, B.L. The Neuropsychiatric Features of Behavioral Variant Frontotemporal Dementia. In Advances in Experimental Medicine and Biology; Springer Science and Business Media LLC: Berlin, Germany, 2021; Volume 1281, pp. 17–31. [Google Scholar]
- Hodges, J.R.; Patterson, K. Semantic dementia: A unique clinicopathological syndrome. Lancet Neurol. 2007, 6, 1004–1014. [Google Scholar] [CrossRef]
- Mion, M.; Patterson, K.; Acosta-Cabronero, J.; Pengas, G.; Izquierdo-Garcia, D.; Hong, Y.T.; Fryer, T.D.; Williams, G.B.; Hodges, J.R.; Nestor, P. What the left and right anterior fusiform gyri tell us about semantic memory. Brain 2010, 133, 3256–3268. [Google Scholar] [CrossRef] [Green Version]
- Kumfor, F.; Landin-Romero, R.; Devenney, E.; Hutchings, R.; Grasso, R.; Hodges, J.R.; Piguet, O. On the right side? A longitudinal study of left- versus right-lateralized semantic dementia. Brain 2016, 139, 986–998. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hsieh, S.; Irish, M.; Daveson, N.; Hodges, J.R.; Piguet, O. When one loses empathy: Its effect on carers of patients with dementia. J. Geriatr. Psychiatry Neurol. 2013, 26, 174–184. [Google Scholar] [CrossRef] [PubMed]
- Irish, M.; Kumfor, F.; Hodges, J.R.; Piguet, O. A tale of two hemispheres: Contrasting socioemotional dysfunction in right- versus left-lateralised semantic dementia. Dement. Neuropsychol. 2013, 7, 88–95. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Irish, M.; Hodges, J.R.; Piguet, O. Right anterior temporal lobe dysfunction underlies theory of mind impairments in semantic dementia. Brain 2014, 137, 1241–1253. [Google Scholar] [CrossRef] [Green Version]
- Bozeat, S.; Gregory, C.A.; Ralph, M.A.L.; Hodges, J.R. Which neuropsychiatric and behavioural features distinguish frontal and temporal variants of frontotemporal dementia from Alzheimer’s disease? J. Neurol. Neurosurg. Psychiatry 2000, 69, 178–186. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kamminga, J.; Kumfor, F.; Burrell, J.; Piguet, O.; Hodges, J.R.; Irish, M. Differentiating between right-lateralised semantic dementia and behavioural-variant frontotemporal dementia: An examination of clinical characteristics and emotion processing. J. Neurol. Neurosurg. Psychiatry 2015, 86, 1082–1088. [Google Scholar] [CrossRef]
- Fletcher, P.D.; Downey, L.E.; Golden, H.L.; Clark, C.; Slattery, C.F.; Paterson, R.W.; Schott, J.; Rohrer, J.; Rossor, M.; Warren, J.D. Auditory hedonic phenotypes in dementia: A behavioural and neuroanatomical analysis. Cortex 2015, 67, 95–105. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hecht, D. Depression and the hyperactive right-hemisphere. Neurosci. Res. 2010, 68, 77–87. [Google Scholar] [CrossRef]
- Gorno-Tempini, M.L.; Hillis, A.E.; Weintraub, S.; Kertesz, A.; Mendez, M.; Cappa, S.F.; Ogar, J.M.; Rohrer, J.D.; Black, S.; Boeve, B.F.; et al. Classification of primary progressive aphasia and its variants. Neurology 2011, 76, 1006–1014. [Google Scholar] [CrossRef] [Green Version]
- McKhann, G.M.; Knopman, D.S.; Chertkow, H.; Hyman, B.T.; Jack, C.R., Jr.; Kawas, C.H.; Klunk, W.E.; Koroshetz, W.J.; Manly, J.J.; Mayeux, R.; et al. The diagnosis of dementia due to Alzheimer’s disease: Recommendations from the National Institute on Aging-Alzheimer’s Association workgroups on diagnostic guidelines for Alzheimer’s disease. Alzheimers Dement. 2011, 7, 263–269. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hsieh, S.; Schubert, S.; Hoon, C.; Mioshi, E.; Hodges, J.R. Validation of the Addenbrooke’s Cognitive Examination III in frontotemporal dementia and Alzheimer’s disease. Dement. Geriatr. Cogn. Disord. 2013, 36, 242–250. [Google Scholar] [CrossRef] [PubMed]
- So, M.; Foxe, D.; Kumfor, F.; Murray, C.; Hsieh, S.; Savage, G.; Ahmed, R.M.; Burrell, J.; Hodges, J.R.; Irish, M.; et al. Addenbrooke’s Cognitive Examination III: Psychometric Characteristics and Relations to Functional Ability in Dementia. J. Int. Neuropsychol. Soc. 2018, 24, 854–863. [Google Scholar] [CrossRef]
- Morris, J.C. Clinical Dementia Rating: A Reliable and Valid Diagnostic and Staging Measure for Dementia of the Alzheimer Type. Int. Psychogeriatr. 1997, 9, 173–176. [Google Scholar] [CrossRef] [PubMed]
- Wedderburn, C.; Wear, H.; Brown, J.; Mason, S.J.; Barker, R.A.; Hodges, J.; Williams-Gray, C. The utility of the Cambridge Behavioural Inventory in neurodegenerative disease. J. Neurol. Neurosurg. Psychiatry 2008, 79, 500–503. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mioshi, E.; Hsieh, S.; Savage, S.; Hornberger, M.; Hodges, J.R. Clinical staging and disease progression in frontotemporal dementia. Neurology 2010, 74, 1591–1597. [Google Scholar] [CrossRef] [PubMed]
- Snaith, R.P.; Hamilton, M.; Morley, S.; Humayan, A.; Hargreaves, D.; Trigwell, P. A Scale for the Assessment of Hedonic Tone the Snaith–Hamilton Pleasure Scale. Br. J. Psychiatry 1995, 167, 99–103. [Google Scholar] [CrossRef] [PubMed]
- Mendez, M.F.; Shapira, J.S. Loss of Insight and Functional Neuroimaging in Frontotemporal Dementia. J. Neuropsychiatry Clin. Neurosci. 2005, 17, 413–416. [Google Scholar] [CrossRef]
- Lovibond, S.H.; Lovibond, P.F. Manual for the Depression Anxiety Stress Scales, 2nd ed.; Psychology Foundation: Sydney, Australia, 1995. [Google Scholar]
- Irish, M.; Piguet, O.; Hodges, J.R.; Hornberger, M. Common and unique grey matter correlates of episodic memory dysfunction in frontotemporal dementia and Alzheimer’s disease. Hum. Brain Mapp. 2014, 35, 1422–1435. [Google Scholar] [CrossRef]
- Smith, S.M. Fast robust automated brain extraction. Hum. Brain Mapp. 2002, 17, 143–155. [Google Scholar] [CrossRef]
- Zhang, Y.; Brady, M.; Smith, S. Segmentation of brain MR images through a hidden Markov random field model and the expectation-maximization algorithm. IEEE Trans. Med. Imaging 2001, 20, 45–57. [Google Scholar] [CrossRef] [PubMed]
- Andersson, J.L.R.; Jenkinson, M.; Smith, S. Non-Linear Optimisation FMRIB Technical Report TR07JA1; University of Oxford FMRIB Centre: Oxford UK, 2007. [Google Scholar]
- Andersson, J.L.R.; Jenkinson, M.; Smith, S. Non-Linear Registration, Aka Spatial Normalisation FMRIB Technical Report TR07JA2; University of Oxford FMRIB Centre: Oxford, UK, 2007. [Google Scholar]
- Hwang, Y.T.; Strikwerda-Brown, C.; El-Omar, H.; Ramanan, S.; Hodges, J.R.; Burrell, J.R.; Piguet, O.; Irish, M. More than words—Longitudinal linguistic changes in the works of a writer diagnosed with semantic dementia. Neurocase 2021, 1–10. [Google Scholar] [CrossRef] [PubMed]
- Ding, J.; Chen, K.; Liu, H.; Huang, L.; Chen, Y.; Lv, Y.; Yang, Q.; Guo, Q.; Han, Z.; Ralph, M.A.L. A unified neurocognitive model of semantics language social behaviour and face recognition in semantic dementia. Nat. Commun. 2020, 11, 2595. [Google Scholar] [CrossRef]
- Rosen, H.J.; Perry, R.J.; Murphy, J.; Kramer, J.H.; Mychack, P.; Schuff, N.; Weiner, M.; Levenson, R.W.; Miller, B.L. Emotion comprehension in the temporal variant of frontotemporal dementia. Brain 2002, 125, 2286–2295. [Google Scholar] [CrossRef]
- Keedwell, P.A.; Andrew, C.; Williams, S.; Brammer, M.J.; Phillips, M.L. The Neural Correlates of Anhedonia in Major Depressive Disorder. Biol. Psychiatry 2005, 58, 843–853. [Google Scholar] [CrossRef]
- Bennett, C.M.; Wolford, G.L.; Miller, M.B. The principled control of false positives in neuroimaging. Soc. Cogn. Affect. Neurosci. 2009, 4, 417–422. [Google Scholar] [CrossRef]
- Chapleau, M.; Aldebert, J.; Montembeault, M.; Brambati, S.M. Atrophy in Alzheimer’s Disease and Semantic Dementia: An ALE Meta-Analysis of Voxel-Based Morphometry Studies. J. Alzheimers Dis. 2016, 54, 941–955. [Google Scholar] [CrossRef] [PubMed]
- Shrout, P.E.; Bolger, N. Mediation in experimental and nonexperimental studies: New procedures and recommendations. Psychol. Methods 2002, 7, 422. [Google Scholar] [CrossRef] [PubMed]
- Hayes, A.F. Introduction to Mediation, Moderation, and Conditional Process Analysis: A Regression-Based Approach; Guilford Publications: New York, NY, USA, 2017; pp. 120–141. [Google Scholar]
- Der-Avakian, A.; Markou, A. The neurobiology of anhedonia and other reward-related deficits. Trends Neurosci. 2012, 35, 68–77. [Google Scholar] [CrossRef] [Green Version]
- Snowden, J.S.; Harris, J.M.; Thompson, J.C.; Kobylecki, C.; Jones, M.; Richardson, A.M.; Neary, D. Semantic dementia and the left and right temporal lobes. Cortex 2018, 107, 188–203. [Google Scholar] [CrossRef] [PubMed]
- Landin-Romero, R.; Tan, R.; Hodges, J.R.; Kumfor, F. An update on semantic dementia: Genetics, imaging, and pathology. Alzheimers Res. Ther. 2016, 8, 1–9. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wong, S.; Irish, M.; Husain, M.; Hodges, J.R.; Piguet, O.; Kumfor, F. Apathy and its impact on carer burden and psychological wellbeing in primary progressive aphasia. J. Neurol. Sci. 2020, 416, 117007. [Google Scholar] [CrossRef] [PubMed]
- Modirrousta, M.; Price, B.H.; Dickerson, B.C. Neuropsychiatric symptoms in primary progressive aphasia: Phenomenology, pathophysiology, and approach to assessment and treatment. Neurodegener. Dis. Manag. 2013, 3, 133–146. [Google Scholar] [CrossRef] [Green Version]
- Gainotti, G. Emotional Behavior and Hemispheric Side of the Lesion. Cortex 1972, 8, 41–55. [Google Scholar] [CrossRef]
- Gainotti, G. The Role of the Right Hemisphere in Emotional and Behavioral Disorders of Patients With Frontotemporal Lobar Degeneration: An Updated Review. Front. Aging Neurosci. 2019, 11, 55. [Google Scholar] [CrossRef] [Green Version]
- Rolls, E.T. The orbitofrontal cortex and emotion in health and disease, including depression. Neuropsychology 2019, 128, 14–43. [Google Scholar] [CrossRef] [Green Version]
- Kumfor, F.; Zhen, A.; Hodges, J.R.; Piguet, O.; Irish, M. Apathy in Alzheimer’s disease and frontotemporal dementia: Distinct clinical profiles and neural correlates. Cortex 2018, 103, 350–359. [Google Scholar] [CrossRef]
- Fellows, L.K. Orbitofrontal contributions to value-based decision making: Evidence from humans with frontal lobe damage. Ann. N. Y. Acad. Sci. USA 2011, 1239, 51–58. [Google Scholar] [CrossRef]
- Kumfor, F.; Irish, M.; Hodges, J.R.; Piguet, O. The orbitofrontal cortex is involved in emotional enhancement of memory: Evidence from the dementias. Brain 2013, 136, 2992–3003. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bertossi, E.; Ciaramelli, E. Ventromedial prefrontal damage reduces mind-wandering and biases its temporal focus. Soc. Cogn. Affect. Neurosci. 2016, 11, 1783–1791. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Irish, M.; Addis, D.R.; Hodges, J.R.; Piguet, O. Considering the role of semantic memory in episodic future thinking: Evidence from semantic dementia. Brain 2012, 135, 2178–2191. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Irish, M.; Addis, D.R.; Hodges, J.R.; Piguet, O. Exploring the content and quality of episodic future simulations in semantic dementia. Neuropsychology 2012, 50, 3488–3495. [Google Scholar] [CrossRef] [PubMed]
- Hsiao, J.J.; Kaiser, N.; Fong, S.S.; Mendez, M.F. Suicidal Behavior and Loss of the Future Self in Semantic Dementia. Cogn. Behav. Neurol. 2013, 26, 85–92. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bush, G.; Luu, P.; Posner, M.I. Cognitive and emotional influences in anterior cingulate cortex. Trends Cogn. Sci. 2000, 4, 215–222. [Google Scholar] [CrossRef]
- Bush, G.; Vogt, B.A.; Holmes, J.; Dale, A.M.; Greve, D.; Jenike, M.A.; Rosen, B.R. Dorsal anterior cingulate cortex: A role in reward-based decision making. Proc. Natl. Acad. Sci. USA 2002, 99, 523–528. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhang, W.-N.; Chang, S.-H.; Guo, L.-Y.; Zhang, K.-L.; Wang, J. The neural correlates of reward-related processing in major depressive disorder: A meta-analysis of functional magnetic resonance imaging studies. J. Affect. Disord. 2013, 151, 531–539. [Google Scholar] [CrossRef] [PubMed]
- Craig, A.D. (Bud) How do you feel now? The anterior insula and human awareness. Nat. Rev. Neurosci. 2009, 10, 59–70. [Google Scholar] [CrossRef]
- Menon, V.; Uddin, L.Q. Saliency, switching, attention and control: A network model of insula function. Brain Struct. Funct. 2010, 214, 655–667. [Google Scholar] [CrossRef] [Green Version]
- Uddin, L. Salience processing and insular cortical function and dysfunction. Nat. Rev. Neurosci. 2015, 16, 55–61. [Google Scholar] [CrossRef]
- Uddin, L.Q.; Nomi, J.S.; Hébert-Seropian, B.; Ghaziri, J.; Boucher, O. Structure and Function of the Human Insula. J. Clin. Neurophysiol. 2017, 34, 300–306. [Google Scholar] [CrossRef]
- Strikwerda-Brown, C.; Ramanan, S.; Goldberg, Z.-L.; Mothakunnel, A.; Hodges, J.R.; Ahmed, R.M.; Piguet, O.; Irish, M. The interplay of emotional and social conceptual processes during moral reasoning in frontotemporal dementia. Brain 2021, 144, 938–952. [Google Scholar] [CrossRef] [PubMed]
- Lam, B.Y.K.; Halliday, G.; Irish, M.; Hodges, J.R.; Piguet, O. Longitudinal white matter changes in frontotemporal dementia subtypes. Hum. Brain Mapp. 2014, 35, 3547–3557. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nakonezny, P.A.; Carmody, T.J.; Morris, D.W.; Kurian, B.T.; Trivedi, M.H. Psychometric evaluation of the Snaith–Hamilton pleasure scale in adult outpatients with major depressive disorder. Int. Clin. Psychopharmacol. 2010, 25, 328–333. [Google Scholar] [CrossRef]
- Erkoyun, H.U.; Groot, C.; Heilbron, R.; Nelissen, A.; Van Rossum, J.; Jutten, R.; Koene, T.; Van Der Flier, W.M.; Wattjes, M.P.; Scheltens, P.; et al. A clinical-radiological framework of the right temporal variant of frontotemporal dementia. Brain 2020, 143, 2831–2843. [Google Scholar] [CrossRef] [PubMed]

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
|
SD-R
(n = 8) |
SD-L
(n = 20) |
AD
(n = 30) |
Control
(n = 30) | Group Effect | Post Hoc Comparisons | |
|---|---|---|---|---|---|---|
| Age, years | 64.9 (8.1) | 67.8 (7.4) | 66.2 (8.2) | 64.8 (7.1) | F = 0.64 n/s | |
| Education, years | 12.6 (3.3) | 13.5 (3.2) | 12.8 (2.8) | 14.6 (3.4) | F = 1.78 n/s | |
| Sex, M:F | 3:5 | 9:11 | 16:14 | 15:15 | χ = 0.79 n/s | |
| Disease duration, years | 5.7 (1.8) | 4.6 (2.3) | 4.8 (1.7) | -- | F = 0.86 n/s | |
| ACE-III Total (100) | 66.9 (17.7) | 67.2 (12.5) | 71.7 (7.8) | 95.3 (3.3) | F = 50.5 *** | Controls > SD-L, SD-R, AD |
| FRS Rasch a | −0.1 (1.1) | 1.2 (1.7) | 0.6 (1.3) | -- | F = 2.64 n/s | |
| CBI-R Total (%) | 35.6 (15.2) | 22.2 (12.5) | 27.4 (15.1) | -- | F = 2.36 n/s | |
| CBI-Motivation | 46.4 (31.7) | 32.9 (26.6) | 35.0 (27.3) | -- | F = 0.63 n/s | |
| DASS-Depression (21) | 6.3 (5.8) | 3.9 (3.3) | 3.7 (3.4) | 1.5 (1.8) | F = 5.57 ** | SD-R > Controls |
| SD Combined (n = 28) | Alzheimer’s Disease (n = 30) | |
|---|---|---|
| Disease duration | −0.32 | −0.15 |
| FRS Rasch Score a | 0.79 *** | 0.46 * |
| ACE-III Total | 0.33 | 0.11 |
| DASS-Depression | −0.27 | −0.45 * |
| CBI-R Motivation | −0.78 *** | −0.39 * |
| CBI-R Total | −0.70 *** | −0.51 ** |
| ZBI | −0.60 ** | −0.25 |
| Contrast | Regions | Side | Cluster Size | Peak MNI Coordinates | t Value | ||
|---|---|---|---|---|---|---|---|
| x | y | z | |||||
| Controls—SD-R | Temporal pole, orbitofrontal cortex, anterior temporal fusiform cortex, anterior superior/middle temporal gyrus, insular cortex, amygdala, putamen, pallidum, hippocampus, parahippocampal gyrus | R | 2954 | 34 | 14 | −36 | 3.97 |
| Temporal pole | L | 136 | −36 | 8 | −30 | 3.01 | |
| Hippocampus, amygdala | L | 60 | −28 | −6 | −24 | 3.46 | |
| Hippocampus, thalamus | R | 58 | 26 | −34 | −6 | 2.59 | |
| Controls—SD-L | Anterior and posterior temporal fusiform cortex, anterior and posterior parahippocampal gyrus, hippocampus, amygdala, temporal occipital fusiform cortex, temporal pole, orbitofrontal cortex, insular cortex, putamen, superior/middle temporal gyrus | L | 5615 | −30 | −8 | −44 | 3.87 |
| Temporal pole, orbitofrontal cortex, anterior parahippocampal gyrus, anterior and posterior temporal fusiform cortex, amygdala, hippocampus | R | 1793 | 38 | 16 | −36 | 3.87 | |
| Controls—AD | Anterior parahippocampal gyrus, temporal pole, amygdala, hippocampus | R | 387 | 24 | 2 | −20 | 3.58 |
| Amygdala, hippocampus, temporal pole, anterior parahippocampal gyrus, orbitofrontal cortex, putamen, pallidum | L | 347 | −26 | −12 | −14 | 3.58 | |
| Angular gyrus, superior parietal lobule, supramarginal gyrus | R | 67 | 23 | 37 | 55 | 3.35 | |
| Thalamus, hippocampus | R | 65 | 18 | −34 | 0 | 3.58 | |
| SD-L—SD-R | No significant clusters | ||||||
| SD-R—SD-L | No significant clusters | ||||||
| Contrast | Regions | Side | Cluster Size | Peak MNI Coordinates | t Value | ||
|---|---|---|---|---|---|---|---|
| x | y | z | |||||
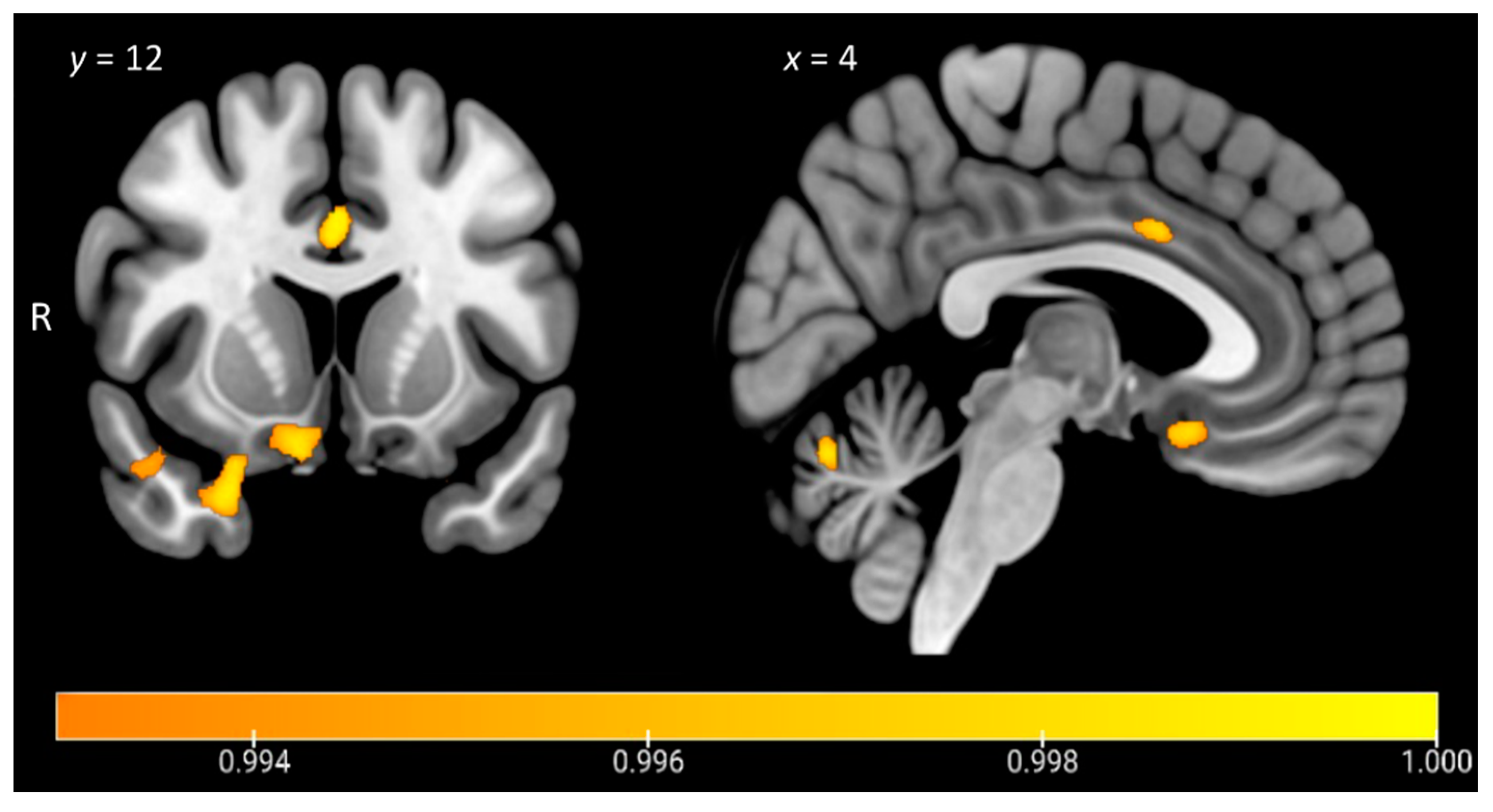
| SHAPS | Orbitofrontal cortex, temporal pole, subcallosal cortex | R | 351 | 18 | 32 | −22 | 3.62 |
| Temporal pole, orbitofrontal cortex | R | 272 | 24 | 10 | −28 | 3.48 | |
| Temporal occipital fusiform cortex, posterior temporal fusiform cortex, posterior parahippocampal gyrus, lingual gyrus | R | 269 | 34 | −46 | −12 | 3.48 | |
| Cerebellum | B | 199 | −6 | −68 | −22 | 3.86 | |
| Posterior temporal fusiform cortex, posterior parahippocampal gyrus | L | 134 | −36 | −42 | −10 | 3.18 | |
| Anterior cingulate gyrus, left paracingulate gyrus | B | 102 | 0 | 12 | 34 | 3.86 | |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Shaw, S.R.; El-Omar, H.; Ramanan, S.; Piguet, O.; Ahmed, R.M.; Whitton, A.E.; Irish, M. Anhedonia in Semantic Dementia—Exploring Right Hemispheric Contributions to the Loss of Pleasure. Brain Sci. 2021, 11, 998. https://doi.org/10.3390/brainsci11080998
Shaw SR, El-Omar H, Ramanan S, Piguet O, Ahmed RM, Whitton AE, Irish M. Anhedonia in Semantic Dementia—Exploring Right Hemispheric Contributions to the Loss of Pleasure. Brain Sciences. 2021; 11(8):998. https://doi.org/10.3390/brainsci11080998
Chicago/Turabian StyleShaw, Siobhán R., Hashim El-Omar, Siddharth Ramanan, Olivier Piguet, Rebekah M. Ahmed, Alexis E. Whitton, and Muireann Irish. 2021. "Anhedonia in Semantic Dementia—Exploring Right Hemispheric Contributions to the Loss of Pleasure" Brain Sciences 11, no. 8: 998. https://doi.org/10.3390/brainsci11080998
APA StyleShaw, S. R., El-Omar, H., Ramanan, S., Piguet, O., Ahmed, R. M., Whitton, A. E., & Irish, M. (2021). Anhedonia in Semantic Dementia—Exploring Right Hemispheric Contributions to the Loss of Pleasure. Brain Sciences, 11(8), 998. https://doi.org/10.3390/brainsci11080998