1. Introduction
Injuries are inevitable for most organisms, yet maintaining a satisfactory level of functionality can be decisive for their survival. The progressive wear of a honeybee’s wings, for example, challenges the insect to sustain its load lift or face less nourishing foraging trips [
1,
2]. Functional robustness is desirable for neural systems as well. For example, while computer devices operate in a regime of near-zero tolerance for physical damage, the middle-aged human brain undergoes significant neuronal losses on a daily basis [
3]. Robustness to injury is often overlooked when analyzing the purpose and function of neural structures while the transmission of maximum information, high signal-to-noise ratio, and low energy consumption are primarily considered [
4]. Analyzing neural information processing in the context of these principles is certainly important, but arguably incomplete. The goal of this work is to examine whether certain neural mechanisms and architectural structures can be understood as adaptive, built-in systems for robustness to brain injury from trauma, aging, and/or other disorders. That is, we examine how biological neural systems are “built to last”. In particular, we explore how certain neural architectures can protect the system’s key downstream outputs (the “deliverables” of the system) from the effects of damage to upstream regions.
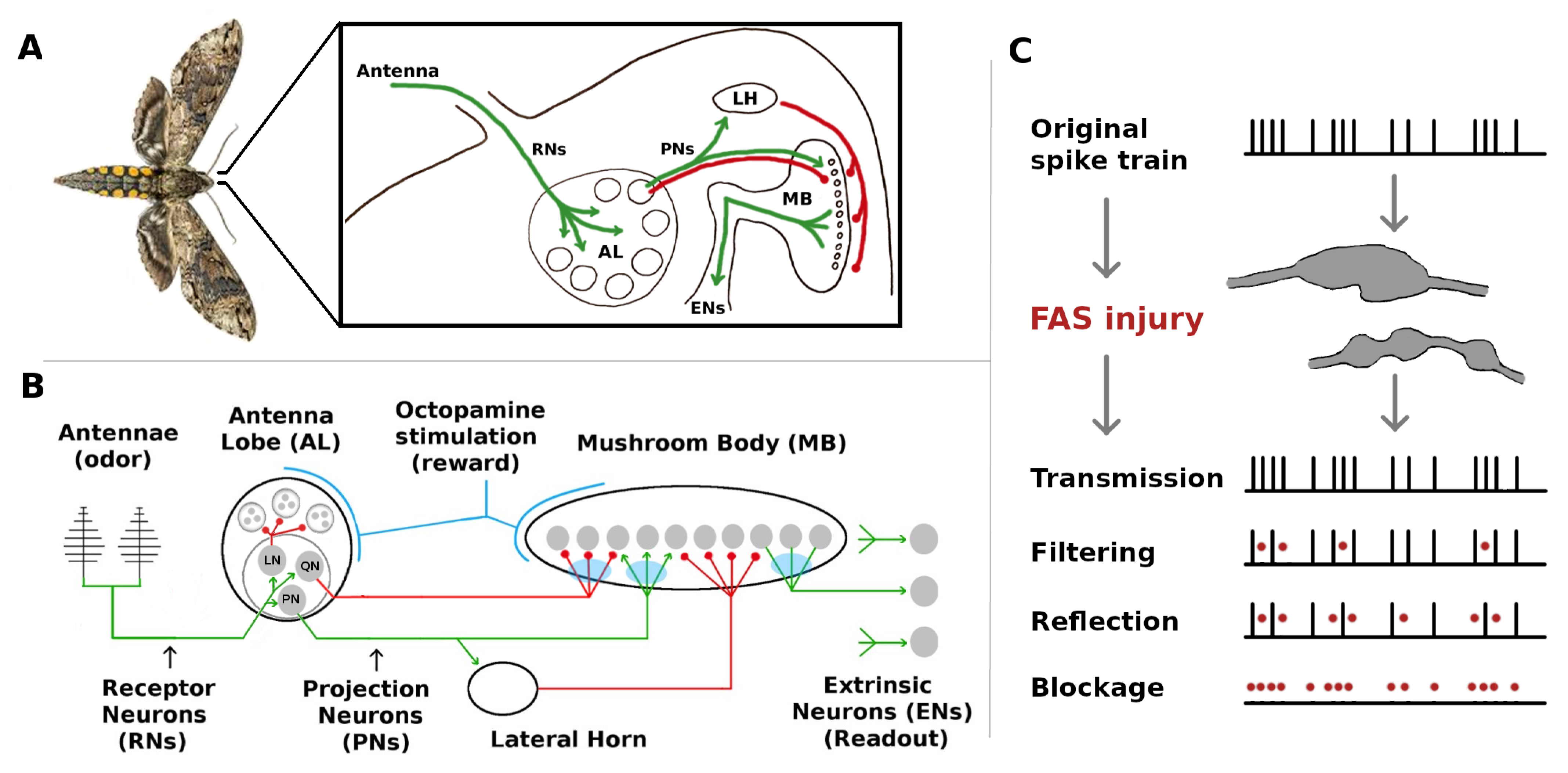
The olfactory system of the
Manduca sexta moth, though simple, shares many neural structures and mechanisms with higher organisms [
5,
6]. These include (i) Hebbian plasticity, (ii) reward-triggered stimulation of neural outputs via neuromodulators, (iii) high noise levels, and (iv) inhibitory feed-forward channels running parallel to excitatory channels. It is thus an ideal model organism to investigate the injury mitigation effects of these elements. MothNet is a computational model of this olfactory network which incorporates known biophysical parameters and which was calibrated to firing rate data recorded during in vivo learning tasks [
7]. MothNet models the food odor-responsive part of the moth’s olfactory network, not the sex pheromone-responsive macroglomerular complex, since the MGC has substantially distinct targets and dynamics (despite small overlaps [
8]). Also, MothNet models learning-induced Hebbian updates in the synaptic connections of existing neurons, not neurogenesis via for example Juvenile Hormone [
9]. See
Figure 1 for a system schematic.
The moth olfactory network (MON) also contains well-defined readout neurons (ENs, for Extrinsic Neurons), which are downstream outputs that deliver key actionable encodings to the rest of its body [
12,
13]. From a functional viewpoint, internal damage is unimportant as long as the key outputs (readouts) of the system are preserved. Thus, to examine injury mitigation effects we ran in silico simulations of neural injury on the MothNet model, and measured how the firing rates (FRs) of readout ENs were affected by injuries and by injury-mitigation mechanisms.
Maia et al. [
14] recently introduced a computational model for the cellular level effects that may distort firing rates due to major forms of neuronal injury. They posit phenomenological input/output rules to transform healthy neuronal spike train responses into injured ones, with filters that can be either discrete-time (for spike trains) or continuous-time (for firing rates) signal processors. These filters were derived modeling the effects of demyelination and Focal Axonal Swellings (FAS), which are present in a broad array of neurological disorders [
15,
16,
17,
18].
Figure 1c exemplifies how a FAS-like injury distorts the propagation of spike trains along the axon, effectively blocking or filtering signals encoded to downstream neurons. In this work, we are agnostic concerning the exact biological underpinnings and pathological mechanisms that may affect an injured/aging moth. Instead, we simply posit that its neurons might be exposed to detrimental effects that can affect their signaling capabilities. In this sense, and as explained in Maia et al. [
14], FAS-based filters provide a more nuanced way to model neuronal malfunction than purely binary ablation which treats a neuron and/or its connections as either fully functional or 100% impaired. Recent computational studies that consider the effects of FAS-like injury in neural networks are providing new insight to decision-making deficits [
19], learning impairments [
20,
21], memory deterioration [
22], and motor-function decline [
23].
While FAS models effects at the level of spike trains, it also has a meaningful representation in Firing Rate models such as MothNet. In particular, unlike ablation FAS causes reduced but still non-zero FRs. In addition, the low-pass filtering effect of FAS, which impacts closely-bunched clusters of spikes more than sparse spikes, in analogous manner impacts high FRs more strongly than low FRs. Thus FAS, applied in a FR model, results in neuron FRs being reduced but not ablated, with high FR neurons affected more strongly than low FR neurons. For a fixed amount of total damage, FAS results in relatively many partially-damaged neurons, while ablation results in relatively fewer fully-destroyed neurons.
In our simulations, we varied the parameters of each network structure-under-test; applied FAS-type injuries to different subnetworks of the system (simulating the outcome of a traumatic brain injury or concussion); and assessed the net effects on EN outputs. We examined two aspects of FR behavior for a single representative EN: (i) changes in raw FR, and (ii) changes in the ability of the EN to discriminate between a trained and untrained odor.
Our experiments led to four main findings concerning injury-mitigation structures in the moth olfactory system:
The learning mechanism, based on the combination of octopamine stimulation and Hebbian growth, can restore both the magnitude and discriminative ability of downstream readout neurons after upstream neurons are injured.
The presence of inhibitory neurons parallel to excitatory neurons connecting the subnetworks can mitigate the effects of injury, possibly via a “canceling out” effect.
A broad noise envelope on neural firing rates protects EN responses from effects of upstream injury, by enabling the strongest neural signals delivered downstream to still exceed action-triggering thresholds.
Simple ablation injury in upstream regions produces downstream effects distinct from the more biologically plausible FAS injury. That is, ablation may be a poor proxy for naturalistic injury in some neural systems.
Concerning item 1 (learning as an injury mitigation mechanism), we note that while it is intuitive that Hebbian plasticity [
24,
25] might
help repair an injured network, it does not follow that Hebbian updates
alone can repair damage: Hebbian “fire together, wire together” updates are proportional to the FRs of both the incoming and the receiving neurons. Hebbian plasticity thus requires that the upstream neurons have sufficiently strong FRs, but precisely these FRs are reduced by injury. Our experiments indicate that octopamine-induced stimulation of the injured upstream neurons is crucial to post-injury plasticity, because it boosts the FRs of injured upstream neurons back to levels that enable non-trivial Hebbian updates. Without such stimulation, the FRs of the injured neurons, as well as the FRs induced by the injured neurons in downstream neurons, are too low to induce gains in synaptic strength via a Hebbian mechanism [
7].
Our computational approach allowed us to quantify the mitigating effect of a neural structure-under-test as a function of injury level, injury location, and structure parameters. We recognize that, as in any computational model, specific quantitative outcomes necessarily depend on the particular parameters and assumptions of the MothNet model. However, MothNet’s architecture is tethered to known biophysical findings, and its parameter values are calibrated to biophysical findings and in vivo FR data [
7]. We believe this approach enables our experimental results to refer back meaningfully to the biological structure.
2. Results
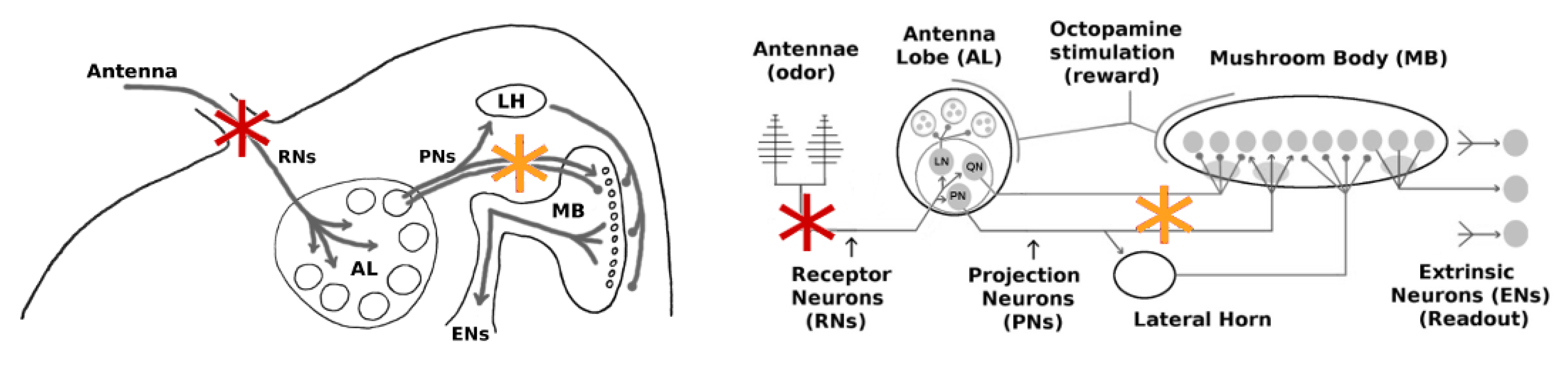
Throughout this work, we targeted two distinct regions with our injury protocols:
(i) the Antennae and
(ii) the channel between the Antennal Lobe (AL) and the Mushroom Body (MB). Their specific locations are shown in
Figure 2.
(i) The antennae comprise the outermost region of the olfactory system and are arguably the most exposed to external environmental shocks. Damage in this location should affect primarily the Receptor Neuron (RN) subpopulation (∼30,000). We note that the hundreds of RNs responsive to a given odor are spread throughout the antennae, ensuring that localized damage to an antenna does not disproportionately reduce the response to a particular odor.
(ii) The AL→MB channel is internal and is a center for signal transfer in the network. Damage in this location would affect both excitatory projection neurons (PNs) and inhibitory projection neurons (QNs) that link the AL to the MB.
2.1. Plasticity-Induced Recovery from Injury
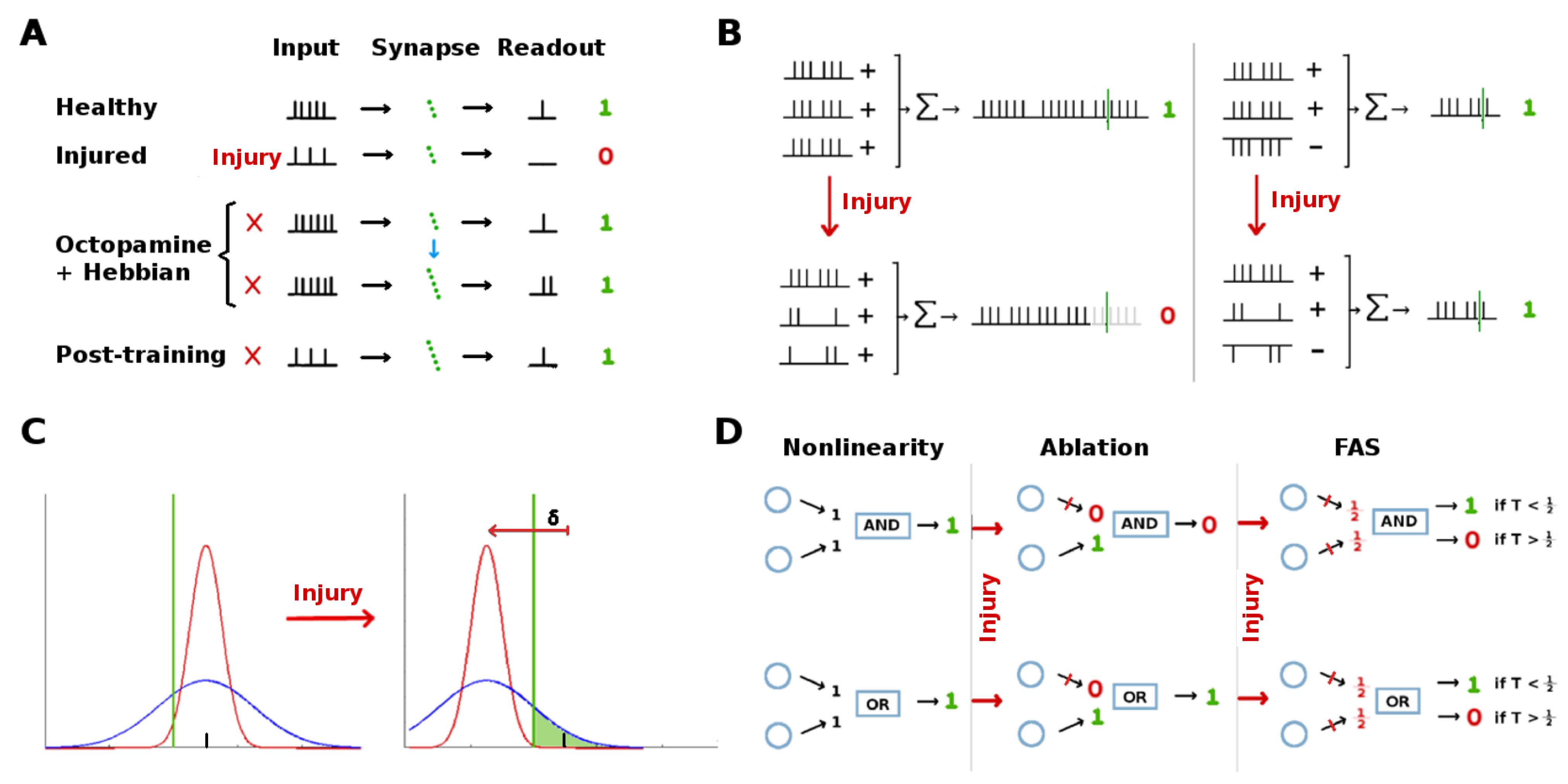
The Moth Olfactory Network (MON) contains some plastic synaptic connections, and it can learn [
26]: In response to reward (sugar at the proboscis), a large neuron releases octopamine widely in the AL and MB. Octopamine stimulation of FRs, combined with Hebbian-like synaptic updates, strengthens the plastic synaptic connections in the AL→MB and MB→EN channels, resulting in stronger EN responses to stimuli.
The goal of this set of experiments was to examine how far the Hebbian learning mechanism can compensate for neural injury, i.e., how far it can operate as a mitigation (as opposed to a learning) mechanism. In the first experiment, RNs in the Antennae→AL channel were injured (
Figure 2, red stars). In the second experiment, PNs in the AL→MB channel were targeted (
Figure 2 orange stars). AL noise was set to naturalistic levels (calibrated per in vivo data [
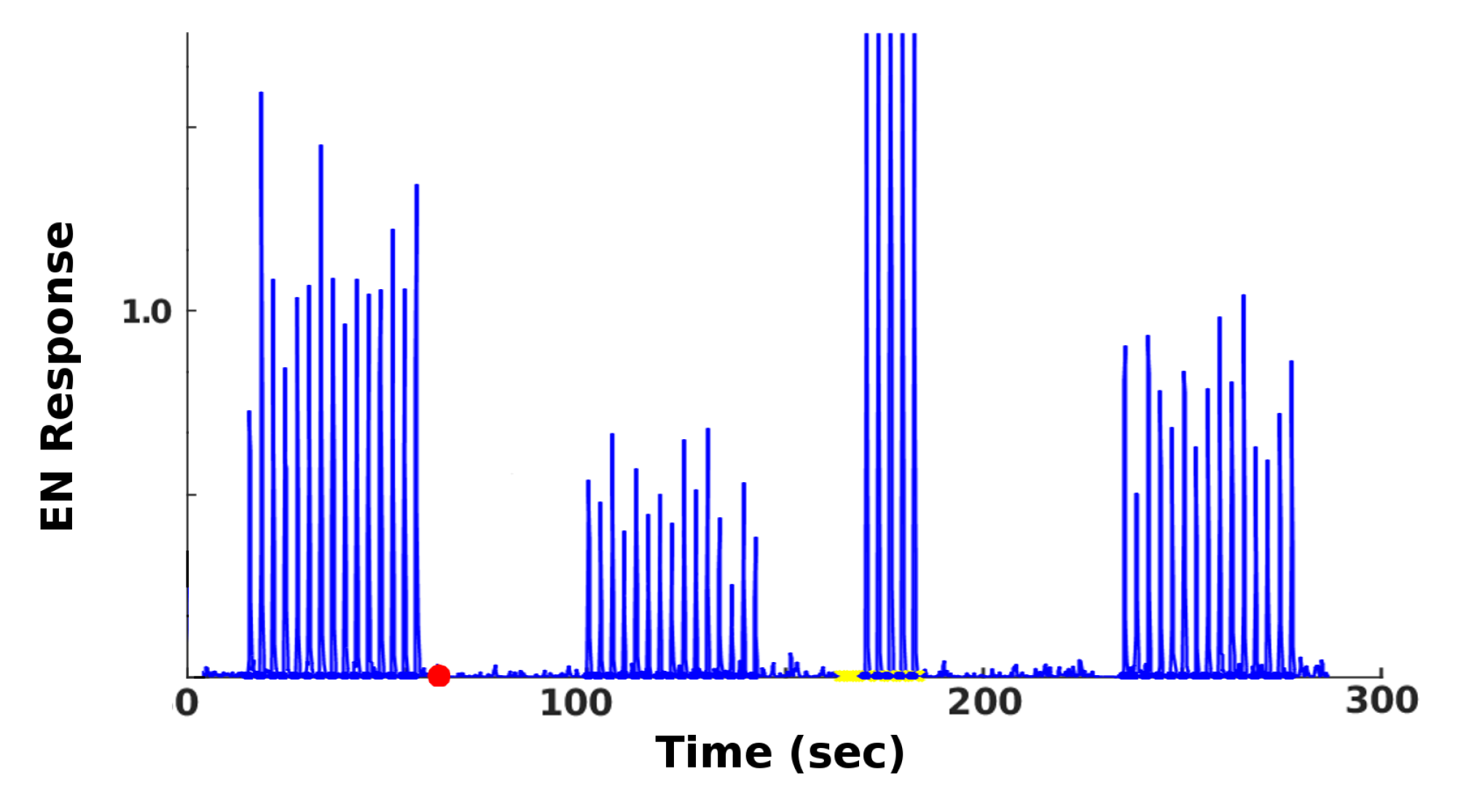
7]) and FAS-like injury levels ranged from 0% to 60%. The MON was subsequently retrained with 5 odor puffs, close to sufficient to max out the allowable synaptic weights. The average EN readout response was recorded, as a key measure of the actionable output of the system. A typical timecourse is shown in
Figure 3. In each experiment, over 30 (
n = 34–38, mean = 36) MothNet instances were generated from template (i.e., a specification of network parameters used to randomly generate MothNet instances) and tested at each injury level.
We examined two properties of the EN readout:
(i) Magnitude of EN output, a basic property relevant to triggering behavioral response to an odor. Examining effects on EN FR magnitude required no pre-training of the network prior to injury.
(ii) Discriminative ability between two odors (one trained and one control), measured as the Fisher linear discriminant
where
are the mean and std. dev. of EN responses to trained and control odors. In these simulations, prior to injury one odor was trained so the system could discriminate the trained odor vs. control odor (
). We either used two randomly-generated odor profiles with broad, overlapping projections onto the AL; or {odor + noise} vs. noise, with mean noise magnitude set between 0.2 and 1.0 times the odor magnitude (in MothNet, odor magnitude is controlled by a scalar which multiplies unit-length odor vectors before they are inputted to the RNs).
Results were as follows:
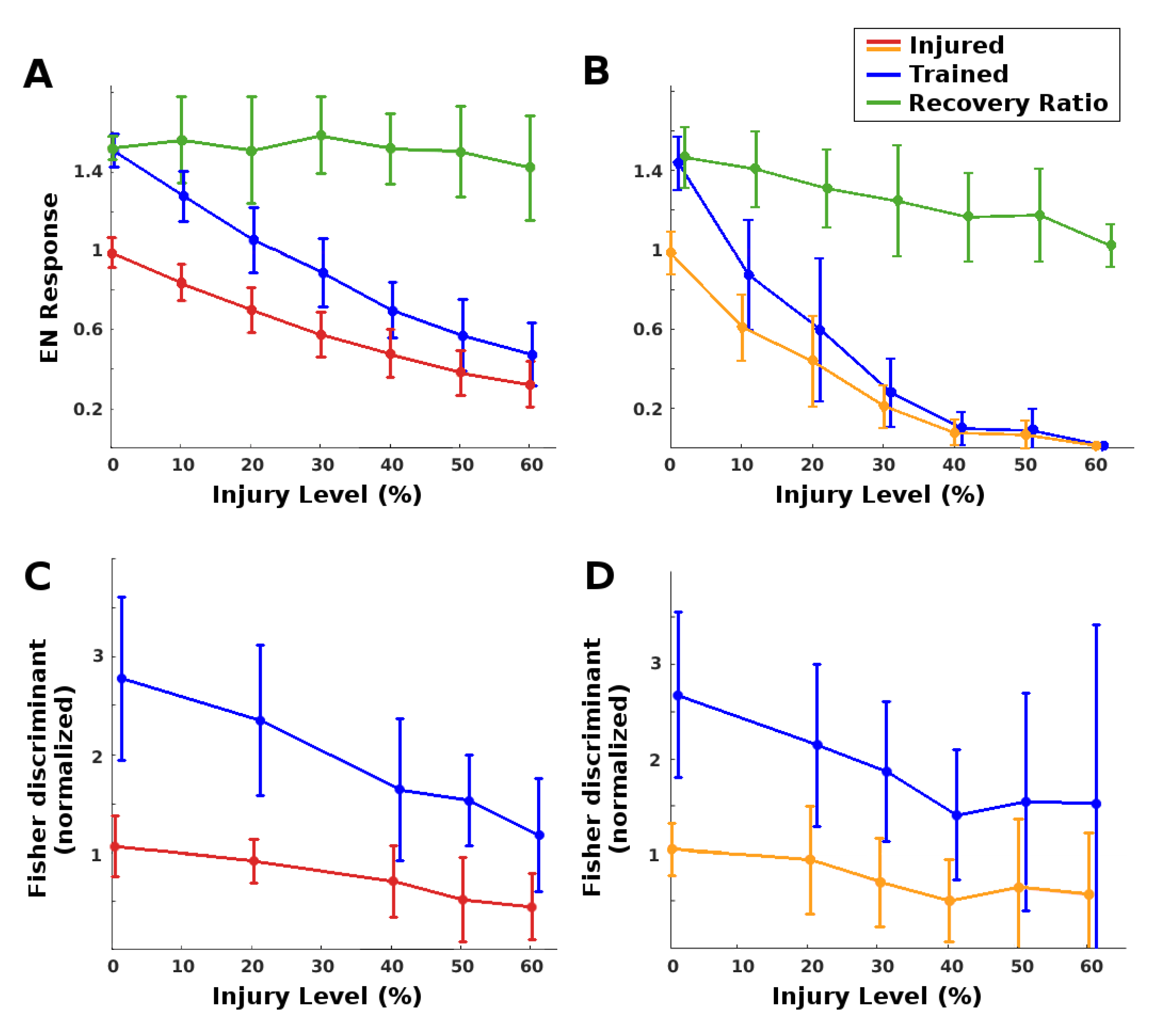
(i) EN magnitude: As expected, injury reduced raw EN response magnitude, and training restored some of this loss. The MON was much more robust to RN (antennae) damage than to PN damage. Complete restoration was achieved (on average) for injury levels below 25% for RN damage and below 8% for PN damage. At these injury values, injury reduced EN odor responses to approximately 70% of the naive baseline, and training restored them to baseline. See the plots in
Figure 4A,B. We note that at low injury levels, the system was able to boost EN output by about 140% to 150%, a value constrained by the model’s saturation parameter for the synaptic connection weights. At high levels of injury to PNs, however, the learning mechanism’s ability to recover EN performance decreased (see green curves in
Figure 4).
(ii) Discrimination: Injury affected EN discriminative ability much less than it did EN magnitude, because it impacted EN responses to both trained odor and control. Given two odors, injury reduced discrimination, while retraining readily restored all losses (Fisher discriminant plots in
Figure 4C,D. Between {odor + noise} vs noise, injury had no effect on discriminative ability at any noise level, likely because the sparsely-firing MB is an effective noise filter [
7]. Post-injury training served to further increase discrimination between {odor + noise} vs. noise (results not shown).
2.2. Inhibitory Neurons and Protective Canceling Out Effect
Each glomerulus in the AL has ≈5 excitatory PNs that feed forward to the MB. The moth also has a smaller number of inhibitory neurons (here called QNs) that also feed forward to the MB, analogous to and in parallel with the PNs. We note that these feed-forward QNs are one of three inhibitory networks in the AL-MB. The other two, viz. lateral inhibitory neurons within the AL [
27] and global sparsity-inducing inhibition onto the MB from the Lateral Horn [
28] (or global self-inhibition by the MB as in
drosophila [
29]), have different functions and are assumed to be non-plastic. Our experiments target the QNs, which innervate only a subset of MB neurons, and are presumed in MothNet to be plastic like PNs.
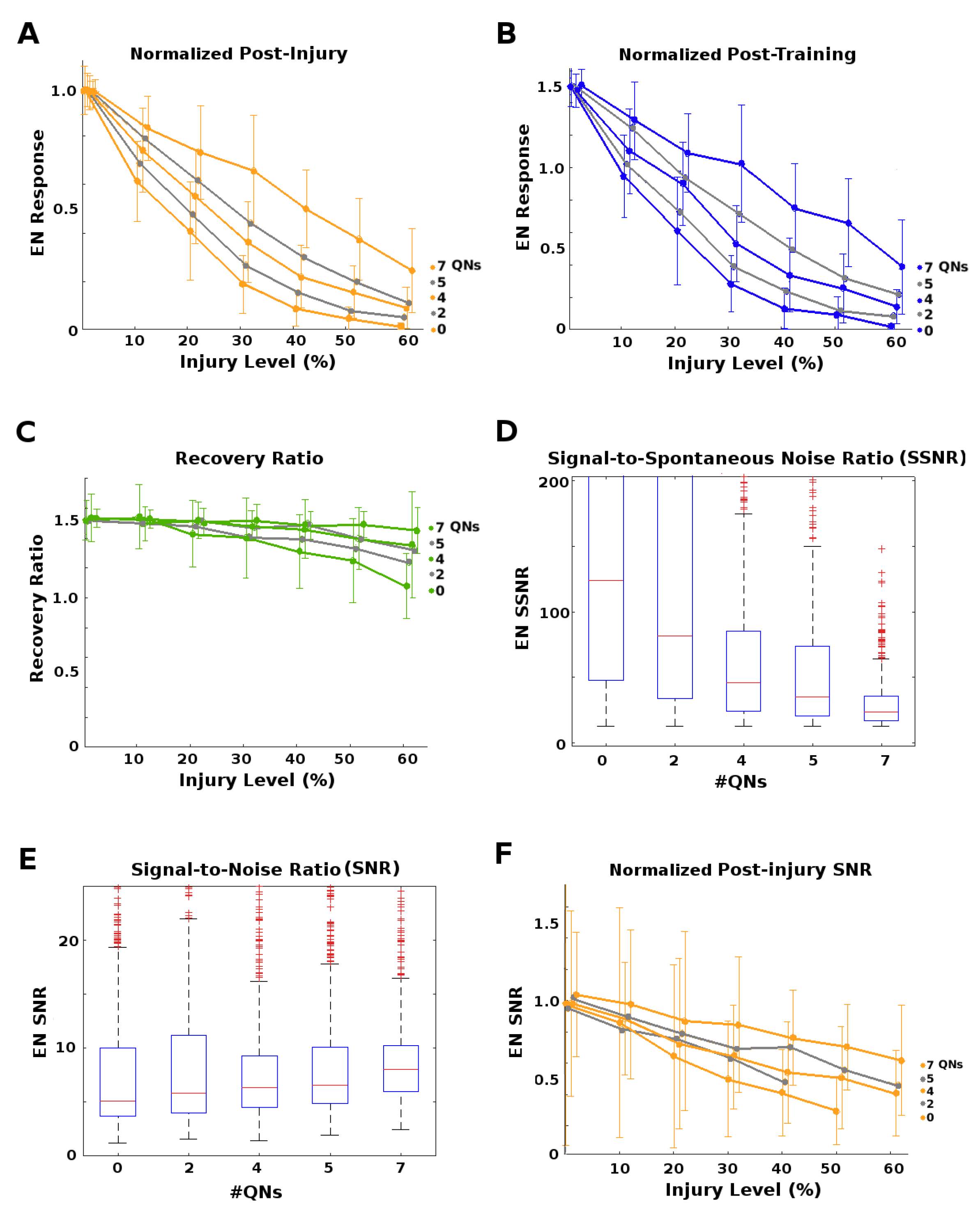
The goal of this set of experiments was to test whether the existence of QNs parallel to PNs might mitigate the effect of injuries applied to this region (orange stars in
Figure 2). We varied the QN:PN ratio (0, 2, 4, 5, and 7 QNs/glomerulus, for 5 PNs/glomerulus) while injuring the AL→MB channel. Each parameter pair (e.g., “4 QNs, 50% FAS injury”) had at least 30 moth instances (31 to 40, mean = 35). We found that higher numbers of QNs correlated strongly with reduced effects on EN output magnitudes from upstream injury, but had no clear effect on discriminative ability (results not shown). Results reported here are for effects on EN response magnitude.
Moths with high QN counts had stronger post-injury EN odor responses (
Figure 5A), and post-injury training sessions allowed them to fully recover from much higher levels of injury than moths with few or no QNs (≈8% injury for QNs = 0, ≈15% injury when QNs = 4, and ≈30% injury when QNs = 7;
Figure 5B,C). High QN counts had another, unexpected advantage regarding the Signal-to-Noise Ratio,
where
and
correspond to the mean and standard deviation, and
is the set of discrete EN responses (peak FR) to a series of odor puffs. Naive SNR values (i.e., pre-injury, pre-training) were similar for all QN counts (
Figure 5E). While post-injury SNR always dropped proportionally to the severity of the injury, high QN counts substantially reduced these losses to SNR (
Figure 5F).
However, high QNs counts also carried a downside. They had a much lower EN Signal-to-Spontaneous Noise Ratio,
where
F is defined as above and
is the spontaneous EN firing rate. The SSNR measures the clarity of the signal with respect to background noise, and their values for different QN counts are shown in
Figure 5D. Many moth instances from high QN count templates were rejected due to untenably high naive spontaneous noise.
P-values corresponding to
Figure 5A,B are given in
Table A7 and
Table A8 in
Appendix B. They indicate that the injury-mitigating effect of high QN ratios was meaningful (we avoid the term “significant” in association with
p-values, following the arguments in [
30]).
Our results demonstrate that the presence of parallel inhibitory neurons in a channel help protect that channel’s signal from the effects of injury (at the cost of decreased SSNR). We hypothesize that QNs achieve this by a “canceling out" mechanism: When inhibitory QNs are injured, the overall transmitted signal increases, offsetting the decreases caused by injury to excitatory PNs.
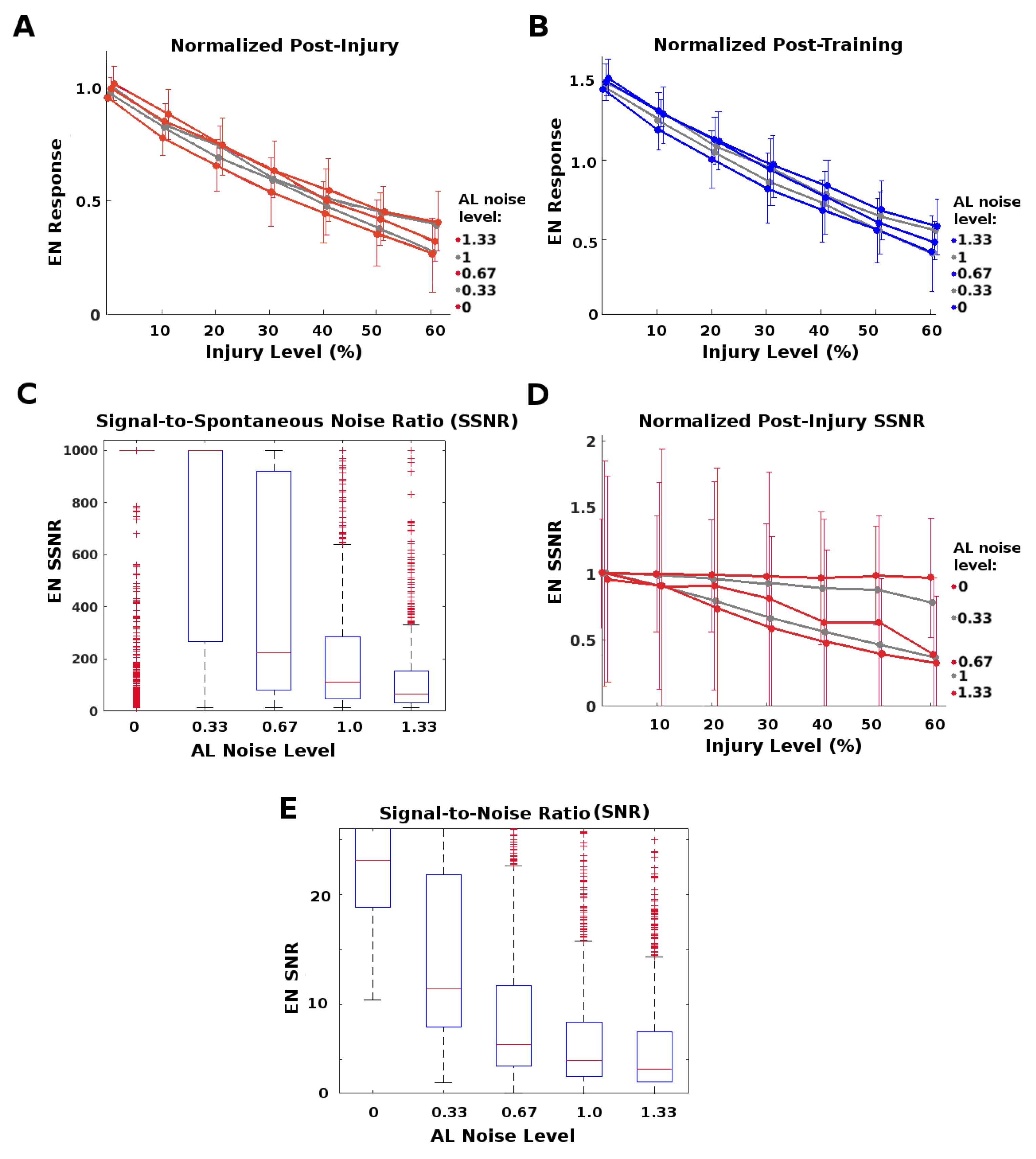
2.3. AL Noise Preserves the Highest EN Responses
The AL is a noisy network. We ask whether this neural noise has injury-mitigation benefits. We suppose that vital odor-related behavior is triggered when a discrete EN response f exceeds some threshold, and that due to AL noise these responses f to a particular odor (at a given concentration) vary as if drawn from a distribution. The moth gets n exposures to a given odor plume, and thus has n discrete responses . Then to induce the behavior, a triggering response (i.e., threshold) is needed for only some, not all, .
In this case, it suffices for the system to protect only the strongest (top-scoring) EN responses from injury-induced attenuation in order to maintain its behavioral response. The goal of this experiment was to examine whether higher AL noise levels might preferentially protect the top-scoring EN responses from injury-induced attenuation. Noting that
F parametrizes a Gaussian
we define this top-scoring tranche as those responses at the top end of the distribution:
. This corresponds to
boosted by fortuitous noise effects and thus most likely to exceed the triggering threshold.
The AL noise level is controlled by a single parameter in MothNet. We adjusted neural noise in the AL to different multiplicative factors of the “natural” AL noise level (i.e., the level calibrated to in vivo data). Factors were 0, 0.33, 0.67, 1.0, and 1.33, where 1.0 is the natural level. Various severities of FAS-like injury were applied to RNs in the Antennae→AL channel (
Figure 2A). Over 30 (31 to 62, mean = 40) moth instances were generated from template for each {AL noise, injury level} datapoint. To measure attenuation in top-scoring responses, we defined the Top-End Preservation
P as:
where
is the set of pre-injury (healthy) responses to repeat applications of some stimulus, and
is the set of discrete responses to the same stimulus post-injury at level
j.
measures how much an injury affects the top-scoring responses (represented by
) when it shrinks the entire response distribution from
to
.
ranges between 1 and 0, where 1 implies no injury-induced attenuation, and 0 implies total attenuation.
Higher AL noise increased the top-end preservation
P of EN responses caused by a given level of injury (
Figure 6A). It also increased the post-training recovery possible: For example, full recovery occurred for injury
when AL noise was greater than natural level, vs.
when AL noise = 0 (
Figure 6B).
However, high AL noise levels had a significant downside, namely, lower SNR (signal to noise ratio) and SSNR (signal to spontaneous noise ratio) values, as seen in
Figure 6C–E. This suggests that the moth must trade-off between robustness to injury and signal quality.
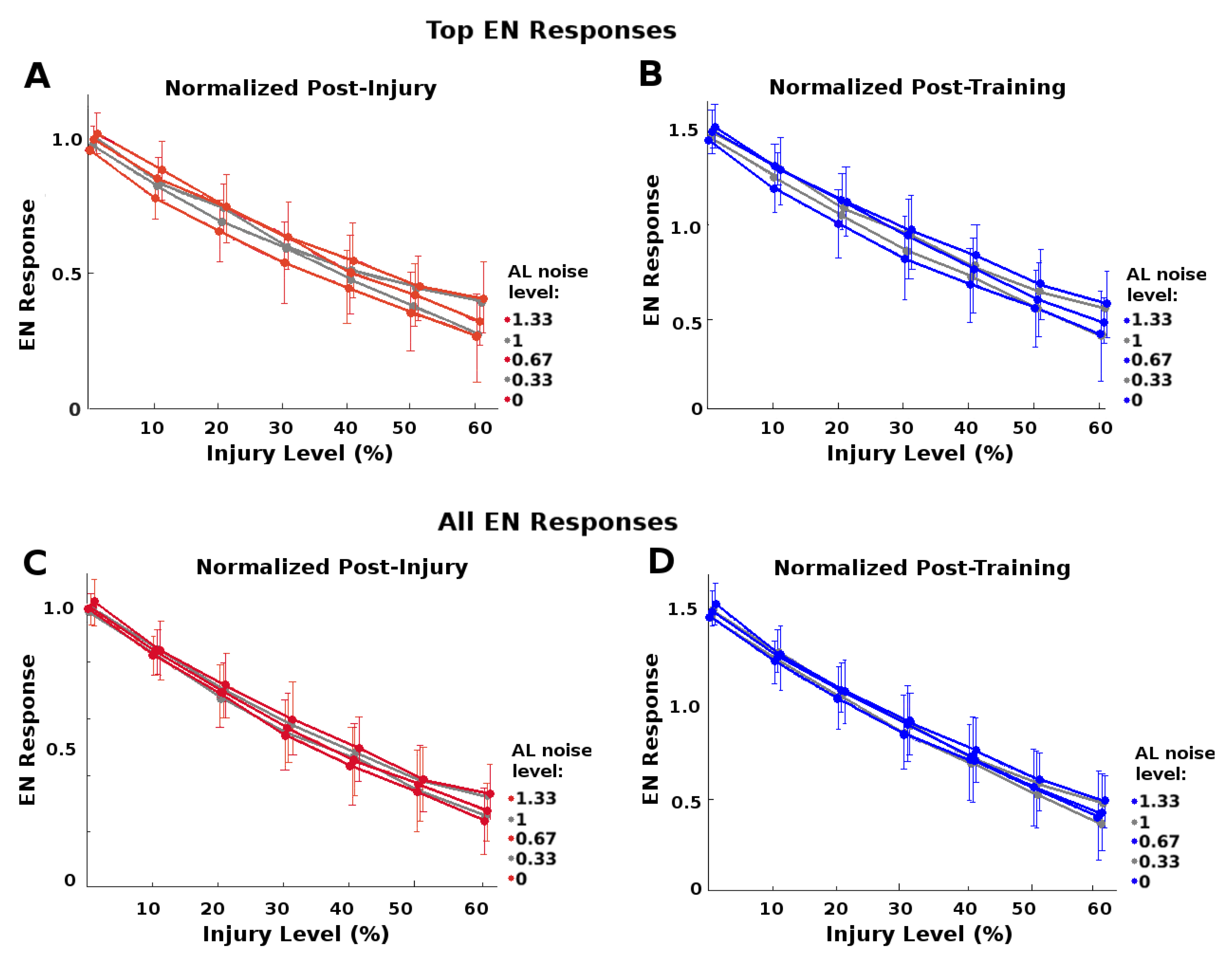
In addition, we found that this protection did not apply to all EN responses: Top-scoring EN responses received more injury-mitigation benefit from higher AL noise levels than did average EN responses, i.e.,
. That is, the extra robustness to injury conferred by higher noise levels was greater for top-scoring responses than for average responses. This meshes with the notion that the system needs not protect all responses, just the ones most likely to exceed triggering threshold.
Figure 7 shows this difference in protective effect, top-scoring vs average.
P-values are given in
Appendix B as follows:
(i) P-values for
Figure 6A,B (and
Figure 7A,B) are given in
Table A1 and
Table A3;
(ii) P-values for
Figure 7C,D are given in
Table A2 and
Table A4;
(iii) P-values comparing injury-mitigating effects on top-scoring vs average responses, (i.e.,
Figure 7, A vs. C and B vs. D) are given in
Table A5 and
Table A6. The
P-values indicate that
(i) increased AL noise correlated meaningfully with increased protective effect on the top-scoring responses;
(ii) the protective effect was noticeably lower for average responses; and
(iii) the protective effect was meaningfully greater for top-scoring than for average responses.
Results given for this set of experiments are for injury’s effects on magnitude of EN response. Injury’s effects on odor discrimination were only slightly affected by varying AL noise, since both trained odor and control were attenuated by injury (results not shown).
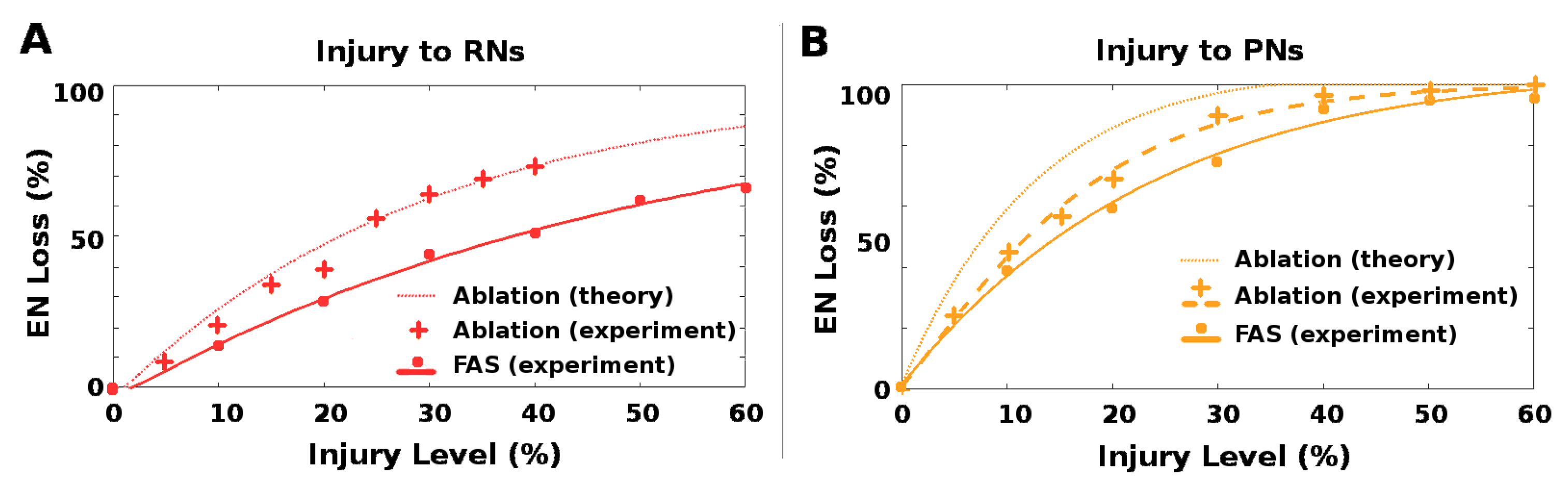
2.4. Ablation Is a Poor Proxy to Biological Injury
Neuronal pathologies are often modeled in a binary way, i.e., by treating a neuron and/or its connections as either fully functional or fully impaired, and ablation injuries are widely studied in theoretical and experimental settings. However, recent FAS studies show that most injured neurons maintain some residual firing rate activity. On large, homogeneous populations of neurons where outputs are pooled, such as the 30,000 RNs in the moth AL-MB, one can expect an approximate equivalence in ablation and FAS, modulo a conversion factor. This is because injuring or ablating any single RN has relatively small effect on the pooled total, and the overall effects of injury to the population can be approximated by average injury ratios. In this case, we estimate that ablation alone is roughly 1.75× more harmful than FAS-like injury, i.e., ablating
n% of neurons in a population causes the same relative drop in total summed FRs as FAS injury to ≈
% of the neuron population. For calculations, see
Section 5.3.3.
However, where neuron numbers are smaller and neural outputs are not pooled, so that individual neurons have relatively unique effects on the system, it is not clear that ablation effects can be reliably mapped to effects of more biologically-plausible FAS-like injuries. Injury to the AL→MB channel (i.e., PNs and QNs) resembles this situation, since there are only 5 PNs per AL glomerulus.
In ablation studies, injury levels are typically measured as percentage of neurons ablated. To assess whether ablation is a good proxy for naturalistic FAS injury, we ran experiments to test whether the impacts on EN response magnitude of ablation injury vs. FAS-like injury had a consistent 1.75× relationship at these two locations, i.e., the RN channel (red stars in
Figure 2) and the PN channel (orange stars in
Figure 2). All parameters were generated from a MothNet template with AL noise at natural levels and number of QNs = 0 (QNs = 2 gave similar results). Half the moths were injured by ablation and half were injured by FAS, with injury levels from 0 to 60%, in order to compare the relative empirical effects on EN outputs. In each experiment, over 30 moth instances (31 to 40, mean = 35) were generated for each injury {type, level, location} datapoint.
The qualitative effects of ablation and FAS were similar, at each injury site. However, the relative quantitative effects (on EN outputs) of the two injury types varied greatly depending on the site of injury. For RN channel damage, ablation effects were roughly in line with that predicted by theory for large homogeneous populations, i.e., 1.75× FAS damage. The match makes sense given the assumptions on number and distribution of RNs stated above, and is seen in
Figure 8A.
In contrast, ablation injury to the PN channel was much less harmful relative to FAS-like injury than predicted by theory. For example, 10% ablation would theoretically induce the same EN loss as 17.5% FAS injury and 20% ablation would correspond to 35% FAS injury. However, in our experiments 10% ablation corresponded to only ∼12% FAS injury (a ratio of 1.2), and 20% ablation corresponded to only ∼25% (a ratio of 1.25). This effect is seen in
Figure 8B by following horizontal lines, which correspond to equivalent EN loss, and comparing the percentage injury levels of (from right to left) experiment FAS, experiment ablation, and theoretical ablation. The experimental ablation levels required to induce a fixed EN loss were much closer to the FAS levels than theory predicted (ratio ∼1.25 instead of 1.75).
We remark that this measured discrepancy between theoretical and actual effects is not at the site of injury, but at the downstream ENs, i.e., after the impact of the upstream injury has been nonlinearly modulated by moving through the cascaded system.
This variability in the ratio of ablation injury to equivalent FAS injury, dependent on which neurons are injured, suggests that ablation may be an unreliable proxy for naturalistic neuronal damage in some contexts.
3. Discussion
Our simulations indicate that the neural mechanisms and motifs we tested have clear injury-mitigation properties. In this section, we suggest mechanisms by which these structures might protect readout neuronal activity from upstream injury. We note that from a functional point of view, overall resilience of a cascaded system is determined by whether downstream units can still transmit key readout signals to the rest of the body despite upstream damage. Cascaded networks are ubiquitous among biological neural systems, so the principles discussed in the moth’s olfactory network may be applicable to other settings. We also discuss the discrepancy between axonal swelling injuries vs. ablation injuries. Finally, we argue that robustness to injury is a key principle of biological neural design.
3.1. Hebbian Plasticity-Induced Recovery from Injury
Learning in the moth olfactory network occurs via a combination of octopamine stimulation and Hebbian growth. Octopamine stimulation temporarily boosts neural firing rates during reinforcement by sugar reward, while Hebbian updates strengthen the synaptic weight
, between two neurons
a and
b, proportionally to the product of their firing rates:
Injuries to the upstream regions of a network result in spike deletions and weaker encodings arriving at downstream neurons. If the damaged region cannot activate downstream neurons with the existing synaptic connection strengths, there is a functional loss of information. However, the combination of octopamine stimulation and a Hebbian update mechanism can evidently mitigate or reverse this effect. The original injured neurons are not themselves repaired, as plasticity only boosts downstream synaptic connections.
The Hebbian mechanism alone is not sufficient to repair damage, because updates to synaptic strengths require sufficiently strong FRs in both the incoming and the receiving neurons. If injury reduces the FRs of upstream neurons, and these reduced inputs lead to lower FRs in downstream neurons, Hebbian updates are significantly degraded. Thus, octopamine-induced stimulation of the injured upstream neurons is crucial to post-injury plasticity, because it temporarily boosts the FRs of injured upstream neurons back to levels that enable the Hebbian mechanism to strengthen the relevant connections.
We propose that degraded firing rates in downstream neurons are restored via the following mechanism (see schematic in
Figure 9A):
Octopamine temporarily increases the firing rates of injured upstream neurons.
The transient boosted encodings are sufficient to trigger firing in the downstream neurons with the existing synaptic connection strengths.
Since neurons on both sides of the plastic connections are firing, Hebbian growth strengthens their connections.
Firing rates from the injured upstream region return to their reduced rate once octopamine is withdrawn. However, due to the stronger synaptic connections, these encodings are now sufficient to trigger the downstream neurons. This restores the transmission of key information to the rest of the system.
Because the {octopamine stimulation + Hebbian updates} learning mechanism is automated, i.e., hard-wired as a reward mechanism for adaptive stimuli such as sugar, it acts as a passive injury mitigation system (absent injury, it serves to boost network responses to adaptive stimuli). Since learning is activated repeatedly throughout life (by any rewarding stimulus), it can be expected to act post-injury as an automatic repair mechanism. Alternately, learning can be viewed as a built-in tuning mechanism that in event of injury serves to restore network responses towards their pre-injury states.
We hesitate to call this learning mechanism homeostatic, even though in the context of injury it automatically moves the system towards a prior state, because the restoration is one-way. Learning will not revert responses that have been previously strengthened (by learning itself). Rather, it is an automated mechanism for tuning a network towards stronger responses to adaptive stimuli, which in the event of injury has a homeostatic effect.
3.2. Inhibitory Neurons and Protective Canceling Out Effect
The moth olfactory network has both excitatory projection neurons and inhibitory projection neurons that feed-forward from the antennal lobe to the mushroom body. We propose a mechanism to explain how this can protect downstream neurons from the effects of upstream damage, assuming downstream dynamics depend on the summed input from upstream neurons:
When FAS-like injury is applied to the PN+QN pipeline in our neural architecture, the net effect on the summed signal reaching downstream target neurons varies according to the proportion of QNs to PNs (
, assuming uniform weights
). When all feed-forward signals are excitatory (i.e.,
= 0), injury will always reduce the summed input reaching a downstream neuron. If QNs exist, however, and both PNs and QNs share the same exposure to injury, then the overall reduction to the summed input will be mitigated on average, since any injury to QNs will increase the summed input, offsetting decreases due to PN injury. A schematic of this “cancelling out” mechanism is shown in
Figure 9B.
The injury resistance provided by high QN counts comes at a cost to other functionalities, e.g., higher spontaneous EN noise relative to odor response. Presumably, biological networks have QN counts which optimally balance the benefits of injury mitigation on one hand versus the need for high signal-noise-ratio, as well as other concerns such as the energy cost to the organism. If the QN counts are low (e.g., QN:PN ≤ 20%, as in the moth), this injury mitigation benefit is likely less important relative to other architectural or functional constraints. We note that learning and plasticity are not pre-requisites for this mechanism.
3.3. Upstream Noise Protects Downstream Behavior
Suppose that the behavioral response is preserved after injury if at least a subset of stimuli elicit downstream responses that exceed action-triggering thresholds. In this case, a large noise envelope on upstream neurons may help protect the network’s functionality.
Assume the firing rate of an upstream neuron FR responds to stimuli following a Gaussian distribution
, and that it needs to exceed a threshold
T to activate downstream neurons. If the neural damage reduces this FR in average by
, a large noise envelope (large
) will ensure that some post-injury responses still exceed threshold, i.e., that
. This idea is sketched in
Figure 9C for two FRs characterized by
and
with
.
Our experiments indicate that AL noise does enable the highest EN responses to exceed threshold after injury, even as the average EN response drops. However, the injury mitigating benefit of increased upstream noise comes at a cost to other system functionalities, e.g., it reduces signal-to-noise ratio. Noise levels in biological networks (such as in the antennal lobe) may represent an evolved/optimized trade-off between injury mitigation effects and negative side-effects such as reduced SNR. We note that the sparsity of the MB acts as a powerful noise filter [
7]. Plasticity is not a pre-requisite to this mechanism.
3.4. Ablation Is a Poor Proxy to Biological Injury
Neuronal injuries are often modeled in a binary way, i.e., by treating a neuron and/or its connections as either fully functional or fully impaired. Our results indicate, however, that in some situations ablations are a poor proxy for more naturalistic FAS-types of injuries regarding effects measured downstream from the injury site.
When the neuron population to be injured is large, and has pooled outputs to the next layers (in our model, the antennae/RNs), ablation maps to FAS injury in a predictable manner due to averaging effects over the population (see
Section 5.3.3). However, when the neuron population is small (in our model, the PNs) the effects of ablation vs. FAS are not predictable. Ablation of PNs had much lower impact than large-population theory would predict.
Our key finding is that ablation effects are inconsistent relative to FAS-like effects, depending on the location and characteristics of the injured neurons. This calls into question the value of ablation as a proxy for naturalistic neural injuries. We suggest that in systems with large numbers of somewhat interchangeable units (e.g., the 30,000 RNs) ablation is a suitable way to model injury; while in systems with smaller numbers of specialized neural units (e.g., the AL with 60 glomeruli) ablation is a poor injury model. This unsuitability is sharpened when the actionable effects of injury are measured downstream from the regions injured, because there is a complex interplay between the injuries and network nonlinearities, making the outcome somewhat unpredictable. Simple examples of possible effects of non-linearities (AND and OR gates) that might cause ablation vs. FAS-like injury models to diverge are shown in
Figure 9D.
3.5. Limitations
Our computational model assumed only one readout neuron and one broadly-activating odor (two for the the discrimination experiments). A more detailed assessment of injury and mitigation might involve several readout neurons to allow for disparate effects on various readouts, as well as several time-varying odor mixtures, more narrowly-activating odors, and concentration gradients. In addition, we model learning-induced Hebbian plasticity in existing neurons, not other neurogenesis mechanisms. We model the effects of octopamine (since it is closely involved in learning), not other neuromodulators such as serotonin. We do not know if the PN+QN channel is a realistic target for injury: We chose it in order to investigate deficits caused by injury to innermost hubs. Our study certainly did not exhaust all potentially interesting combinations of structures-under-test and injuries. More elaborate models could incorporate extra-neural factors such as moth physiology and physiological status, and external environmental conditions such as temperature and humidity. These conditions are beyond the scope of MothNet and the in vivo experiments that inform it.
4. Conclusions
We investigated the moth olfactory network with the goal of understanding how its basic architectural components serve to make sensory processing robust to injury. Since most organisms are exposed to neuronal damage throughout their lives, it is important to understand how such neuronal circuits are structured to maintain functionality despite impairments. In this work, we showed explicitly how certain structural and functional network motifs act as injury mitigation mechanisms. Specifically, we examined how (i) Hebbian learning, (ii) high levels of noise, and (iii) presence of parallel inhibitory and excitatory connections, can support overall robustness to injury in the olfactory system in the Manduca sexta moth.
Our findings indicate that, in addition to accurate sensory processing, biological neural networks such as those found in the moth olfactory system hold robustness to injury as a central design principle. Our findings also suggest an additional hypothesis: Plasticity coupled with neuromodulatory stimulation, now central to learning, may have originally evolved as a repair mechanism for neural systems to offset injury and maintain function, and was only later ported to the task of developing responses to new information (exaptation). If this is the case, then the gift of learning is due originally to the exigencies of brain damage.
Our results also show that these architectures can in fact cause worse performance by some other performance metrics, e.g., SNR. Thus, trying to explain these architectures from the point-of-view of, for example, information theory risks running against the fact that they are actually suboptimal according to that particular lens. A more comprehensive and nuanced framing of the neural signal processing task, positing multiple design goals including injury mitigation, can enable better understanding of neurosensory processing. That is, a neural architecture can be understood only if its injury mitigation function, and the trade-offs between this and other desired functions, are considered. Indeed, it is possible that some neural structures and mechanisms, including the ability to learn, are best understood as evolutionary solutions to the challenge of maintaining function despite injury.




{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}








