Differential Expression of CD31 and Von Willebrand Factor on Endothelial Cells in Different Regions of the Human Brain: Potential Implications for Cerebral Malaria Pathogenesis


Abstract
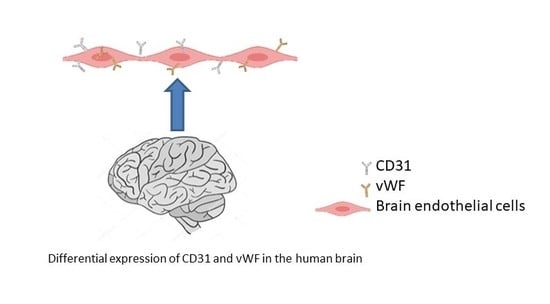
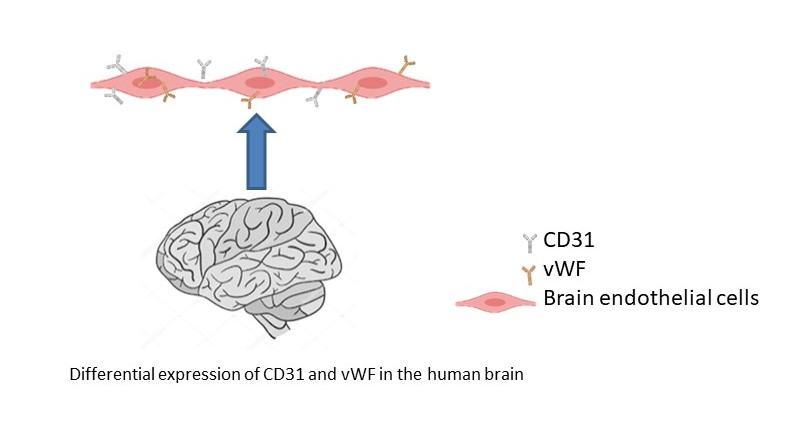
1. Introduction
2. Materials and Methods
2.1. Acquisition of Human Tissue Samples
2.2. Immunohistochemistry
2.2.1. Single Staining
2.2.2. Double Staining of CD31 and vWF
2.3. Imaging and Analysis
2.4. Percentage Area Quantification of CD31 and vWF Expression
2.5. Statistical Analysis
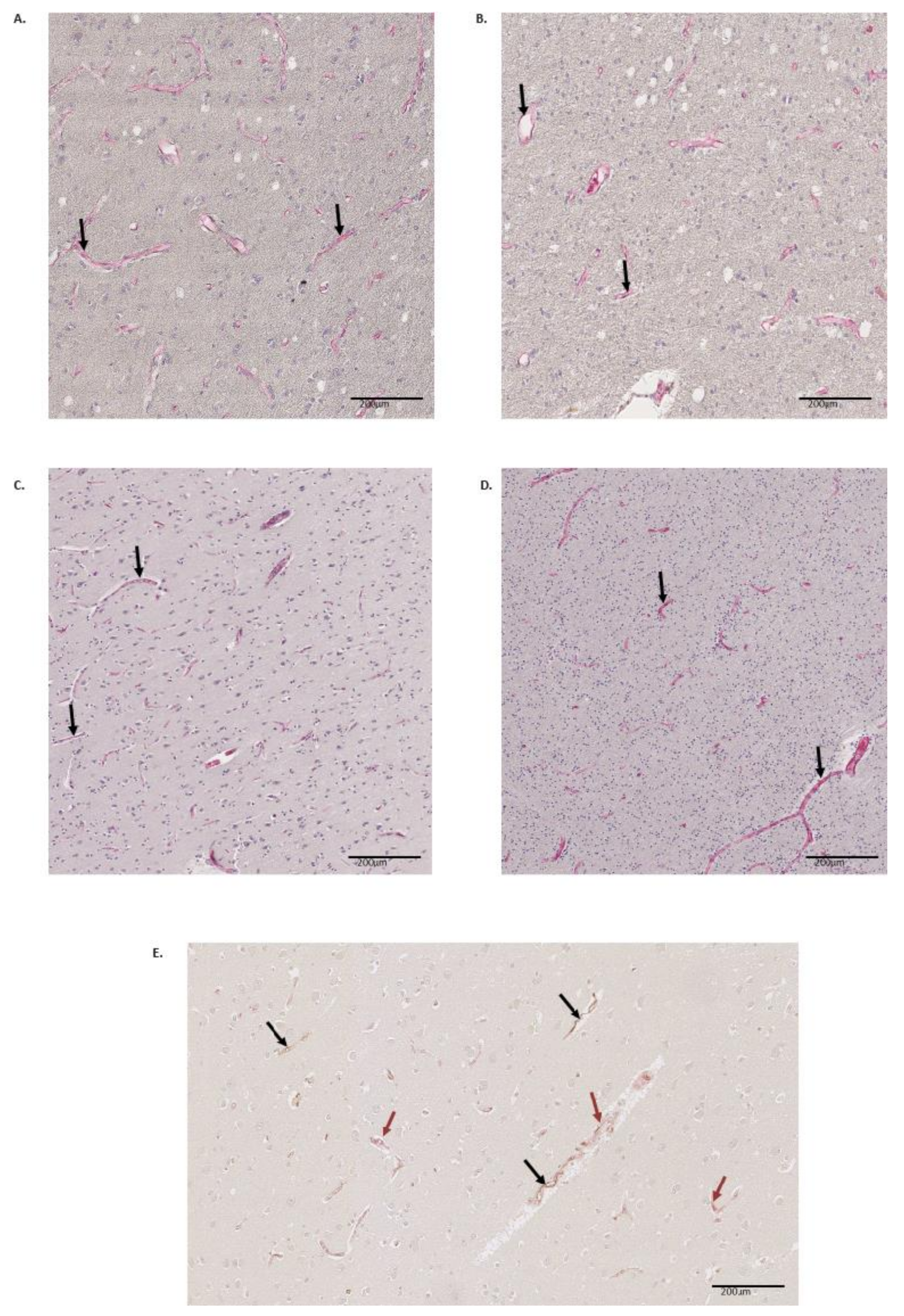
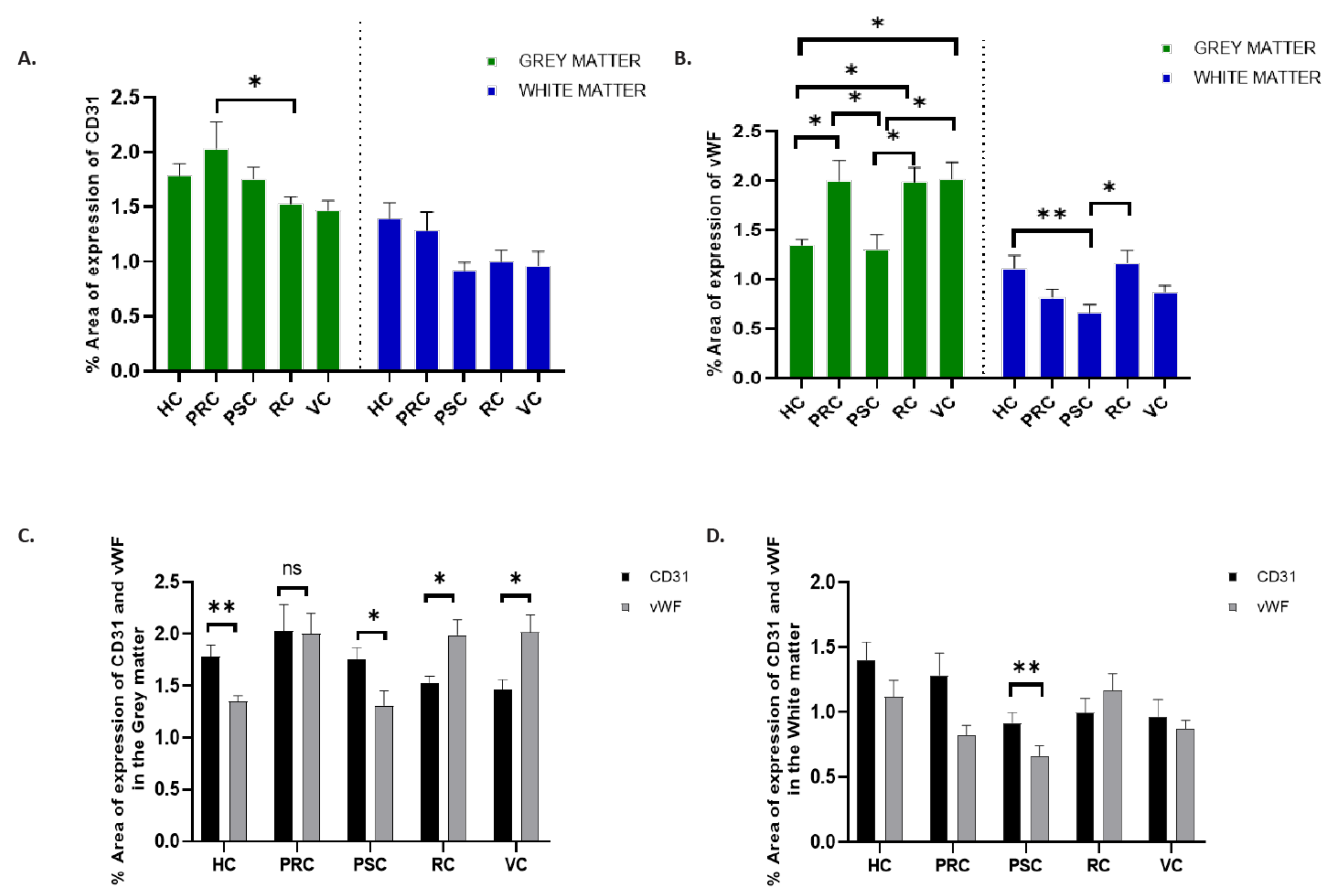
3. Results
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Chandra, A.; Li, W.A.; Stone, C.R.; Geng, X.; Ding, Y. The cerebral circulation and cerebrovascular disease I: Anatomy. Brain Circ. 2017, 3, 45–56. [Google Scholar] [CrossRef] [PubMed]
- Kisler, K.; Lazic, D.; Sweeney, M.D.; Plunkett, S.; El Khatib, M.; Vinogradov, S.A.; Boas, D.A.; Sakadži, S.; Zlokovic, B.V. In vivo imaging and analysis of cerebrovascular hemodynamic responses and tissue oxygenation in the mouse brain. Nat. Protoc. 2018, 13, 1377–1402. [Google Scholar] [CrossRef] [PubMed]
- Linninger, A.A.; Gould, I.G.; Marrinan, T.; Hsu, C.-Y.; Chojecki, M.; Alaraj, A. Cerebral microcirculation and oxygen tension in the human secondary cortex. Ann. Biomed. Eng. 2013, 41, 2264–2284. [Google Scholar] [CrossRef] [PubMed]
- Willie, C.K.; Tzeng, Y.-C.; Fisher, J.A.; Ainslie, P.N. Integrative regulation of human brain blood flow. J. Physiol. 2014, 592, 841–859. [Google Scholar] [CrossRef] [PubMed]
- Nyúl-Tóth, Á.; Suciu, M.; Molnár, J.; Fazakas, C.; Haskó, J.; Herman, H.; Farkas, A.E.; Kaszaki, J.; Hermenean, A.; Wilhelm, I.; et al. Differences in the molecular structure of the blood-brain barrier in the cerebral cortex and white matter: An in silico, in vitro, and ex vivo study. Am. J. Physiol. Heart Circ. Physiol. 2016, 310, H1702–H1714. [Google Scholar] [CrossRef] [PubMed]
- Peterson, E.C.; Wang, Z.; Britz, G. Regulation of cerebral blood flow. Int. J. Vasc. Med. 2011, 2011, 823525. [Google Scholar] [CrossRef]
- McCarron, R.M.; Chen, Y.; Tomori, T.; Strasser, A.; Mechoulam, R.; Shohami, E.; Spatz, M. Endothelial-mediated regulation of cerebral microcirculation. J. Physiol. Pharm. 2006, 57 (Suppl. S11), 133–144. [Google Scholar]
- Kolinko, Y.; Krakorova, K.; Cendelin, J.; Tonar, Z.; Kralickova, M. Microcirculation of the brain: Morphological assessment in degenerative diseases and restoration processes. Rev. Neurosci. 2015, 26, 75–93. [Google Scholar] [CrossRef]
- Chandra, A.; Stone, C.R.; Li, W.A.; Geng, X.; Ding, Y. The cerebral circulation and cerebrovascular disease II: Pathogenesis of cerebrovascular disease. Brain Circ. 2017, 3, 57–65. [Google Scholar] [CrossRef]
- Chen, Z.; Venkat, P.; Seyfried, D.; Chopp, M.; Yan, T.; Chen, J. Brain-heart interaction: Cardiac complications after stroke. Circ. Res. 2017, 121, 451–468. [Google Scholar] [CrossRef]
- Sweeney, M.D.; Sagare, A.P.; Zlokovic, B.V. Blood–brain barrier breakdown in Alzheimer’s disease and other neurodegenerative disorders. Nat. Rev. Neurol. 2018, 14, 133–150. [Google Scholar] [CrossRef] [PubMed]
- Sandsmark, D.K.; Bogoslovsky, T.; Qu, B.-X.; Haber, M.; Cota, M.R.; Davis, C.; Butman, J.A.; Latour, L.L.; Diaz-Arrastia, R. Changes in Plasma von Willebrand Factor and Cellular Fibronectin in MRI-Defined Traumatic Microvascular Injury. Front. Neurol. 2019, 10, 246. [Google Scholar] [CrossRef] [PubMed]
- Nagatake, T.; Hoang, V.T.; Tegoshi, T.; Rabbege, J.; Ann, T.K.; Aikawa, M. Pathology of falciparum malaria in Vietnam. Am. J. Trop. Med. Hyg. 1992, 47, 259–264. [Google Scholar] [CrossRef] [PubMed]
- Pongponratn, E.; Turner, G.D.H.; Day, N.P.J.; Phu, N.H.; Simpson, J.A.; Stepniewska, K.; Mai, N.T.H.; Viriyavejakul, P.; Looareesuwan, S.; Hien, T.T.; et al. An ultrastructural study of the brain in fatal Plasmodium falciparum malaria. Am. J. Trop. Med. Hyg. 2003, 69, 345–359. [Google Scholar] [CrossRef]
- Pusztaszeri, M.P.; Seelentag, W.; Bosman, F.T. Immunohistochemical expression of endothelial markers CD31, CD34, von Willebrand factor, and Fli-1 in normal human tissues. J. Histochem. Cytochem. 2006, 54, 385–395. [Google Scholar] [CrossRef]
- Middleton, J.; Americh, L.; Gayon, R.; Julien, D.; Mansat, M.; Mansat, P.; Anract, P.; Cantagrel, A.; Cattan, P.; Reimund, J.-M.; et al. A comparative study of endothelial cell markers expressed in chronically inflamed human tissues: MECA-79, Duffy antigen receptor for chemokines, von Willebrand factor, CD31, CD34, CD105 and CD146. J. Pathol. 2005, 206, 260–268. [Google Scholar] [CrossRef]
- Rakocevic, J.; Orlic, D.; Mitrovic-Ajtic, O.; Tomasevic, M.; Dobric, M.; Zlatic, N.; Milasinovic, D.; Stankovic, G.; Ostojić, M.; Labudovic-Borovic, M. Endothelial cell markers from clinician’s perspective. Exp. Mol. Pathol. 2017, 102, 303–313. [Google Scholar] [CrossRef]
- Hao, Q.; Chen, X.-L.; Ma, L.; Wang, T.-T.; Hu, Y.; Zhao, Y.-L. Procedure for the Isolation of Endothelial Cells from Human Cerebral Arteriovenous Malformation (cAVM) Tissues. Front. Cell. Neurosci. 2018, 12, 2426. [Google Scholar] [CrossRef]
- Noumbissi, M.E.; Galasso, B.; Stins, M.F. Brain vascular heterogeneity: Implications for disease pathogenesis and design of in vitro blood-brain barrier models. Fluids Barriers CNS 2018, 15, 12. [Google Scholar] [CrossRef]
- Cucullo, L.; Hossain, M.; Puvenna, V.; Marchi, N.; Janigro, D. The role of shear stress in Blood-Brain Barrier endothelial physiology. BMC Neurosci. 2011, 12, 3915. [Google Scholar] [CrossRef]
- Wilhelm, I.; Nyúl-Tóth, Á.; Suciu, M.; Hermenean, A.; Krizbai, I.A. Heterogeneity of the blood-brain barrier. Tissue Barriers 2016, 4, e1143544. [Google Scholar] [CrossRef] [PubMed]
- Macdonald, J.A.; Murugesan, N.; Pachter, J.S. Endothelial cell heterogeneity of blood-brain barrier gene expression along the cerebral microvasculature. J. Neurosci. Res. 2010, 88, 1457–1474. [Google Scholar] [CrossRef] [PubMed]
- Navone, S.; Marfia, G.; Nava, S.; Invernici, G.; Cristini, S.; Balbi, S.; Sangiorgi, S.; Ciusani, E.; Bosutti, A.; Alessandri, G.; et al. Human and mouse brain-derived endothelial cells require high levels of growth factors medium for their isolation, in vitro maintenance and survival. Vasc. Cell 2013, 5, 10. [Google Scholar] [CrossRef] [PubMed]
- Goncharov, N.V.; Nadeev, A.D.; Jenkins, R.O.; Avdonin, P.V. Markers and Biomarkers of Endothelium: When Something Is Rotten in the State. Oxidative Med. Cell. Longev. 2017, 2017, 1–27. [Google Scholar] [CrossRef] [PubMed]
- Navone, S.E.; Marfia, G.; Invernici, G.; Cristini, S.; Nava, S.; Balbi, S.; Sangiorgi, S.; Ciusani, E.; Bosutti, A.; Alessandri, G.; et al. Isolation and expansion of human and mouse brain microvascular endothelial cells. Nat. Protoc. 2013, 8, 1680–1693. [Google Scholar] [CrossRef]
- Ambrose, C.T. The Role of Capillaries in the Lesser Ailments of Old Age and in Alzheimer’s Disease and Vascular Dementia: The Potential of Pro-Therapeutic Angiogenesis. J. Alzheimers Dis. 2016, 54, 31–43. [Google Scholar] [CrossRef]
- Yamazaki, Y.; Baker, D.J.; Tachibana, M.; Liu, C.-C.; van Deursen, J.M.; Brott, T.G.; Bu, G.; Kanekiyo, T. Vascular Cell Senescence Contributes to Blood-Brain Barrier Breakdown. Stroke 2016, 47, 1068–1077. [Google Scholar] [CrossRef]
- Chen, M.B.; Yousef, H.; Yang, A.C.; Lee, D.; Lehallier, B.; Schaum, N.; Quake, S.R.; Wyss-Coray, T. Brain endothelial cells are exquisite sensors of age-related circulatory cues. Cell Rep. 2019. [Google Scholar] [CrossRef]
- Xu, X.; Wang, B.; Ren, C.; Hu, J.; Greenberg, D.A.; Chen, T.; Xie, L.; Jin, K. Age-related Impairment of Vascular Structure and Functions. Aging Dis. 2017, 8, 590–610. [Google Scholar] [CrossRef]
- Csiszar, A.; Tarantini, S.; Fülöp, G.A.; Kiss, T.; Valcarcel-Ares, M.N.; Galvan, V.; Ungvari, Z.; Yabluchanskiy, A. Hypertension impairs neurovascular coupling and promotes microvascular injury: Role in exacerbation of Alzheimer’s disease. Geroscience 2017, 39, 359–372. [Google Scholar] [CrossRef]
- Park, J.-H.; Hong, J.-H.; Lee, S.-W.; Ji, H.D.; Jung, J.-A.; Yoon, K.-W.; Lee, J.-I.; Won, K.S.; Song, B.-I.; Kim, H.W. The effect of chronic cerebral hypoperfusion on the pathology of Alzheimer’s disease: A positron emission tomography study in rats. Sci. Rep. 2019, 9, 14102. [Google Scholar] [CrossRef] [PubMed]
- Ambrose, C.T. Neuroangiogenesis: A vascular basis for Alzheimer’s disease and cognitive decline during aging. J. Alzheimers Dis. 2012, 32, 773–788. [Google Scholar] [CrossRef] [PubMed]
- Mühlberger, N.; Jelinek, T.; Behrens, R.H.; Gjørup, I.; Coulaud, J.P.; Clerinx, J.; Puente, S.; Burchard, G.; Gascon, J.; Grobusch, M.P.; et al. Age as a risk factor for severe manifestations and fatal outcome of falciparum malaria in European patients: Observations from TropNetEurop and SIMPID Surveillance Data. Clin. Infect. Dis. 2003, 36, 990–995. [Google Scholar] [CrossRef] [PubMed]
- Hammarlund-Udenaes, M.; Fridén, M.; Syvänen, S.; Gupta, A. On the rate and extent of drug delivery to the brain. Pharm. Res. 2008, 25, 1737–1750. [Google Scholar] [CrossRef]
- Moxon, C.A.; Wassmer, S.C.; Milner, D.A.; Chisala, N.V.; Taylor, T.E.; Seydel, K.B.; Molyneux, M.E.; Faragher, B.; Esmon, C.T.; Downey, C.; et al. Loss of endothelial protein C receptors links coagulation and inflammation to parasite sequestration in cerebral malaria in African children. Blood 2013, 122, 842–851. [Google Scholar] [CrossRef]
- MacPherson, G.G.; Warrell, M.J.; White, N.J.; Looareesuwan, S.; Warrell, D.A. Human cerebral malaria. A quantitative ultrastructural analysis of parasitized erythrocyte sequestration. Am. J. Pathol. 1985, 119, 385–401. [Google Scholar]
- Dorovini-Zis, K.; Schmidt, K.; Huynh, H.; Fu, W.; Whitten, R.O.; Milner, D.; Kamiza, S.; Molyneux, M.; Taylor, T.E. The neuropathology of fatal cerebral malaria in malawian children. Am. J. Pathol. 2011, 178, 2146–2158. [Google Scholar] [CrossRef]
- Taylor, T.E.; Fu, W.J.; Carr, R.A.; Whitten, R.O.; Mueller, J.S.; Fosiko, N.G.; Lewallen, S.; Liomba, N.G.; Molyneux, M.E.; Mueller, J.G. Differentiating the pathologies of cerebral malaria by postmortem parasite counts. Nat. Med. 2004, 10, 143–145. [Google Scholar] [CrossRef]
- Van Horssen, J.; Brink, B.P.; de Vries, H.E.; van der Valk, P.; Bø, L. The blood-brain barrier in cortical multiple sclerosis lesions. J. Neuropathol. Exp. Neurol. 2007, 66, 321–328. [Google Scholar] [CrossRef]
- Prins, M.; Schul, E.; Geurts, J.; van der Valk, P.; Drukarch, B.; van Dam, A.-M. Pathological differences between white and grey matter multiple sclerosis lesions. Ann. N. Y. Acad. Sci. 2015, 1351, 99–113. [Google Scholar] [CrossRef]
- Wimmer, I.; Tietz, S.; Nishihara, H.; Deutsch, U.; Sallusto, F.; Gosselet, F.; Lyck, R.; Muller, W.A.; Lassmann, H.; Engelhardt, B. PECAM-1 Stabilizes Blood-Brain Barrier Integrity and Favors Paracellular T-Cell Diapedesis Across the Blood-Brain Barrier During Neuroinflammation. Front. Immunol. 2019, 10, 711. [Google Scholar] [CrossRef] [PubMed]
- Harada, N.; Masuda, M.; Fujiwara, K. Fluid flow and osmotic stress induce tyrosine phosphorylation of an endothelial cell 128 kDa surface glycoprotein. Biochem. Biophys. Res. Commun. 1995, 214, 69–74. [Google Scholar] [CrossRef] [PubMed]
- Tai, L.-K.; Zheng, Q.; Pan, S.; Jin, Z.-G.; Berk, B.C. Flow activates ERK1/2 and endothelial nitric oxide synthase via a pathway involving PECAM1, SHP2, and Tie2. J. Biol. Chem. 2005, 280, 29620–29624. [Google Scholar] [CrossRef] [PubMed]
- Chen, Z.; Tzima, E. PECAM-1 is necessary for flow-induced vascular remodeling. Arterioscler. Thromb. Vasc. Biol. 2009, 29, 1067–1073. [Google Scholar] [CrossRef]
- Privratsky, J.R.; Newman, P.J. PECAM-1: Regulator of endothelial junctional integrity. Cell Tissue Res. 2014, 355, 607–619. [Google Scholar] [CrossRef]
- Woodfin, A.; Voisin, M.-B.; Nourshargh, S. PECAM-1: A multi-functional molecule in inflammation and vascular biology. Arterioscler. Thromb. Vasc. Biol. 2007, 27, 2514–2523. [Google Scholar] [CrossRef]
- Yan, B.C.; Xu, P.; Gao, M.; Wang, J.; Jiang, D.; Zhu, X.; Won, M.-H.; Su, P.Q. Changes in the Blood-Brain Barrier Function Are Associated With Hippocampal Neuron Death in a Kainic Acid Mouse Model of Epilepsy. Front. Neurol. 2018, 9, 775. [Google Scholar] [CrossRef]
- Brown, H.; Hien, T.T.; Day, N.; Mai, N.T.; Chuong, L.V.; Chau, T.T.; Loc, P.P.; Phu, N.H.; Bethell, D.; Farrar, J.; et al. Evidence of blood-brain barrier dysfunction in human cerebral malaria. Neuropathol. Appl. Neurobiol. 1999, 25, 331–340. [Google Scholar] [CrossRef]
- Treutiger, C.J.; Heddini, A.; Fernandez, V.; Muller, W.A.; Wahlgren, M. PECAM-1/CD31, an endothelial receptor for binding Plasmodium falciparum-infected erythrocytes. Nat. Med. 1997, 3, 1405–1408. [Google Scholar] [CrossRef]
- Nishanth, G.; Schlüter, D. Blood-Brain Barrier in Cerebral Malaria: Pathogenesis and Therapeutic Intervention. Trends Parasitol. 2019, 35, 516–528. [Google Scholar] [CrossRef]
- Nacer, A.; Movila, A.; Sohet, F.; Girgis, N.M.; Gundra, U.M.; Loke, P.N.; Daneman, R.; Frevert, U. Experimental cerebral malaria pathogenesis—Hemodynamics at the blood brain barrier. PLoS Pathog. 2014, 10, e1004528. [Google Scholar] [CrossRef] [PubMed]
- Ohashi, J.; Naka, I.; Hananantachai, H.; Patarapotikul, J. Association of PECAM1/CD31 polymorphisms with cerebral malaria. Int. J. Mol. Epidemiol. Genet. 2016, 7, 87–94. [Google Scholar] [PubMed]
- Mandyam, C.D.; Villalpando, E.G.; Steiner, N.L.; Quach, L.W.; Fannon, M.J.; Somkuwar, S.S. Platelet Endothelial Cell Adhesion Molecule-1 and Oligodendrogenesis: Significance in Alcohol Use Disorders. Brain Sci. 2017, 7, 131. [Google Scholar] [CrossRef] [PubMed]
- Suidan, G.L.; Brill, A.; de Meyer, S.F.; Voorhees, J.R.; Cifuni, S.M.; Cabral, J.E.; Wagner, D.D. Endothelial Von Willebrand factor promotes blood-brain barrier flexibility and provides protection from hypoxia and seizures in mice. Arterioscler. Thromb. Vasc. Biol. 2013, 33, 2112–2120. [Google Scholar] [CrossRef] [PubMed]
- Dhanesha, N.; Prakash, P.; Doddapattar, P.; Khanna, I.; Pollpeter, M.J.; Nayak, M.K.; Staber, J.M.; Chauhan, A.K. Endothelial cell-derived VWF is the major determinant that mediates VWF-dependent acute ischemic stroke by promoting post-ischemic thrombo-inflammation. Arterioscler. Thromb. Vasc. Biol. 2016, 36, 1829–1837. [Google Scholar] [CrossRef] [PubMed]
- Noubade, R.; del Rio, R.; McElvany, B.; Zachary, J.F.; Millward, J.M.; Wagner, D.D.; Offner, H.; Blankenhorn, E.P.; Teuscher, C. Von-Willebrand Factor Influences Blood Brain Barrier Permeability and Brain Inflammation in Experimental Allergic Encephalomyelitis. Am. J. Pathol. 2008, 173, 892–900. [Google Scholar] [CrossRef]
- Wu, Y.; Liu, W.; Zhou, Y.; Hilton, T.; Zhao, Z.; Wang, M.; Yeon, J.; Houck, K.; Thiagarajan, P.; Zhang, F.; et al. von Willebrand factor enhances microvesicle-induced vascular leakage and coagulopathy in mice with traumatic brain injury. Blood 2018, 132, 1075–1084. [Google Scholar] [CrossRef]
- Hollestelle, M.J.; Donkor, C.; Mantey, E.A.; Chakravorty, S.J.; Craig, A.; Akoto, A.O.; O’Donnell, J.; van Mourik, J.A.; Bunn, J. von Willebrand factor propeptide in malaria: Evidence of acute endothelial cell activation. Br. J. Haematol. 2006, 133, 562–569. [Google Scholar] [CrossRef]
- Pais, T.F.; Penha-Gonçalves, C. Brain Endothelium: The “Innate Immunity Response Hypothesis” in Cerebral Malaria Pathogenesis. Front. Immunol. 2018, 9, 3100. [Google Scholar] [CrossRef]
- Moxon, C.A.; Heyderman, R.S.; Wassmer, S.C. Dysregulation of coagulation in cerebral malaria. Mol. Biochem. Parasitol. 2009, 166, 99–108. [Google Scholar] [CrossRef]
- Müller, A.M.; Hermanns, M.I.; Skrzynski, C.; Nesslinger, M.; Müller, K.-M.; Kirkpatrick, C.J. Expression of the Endothelial Markers PECAM-1, vWf, and CD34 in Vivo and in Vitro. Exp. Mol. Pathol. 2002, 72, 221–229. [Google Scholar] [CrossRef] [PubMed]
- Dmitrieva, N.I.; Burg, M.B. Secretion of von Willebrand factor by endothelial cells links sodium to hypercoagulability and thrombosis. Proc. Natl. Acad. Sci. USA 2014, 111, 6485–6490. [Google Scholar] [CrossRef] [PubMed]
- Kikuchia, M.; Looareesuwanb, S.; Ubaleea, R.; Tasanorc, O.; Suzukia, F. Association of adhesion molecule PECAM-1 CD31 polymorphism with susceptibility to cerebral malaria in Thais. Parasitol. Int. 2001, 50, 235–239. [Google Scholar] [CrossRef]
- Wolters, F.J.; Boender, J.; de Vries, P.S.; Sonneveld, M.A.; Koudstaal, P.J.; de Maat, M.P.; Franco, O.H.; Ikram, M.K.; Leebeek, F.W.; Ikram, M.A. Von Willebrand factor and ADAMTS13 activity in relation to risk of dementia: A population-based study. Sci. Rep. 2018, 8, 5474. [Google Scholar] [CrossRef]
- Erickson, M.A.; Banks, W.A. Age-Associated Changes in the Immune System and Blood–Brain Barrier Functions. Int. J. Mol. Sci. 2019, 20, 1632. [Google Scholar] [CrossRef]
- Schwartz, E.; Sadetzki, S.; Murad, H.; Raveh, D. Age as a risk factor for severe Plasmodium falciparum malaria in nonimmune patients. Clin. Infect. Dis. 2001, 33, 1774–1777. [Google Scholar] [CrossRef]
- Allen, N.; Bergin, C.; Kennelly, S.P. Malaria in the returning older traveler. Trop. Dis. Travel Med. Vaccines 2016, 2, 2. [Google Scholar] [CrossRef]



{kind=link}
{kind=link}
{kind=link}
| Biomarkers | CD31 (Mean ± SEM) | vWF (Mean ± SEM) | ||||
|---|---|---|---|---|---|---|
| Brain Regions | Grey Matter (%) | White Matter (%) | p Values | Grey Matter (%) | White Matter (%) | p Values |
| Hippocampus | 1.782 ± 0.11 | 1.394 ± 0.14 | 0.0105 | 1.346 ± 0.06 | 1.116 ± 0.13 | <0.0001 |
| Precentral cortex | 2.032 ± 0.23 | 1.286 ± 0.17 | 0.1408 | 1.998 ± 0.20 | 0.8186 ± 0.08 | <0.0001 |
| Postcentral cortex | 1.759 ± 0.11 | 0.918 ± 0.08 | <0.0001 | 1.310 ± 0.14 | 0.6622 ± 0.08 | 0.0011 |
| Rhinal cortex | 1.527 ± 0.07 | 0.996 ± 0.11 | <0.0001 | 1.987 ± 0.15 | 1.169 ± 0.13 | <0.0001 |
| Visual cortex | 1.466 ± 0.90 | 0.964 ± 0.13 | <0.0001 | 2.013 ± 0.17 | 0.8761 ± 0.06 | <0.0001 |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Mbagwu, S.I.; Filgueira, L. Differential Expression of CD31 and Von Willebrand Factor on Endothelial Cells in Different Regions of the Human Brain: Potential Implications for Cerebral Malaria Pathogenesis. Brain Sci. 2020, 10, 31. https://doi.org/10.3390/brainsci10010031
Mbagwu SI, Filgueira L. Differential Expression of CD31 and Von Willebrand Factor on Endothelial Cells in Different Regions of the Human Brain: Potential Implications for Cerebral Malaria Pathogenesis. Brain Sciences. 2020; 10(1):31. https://doi.org/10.3390/brainsci10010031
Chicago/Turabian StyleMbagwu, Smart Ikechukwu, and Luis Filgueira. 2020. "Differential Expression of CD31 and Von Willebrand Factor on Endothelial Cells in Different Regions of the Human Brain: Potential Implications for Cerebral Malaria Pathogenesis" Brain Sciences 10, no. 1: 31. https://doi.org/10.3390/brainsci10010031
APA StyleMbagwu, S. I., & Filgueira, L. (2020). Differential Expression of CD31 and Von Willebrand Factor on Endothelial Cells in Different Regions of the Human Brain: Potential Implications for Cerebral Malaria Pathogenesis. Brain Sciences, 10(1), 31. https://doi.org/10.3390/brainsci10010031