Effect of Cell-Free Supernatant from Aeromonas sobria on the Spoilage of Shewanella putrefaciens in Pacific White Shrimp (Litopenaeus vannamei) with the Influence of Temperature Fluctuation



Abstract
:Featured Application
Abstract
1. Introduction
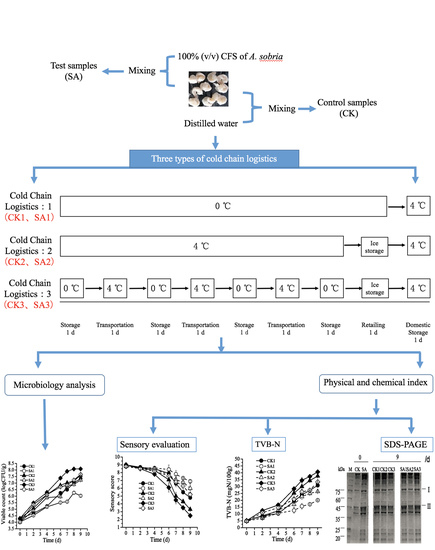
2. Materials and Methods
2.1. Bacterial Strains
2.2. Preparation of CFS
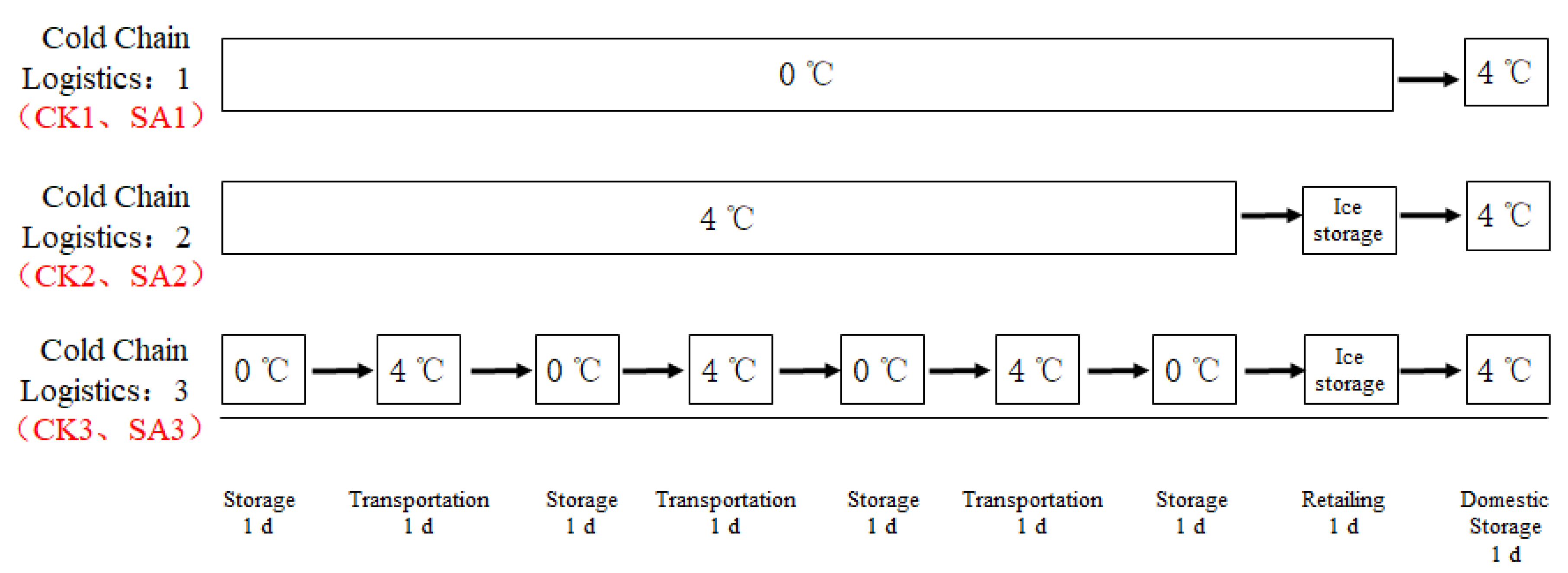
2.3. Sample Preparation
2.4. Microbiological Analysis
2.5. Sensory Evaluation
2.6. Determination of Total Volatile Basic Nitrogen (TVB-N)
2.7. Biogenic Amines Analysis
2.8. Amino Acid Analysis
2.9. Extraction of Muscle Proteins and Electrophoresis
2.10. Statistical Analysis
3. Results
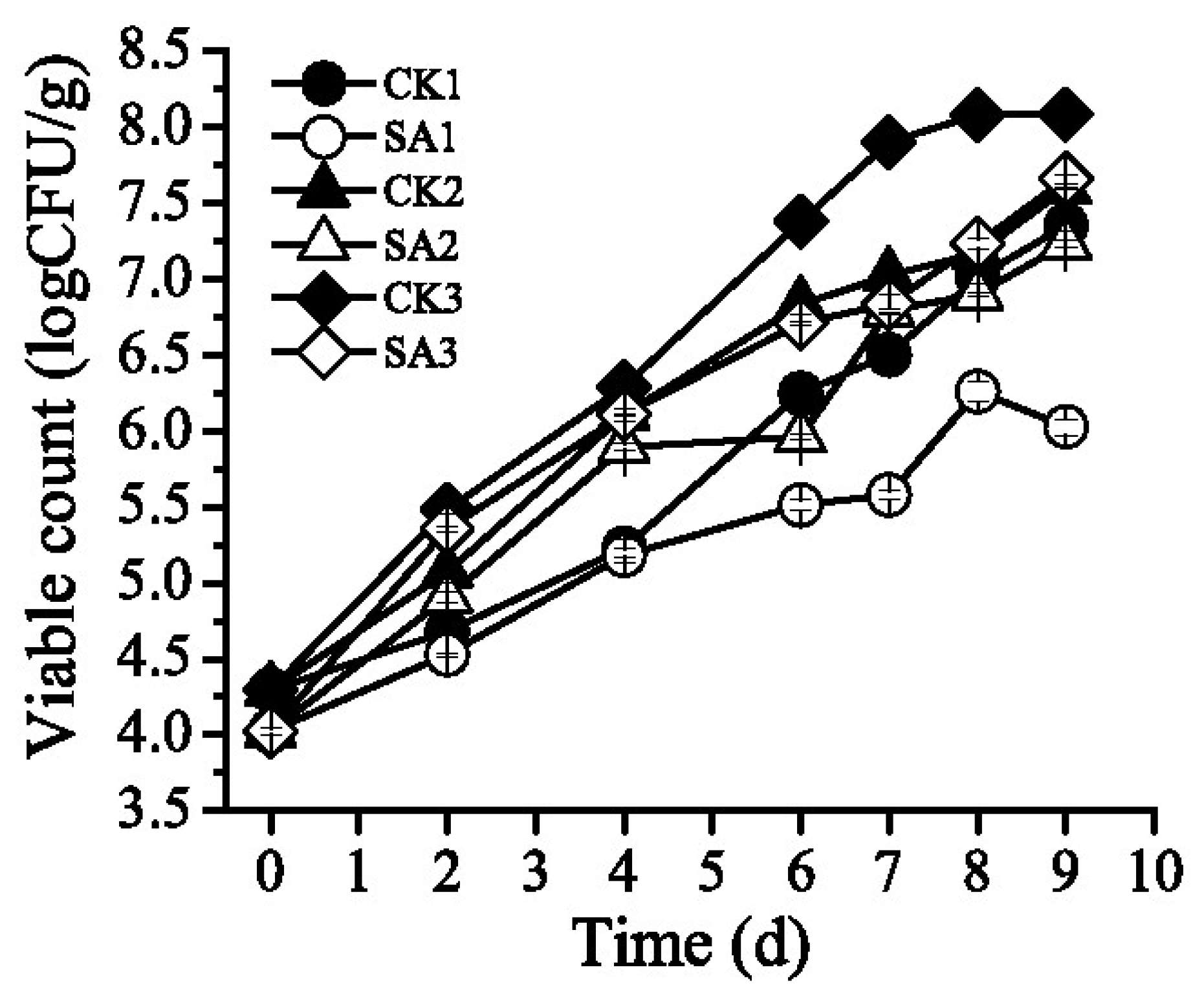
3.1. Effect of the CFS on the Growth of S. putrefaciens in Pacific White Shrimp
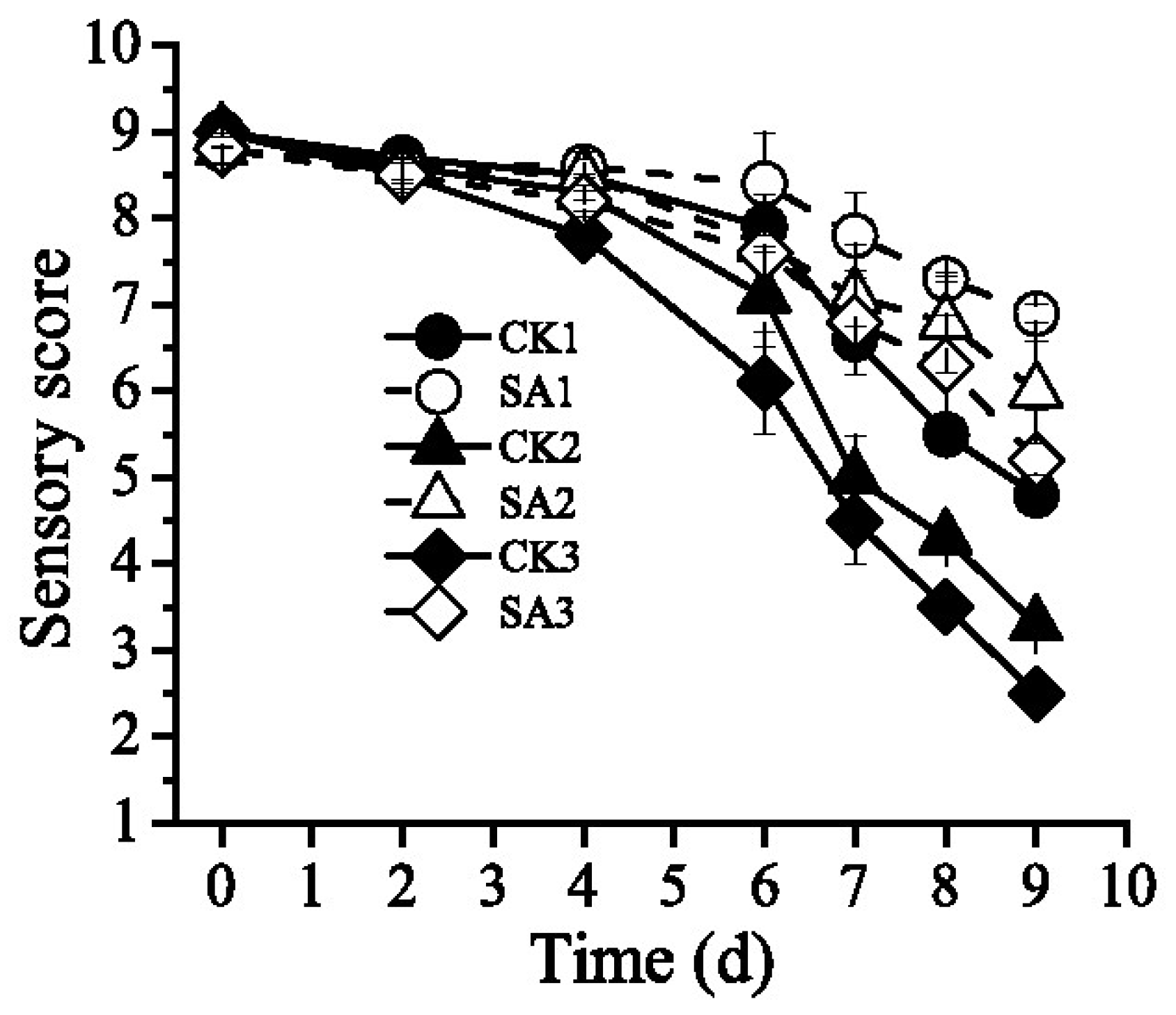
3.2. Effect of the CFS on Sensory Quality of Pacific White Shrimp
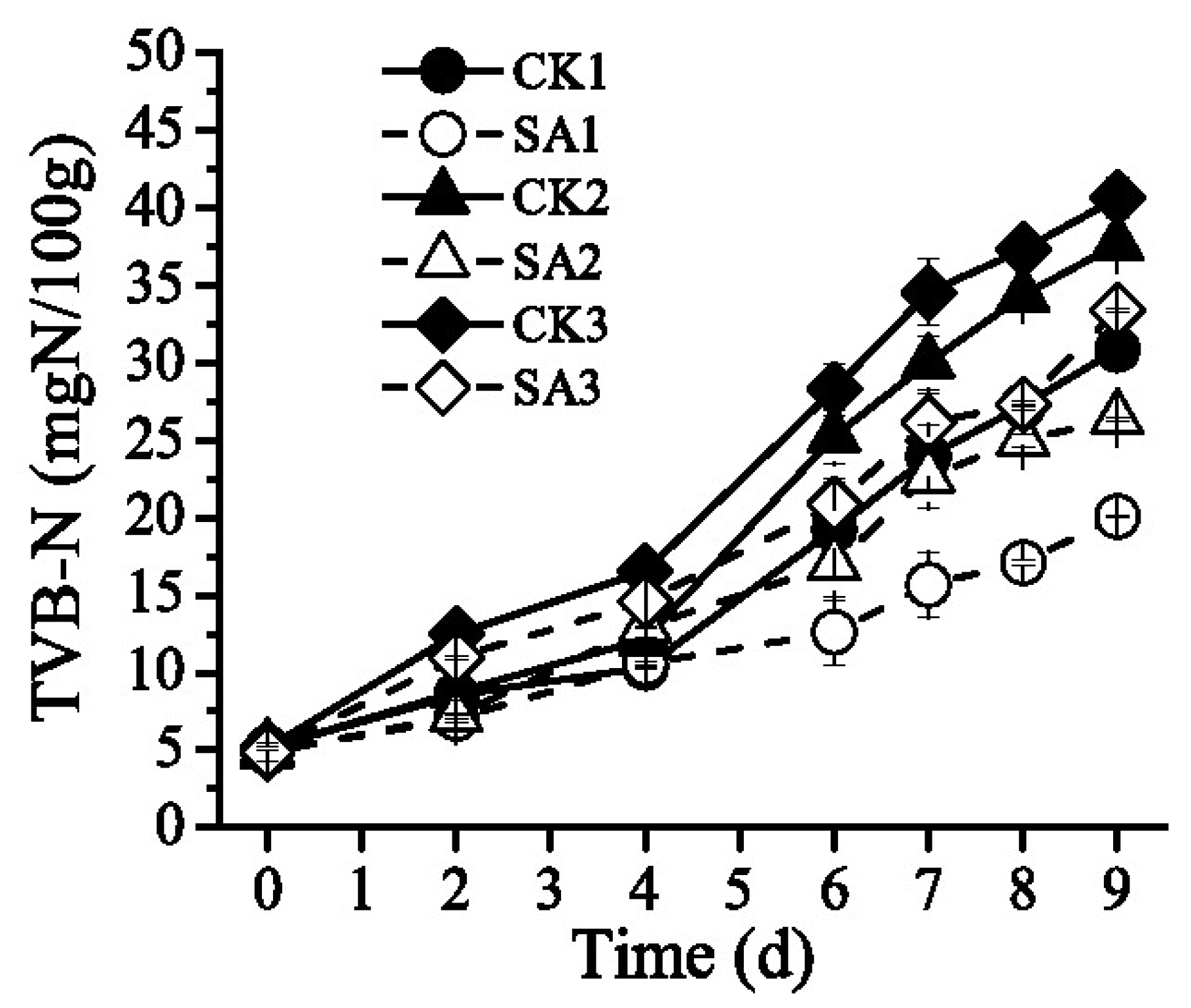
3.3. Effect of the CFS on TVB-N Content of Pacific White Shrimp
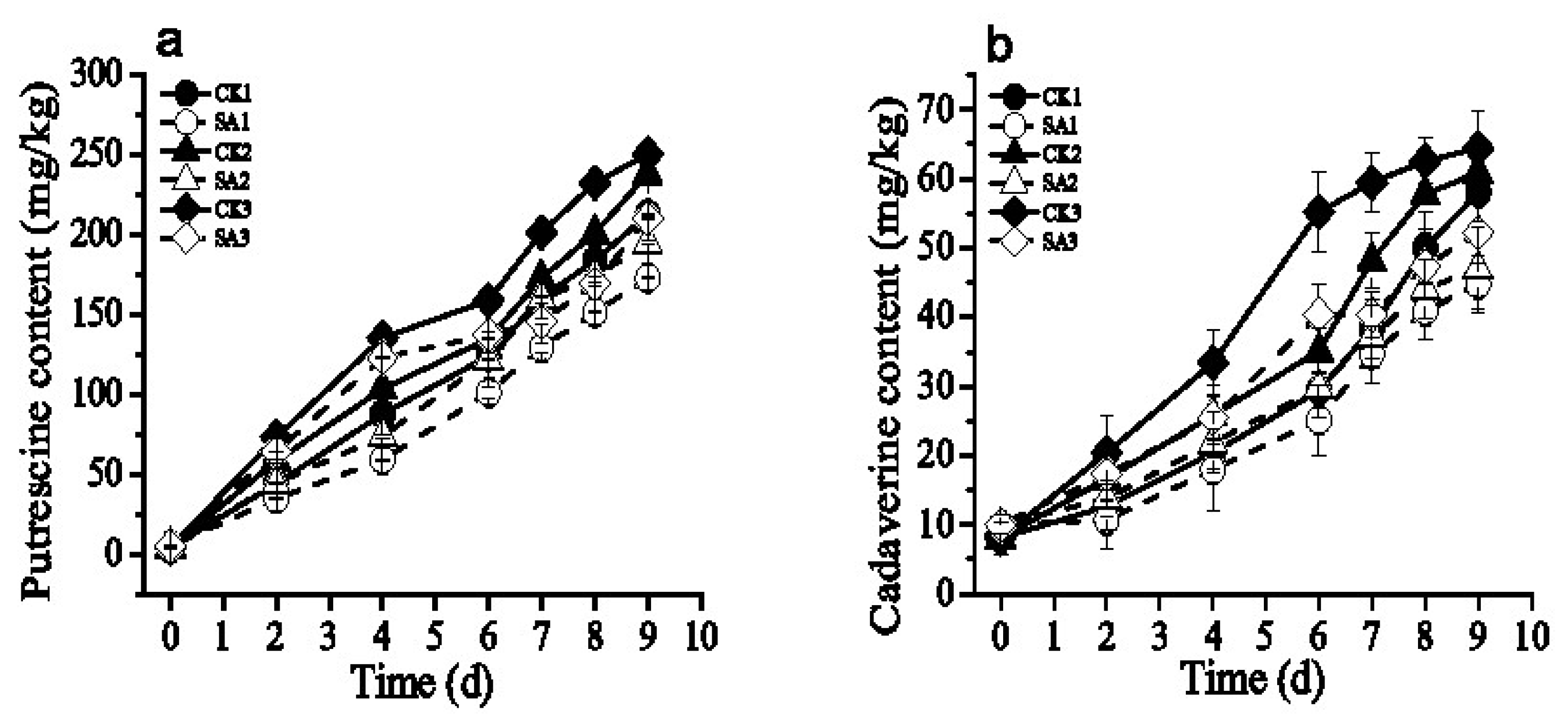
3.4. Effect of the CFS on Biogenic Amine Production of Pacific White Shrimp
3.5. Effect of the CFS on Amino Acid Content of Pacific White Shrimp
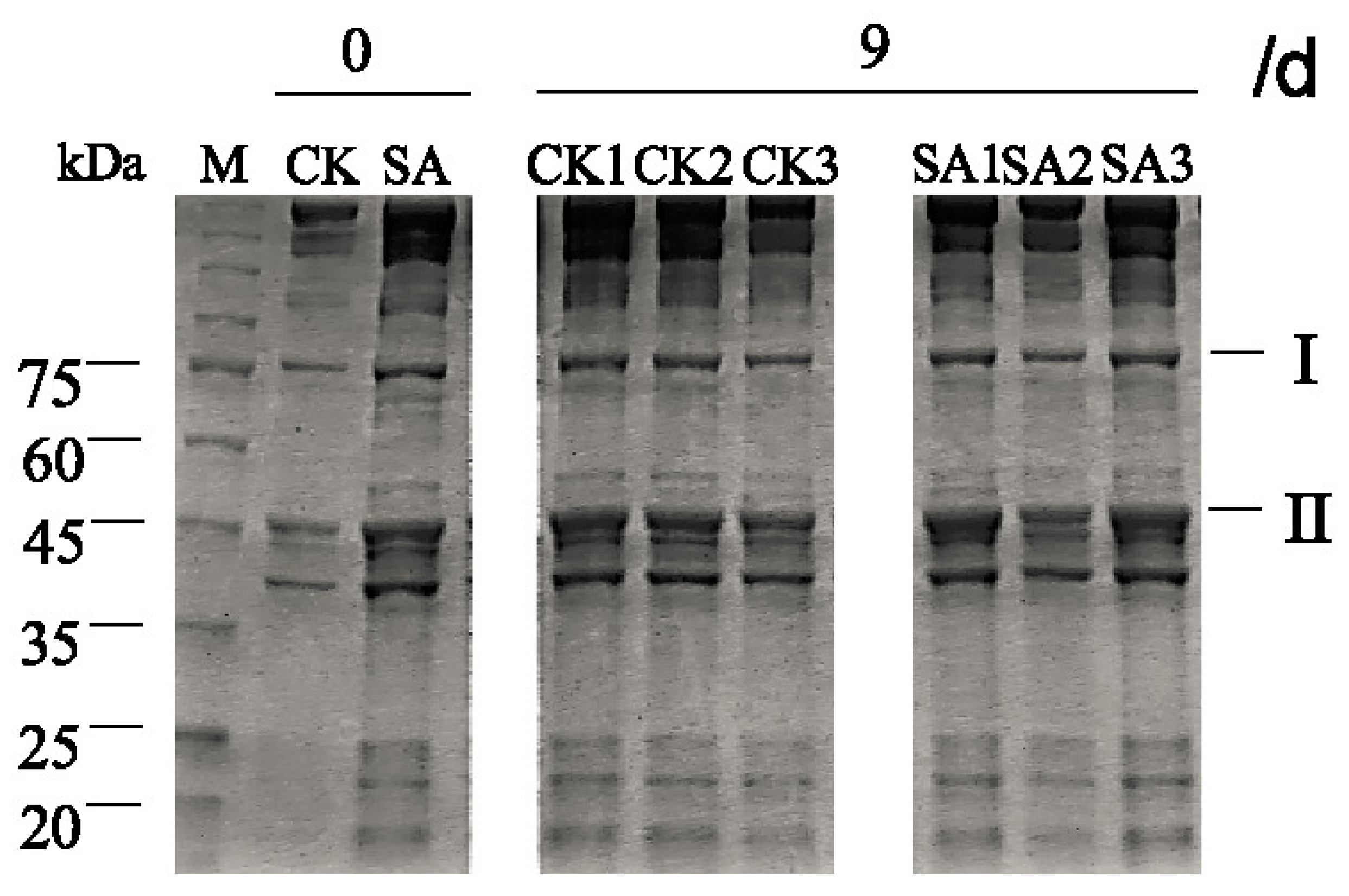
3.6. Effect of the CFS on the Sodium Dodecyl Sulfate-polyacrylamide Gel Electrophoresis (SDS-PAGE) Profile of the Myofibrillar Proteins of Pacific White Shrimp
4. Discussions
5. Conclusions
Author Contributions
Funding
Conflicts of Interest
References
- Jescovitch, L.N.; Ullman, C.; Rhodes, M.; Davis, D.A. Effects of different feed management treatments on water quality for Pacific white shrimp Litopenaeus vannamei. Aquac. Res. 2018, 49, 526–531. [Google Scholar] [CrossRef]
- Qian, Y.-F.; Xie, J.; Yang, S.-P.; Huang, S.; Wu, W.-H.; Li, L. Inhibitory effect of a quercetin-based soaking formulation and modified atmospheric packaging (MAP) on muscle degradation of Pacific white shrimp (Litopenaeus vannamei). LWT-Food Sci. Technol. 2015, 63, 1339–1346. [Google Scholar] [CrossRef]
- Okpala, C.O.R. Investigation of quality attributes of ice-stored Pacific white shrimp (Litopenaeus vannamei) as affected by sequential minimal ozone treatment. LWT-Food Sci. Technol. 2014, 57, 538–547. [Google Scholar] [CrossRef]
- Gram, L.; Huss, H.H. Microbiological spoilage of fish and fish products. Int. J. Food Microbiol. 1996, 33, 121–137. [Google Scholar] [CrossRef]
- Fall, P.-A.; Leroi, F.; Cardinal, M.; Chevalier, F.; Pilet, M.-F. Inhibition of Brochothrix thermosphacta and sensory improvement of tropical peeled cooked shrimp by Lactococcus piscium CNCM I-4031. Lett. Appl. Microbiol. 2010, 50, 357–361. [Google Scholar] [CrossRef]
- Dabadé, D.S.; den Besten, H.M.; Azokpota, P.; Nout, M.R.; Hounhouigan, D.J.; Zwietering, M.H. Spoilage evaluation, shelf-life prediction, and potential spoilage organisms of tropical brackish water shrimp (Penaeus notialis) at different storage temperatures. Food Microbiol. 2015, 48, 8–16. [Google Scholar] [CrossRef]
- Skandamis, P.N.; Nychas, G.-J.E. Quorum sensing in the context of food microbiology. Appl. Environ. Microbiol. 2012, 78, 5473–5482. [Google Scholar] [CrossRef]
- Ginson, J.; Panda, S.K.; Bindu, J.; Kamalakanth, C.K.; Gopal, T.K.S. Effect of high pressure treatment on microbiological quality of Indian white prawn (Fenneropenaeus indicus) during chilled storage. Food Microbiol. 2015, 46, 596–603. [Google Scholar] [CrossRef]
- Yang, S.-P.; Xie, J.; Qian, Y.-F. Determination of spoilage microbiota of Pacific white shrimp during ambient and cold storage using next-generation sequencing and culture-dependent method. J. Food Sci. 2017, 82, 1178–1183. [Google Scholar] [CrossRef]
- Gram, L.; Dalgaard, P. Fish spoilage bacteria—Problems and solutions. Curr. Opin. Biotechnol. 2002, 13, 262–266. [Google Scholar] [CrossRef]
- Fan, H.; Luo, Y.; Yin, X.; Bao, Y.; Feng, L. Biogenic amine and quality changes in lightly salt-and sugar-salted black carp (Mylopharyngodon piceus) fillets stored at 4 °C. Food Chem. 2014, 159, 20–28. [Google Scholar] [CrossRef]
- Zhao, A.; Zhu, J.; Ye, X.; Ge, Y.; Li, J. Inhibition of biofilm development and spoilage potential of Shewanella baltica by quorum sensing signal in cell-free supernatant from Pseudomonas fluorescens. Int. J. Food Microbiol. 2016, 230, 73–80. [Google Scholar] [CrossRef]
- Jorgensen, L.V.; Dalgaard, P.; Huss, H.H. Multiple compound quality index for cold-smoked salmon (Salmo salar) developed by multivariate regression of biogenic amines and pH. J. Agric. Food Chem. 2000, 48, 2448–2453. [Google Scholar] [CrossRef]
- Macé, S.; Cardinal, M.; Jaffrès, E.; Cornet, J.; Lalanne, V.; Chevalier, F.; Sérot, T.; Pilet, M.-F.; Dousset, X.; Joffraud, J.-J. Evaluation of the spoilage potential of bacteria isolated from spoiled cooked whole tropical shrimp (Penaeus vannamei) stored under modified atmosphere packaging. Food Microbiol. 2014, 40, 9–17. [Google Scholar] [CrossRef]
- Laursen, B.G.; Leisner, J.J.; Dalgaard, P. Carnobacterium species: Effect of metabolic activity and interaction with Brochothrix thermosphacta on sensory characteristics of modified atmosphere packed shrimp. J. Agric. Food Chem. 2006, 54, 3604–3611. [Google Scholar] [CrossRef]
- Gui, M.; Wu, R.; Liu, L.; Wang, S.; Zhang, L.; Li, P. Effects of quorum quenching by AHL lactonase on AHLs, protease, motility and proteome patterns in Aeromonas veronii LP-11. Int. J. Food Microbiol. 2017, 252, 61–68. [Google Scholar] [CrossRef]
- Jaffres, E.; Lalanne, V.; Mace, S.; Cornet, J.; Cardinal, M.; Serot, T.; Dousset, X.; Joffraud, J.-J. Sensory characteristics of spoilage and volatile compounds associated with bacteria isolated from cooked and peeled tropical shrimps using SPME-GC-MS analysis. Int. J. Food Microbiol. 2011, 147, 195–202. [Google Scholar] [CrossRef]
- Yue, J.; Liu, L.; Li, Z.; Li, D.; Fu, Z. Improved quality analytical models for aquatic products at the transportation in the cold chain. Math. Comp. Model. Dyn. 2013, 58, 474–479. [Google Scholar] [CrossRef]
- Shi, C.; Yang, X.; Han, S.; Fan, B.; Zhao, Z.; Wu, X.; Qian, J. Nondestructive prediction of tilapia fillet freshness during storage at different temperatures by integrating an electronic nose and tongue with radial basis function neural networks. Food Bioprocess. Technol. 2018, 11, 1840–1852. [Google Scholar] [CrossRef]
- Zhang, Y.; Ertbjerg, P. Effects of frozen-then-chilled storage on proteolytic enzyme activity and water-holding capacity of pork loin. Meat Sci. 2018, 145, 375–382. [Google Scholar] [CrossRef]
- Surette, M.G.; Bassler, B.L. Quorum sensing in Escherichia coli and Salmonella typhimurium. Proc. Natl. Acad. Sci. USA 1998, 95, 7046–7050. [Google Scholar] [CrossRef]
- Qian, Y.-F.; Xie, J.; Yang, S.-P.; Wu, W.-H.; Xiong, Q.; Gao, Z.-L. In vivo study of spoilage bacteria on polyphenoloxidase activity and melanosis of modified atmosphere packaged Pacific white shrimp. Food Chem. 2014, 155, 126–131. [Google Scholar] [CrossRef]
- Jeyasekaran, G.; Ganesan, P.; Anandaraj, R.; Shakila, R.J.; Sukumar, D. Quantitative and qualitative studies on the bacteriological quality of Indian white shrimp (Penaeus indicus) stored in dry ice. Food Microbiol. 2006, 23, 526–533. [Google Scholar] [CrossRef]
- Qian, Y.-F.; Xie, J.; Yang, S.-P.; Wu, W.-H. Study of the quality changes and myofibrillar proteins of white shrimp (Litopenaeus vannamei) under modified atmosphere packaging with varying CO2 levels. Eur. Food Res. Technol. 2013, 236, 629–635. [Google Scholar] [CrossRef]
- Luzuriaga, D.A.; Korel, F.; Balaban, M.O. Odor evaluation of shrimp treated with different chemicals using an electronic nose and a sensory panel. J. Aquat. Food Prod. Technol. 2007, 16, 57–75. [Google Scholar] [CrossRef]
- Ikonic, P.; Tasic, T.; Petrovic, L.; Skaljac, S.; Jokanovic, M.; Mandic, A.; Ikonic, B. Proteolysis and biogenic amines formation during the ripening of Petrovska klobasa, traditional dry-fermented sausage from Northern Serbia. Food Control 2013, 30, 69–75. [Google Scholar] [CrossRef]
- Tasic, T.; Ikonic, P.; Mandic, A.; Jokanovic, M.; Tomovic, V.; Savatic, S.; Petrovic, L. Biogenic amines content in traditional dry fermented sausage Petrovska klobasa as possible indicator of good manufacturing practice. Food Control 2012, 23, 107–112. [Google Scholar] [CrossRef]
- Qian, Y.-F.; Ye, J.-X.; Yang, S.-P.; Lin, Z.-Q.; Cao, W.; Xie, J. Evaluation of the spoilage potential of Shewanella putrefaciens, Aeromonas hydrophila, and Aeromonas sobria isolated from spoiled Pacific white shrimp (Litopenaeus vannamei) during cold storage. J. Food Saf. 2018, 38, e12550. [Google Scholar] [CrossRef]
- Deng, Y.; Luo, Y.; Wang, Y.; Zhao, Y. Effect of different drying methods on the myosin structure, amino acid composition, protein digestibility and volatile profile of squid fillets. Food Chem. 2015, 171, 168–176. [Google Scholar] [CrossRef]
- Manju, S.; Jose, L.; Gopal, T.K.S.; Ravishankar, C.N.; Lalitha, K.V. Effects of sodium acetate dip treatment and vacuum-packaging on chemical, microbiological, textural and sensory changes of Pearlspot (Etroplus suratensis) during chill storage. Food Chem. 2007, 102, 27–35. [Google Scholar] [CrossRef]
- Ozogul, Y.; Durmus, M.; Ucar, Y.; Ozogul, F.; Regenstein, J.M. Comparative study of nanoemulsions based on commercial oils (sunflower, canola, corn, olive, soybean, and hazelnut oils): Effect on microbial, sensory, and chemical qualities of refrigerated farmed sea bass. Innov. Food Sci. Emerg. Technol. 2016, 33, 422–430. [Google Scholar] [CrossRef]
- Coffino, P. Regulation of cellular polyamines by antizyme. Nat. Rev. Mol. Cell Biol. 2001, 2, 188–194. [Google Scholar] [CrossRef]
- Ruiz-Capillas, C.; Moral, A. Free amino acids in muscle of Norway lobster (Nephrops novergicus (L.)) in controlled and modified atmospheres during chilled storage. Food Chem. 2004, 86, 85–91. [Google Scholar] [CrossRef]

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Time (days) | Microbiology Analysis | |||||
|---|---|---|---|---|---|---|
| CK1 | SA1 | CK2 | SA2 | CK3 | SA3 | |
| 0 | 4.29 ± 0.02 a | 4.02 ± 0.02 a | 4.29 ± 0.03 a | 4.02 ± 0.02 a | 4.29 ± 0.03 a | 4.02 ± 0.02 a |
| 2 | 4.68 ± 0.07 a | 4.53 ± 0.01 a | 5.07 ± 0.06 a | 4.90 ± 0.03 a | 5.48 ± 0.03 a | 5.35 ± 0.02 a |
| 4 | 5.23 ± 0.05 d | 5.18 ± 0.04e | 6.11 ± 0.05 b | 5.89 ± 0.01 c | 6.29 ± 0.04 a | 6.10 ± 0.04 b c |
| 6 | 6.24 ± 0.03 d | 5.51 ± 0.03 f | 6.83 ± 0.06 b | 5.96 ± 0.02e | 7.37 ± 0.10 a | 6.71 ± 0.01 c |
| 7 | 6.50 ± 0.13 c | 5.57 ± 0.03 d | 7.02 ± 0.04 b | 6.79 ± 0.01 c | 7.90 ± 0.05 a | 6.83 ± 0.06 c |
| 8 | 7.01 ± 0.04 b | 6.25 ± 0.06 c | 7.17 ± 0.09 b | 6.90 ± 0.01 b | 8.08 ± 0.02 a | 7.23 ± 0.03 b |
| 9 | 7.35 ± 0.03 c | 6.03 ± 0.05 d | 7.59 ± 0.04 b c | 7.23 ± 0.02 c | 8.08 ± 0.01 a | 7.66 ± 0.04 b |
| Time(days) | Sensory Score | |||||
|---|---|---|---|---|---|---|
| CK1 | SA1 | CK2 | SA2 | CK3 | SA3 | |
| 0 | 9.0 ± 0.15 a | 8.8 ± 0.18 a | 9.0 ± 0.18 a | 8.8 ± 0.14 a | 9.0 ± 0.18 a | 8.8 ± 0.14 a |
| 2 | 8.7 ± 0.02 a | 8.6 ± 0.19 a | 8.6 ± 0.19 a | 8.6 ± 0.15 a | 8.5 ± 0.19 a | 8.5 ± 0.15 a |
| 4 | 8.5 ± 0.18 a | 8.6 ± 0.22 a | 8.3 ± 0.22 a | 8.5 ± 0.28 a | 7.8 ± 0.22 a,b | 8.2 ± 0.28 a |
| 6 | 7.9 ± 0.28 b | 8.4 ± 0.58 b | 7.1 ± 0.58 b | 7.7 ± 0.57 b | 6.1 ± 0.58 b,* | 7.6 ± 0.57 b |
| 7 | 6.6 ± 0.15 c | 7.8 ± 0.49 c | 5.0 ± 0.49 c | 7.1 ± 0.60 c | 4.5 ± 0.49 c | 6.8 ± 0.61 c |
| 8 | 5.5 ± 0.18 d | 7.3 ± 0.03 d | 4.3 ± 0.03 d | 6.8 ± 0.58 d | 3.5 ± 0.03 d | 6.3 ± 0.58 d |
| 9 | 4.8 ± 0.23 e | 6.9 ± 0.10 e | 3.3 ± 0.14 e | 6.0 ± 0.59 e | 2.5 ± 0.13 e | 5.2 ± 0.59 e |
| Time (days) | The Content of TVB-N | |||||
|---|---|---|---|---|---|---|
| CK1 | SA1 | CK2 | SA2 | CK3 | SA3 | |
| 0 | 5.13 ± 0.16 a | 4.86 ± 0.58 b | 5.13 ± 0.16 a | 4.86 ± 0.58 b | 5.13 ± 0.16 a | 4.86 ± 0.58 b |
| 2 | 8.56 ± 0.36 c | 7.05 ± 0.29 d | 8.76 ± 0.30 c | 7.21 ± 0.11 d | 12.48 ± 0.17 a | 10.97 ± 0.08 b |
| 4 | 10.34 ± 0.06 f | 10.575 ± 0.11 e | 12.17 ± 0.06 d | 12.975 ± 1.12 c | 16.54 ± 0.03 a | 14.61 ± 0.00 b |
| 6 | 19.26 ± 1.08 c,d | 12.59 ± 1.06 f | 25.27 ± 1.73 a,b | 16.98 ± 1.12 d,e | 28.26 ± 1.64 a | 20.90 ± 1.67 c |
| 9 | 30.87 ± 0.10 c,d | 20.09 ± 0.06 f | 37.67 ± 0.25 b | 26.41 ± 0.11 e | 40.67 ± 1.31 a | 33.41 ± 0.08 c |
| Time (days) | The Content of Putrescine | |||||
|---|---|---|---|---|---|---|
| CK1 | SA1 | CK2 | SA2 | CK3 | SA3 | |
| 9 | 212.72 ± 0.23 c | 173.04 ± 0.32 d,e | 238.19 ± 0.13 b | 194.81 ± 1.09 d | 250.56 ± 0.31 a | 210.34 ± 0.97 c |
| Time (days) | The Content of Cadaverine | |||||
|---|---|---|---|---|---|---|
| CK1 | SA1 | CK2 | SA2 | CK3 | SA3 | |
| 6 | 29.43 ± 1.92 c | 25.12 ± 3.01 d | 35.12 ± 1.21 c | 29.81 ± 0.23 c | 55.23 ± 3.70 a | 40.45 ± 2.32 b |
| 9 | 58.05 ± 2.23 b | 44.81 ± 2.1 c | 60.91 ± 1.56 a,b | 47.02 ± 3.89 c | 64.45 ± 3.34 a | 52.23 ± 2.44 b,c |
| Amino Acid | 0 day | 9 days | ||||||
|---|---|---|---|---|---|---|---|---|
| CK | SA | CK1 | CK2 | CK3 | SA1 | SA2 | SA3 | |
| Asp | 621.56 ± 0.18 | 645.59 ± 0.04 | 519.92 ± 0.43 | 479.02 ± 0.18 | 457.21 ± 0.12 | 587.89 ± 0.11 | 548.36 ± 0.48 | 516.02 ± 0.82 |
| Thr | 182.45 ± 0.53 | 163.15 ± 0.39 | 129.91 ± 0.24 | 114.47 ± 0.41 | 101.24 ± 0.26 | 156.50 ± 0.33 | 148.37 ± 0.08 | 130.11 ± 0.16 |
| Ser | 259.32 ± 0.36 | 264.18 ± 0.76 | 221.48 ± 0.22 | 212.30 ± 0.17 | 201.45 ± 0.36 | 244.06 ± 0.23 | 230.92 ± 0.11 | 226.12 ± 0.20 |
| Glu | 982.76 ± 0.08 | 992.48 ± 1.86 | 870.64 ± 0.24 | 829.4 ± 0.04 | 810.81 ± 0.17 | 891.34 ± 0.07 | 869.34 ± 0.89 | 856.17 ± 0.08 |
| Gly | 351.49 ± 0.74 | 369.01 ± 0.01 | 312.10 ± 0.26 | 289.35 ± 0.58 | 271.78 ± 0.37 | 331.35 ± 0.76 | 326.42 ± 0.14 | 310.45 ± 0.23 |
| Ala | 498.39 ± 0.67 | 480.02 ± 0.07 | 441.31 ± 0.04 | 353.50 ± 0.18 | 333.41 ± 0.32 | 401.34 ± 0.53 | 391.39 ± 0.11 | 370.02 ± 0.20 |
| Val | 129.18 ± 0.82 | 141.39 ± 0.74 | 131.93 ± 0.45 | 129.71 ± 0.36 | 127.40 ± 0.14 | 113.04 ± 0.33 | 108.35 ± 0.26 | 105.09 ± 0.33 |
| Met | 181.03 ± 0.31 | 167.14 ± 0.17 | 149.45 ± 0.57 | 135.30 ± 0.48 | 132.37 ± 0.23 | 131.91 ± 0.72 | 129.49 ± 0.27 | 126.34 ± 0.82 |
| Ile | 94.34 ± 0.33 | 95.01 ± 0.36 | 92.40 ± 0.25 | 69.32 ± 0.20 | 79.01 ± 0.26 | 83.91 ± 0.81 | 78.81 ± 0.14 | 80.02 ± 0.09 |
| Leu | 430.61 ± 0.24 | 451.88 ± 0.04 | 323.82 ± 0.16 | 300.45 ± 0.45 | 278.30 ± 0.51 | 389.01 ± 0.49 | 357.38 ± 0.53 | 329.71 ± 0.81 |
| Tyr | 221.91 ± 0.14 | 239.07 ± 0.32 | 170.51 ± 0.14 | 167.01 ± 0.39 | 140.40 ± 0.32 | 189.39 ± 0.81 | 179.04 ± 0.30 | 161.34 ± 0.41 |
| Phe | 231.94 ± 0.14 | 241.06 ± 0.23 | 170.32 ± 0.29 | 169.96 ± 0.10 | 140.92 ± 0.14 | 183.92 ± 0.32 | 177.34 ± 0.42 | 165.80 ± 0.88 |
| Lys | 460.73 ± 0.76 | 453.74 ± 0.29 | 395.28 ± 0.41 a | 367.51 ± 0.26 c | 338.91 ± 0.76 d | 401.49 ± 0.49 a | 394.20 ± 0.14 a | 378.91 ± 0.72 b |
| His | 113.01 ± 0.54 | 120.19 ± 0.48 | 110.49 ± 0.23 | 90.91 ± 0.48 | 80.81 ± 0.81 | 103.45 ± 0.07 | 94.35 ± 0.67 | 87.01 ± 0.81 |
| Arg | 505.51 ± 0.49 | 490.30 ± 0.14 | 437.18 ± 0.32 b | 412.19 ± 0.14 c | 389.30 ± 0.27 d | 476.02 ± 0.14 a | 434.35 ± 0.36 b | 419.45 ± 0.68 c |
| Total | 5264.23 | 4834.19 | 4476.74 | 3985.1 | 3883..32 | 4684.62 | 4468.11 | 4262.56 |
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Ye, J.-X.; Yang, S.-P.; Qian, Y.-F.; Xie, J. Effect of Cell-Free Supernatant from Aeromonas sobria on the Spoilage of Shewanella putrefaciens in Pacific White Shrimp (Litopenaeus vannamei) with the Influence of Temperature Fluctuation. Appl. Sci. 2019, 9, 587. https://doi.org/10.3390/app9030587
Ye J-X, Yang S-P, Qian Y-F, Xie J. Effect of Cell-Free Supernatant from Aeromonas sobria on the Spoilage of Shewanella putrefaciens in Pacific White Shrimp (Litopenaeus vannamei) with the Influence of Temperature Fluctuation. Applied Sciences. 2019; 9(3):587. https://doi.org/10.3390/app9030587
Chicago/Turabian StyleYe, Jing-Xin, Sheng-Ping Yang, Yun-Fang Qian, and Jing Xie. 2019. "Effect of Cell-Free Supernatant from Aeromonas sobria on the Spoilage of Shewanella putrefaciens in Pacific White Shrimp (Litopenaeus vannamei) with the Influence of Temperature Fluctuation" Applied Sciences 9, no. 3: 587. https://doi.org/10.3390/app9030587
APA StyleYe, J.-X., Yang, S.-P., Qian, Y.-F., & Xie, J. (2019). Effect of Cell-Free Supernatant from Aeromonas sobria on the Spoilage of Shewanella putrefaciens in Pacific White Shrimp (Litopenaeus vannamei) with the Influence of Temperature Fluctuation. Applied Sciences, 9(3), 587. https://doi.org/10.3390/app9030587