1. Introduction
In today’s world, Autonomous Underwater Vehicles (AUVs) are implemented for a variety of tasks, including ocean surveys, mine clearance and data collection in the ocean and river environments [
1]. It is a well known fact that, in nature, the fish propels itself by the coordinate motion of its body, fins, and tail, achieving higher propulsive efficiency and better maneuverability than the conventional AUVs powered by rotary propellers with the same power consumption [
2]. To improve the efficiency of AUVs, more research studies are performedthe research on application, design, and control of underwater robotic fish in recent years [
3,
4,
5,
6,
7,
8,
9].
A single robot with multiple capabilities cannot necessarily accomplish a complicated task successfully, an intended job whereas a robot swarm in which each robot has its own functions, can be more flexible, robust, and cost-effective and do complex tasks [
10]. Recent studies [
11,
12,
13,
14,
15] show that robotic fish formation can improve swimming efficiency. However, in the underwater environment, the velocity and viscosity of fluids, the complex geometry environment condition and even the interaction between robots can affect the stability of underwater robots formation. It is necessary for a robotic fish in a formation to get basic environmental information around itself, such as the inlet flow velocity and the spacing between itself and other robotic fish, which are useful for stable formation. With an underwater robot formation, unlike air or land robots, it is difficult to obtain the information through traditional means such as communication, GPS, and acoustic or visual systems [
1,
16].
Fish have a unique sensory system, lateral line system (LLS), which helps them sense fluid changes and the spacing between themselves and other objects. It has a great significance for predation, avoiding enemies, breeding and formation swimming [
17,
18,
19,
20]. In recent years, researchers try to incorporate LLS into engineering applications as an alternative method for sensing the fluid environment around AUV. Venturelli et al. [
21] adopted laterally distributed parallel pressure sensor arrays and presented methods for the analysis of the surrounding steady and unsteady flows to provide a simplistic mimic of the fish’s posterior lateral line organ, which can extract control-related information. Yanagitsuru et al. [
22] used pressure sensors to detect flow parameters (flow speed, vortex shedding frequency, and cylinder diameter), and the results provide insight into the sensory ecology of schools of fish and give implications for the design of autonomous underwater vehicles. Inspired by the lateral line of fish, Tang et al. [
23] proposed a new kind of near-field detection system to be applied to detect impending walls, approaching obstacles and other target objects. Nawi et al. [
24] could detect the moving objects located in front of the sensor by an artificial lateral line flow sensor.
Our work is inspired by the LLS of fish and we try to construct the predictive model of flow velocity and the judgement model of spacing for robotic fish formation based on the monitoring sensors attached on the robotic fish surface. To get enough monitoring data for analysis, a large number of experiments should be conducted with different physical conditions. At the same time, eigenvalues should be sorted out to improve the data quality. In traditional physics experiments, the data monitored by sensors are difficult to collect, and the errors caused by instruments and materials are inevitable. In addition, some specific conditions are difficult to achieve in real physical experiments. Therefore, we use the numerical simulation method based on Computational Fluid Dynamics (CFD) in this work.
Zhou et al. [
25] used CFD methods to investigate the hydrodynamics of bio-inspired fish swimming, especially the spatial distribution and temporal variation of the near-body pressure of fish are studied. They also provided an algorithm to predict the inlet flow velocity by using the near-body pressure at distributed spatial points. Lin et al. [
26] numerically studied the hydrodynamics of AUV at different speeds and analyzed the spatial distribution of the near-body pressure over the whole computational domain. Husaini et al. [
27] studied the drag variation by choosing two position arrangement of cooperative AUV. Jagadeesh [
28] presented a comparative assessment of the variation of drag characteristics over cooperative axisymmetric bodies at zero angle of attack. However, these original studies are all based on a single entity or the conventional AUVs formation. In robotic fish formation, the disturbance of flow caused by the swing of fish affect the data measured by sensors. It is necessary to study the perception of robotic fish formation further. Therefore, this paper studies the relationship between the pressure of fish surface and the inlet flow velocity, the spacing between individual members in simple fish formations. To be specific, the main contributions are summarized as follows:
We construct the models of basic environment perception for different robotic fish formations. Two specific formations, tandem and phalanx, are discussed in detail. We analyze the hydrodynamic phenomena of through the flow fields, including the wake effects in the tandem formation and the pressure distribution in the phalanx formation. For the tandem formation, the robot swimming in the front will generate flow in the wake, thus the fish swimming downstream will be affected by the unsteady flow. For the phalanx configuration, the effect of the lateral flow oscillating will be considered.
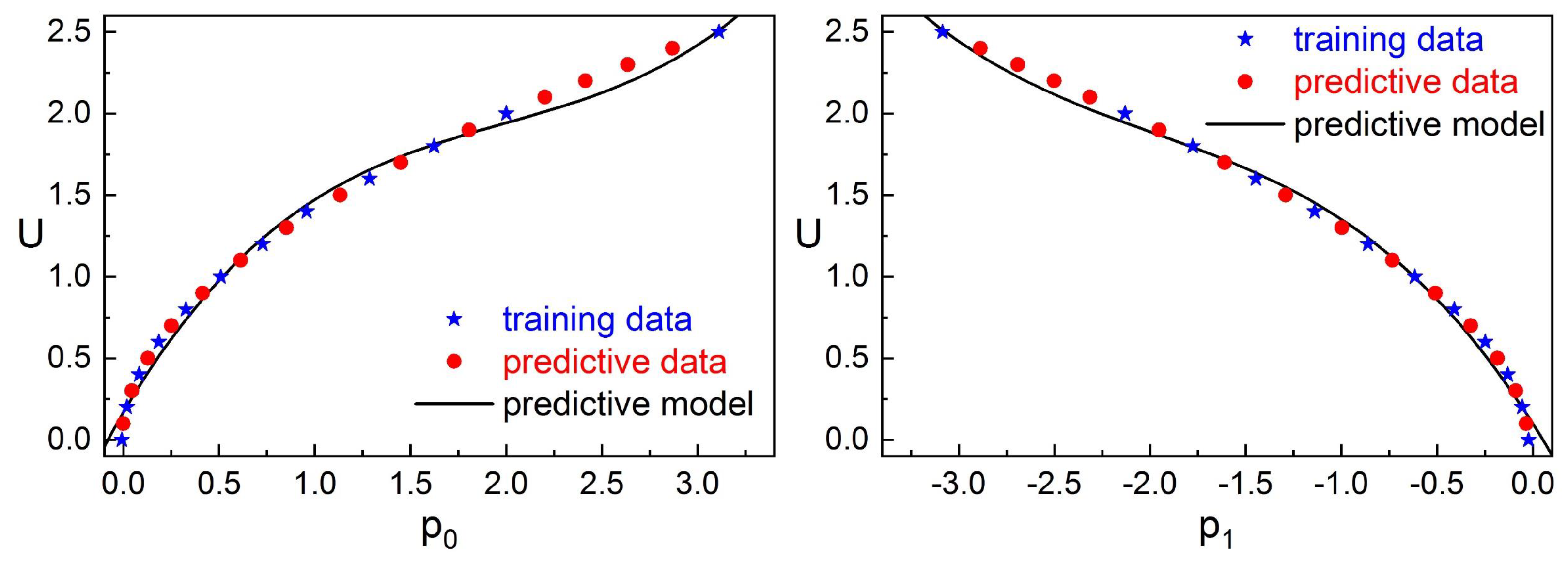
We adopt machine learning methods to generate our models using the simulation results as the sample data. Specifically, the predictive model of the flow velocity is constructed by polynomial fitting and the judgement model of spacing between robotic fish platforms is developed based on the method of neural networks. Our predictive models could reduce the error to , and the spacing judgement accuracy could reach at least 80%.
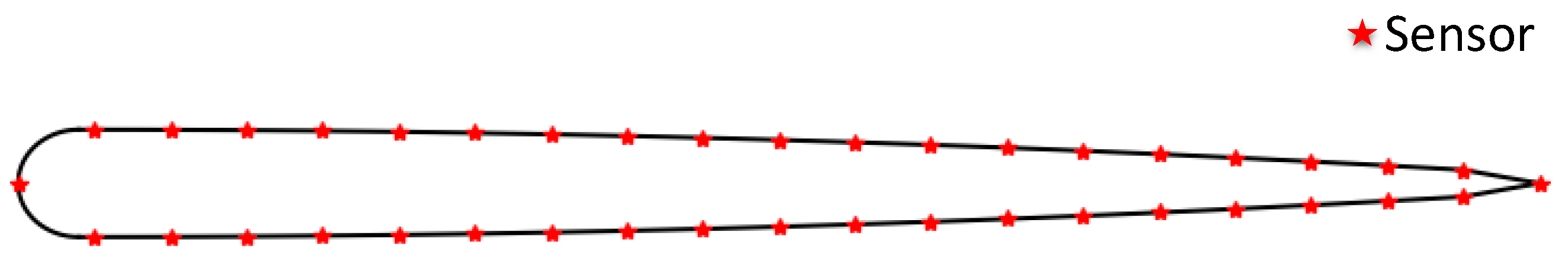
We analyze the data monitored by sensors at different positions and find that sensors located from the head to about one-third point of the front body are more useful, while the data monitored by sensors near the tail is unstable due to the large swing amplitude at the latter part of the fish body.
[In this study, we present simulations the robotic fish usingWe outline and simulate the robotic fish based on the open-source software OpenFOAM [
29,
30,
31], and construct the mathematical models by the methods of polynomial fitting and neural networks. The rest of the paper is organized as follows.
Section 2 describes the kinematic and geometry models of robotic fish, and the CFD numerical method.
Section 3 constructs the predictive model of the inlet flow velocity for single robotic fish.
Section 4 constructs the predictive models of the inlet flow velocity for different formations separately.
Section 5 gives the judgement models of the spacing between individual platforms for different formations separately. Conclusions are drawn in the final section.
5. The Spacing Judgement of Two Robotic Fishes
In this section, we judge the spacing between two robotic fishes by the kinematic pressures of the fish surface. Our purpose is to determine the relationship between the current spacing and the presetting spacing
. We only need to know whether the current spacing is greater or less than the presetting spacing
, but we do not need to know the exact value of the current spacing. Therefore, we choose neural networks to binary classification and construct the judgement model. The simulated experiment settings are the same as
Section 4. The tandem formation and phalanx formation have different phenomena, thus we discuss the tandem formation and phalanx formation separately.
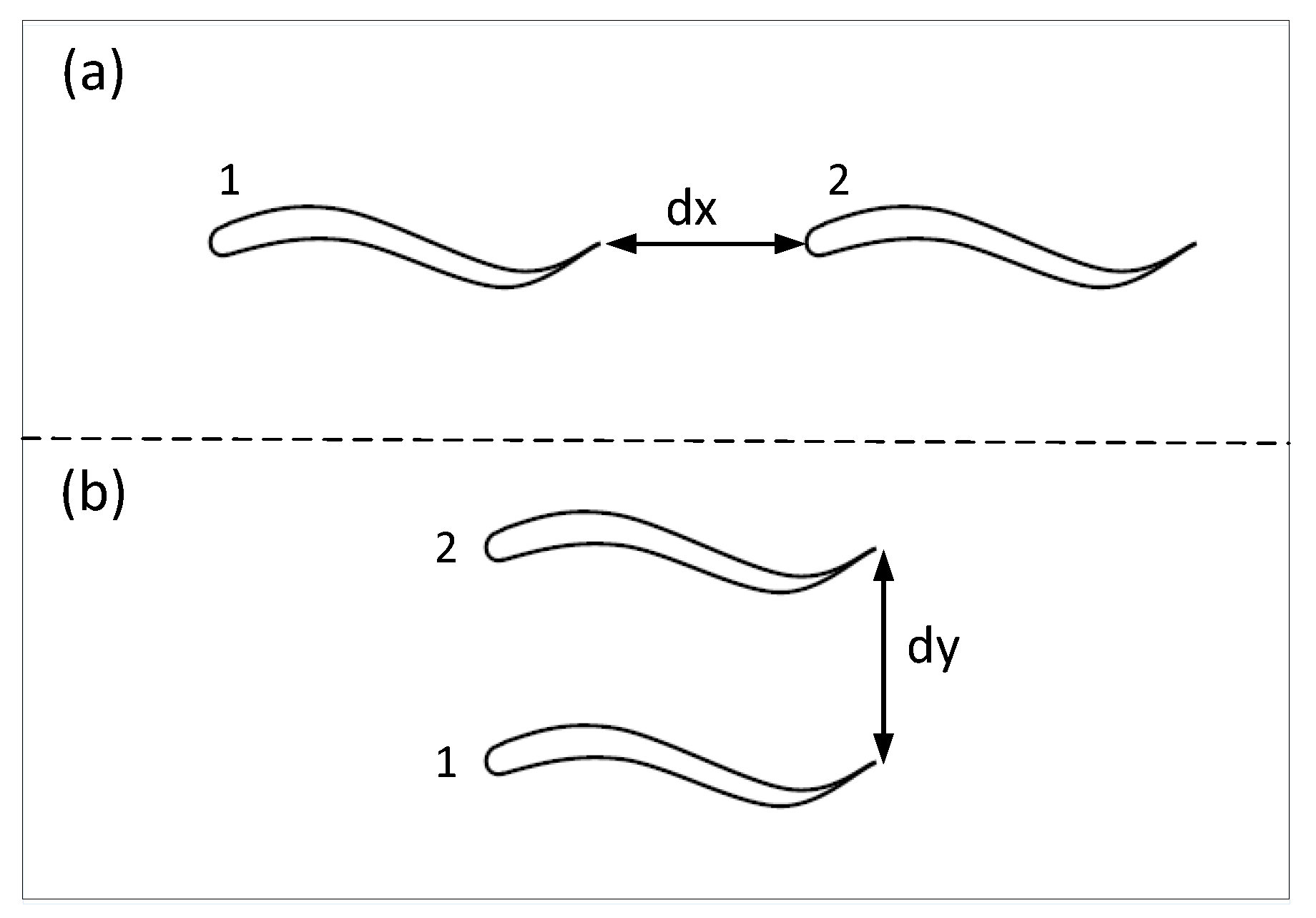
5.1. The Tandem Formation
In the tandem formation, the fish in the front (No. 1) does not need to consider the spacing between two fishes. To avoid collision and maintain the appropriate spacing in the formation, the fish at the back (No. 2) needs to judge the spacing between the two fishes. Thus, we are concerned about the kinematic pressures on No. 2 fish surface and judge the relationship between the current spacing and the presetting appropriate spacing by the kinematic pressure on No. 2 fish surface.
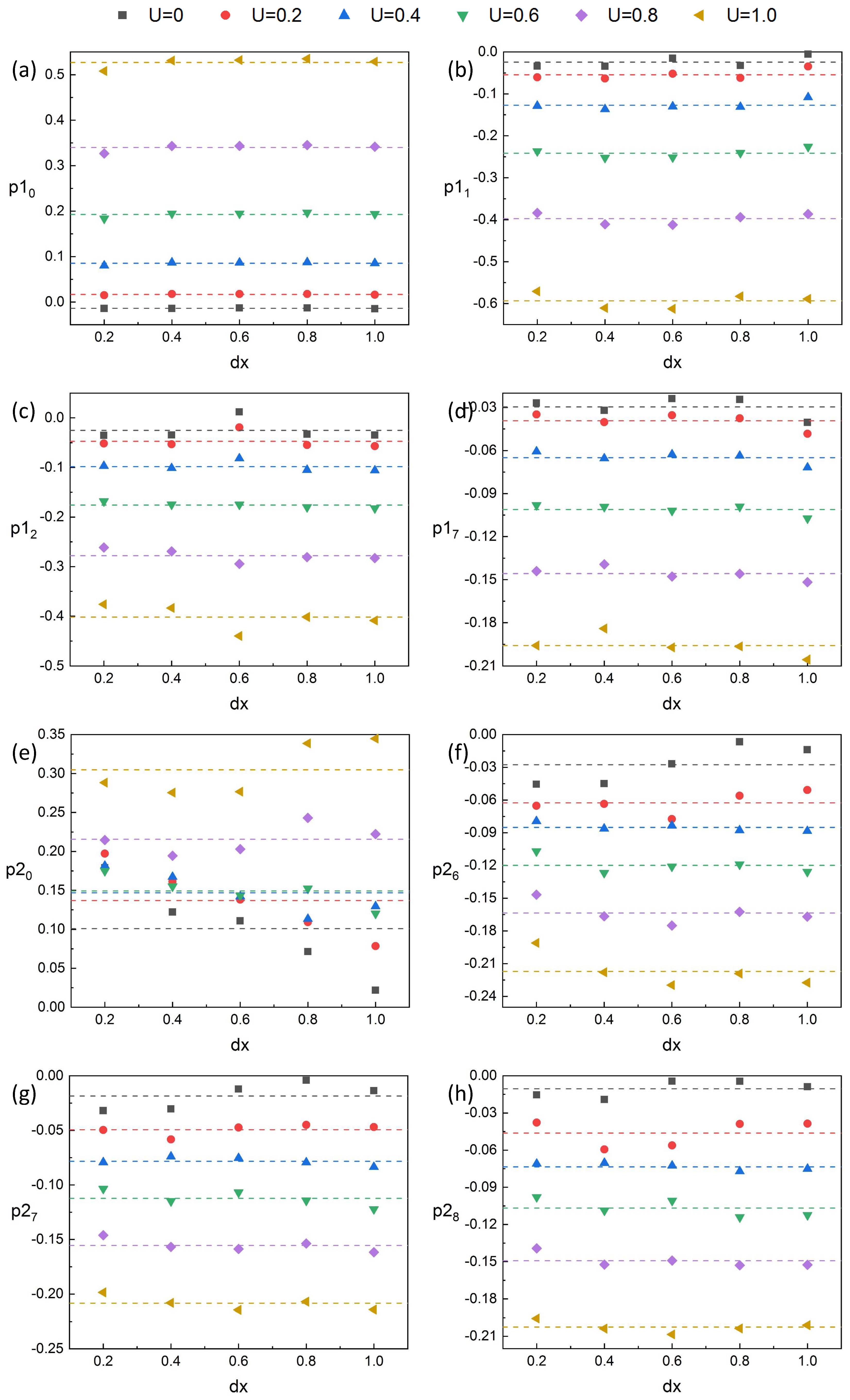
5.1.1. The Results Analysis
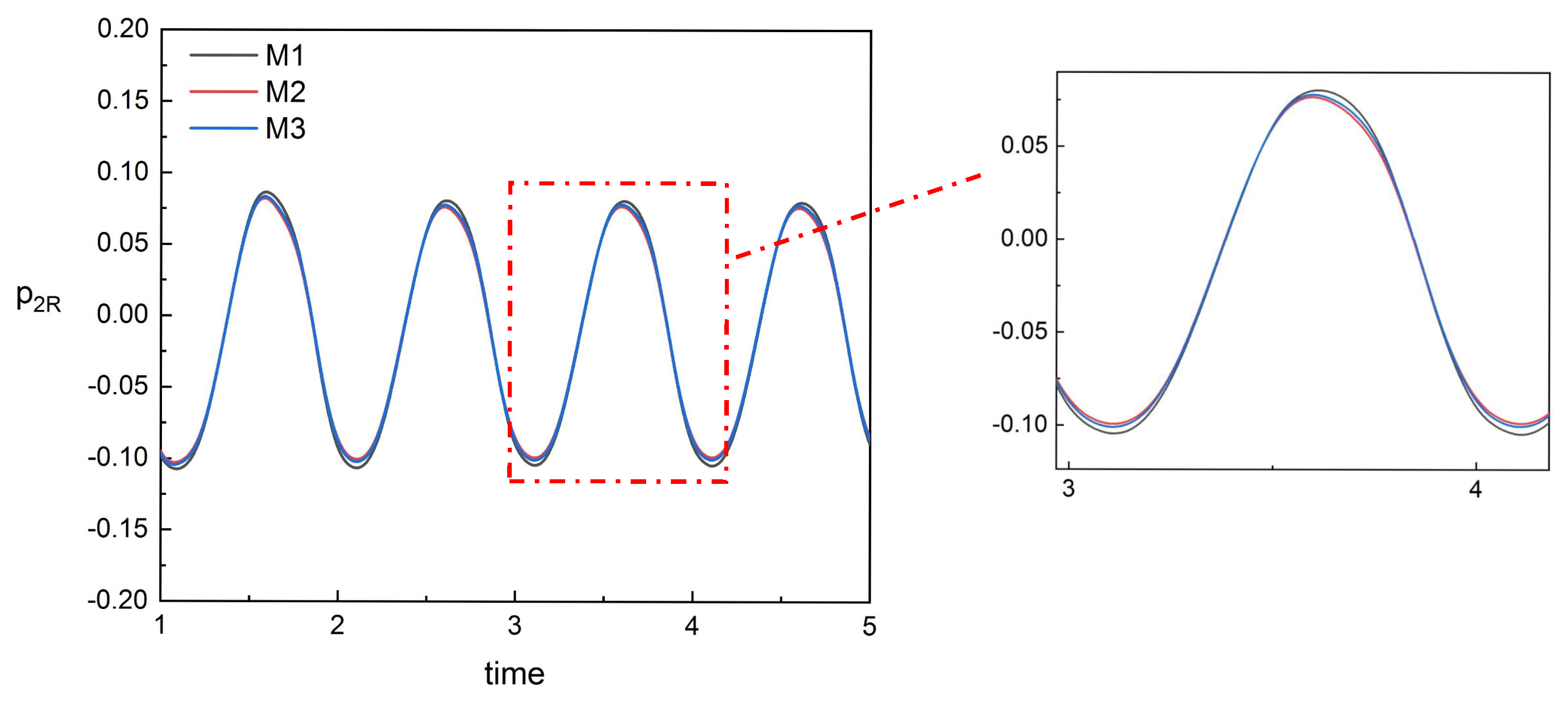
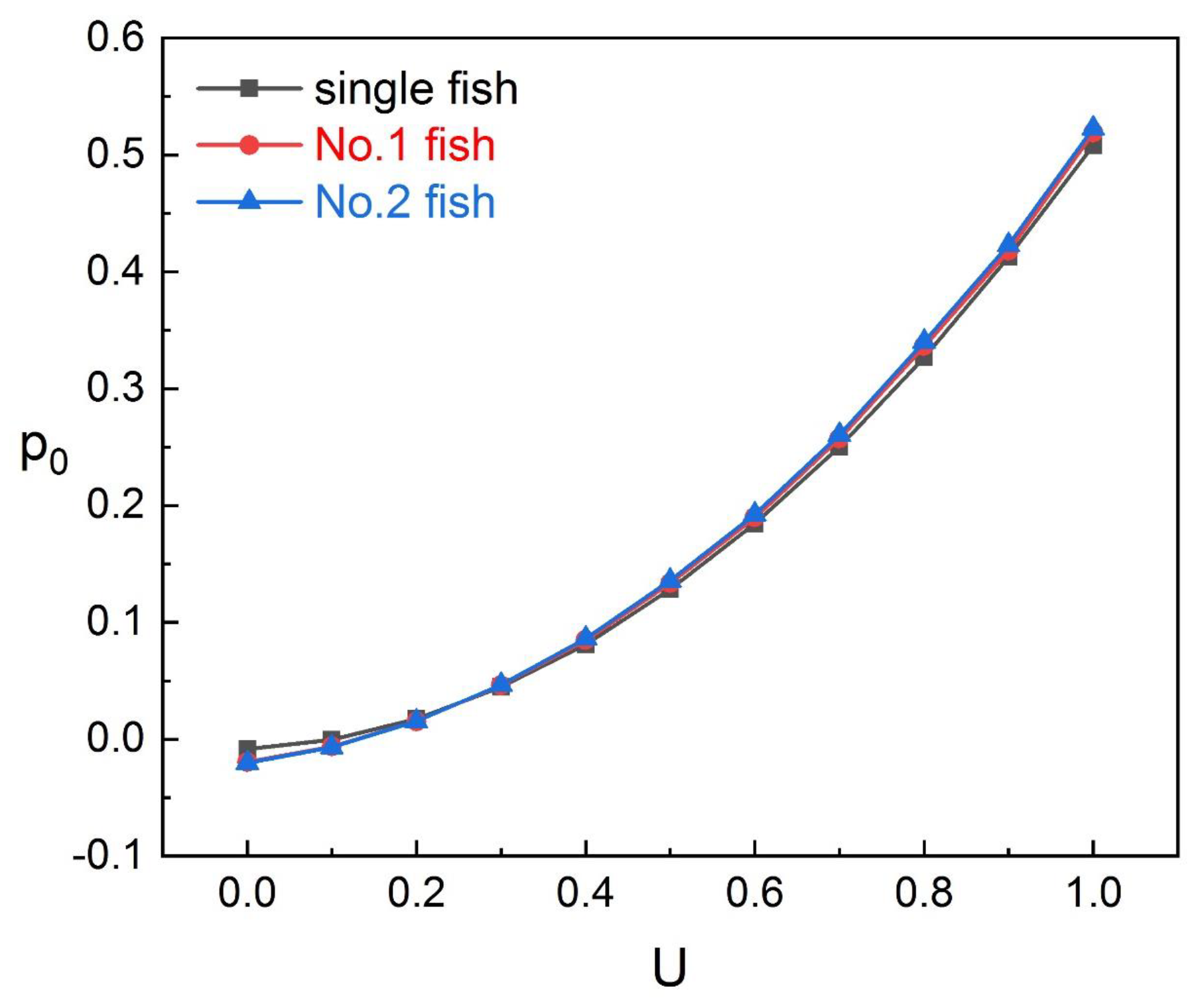
As presented in
Section 4, we found that, for No. 2 fish in the tandem, the data monitored by sensor
are less affected by spacing and we used
to construct the predictive model of inlet flow velocity for No. 2 fish. Since the wake of No. 1 fish mainly affects the head of No. 2 fish and its surroundings, the data monitored by sensors after
are almost unaffected by spacing. The head of No. 2 fish is directly impacted by the wake. The data monitored by the sensor here (
) are noisy and inaccurate, as shown on the left of
Figure 16. We chose the data monitored by sensors
to judge the spacing. This position can be affected by the wake, and, because of the swing of No. 2 fish itself, the impact of the wake is not as severe as that of the head and the data monitored by these sensors
are stable, as shown on the right of
Figure 16, which are suitable for judging the spacing.
5.1.2. The Construction and Validation of Judgement Models
Through the results on right side of
Figure 16, we found that, at the different inlet flow velocity
U, the relationship between
and the spacing
is different. Thus, we also need to use the kinematic pressure
, which is insensitive to the spacing as a feature for eliminating the effect of inlet flow velocity. In this work, for the presetting appropriate spacing
, we divide the data into two categories: the label is equal to 0 when the current spacing
is less than
, and the label is equal to 1 when the current spacing
is greater than
. We construct a binary classification model by using the method of neural networks [
44,
45]. The model can be used to judge the relationship between the current spacing
and the presetting appropriate spacing
. It has one input layer, two hidden layers and one output layer, as shown in
Figure 17. The input layer contains two features,
and
. Each hidden layer has 128 neurons and the activation function is Rectified Linear Unit (ReLU). The output layer is the result of classification, which is equal to 0 or 1 and the activation function is Sigmoid. The loss function of the model is binary_crossentropy, and the optimizer is Adam. We used the data simulated by settings in
Table 4 to train the classification model, and used the data simulated by settings in
Table 6 to validate the model. We used several
to construct the judgement models, and the effects are shown in
Table 9.
Through the above neural network structure, we can train a judgement model to compare the current spacing with any in a certain range.
5.2. The Phalanx Formation
In the phalanx formation, to avoid collision and maintain the appropriate spacing in the formation, the fish, whether on the left (No. 1) or on the right (No. 2), needs to know the spacing between the two fishes. We are concerned about the kinematic pressure on each fish surface and judge the relationship between the current spacing and the presetting appropriate spacing by the kinematic pressure on each fish surface.
5.2.1. The Results Analysis
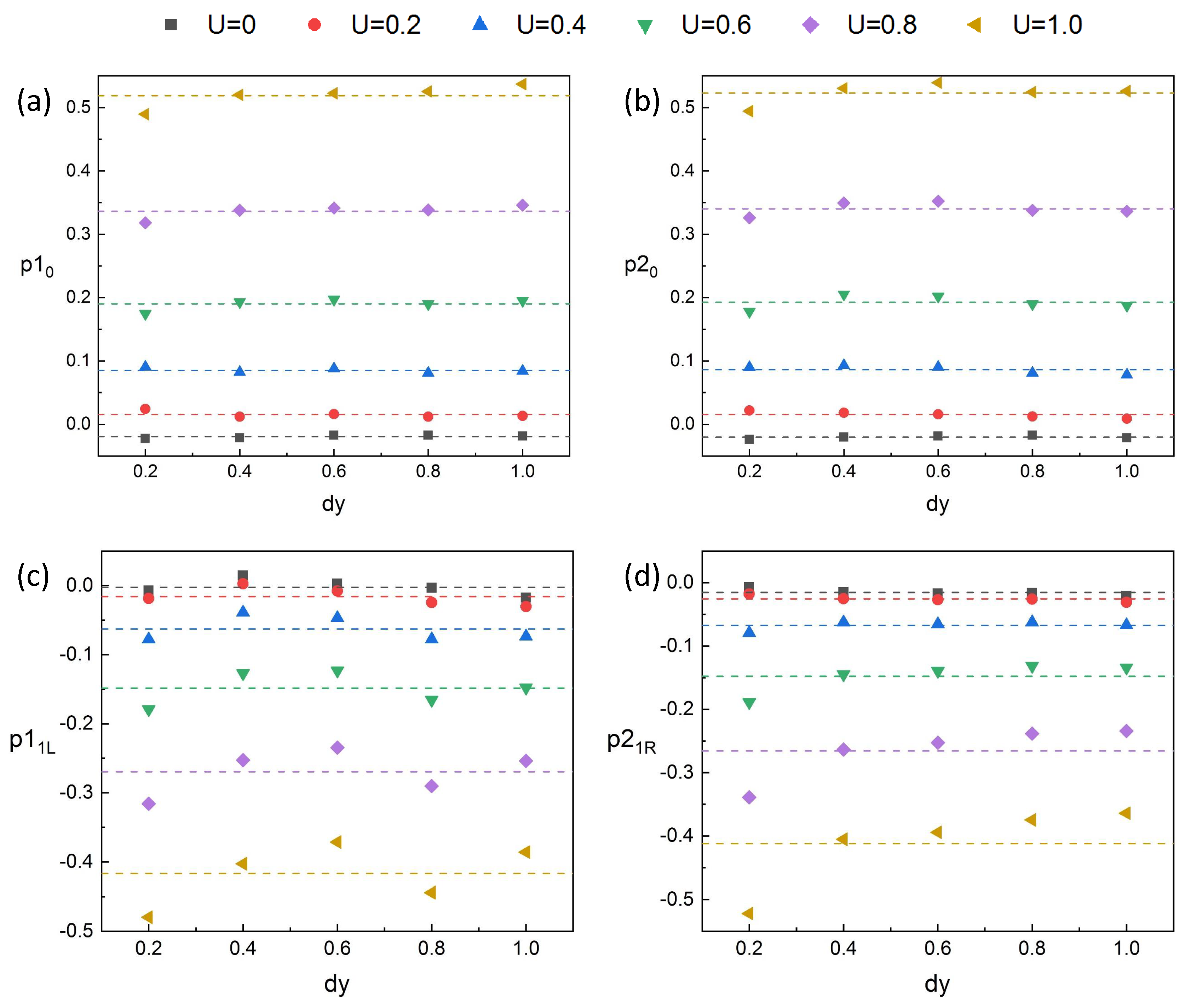
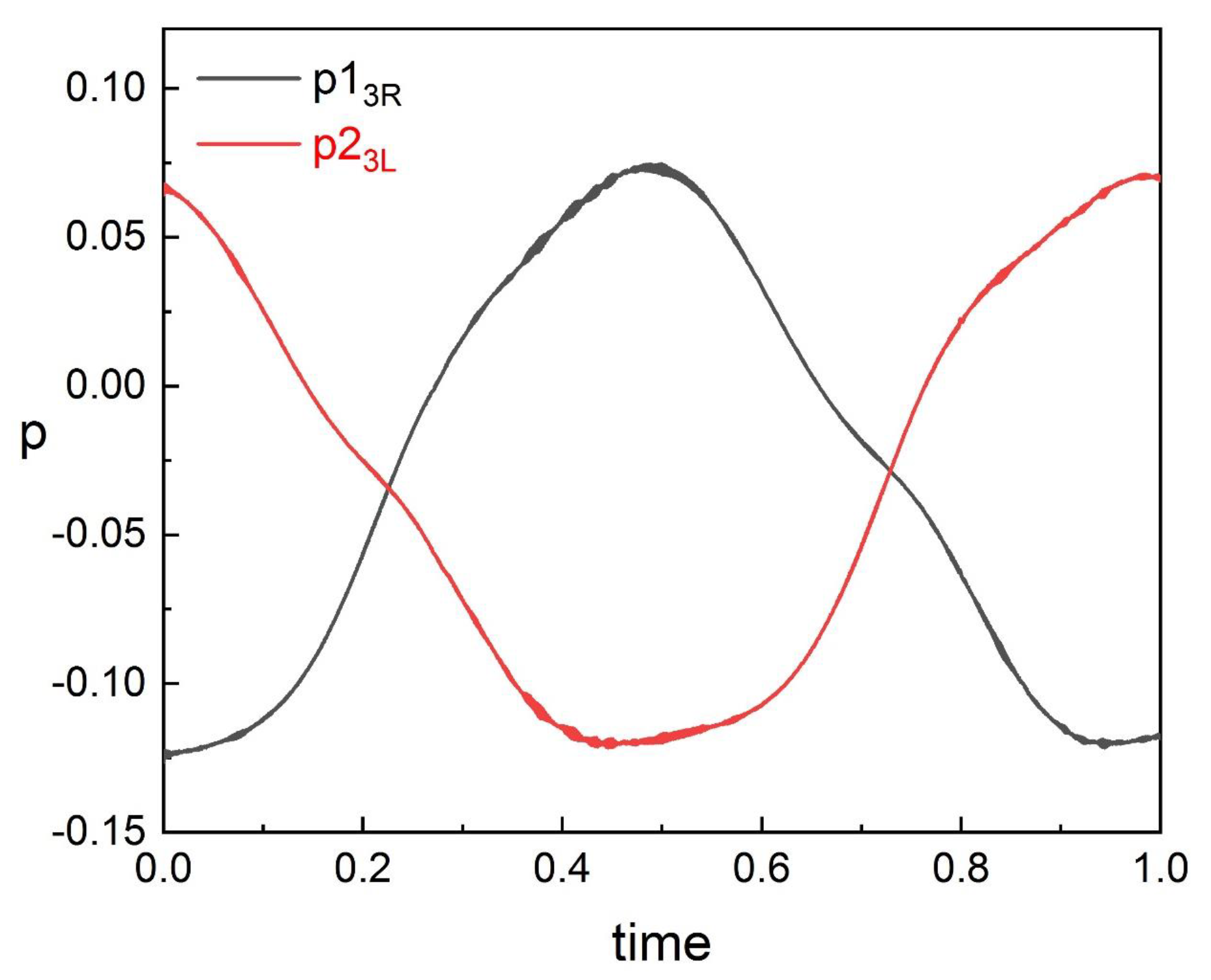
Through the results in
Figure 13 (
Section 4), we found that the pressure fields on the adjacent side of the two fishes overlap greatly at a close spacing in the phalanx formation.
and
are more susceptible to spacing than pressures on the other side (
and
). Therefore, we calculated the kinematic pressures of the specific side of fish, which are
and
. We found that
and
are symmetrical in a cycle of fish swimming, as shown in
Figure 18. The data in
Figure 18 are monitored at a suitable spacing (
), which can ensure the representativeness of the rules of data at different spacing. When the inlet flow velocity is 0, the interaction of fish body swing is large, thus the rules of the data are representative at different
U. Although there are some slight jitters in the data,
and
are approximately equal and we calculated the average of
and
as
.
In this work, the data we mainly used are monitored by sensors
,
and
. To save the cost of sensors placement, we give priority to the data at these positions. We used the data (
and
) monitored by
, which are almost unaffected by spacing to predictive the inlet flow velocity for the phalanx formation. Thus, we consider the data monitored by
and
in this part. In the phalanx formation, we calculated
and
to discuss the relationship between the kinematic pressure and the spacing
. As shown in
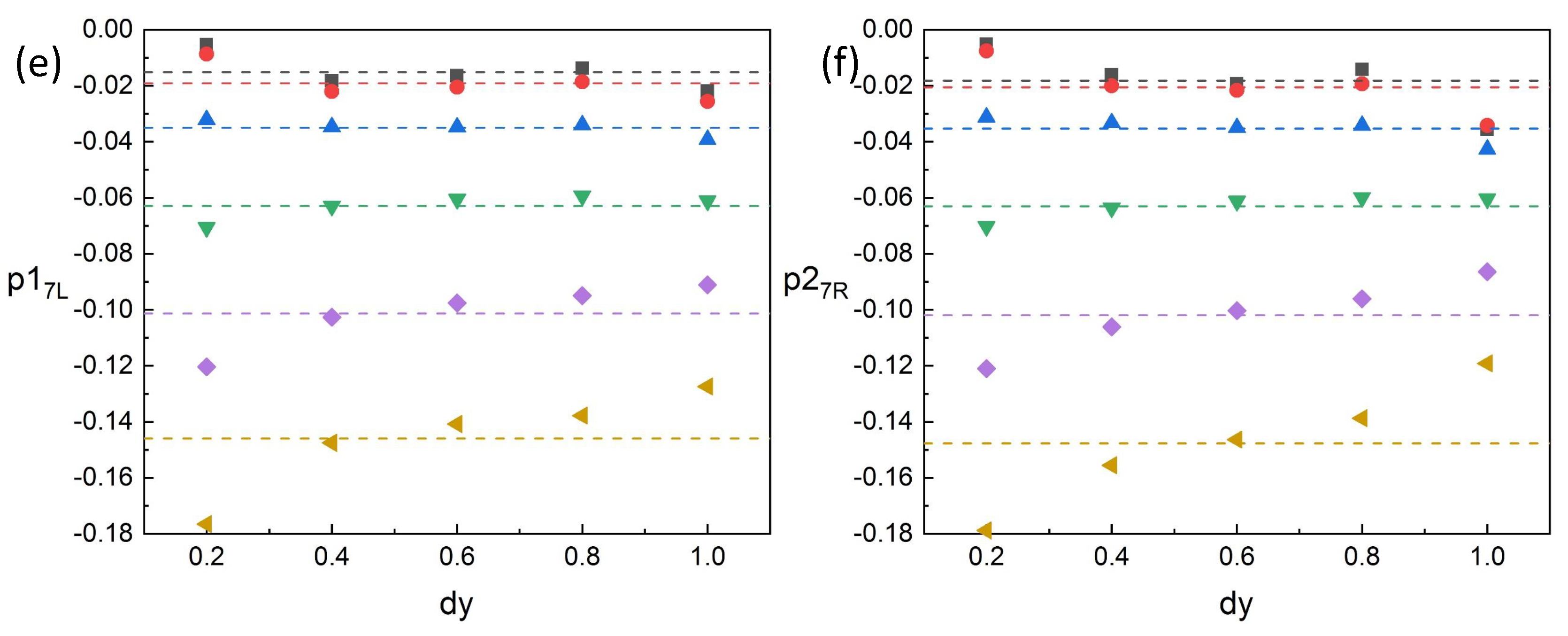
Figure 19, when the spacing
is
, the kinematic pressures differ greatly from that of other spacings at a certain
U. This is because the spacing is close, and the impact caused by the swing of two fishes is greater than that of large spacings. However, we found that
is monotonic with
increasing at a certain
U as shown in the left of
Figure 19. Thus, we can use
to judge the spacing.
5.2.2. The Construction and Validation of Judgement Models
Through the results on the left side of
Figure 19, we found that, at different inlet flow velocity
U, the relationship between
and the spacing
is different. Thus, we also need to use the kinematic pressure
or
, which is insensitive to the spacing as a feature for eliminating the effect of inlet flow velocity. For the presetting appropriate spacing
, we divide the data into two categories: the label is equal to 0 when the current spacing
is less than
, and the label is equal to 1 when the current spacing
is greater than
. We used the same neural network structure above. We used the data simulated by settings in
Table 4 to train the classification model, and used the data simulated by settings in
Table 6 to validate the model. When we construct the model, the input layer contains two features
and
(which is about equal to
). However, when we validated the model, we used the kinematic pressures of each fish, and two features in the input layer are
,
or
,
. We used several
to construct the judgement models, and the effects of each fish are shown in
Table 10.
In this section, we construct some binary classification models to judge the relationship between the current spacing and the presetting appropriate spacing . Through the results of judging, the robotic fishes can adjust their actions to maintain a stable formation at the appropriate spacing .
6. Conclusions
In this work, we construct the models for single robotic fish, the tandem formation and the phalanx formation to predict the inlet flow velocity and judge the spacing between individual platforms in the formation. The results show that the best predictive model of the inlet flow velocity for single fish is calculated by the kinematic pressures monitored by sensors at position . The predictive models of the inlet flow velocity for the front robotic fish in the tandem formation and two fishes in the phalanx formation are the same as that of single fish calculated by the kinematic pressures at the fish head. The predictive model of the inlet flow velocity for robotic fish at the back in the tandem is calculated by the kinematic pressures monitored by the sensors at position , which is different from the best model for single fish. In the tandem formation, only the fish at the back need to know the spacing with the other fish. The spacing is judged by the kinematic pressures monitored by the sensors at position . In the phalanx, both fishes need to know the spacing. For the fish on the left, the spacing is judged by the data monitored by the right sensors at position . For the right fish, the spacing is judged by the data monitored by the left sensors at position . The robotic fishes can know basic environmental information, such as the inlet flow velocity and the spacing between the robotic fish itself and the other fish in the formation through these models. It is useful for maintaining a stable formation to perform more complex tasks without communication. In addition, the results can provide guidance for sensor installation position on the robotic fish surface. In the future, we will further consider more complex formations with more robotic fishes and change the phase difference of fishes swing. We will try to improve the applicability of the predictive and judgement models. To make the simulation closer to reality, we will also consider more control algorithms and cooperate with colleagues who make robots to apply our simulation results to actual robots.
























{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}