

Risk Element Interaction with an Emerging Contaminant in Mammals: The Case of Arsenic and Carbamazepine Orally Taken up by Microtus arvalis L. as a Model Organism

 , , ,
, , ,  , ,
, , 
Abstract
1. Introduction
- (a)
- Assess the impact of the sole and co-exposure effects of CBZ and As oral uptake by M. arvalis by evaluating their accumulation and biotransformation (methylation) within the investigated animal organism as compared to the control group and in relation to each other. In this context, CBZ was present in the drinking water, whereas As-bearing soil was incorporated into the diet of voles to simulate unintentional consumption of soil under natural conditions.
- (b)
- Determine the activity of glutathione peroxidase (GPx) in blood plasma, and use it as the main indicator to assess the antioxidant enzyme defence response to the As and CBZ individual and combined uptake by the animals.
2. Materials and Methods
2.1. Ethical Approval
2.2. Animal Experiment
2.2.1. Experimental Design
2.2.2. Characteristics of Experimental Soils
2.2.3. Preparations of Experimental Diet and CBZ Solution
2.2.4. Tissue Sample Collection
2.3. Analytical Procedures
2.3.1. Determination of Total As Content in Animal Tissues and Experimental Diets
2.3.2. Extraction of As Species and As Speciation Using High-Performance Liquid Chromatography in the Liver
2.3.3. CBZ and Its Metabolites Determination
2.3.4. Glutathione Peroxidase (GPx) Activity in Blood Plasma
2.4. Calculation of Inorganic As Methylation Capacities of the Experimental Animal Groups
2.5. Data Processing and Statistical Analyses
3. Results
3.1. Body Weights of the Experimental Animal Groups
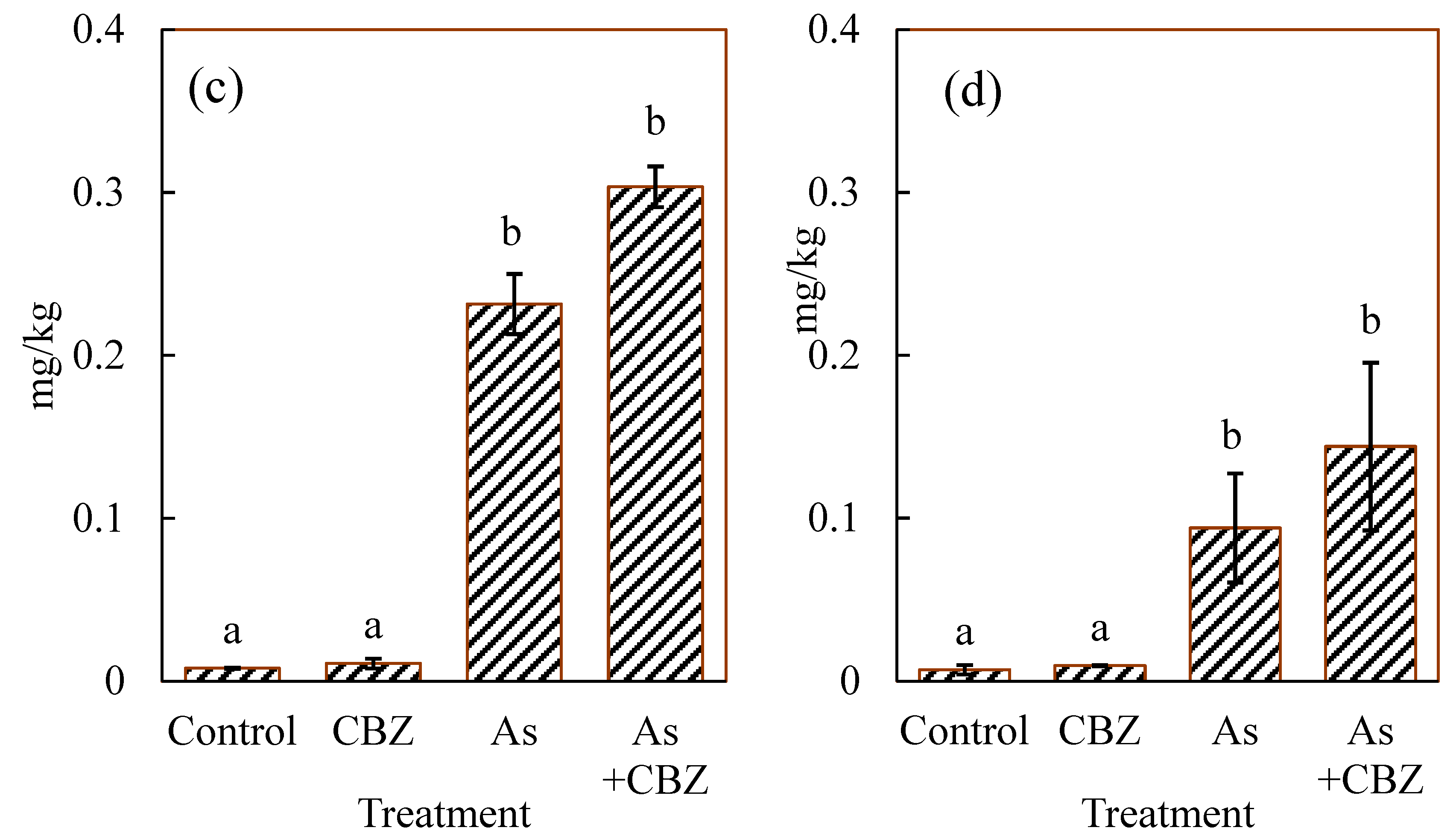
3.2. Total As Concentrations in the Various Organs of the Experimental Animals
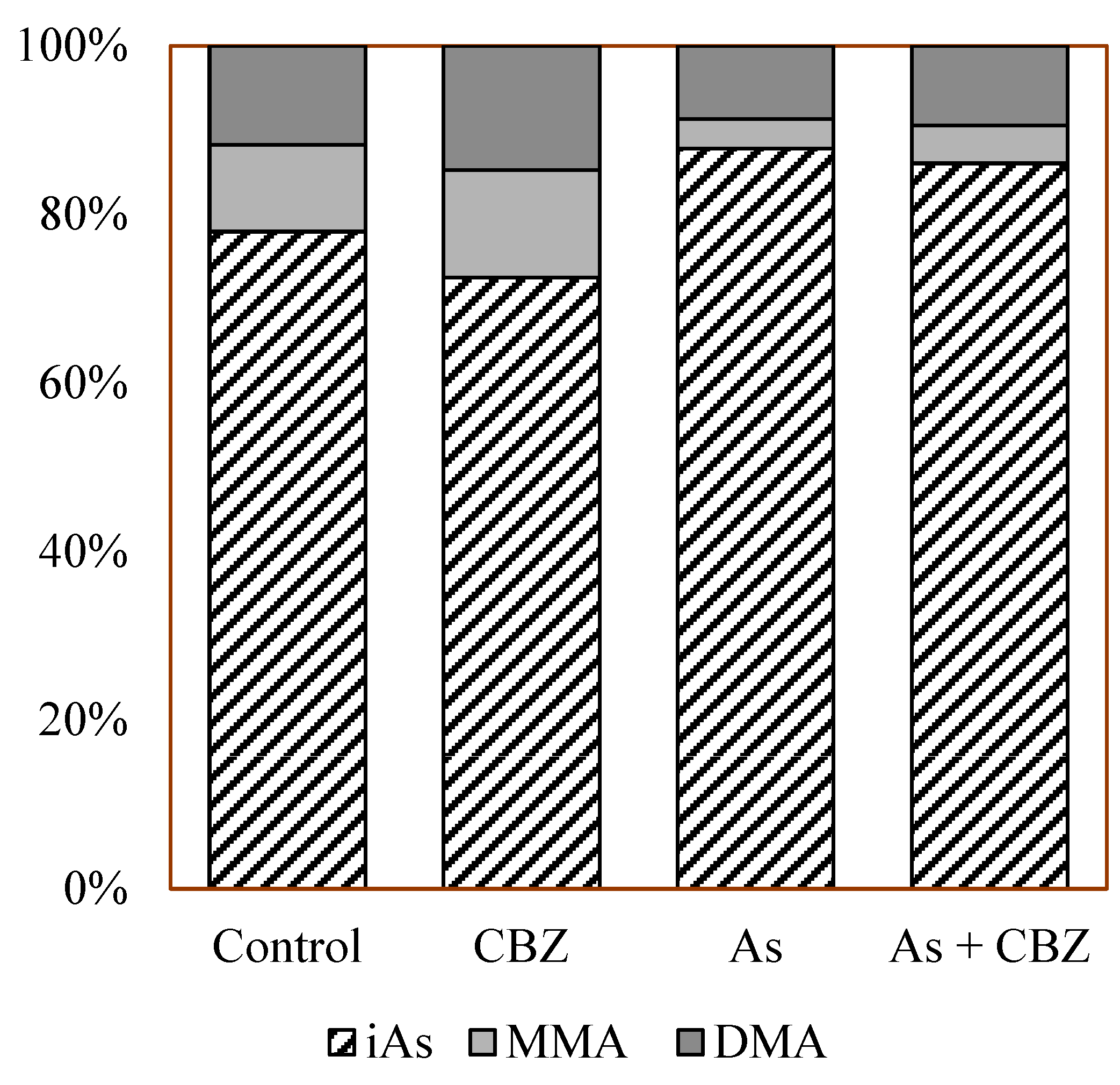
3.3. Individual As Species Concentrations and Methylation Capacities in the Liver of Experimental Animals
3.4. CBZ and Its Metabolites in the Liver of the Experimental Animals
3.5. Glutathione Peroxidase (GPx) Activity in the Blood Plasma of the Experimental Animals
4. Discussion
4.1. Total As Concentrations in Different Organs and As Species Concentrations in the Liver of the Experimental Animals
4.2. Inorganic As Methylation in the Liver of the Experimental Animals
4.3. Concentrations of CBZ and Its Metabolites in the Liver of Experimental Animals
4.4. The Activity of the Antioxidant Enzyme Glutathione Activity (GPx) in the Blood Plasma of the Experimental Animal Groups
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Geissen, V.; Mol, H.; Klumpp, E.; Umlauf, G.; Nadal, M.; van der Ploeg, M.; van de Zee, S.E.; Ritsema, C.J. Emerging pollutants in the environment: A challenge for water resource management. Int. Soil. Water Conserv. Res. 2015, 3, 57–65. [Google Scholar] [CrossRef]
- Naidu, R.; Arias Espana, V.A.; Liu, Y.; Jit, J. Emerging contaminants in the environment: Risk-based analysis for better management. Chemosphere 2016, 154, 350–357. [Google Scholar] [CrossRef] [PubMed]
- Chen, H.; Gu, X.; Zeng, Q.; Mao, Z.; Liang, X.; Martyniuk, C.J. Carbamazepine disrupts molting hormone signaling and inhibits molting and growth of Eriocheir sinensis at environmentally relevant concentrations. Aquat. Toxicol. 2019, 208, 138–145. [Google Scholar] [CrossRef]
- Wilkinson, J.L.; Boxall, A.B.A.; Kolpin, D.W.; Leung, K.M.Y.; Lai, R.W.S.; Wong, D.; Ntchantcho, R.; Pizarro, J.; Mart, J.; Echeverr, S.; et al. Pharmaceutical pollution of the world’s rivers. Proc. Natl. Acad. Sci. USA 2022, 119, e2113947119. [Google Scholar] [CrossRef]
- Ambrosio, A.F.; Soares-da-Silva, P.; Carvalho, C.M.; Carvalho, A.P. Mechanisms of action of carbamazepine and its derivatives, oxcarbazepine, BIA 2-093, and BIA 2-024. Neurochem. Res. 2002, 27, 121–130. [Google Scholar] [CrossRef]
- Deng, Y.; Ok, Y.S.; Mohan, D.; Pittman, C.U.; Dou, X. Carbamazepine removal from water by carbon dot-modified magnetic carbon nanotubes. Environ. Res. 2019, 169, 434–444. [Google Scholar] [CrossRef] [PubMed]
- Frickle-Galindo, I.; LLerena, A.; Jung-Cook, H.; Lopez-Lopez, M. Carbamazepine adverse drug reactions. Expert. Rev. Clin. Pharmacol. 2018, 11, 705–718. [Google Scholar] [CrossRef]
- Zhang, Y.; Geißen, S.U.; Gal, C. Carbamazepine and diclofenac: Removal in wastewater treatment plants and occurrence in water bodies. Chemosphere 2008, 73, 1151–1161. [Google Scholar] [CrossRef]
- Brezina, E.; Prasse, C.; Meyer, J.; Mückter, H.; Ternes, T.A. Investigation and risk evaluation of the occurrence of carbamazepine, oxcarbazepine, their human metabolites and transformation products in the urban water cycle. Environ. Pollut. 2017, 225, 261–269. [Google Scholar] [CrossRef]
- Ebrahimzadeh, S.; Castiglioni, S.; Riva, F.; Zuccato, E.; Azzellino, A. Carbamazepine levels related to the demographic indicators in groundwater of densely populated area. Water 2021, 13, 2539. [Google Scholar] [CrossRef]
- Mezzelani, M.; Gorbi, S.; Regoli, F. Pharmaceuticals in the aquatic environments: Evidence of emerged threat and future challenges for marine organisms. Mar. Envl Res. 2018, 140, 41–60. [Google Scholar] [CrossRef] [PubMed]
- Wu, M.; Xiang, J.; Que, C.; Chen, F.; Xu, G. Occurrence and fate of psychiatric pharmaceuticals in the urban water system of Shanghai, China. Chemosphere 2015, 138, 486–493. [Google Scholar] [CrossRef]
- Lindim, C.; de Zwart, D.; Cousins, I.T.; Kutsarova, S.; Kühne, R.; Schüürmann, G. Exposure and ecotoxicological risk assessment of mixtures of top prescribed pharmaceuticals in Swedish freshwaters. Chemosphere 2019, 220, 344–352. [Google Scholar] [CrossRef] [PubMed]
- Meyer, W.; Reich, M.; Beier, S.; Behrendt, J.; Gulyas, H.; Otterpohl, R. Measured and predicted environmental concentrations of carbamazepine, diclofenac, and metoprolol in small and medium rivers in northern Germany. Environ. Monit. Assess. 2016, 188, 487. [Google Scholar] [CrossRef] [PubMed]
- Almeida, Â.; Soares, A.M.V.M.; Esteves, V.I.; Freitas, R. Occurrence of the antiepileptic carbamazepine in water and bivalves from marine environments: A review. Environ. Toxicol. Pharmacol. 2021, 86, 103661. [Google Scholar] [CrossRef]
- Musolff, A.; Leschik, S.; Möder, M.; Strauch, G.; Reinstorf, F.; Schirmer, M. Temporal and spatial patterns of micropollutants in urban receiving waters. Environ. Pollut. 2009, 157, 3069–3077. [Google Scholar] [CrossRef]
- Osenbrück, K.; Gläser, H.R.; Knöller, K.; Weise, S.M.; Möder, M.; Wennrich, R.; Schirmer, M.; Reinstorf, F.; Busch, W.; Strauch, G. Sources and transport of selected organic micropollutants in urban groundwater underlying the city of Halle (Saale), Germany. Water Res. 2007, 41, 3259–3270. [Google Scholar] [CrossRef] [PubMed]
- Vulliet, E.; Cren-Olivé, C. Screening of pharmaceuticals and hormones at the regional scale, in surface and groundwaters intended to human consumption. Environ. Pollut. 2011, 159, 2929–2934. [Google Scholar] [CrossRef]
- Riva, F.; Castiglioni, S.; Fattore, E.; Manenti, A.; Davoli, E.; Zuccato, E. Monitoring emerging contaminants in the drinking water of Milan and assessment of the human risk. Int. J. Hyg. Environ. Health 2018, 221, 451–457. [Google Scholar] [CrossRef]
- Simazaki, D.; Kubota, R.; Suzuki, T.; Akiba, M.; Nishimura, T.; Kunikane, S. Occurrence of selected pharmaceuticals at drinking water purification plants in Japan and implications for human health. Water Res. 2015, 76, 187–200. [Google Scholar] [CrossRef] [PubMed]
- Wang, C.; Ye, D.; Li, X.; Jia, Y.; Zhao, L.; Liu, S.; Xu, J.; Du, J.; Tian, L.; Li, J.; et al. Occurrence of pharmaceuticals and personal care products in bottled water and assessment of the associated risks. Environ. Int. 2021, 155, 106651. [Google Scholar] [CrossRef] [PubMed]
- Datel, J.V.; Hrabankova, A. Pharmaceuticals load in the Svihov water reservoir (Czech Republic) and impacts on quality of treated drinking water. Water 2020, 12, 1387. [Google Scholar] [CrossRef]
- Kozisek, F.; Pomykacova, I.; Jeligova, H.; Cadek, V.; Svobodova, V. Survey of human pharmaceuticals in drinking water in the Czech Republic. J. Water Health 2013, 11, 84–97. [Google Scholar] [CrossRef]
- Kaushik, G.; Huber, D.P.; Aho, K.; Finney, B.; Bearden, S.; Zarbalis, K.S.; Thomas, M.A. Maternal exposure to carbamazepine at environmental concentrations can cross intestinal and placental barriers. Biochem. Biophysl Res. Commun. 2016, 474, 291–295. [Google Scholar] [CrossRef]
- Kohl, A.; Golan, N.; Cinnamon, Y.; Genin, O.; Chefetz, B.; Sela-Donenfeld, D. A proof of concept study demonstrating that environmental levels of carbamazepine impair early stages of chick embryonic development. Environ. Int. 2019, 129, 583–594. [Google Scholar] [CrossRef]
- Santariová, M.; Zadinová, K.; Vostrá-Vydrová, H.; Kolářová, M.F.; Kurhan, S.; Chaloupková, H. Effect of environmental concentration of carbamazepine on the behaviour and gene expression of laboratory rats. Animals 2023, 13, 2097. [Google Scholar] [CrossRef]
- Dean, J.R.; Ma, R. Approaches to assess the oral bioaccessibility of persistent organic pollutants: A critical review. Chemosphere 2007, 68, 1399–1407. [Google Scholar] [CrossRef] [PubMed]
- Petruzzelli, G.; Pedron, F.; Rosellini, I. Bioavailability and bioaccessibility in soil: A short review and a case study. AIMS Environ. Sci. 2020, 7, 208–224. [Google Scholar] [CrossRef]
- Tang, X.; Tang, L.; Zhu, Y.; Xing, B.; Duan, J.; Zheng, M. Assessment of the bioaccessibility of polycyclic aromatic hydrocarbons in soils from Beijing using an intro test. Environ. Pollut. 2006, 140, 279–285. [Google Scholar] [CrossRef]
- Zhang, Y.; Yang, X.; Gu, C.; Bian, Y.; Liu, Z.; Jia, M.; Wang, F.; Wang, D.; Jiang, X. Prediction of polycyclic aromatic hydrocarbon bioaccessibility to earthworms in spiked soils by composite extraction with hydroxypropyl-ß-cyclodextrin and organic acids. Pedosphere 2017, 27, 502–510. [Google Scholar] [CrossRef]
- Chen, F.; Muhammad, F.G.; Khan, Z.I.; Ahmad, K.; Malik, I.S.; Ashfaq, A.; Naeem, M.; Nadeem, M.; Ma, J.; Awan, M.U.M.; et al. Bioaccumulation and transfer of Zinc in soil plant and animal system: A health risk assessment for grazing animals. Environ. Sci. Pollut. Res. 2022, 29, 2718–2727. [Google Scholar] [CrossRef] [PubMed]
- Guo, Y.L.; Lambert, G.H.; Hsu, C.C.; Hsu, M.M.L. Yucheng: Health effects of prenatal exposure to polychlorinated biphenyls and dibenzofurans. Int. Arch. Occup. Environ. Health 2004, 77, 153–158. [Google Scholar] [CrossRef] [PubMed]
- Misaki, K.; Takamura–Enya, T.; Ogawa, H.; Takamori, K.; Yanagida, M. Tumour-promoting activity of polycyclic aromatic hydrcarbons and their oxygenated or nitrated derivative. Mutagenesis 2016, 31, 205–213. [Google Scholar] [CrossRef] [PubMed]
- Alka, S.; Shahir, S.; Ibrahim, N.; Ndejiko, M.J.; Vo, D.V.N.; Manan, F.A. Arsenic removal technologies and future trends: A mini review. J. Clean. Prod. 2021, 278, 123805. [Google Scholar] [CrossRef]
- Podgorski, J.; Berg, M. Global threat of arsenic in groundwater. Science 2020, 368, 845–850. [Google Scholar] [CrossRef]
- Tseng, C.H.; Huang, Y.K.; Huang, Y.L.; Chung, C.J.; Yang, M.H.; Chen, C.J.; Hsueh, Y.M. Arsenic exposure, urinary arsenic speciation, and peripheral vascular disease in blackfoot disease-hyperendemic villages in Taiwan. Toxicol. Appl. Pharmacol. 2005, 206, 299–308. [Google Scholar] [CrossRef]
- Xu, M.; Rui, D.; Yan, Y.; Xu, S.; Niu, Q.; Feng, G.; Wang, Y.; Li, S.; Jing, M. Oxidative Damage Induced by Arsenic in Mice or Rats: A Systematic Review and Meta-Analysis. Biol. Trace Elem. Res. 2017, 176, 154–175. [Google Scholar] [CrossRef]
- Gasca-Pérez, E.; Galar-Martínez, M.; García-Medina, S.; Pérez-Coyotl, I.A.; Ruiz-Lara, K.; Cano-Viveros, S.; Pérez-Pastén, B.R.; Gómez-Oliván, L.M. Short-term exposure to carbamazepine causes oxidative stress on common carp (Cyprinus carpio). Environ. Toxicol. Pharmacol. 2019, 66, 96–103. [Google Scholar] [CrossRef]
- Papaioannou, D.; Koukoulakis, P.H.; Lambropoulou, D.; Papageorgiou, M.; Kalavrouziotis, I.K. The dynamics of the pharmaceutical and personal care product interactive capacity under the effect of artificial enrichment of soil with heavy metals and of wastewater reuse. Sci. Total Environ. 2019, 662, 537–546. [Google Scholar] [CrossRef] [PubMed]
- Gall, J.E.; Boyd, R.S.; Rajakaruna, N. Transfer of heavy metals through terrestrial food webs: A review. Environ. Monit. Assess. 2015, 187, 201. [Google Scholar] [CrossRef] [PubMed]
- Skácel, F.; Pekárek, J. Monitoring of lead, cadmium and mercury in environmental samples at the regional station of the integrated background monitoring network of GEMS in Czechoslovakia. Sci. Total Environ. 1992, 115, 261–276. [Google Scholar] [CrossRef]
- Hamers, T.; van den Berg, J.H.J.; van Gestel, C.A.M.; van Schooten, F.J.; Murk, A.J. Risk assessment of metals and organic pollutants for herbivorous and carnivorous small mammal food chains in a polluted floodplain (Biesbosch, The Netherlands). Environ. Pollut. 2006, 144, 581–595. [Google Scholar] [CrossRef] [PubMed]
- Rodríguez-Estival, J.; Smits, J.E.G. Small mammals as sentinels of oil sands related contaminants and health effects in northeastern Alberta, Canada. Ecotoxicol. Environ. Safe 2016, 124, 285–295. [Google Scholar] [CrossRef]
- Yudovich, E.Y.; Ketris, M.P. Arsenic in coal: A review. Int. J. Coal Geol. 2005, 61, 141–196. [Google Scholar] [CrossRef]
- Skála, J.; Boahen, F.; Száková, J.; Vácha, R.; Tlustoš, P. Arsenic and lead in soil: Impacts on element mobility and bioaccessibility. Environ. Geochem. Health 2022, 44, 943–959. [Google Scholar] [CrossRef] [PubMed]
- Zadrapova, D.; Titera, A.; Szakova, J.; Cadkova, Z.; Cudlin, O.; Najmanova, J.; Tlustos, P. Mobility and bioaccessibility of risk elements in the area affected by the long-term opencast coal mining. J. Environ. Sci. Health A Tox Hazard. Subst. Environ. Eng. 2019, 54, 1159–1169. [Google Scholar] [CrossRef]
- Zita, L.; Kurhan, S.; Krunt, O.; Chmelíková, E.; Kraus, A.; Čítek, J.; Klouček, P.; Stupka, R. The effect of carbamazepine on performance, carcass value, hematological and biochemical blood parameters, and detection of carbamazepine and its metabolites in tissues, internal organs, and body fluids in growing rabbits. Animals 2023, 13, 2041. [Google Scholar] [CrossRef]
- European Commission. Joint Research Centre (JRC) (2019): EURL ECVAM Dataset on Alternative Methods to Animal Experimentation (DB-ALM). European Commission, Joint Research Centre (JRC) [Dataset] PID. Available online: http://data.europa.eu/89h/b7597ada-148d-4560-9079-ab0a5539cad3 (accessed on 8 January 2025).
- Mercl, F.; Košnář, Z.; Maršík, P.; Vojtíšek, M.; Dušek, J.; Száková, J.; Tlustoš, P. Pyrolysis of biosolids as an effective tool to reduce the uptake of pharmaceuticals by plants. J. Hazard. Mat. 2021, 405, 124278. [Google Scholar] [CrossRef]
- Public Notice No.153/2016 About the Conditions for the Protection of the Agricultural Soil Quality; Legal code of The Czech Republic: Brno, Czech Republic, 2016; pp. 2692–2699.
- Tremlová, J.; Száková, J.; Tlustoš, P. An assessment of possible effect of risk elements contained in soil on human organism. Chem. Listy 2010, 104, 349–352. (In Czech) [Google Scholar]
- Vlčková, V.; Malinová, M.; Koubková, B.; Száková, J.; Zídek, V.; Fučíková, A.; Zídková, J.; Kolihová, D.; Tlustoš, P. The long-term effect of diet amended by risk elements contaminated soils on risk element penetration and physiological parameters of rats. Czech J. Anim. Sci. 2014, 59, 416–427. [Google Scholar] [CrossRef]
- Lin, Y.; Sun, Y.; Wang, X.; Chen, S.; Wu, Y.; Fu, F. Universal method for the speciation analysis in various seafood based on microwave-assisted extraction and ion chromatography-inductively coupled plasma mass spectrometry. Microchem. J. 2020, 159, 105592. [Google Scholar] [CrossRef]
- Button, M.; Moriarty, M.M.; Watts, M.J.; Zhang, J.; Koch, I.; Reimer, K.J. Arsenic speciation in field-collected and laboratory-exposed earthworms lumbricus terrestris. Chemosphere 2011, 85, 1277–1283. [Google Scholar] [CrossRef] [PubMed]
- Lawrence, R.A.; Burk, R.F. Glutathione peroxidase activity in selenium—Deficient rat liver. Biochem. Biophys. Res. Commun. 1974, 71, 952–958. [Google Scholar] [CrossRef]
- Lawrence, R.A.; Burk, R.F. Species, tissue and subcellular distribution of non-Se-dependent glutathione peroxidase activity. J. Nutr. 1978, 108, 211–215. [Google Scholar] [CrossRef] [PubMed]
- EC 1.11.19; Enzymatic Assay of Gluthathione Peroxidase. Sigma-Aldrich: Burlington, MA, USA, 2000.
- Hsieh, R.; Huang, Y.; Shiue, H.; Huang, S.; Lin, M.; Mu, S.; Chung, C.; Hsueh, Y. Arsenic methylation capacity and developmental delay in preschool children in Taiwan. Int. J. Hyg. Environ. Health 2014, 217, 678–686. [Google Scholar] [CrossRef]
- Stýblo, M.; Douillet, C.; Bangma, J.; Eaves, L.A.; de Villena, F.P.; Fry, R. Differential metabolism of inorganic arsenic in mice from genetically diverse Collaborative Cross strains. Arch. Toxicol. 2019, 93, 2811–2822. [Google Scholar] [CrossRef] [PubMed]
- Katoh, H. International Harmonization of Laboratory Animals. In Microbial Status and Genetic Evaluation of Mice and Rats: Proceedings of the 1999 US/Japan Conference; National Academies Press (US): Washington, DC, USA, 2000. [Google Scholar]
- EPA. Guidelines for Developmental Toxicity Risk Assessment. Fed. Regist. 1991, 56, 63798–63826. [Google Scholar]
- EFSA. Statistical Significance and Biological Relevance. EFSA J. 2011, 9, 2372. [Google Scholar]
- Hughes, M.F.; Kenyon, E.M.; Edwards, B.C.; Mitchell, C.T.; Razo, L.M.D.; Thomas, D.J. Accumulation and metabolism of arsenic in mice after repeated oral administration of arsenate. Toxicol. Appl. Pharmacol. 2003, 191, 202–210. [Google Scholar] [CrossRef] [PubMed]
- Souza, J.M.O.; Grotto, D.; Batista, B.L.; Júnior, F.B. Distribution of arsenic and oxidative stress in mice after rice ingestion. J. Trace Elem. Med. Biol. 2017, 44, 192–200. [Google Scholar] [CrossRef]
- Li, J.; Guo, Y.; Duan, X.; Li, B. Tissue- and region-specific accumulation of arsenic species, especially in the brain of mice, after long-term arsenite exposure in drinking water. Biol. Trace Elem. Res. 2020, 198, 168–176. [Google Scholar] [CrossRef]
- Shao, J.; Li, X.; Luo, Y.; Fang, H.; Lin, F.; Zheng, G.; Lu, F.; Guo, L.; Sun, Y. Distribution of arsenic species and pathological characteristics of tissues of the mice fed with arsenic–supplemented food simulating rice. J. Toxicol. Sci. 2021, 46, 539–551. [Google Scholar] [CrossRef] [PubMed]
- Ratnaike, R.N. Acute and chronic arsenic toxicity. Postgrad. Med. J. 2003, 79, 391–396. [Google Scholar] [CrossRef]
- Zhou, Q.; Xi, S. A review on arsenic carcinogenesis: Epidemiology, metabolism, genotoxicity and epigenetic changes. Regul. Toxicol. Pharmacol. 2018, 99, 78–88. [Google Scholar] [CrossRef] [PubMed]
- Moreno-Jiménez, E.; Esteban, E.; Peñalosa, J.M. The fate of arsenic in soil-plant systems. Rev. Environ. Contam. Toxicol. 2012, 215, 1–37. [Google Scholar] [PubMed]
- Zhao, F.J.; McGrath, S.P.; Meharg, A.A. Arsenic as a food-chain contaminant: Mechanisms of plant uptake and metabolism and mitigation strategies. Ann. Rev. Plant Biol. 2010, 61, 535–559. [Google Scholar] [CrossRef]
- Maciaszczyk-Dziubinska, E.; Wawrzycka, D.; Wysocki, R. Arsenic and antimony transporters in eukaryotes. Int. J. Mol. Sci. 2012, 13, 3527–3548. [Google Scholar] [CrossRef]
- Kenyon, E.M.; Hughes, M.F.; Adair, B.M.; Highfill, J.H.; Crecelius, E.A.; Clewell, H.J.; Yager, J.W. Tissue distribution and urinary excretion of inorganic arsenic and its methylated metabolites in C57BL6 mice following subchronic exposure to arsenate in drinking water. Toxicol. Appl. Pharmacol. 2008, 232, 448–455. [Google Scholar] [CrossRef]
- Khairul, I.; Wang, Q.Q.; Jiang, Y.H.; Wang, C.; Naranmandura, H. Metabolism, toxicity and anticancer activities of arsenic compounds. Oncotarget 2017, 8, 23905–23926. [Google Scholar] [CrossRef]
- Thomas, D.J.; Li, J.; Waters, S.B.; Xing, W.; Adair, B.M.; Drobna, Z.; Devesa, V.; Styblo, M. Arsenic (+3 oxidation state) methyltransferase and the methylation of arsenicals. Exp. Biol. Med. 2007, 232, 3–13. [Google Scholar]
- Cui, X.; Okayasu, R. Arsenic accumulation, elimination, and interaction with copper, zinc and manganese in liver and kidney of rats. Food Chem. Toxicol. 2008, 46, 3646–3650. [Google Scholar] [CrossRef]
- Soria, E.A.; Pérez, R.D.; Queralt, I.; Pérezd, C.A.; Bongiovannie, G.A. Immunotoxicological effects of arsenic bioaccumulation on spatial metallomics and cellular enzyme response in the spleen of male Wistar rats after oral intake. Toxicol. Lett. 2017, 266, 65–73. [Google Scholar] [CrossRef]
- Putila, J.J.; Guo, N.L. Association of arsenic exposure with lung cancer incidence rates in the United States. PLoS ONE 2011, 6, e25886. [Google Scholar] [CrossRef]
- Ren, C.; Zhou, Y.; Liu, W.; Wang, Q. Paraxodical effects of arsenic in the lungs. Environ. Health Prevent Med. 2021, 26, 80. [Google Scholar] [CrossRef] [PubMed]
- Yi, Y.; Gao, S.; Xia, J.; Li, C.; Zhao, Y.; Zhang, Y.; Liang, A.; Ji, S. Study of the accumulation and distribution of arsenic species and association with arsenic toxicity in rats after 30 days of oral realgar administration. J. Ethnopharmacol. 2020, 247, 111576. [Google Scholar] [CrossRef] [PubMed]
- Twaddle, N.C.; Vanlandingham, M.; Churchwell, M.I.; Doerge, D.R. Metabolism and disposition of arsenic species from controlled oral dosing with sodium arsenite in adult female CD-1 mice. I. Pilot study to determine dosing, analytical measurements, and sampling strategies. Food Chem. Toxicol. 2018, 111, 482–493. [Google Scholar] [CrossRef]
- Watanabe, T.; Hirano, S. Metabolism of arsenic and its toxicological relevance. Arch. Toxicol. 2013, 87, 969–979. [Google Scholar] [CrossRef] [PubMed]
- Bogdan, G.M.; Sampayo-Reyest, A.; Aposhian, H.V. Arsenic binding protins of mammalian systems: I. Isolation of three arsnite-binding proteins of rabbit liver. Toxicology 1994, 93, 175–193. [Google Scholar] [CrossRef] [PubMed]
- Doerge, D.R.; Twaddle, N.C.; Churchwell, M.I.; Beland, F.A. Reduction by, ligand exchange among, and covalent binding to glutathione and cellular thiols link metabolism and disposition of dietary arsenic species with toxicity. Environ. Int. 2020, 144, 106086. [Google Scholar] [CrossRef]
- Drobna, Z.; Naranmandura, H.; Kubachka, K.M.; Edwards, B.C.; Herbin-Davis, K.; Styblo, M.; Le, X.C.; Creed, J.T.; Maeda, N.; Hughes, M.F.; et al. Disruption of the arsenic (+3 oxidation state) methyltransferase gene in the mouse alters the phenotype for methylation of arsenic and affects distribution and retention of orally administered arsenate. Chem. Res. Toxicol. 2009, 22, 1713–1720. [Google Scholar] [CrossRef] [PubMed]
- Thomas, D.J. Arsenic methylation—Lessons from three decades of research. Toxicology 2021, 457, 152800. [Google Scholar] [CrossRef]
- Styblo, M.; Del Razo, L.M.; Vega, L.; Germolec, D.R.; LeCluyse, E.L.; Hamilton, G.A.; Reed, W.; Wang, C.; Cullen, W.R.; Thomas, D.J. Comparative toxicity of trivalent and pentavalent inorganic and methylated arsenicals in rat and human cells. Arch. Toxicol. 2000, 74, 289–299. [Google Scholar] [CrossRef] [PubMed]
- Sun, G.; Xu, Y.; Li, X.; Jin, Y.; Li, B.; Sun, X. Urinary Arsenic Metabolites in Children and Adults Exposed to Arsenic in Drinking Water in Inner Mongolia, China. Environ. Health Perspect. 2007, 115, 648–652. [Google Scholar] [CrossRef] [PubMed]
- Peña, L.C.S.; Hernández, A.B.; Razo, L.M.D. Decreased arsenic disposition and alteration of its metabolic profile in mice coexposed to fluoride. Biol. Trace Elem. Res. 2024, 202, 1594–1602. [Google Scholar] [CrossRef] [PubMed]
- Wang, J.; Zhang, G.; Lin, Z.; Luo, Y.; Fang, H.; Yang, L.; Xie, J.; Guo, L. Determination of arsenicals in mouse tissues after simulated exposure to arsenic from rice for sixteen weeks and the effects on histopathological features. Ecotoxicol. Environ. Safe 2020, 200, 110742. [Google Scholar] [CrossRef]
- Chen, F.; Gong, Z.; Kelly, B.C. Rapid analysis of pharmaceuticals and personal care products in fish plasma micro-aliquots using liquid chromatography tandem mass spectrometry. J. Chromatogr. 2015, 1383, 104–111. [Google Scholar] [CrossRef]
- Daniele, G.; Fieu, M.; Joachim, S.; Bado-Nilles, A.; Beaudouin, R.; Baudoin, P.; James-Casas, A.; Andres, S.; Bonnard, M.; Bonnard, I.; et al. Determination of carbamazepine and 12 degradation products in various compartments of an outdoor aquatic mesocosm by reliable analytical methods based on liquid chromatography-tandem mass spectrometry. Environ. Sci. Pollut. Res. 2017, 24, 16893–16904. [Google Scholar] [CrossRef] [PubMed]
- Valdés, M.E.; Amé, M.V.; Bistoni, M.A.; Wunderlin, D.A. Occurrence and bioaccumulation of pharmaceutical in a fish species inhabiting the Suquía River basin (Córdoba, Argentina). Sci. Total Environ. 2014, 472, 389–396. [Google Scholar] [CrossRef]
- Higuchi, S.; Yano, A.; Takai, S.; Tsuneyama, K.; Fukami, T.; Nakajima, M.; Yokoi, T. Metabolic activation and inflammation reactions involved in carbamazepine-induced liver injury. Toxicol. Sci. 2012, 130, 4–16. [Google Scholar] [CrossRef] [PubMed]
- Pei, J.; Pan, X.; Wei, G.; Hua, Y. Research progress of glutathione peroxidase family (GPX) in redoxidation. Front. Pharmacol. 2023, 14, 1147414. [Google Scholar] [CrossRef] [PubMed]
- Yang, L.; Chen, Y.; Liu, Y.; Xing, Y.; Miao, C.; Zhao, Y.; Chang, X.; Zhang, Q. The role of oxidative stress and natural antioxidants in ovarian aging. Front. Pharmacol. 2021, 14, 617843. [Google Scholar] [CrossRef]
- Nandi, D.; Patra, R.; Swarup, D. Oxidative stress indices and plasma biochemical parameters during oral exposure to arsenic in rats. Food Chem. Toxicol. 2006, 44, 1579–1584. [Google Scholar] [CrossRef] [PubMed]
- Das, S.; Santra, A. Implications of oxidative stress and hepatic cytokine (TNF-α and IL-6) response in the pathogenesis of hepatic collagenesis in chronic arsenic toxicity. Toxicol. Appl. Pharmacol. 2005, 204, 18–26. [Google Scholar] [CrossRef] [PubMed]
- Santra, A.; Maiti, A.; Das, S.; Lahiri, S.; Charkaborty, S.K.; Mazumder, D.N. Hepatic damage caused by chronic arsenic toxicity in experimental animals. J. Clin. Toxicol. 2000, 38, 395–405. [Google Scholar] [CrossRef] [PubMed]
- Cavar, S.; Bošnjak, Z.; Klapec, T.; Barišić, K.; Cepelak, I.; Jurasović, J.; Milić, M. Blood selenium, glutathione peroxidase activity and antioxidant supplementation of subjects exposed to arsenic via drinking water. Environ. Toxicol. Pharmacol. 2010, 29, 138–143. [Google Scholar] [CrossRef] [PubMed]
- Shila, S.; Subathra, M.; Devi, M.A.; Panneerselvam, C. Arsenic intoxication-induced reduction of glutathione level and of the activity of related enzymes in rat brain regions: Reversal by DL-a-lipoic acid. Arch. Toxicol. 2005, 79, 140–146. [Google Scholar] [CrossRef]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Group | As Contained Diet | CBZ Enriched Water | n | Total Mean Feed Consumption (g per Animal) | Individual Water Consumption (mL per Animal) |
|---|---|---|---|---|---|
| Control | Very low | No | 11 | 269 | 492 |
| CBZ | Very low | Yes | 14 | 252 | 436 |
| As | Very high | No | 13 | 268 | 457 |
| As + CBZ | Very high | Yes | 14 | 192 | 328 |
| Elements | Cítov Soil mg/kg | Mokrsko Soil mg/kg | Preventive Values * mg/kg |
|---|---|---|---|
| As | 12 ± 1.1 | 1304 ± 131 | 20.0 |
| Be | 0.778 ± 0.014 | 0.545 ± 0.018 | 2.0 |
| Cd | 0.165 ± 0.046 | 0.265 ± 0.005 | 0.5 |
| Co | 6.08 ± 0.36 | 11.7 ± 0.0 | 30.0 |
| Cr | 25.3 ± 0.4 | 16.0 ± 0.1 | 90.0 |
| Cu | 10.7 ± 0.1 | 30.8 ± 6.4 | 60.0 |
| Ni | 14.0 ± 0.5 | 8.23 ± 0.46 | 50.0 |
| Pb | 12.4 ± 0.6 | 10.2 ± 0.8 | 60.0 |
| V | 34.7 ± 0.0 | 91.3 ± 1.3 | 130.0 |
| Zn | 39.4 ± 0.7 | 76.6 ± 0.2 | 120.0 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2025 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Boahen, F.; Száková, J.; Čadková, Z.; Štanderová, J.; Schwarz, C.; Gierus, M.; Praus, L.; Mercl, F.; Tlustoš, P. Risk Element Interaction with an Emerging Contaminant in Mammals: The Case of Arsenic and Carbamazepine Orally Taken up by Microtus arvalis L. as a Model Organism. Appl. Sci. 2025, 15, 1662. https://doi.org/10.3390/app15031662
Boahen F, Száková J, Čadková Z, Štanderová J, Schwarz C, Gierus M, Praus L, Mercl F, Tlustoš P. Risk Element Interaction with an Emerging Contaminant in Mammals: The Case of Arsenic and Carbamazepine Orally Taken up by Microtus arvalis L. as a Model Organism. Applied Sciences. 2025; 15(3):1662. https://doi.org/10.3390/app15031662
Chicago/Turabian StyleBoahen, Frank, Jiřina Száková, Zuzana Čadková, Jana Štanderová, Christiane Schwarz, Martin Gierus, Lukáš Praus, Filip Mercl, and Pavel Tlustoš. 2025. "Risk Element Interaction with an Emerging Contaminant in Mammals: The Case of Arsenic and Carbamazepine Orally Taken up by Microtus arvalis L. as a Model Organism" Applied Sciences 15, no. 3: 1662. https://doi.org/10.3390/app15031662
APA StyleBoahen, F., Száková, J., Čadková, Z., Štanderová, J., Schwarz, C., Gierus, M., Praus, L., Mercl, F., & Tlustoš, P. (2025). Risk Element Interaction with an Emerging Contaminant in Mammals: The Case of Arsenic and Carbamazepine Orally Taken up by Microtus arvalis L. as a Model Organism. Applied Sciences, 15(3), 1662. https://doi.org/10.3390/app15031662