Featured Application
The Utilization of Carotenoids, Vitamin A, and its Vitaminoids from various sources in several Nutricosmetic, Cosmeceutical, and Cosmetic Applications with Skin-Health Promoting Properties.
Abstract
Following the significant advancement in the cosmetic field over the past few decades, carotenoids, vitamin A, and vitaminoids have emerged as pivotal components in the formulation of cosmetic products due to their diverse bioactive properties. Delving into a general approach on vitamin A and its derivatives’ structure, activity, biochemical way of action, and benefits, their role towards promoting mainly skin health is thoroughly detailed. For this purpose, vitamin A, vitaminoids, and carotenoids of animal, marine, plant, herb, and microorganism sources were extensively reviewed in order to evaluate their health benefits regarding skin protection. Vitamin A and its derivatives of any source heavily contributed to specific skin-related functions, including their anti-aging, skin regeneration, wound healing, hyperpigmentation, and acne treatment activity, by primarily supporting hydration, skin elasticity, and barrier repair. This review also entails recent advances in the delivery systems of these compounds, such as microencapsulation and nanoemulsions, while their potential side effects are addressed as well. Ultimately, limitations and future perspectives of vitamin A, vitaminoids, and carotenoids, considering their utilization in nutricosmetic, cosmeceutical, and cosmetic products, are further discussed.
Keywords:
vitamin A; vitaminoids; carotenoids; retinol; retinal; retinoic acid; cosmetics; anti-aging; antioxidant; skin health 1. Introduction
According to ancient Egyptian writings, the study upon vitamin A’s significance dates back to around 1500 BC. The year 1913 has specifically marked the beginning of the contemporary study on vitamin A, while in the year span of 1913 to the 1950s, many important findings led the scientific community to delve more into this compound’s unique profile. Because of the colored plant lipids’ and colorless liver extracts’ biological activity, Steenbock [1,2] proposed the interconversion of two vitamin types (1919), and it was only ten years later when Moore [3] exhibited how liver tissue changed the plant pigment β-carotene into vitamin A’s colorless form [1,3]. Those research findings were enough for scientists to shortly after discover that a vitamin A shortage also existed in children and that the retinene present in the eye’s visual pigments was the same compound as the chemical substance retinaldehyde [1,3]. Moreover, in the 1940s, Otto Isler and his colleagues [1,4] synthesized the all-trans vitamin A from the less expensive precursor β-ionone, which led to a decrease in its price in the global market, while in the 1950s, the discovery of the radioactive vitamin A isotopes further clarified its metabolic pathways and behavior. Once the specifics of the rhodopsin-related visual cycle in the eye were introduced, scientific research shifted to vitamin A’s potential biological role in cellular differentiation and growth, promoting skin’s well-being [1,4].
As apparent, the pursuit of youthful and healthy skin has long been a driving force in cosmetic science. As consumers increasingly seek formulations that are both effective and natural, bioactive compounds such as carotenoids, vitamin A, and its vitaminoids have gained substantial attention in the cosmetic industry. These compounds, derived from natural or synthetic sources, have several biological effects and have offered multifaceted benefits, such as antioxidant protection, skin repair, and anti-aging properties. Their integration into cosmeceuticals reflects a growing alignment between dermatological science and consumer expectations for more effective skincare solutions [5,6,7]. A glimpse of these bioactives’ skin health-related activity is depicted in Figure 1.
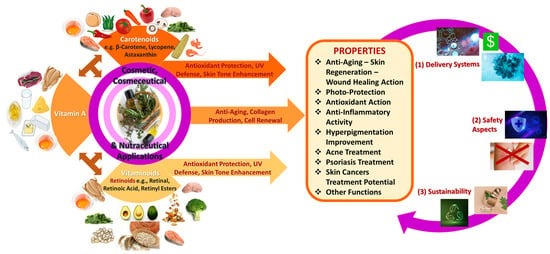
Figure 1.
The beneficial use of carotenoids, vitamin A, and vitaminoids in cosmetic products.
Carotenoids, mainly found in plants and algae, including β-carotene, lutein, and lycopene, have the ability to neutralize reactive oxygen species (ROS), hence to protect skin cells from oxidative damage induced by environmental factors like ultraviolet radiation (UV) and pollution. Furthermore, carotenoids may enhance skin tone and radiance in plant-based, eco-conscious formulations [1,5,6,7,8,9]. Following, vitamin A and its derivatives, such as retinol and retinoic acid, are responsible for regulating cell turnover, stimulating collagen production, and addressing several skin concerns, like fine lines, wrinkles, and hyperpigmentation [3,4,10,11]. However, challenges such as irritation and photosensitivity have recently been managed by advancements in vitamin A encapsulation and slow-release delivery systems [1,11]. Lastly, vitaminoids, mainly in the form of retinoids, are integral factors to skin hydration, barrier function, and repair mechanisms, being compatible with many skin-related formulations [12,13,14].
Recent innovations in cosmetic science have focused on improving the stability, bioavailability, and delivery of these active ingredients by investigating their structure, molecular activity, biochemical behavior, and skin-health benefits [1,6]. By examining bioactives like vitamin A, vitaminoids, and carotenoids of animal, marine, plant, and microorganism origin, their skin regeneration, wound healing, hyperpigmentation, and acne treatment activity, as well as their ability to promote hydration, skin elasticity, and barrier repair, have been quite supported [6]. Interestingly, newly emerged techniques such as nanoemulsions and microencapsulation have enhanced their efficacy and synergistic effects in skincare while minimizing any adverse reactions [1,6,15].
This review aims to provide a comprehensive overview of the roles of carotenoids, vitamin A, and vitaminoids in cosmetic formulations. By delving into their mechanisms of action and recent advancements, without overlooking any safety considerations, this review highlights their potential to revolutionize skincare. As the demand for natural, effective, and sustainable products grows, these bioactives stand at the forefront of innovations that merge science and consumer preferences.
2. Materials and Methods
The Scopus database was predominantly utilized for obtaining relevant literature. Some of the main keywords and key phrases used in this review are the following: “vitamins”, “vitamin A”, “vitaminoids”, “carotenoids”, “provitamins”, “vitamin A derivatives”, “β-carotene”, “retinol”, “retinal”, “retinoic acid”, “cosmetics”, “cosmeceuticals”, “nutricosmetics”, “nutraceuticals”, “pharmaceuticals”, “anti-aging”, “antioxidant”, “skin health”, “acne treatment”, “skin regeneration”, “hyperpigmentation”, “adaptive immunity”, “health benefits”, “metabolism”, “wound healing”, “microencapsulation”, “nanoemulsions”, “animal sources”, “marine sources”, “plant origin”, “microorganism sources”, “side effects”, “precautions”, with the use of combinations of these keywords by using the AND and/or OR terms in each query so as to retrieve information from relevant scientific databases, like Scopus, Science Direct, Google Scholar, PubMed, and Research Gate.
This search process was concluded during September–December 2024, considering the last 5–10 years, and the selection criteria were determined by applying the available metadata from these databases, with the eligible review studies meeting the corresponding criteria: (i) being exclusively research articles; (ii) written in the English language; and (iii) being published between 2018 and 2024. Moreover, articles’ quality and relevance were evaluated by reviewing their titles, abstracts, and keywords and then excluding duplicates retrieved from different databases unrelated to the topic.
Subsequently, the selected articles were extensively analyzed in order to determine whether they met the predefined inclusion criteria and provided pertinent information. Conference papers, books, old reviews, and short surveys, as well as publications written in other languages, were excluded. A limited number of important articles prior to 2018 were also included, since they had not been previously reviewed thoroughly.
3. Vitamin A, Its Vitaminoids and Carotenoids’ General Profile and Health Benefits
Vitamin A, vitaminoids, and carotenoids are fat-soluble hydrocarbons that act as micronutrients and antioxidant factors, primarily functioning against oxidative stress and its consequences, with a profound impact on human health. Such bioactives are integral to several physiological processes (e.g., vision, immune function, cellular communication, and antioxidant defense). While vitamin A refers to a group of retinoids (e.g., retinol, retinal, retinoic acid), carotenoids like β-carotene serve as provitamins able to be converted into active vitamin A. Vitaminoids plus encompass compounds structurally or functionally related to vitamins, often acting as precursors or analogs with distinct activity [16,17,18,19]. The dietary sources of these compounds range from animal sources rich in retinoids to plant-based sources abundant in carotenoids. Also, they exhibit therapeutic benefits such as decreasing chronic diseases like cancer risk, supporting skin health, and mitigating oxidative stress [16]. This section explores the general profile of vitamin A, vitaminoids, and carotenoids, emphasizing their biochemical and health-related significance.
3.1. Chemical Structures, Origin, and Isolation of Vitamin A and Its Vitaminoids
The vitamin A group is comprised of compounds of several origins and a basic structure of an unsaturated isoprenoid chain that provides them with the same functional abilities. It is a 20-carbon (C) molecule comprised of a cyclohexenyl ring with methyl substitution (β-ionone ring) and a conjugated polyene (isoprenoid side chain) chain structure (-C=C-). Vitamin A compounds are, in fact, retinoids (vitaminoids) that own four isoprenoid units of natural or organic origin and share a mutual interaction with retinoid receptors [3,20]. First-generation retinoids are dietary-derived metabolites, including retinol (vitamin A), retinal (all-trans-retinaldehyde), all-trans-retinoid acid (ATRA, tretinoin, or all-trans-RA), all-trans-retinyl ester, 9-cis-retinoic acid (alitretinoin), and 13-cis-retinoic acid (isotretinoin), while 2nd (etretinate and acitretin), 3rd (adapalene, tazarotene, and bexarotene), and 4th (trifarotene) generation ones are used in many diseases’ treatments. Vitamin A is either obtained from animals (retinol and retinoids) or as provitamin A, otherwise carotenoids, from consumable plant sources [3,20]. Moreover, it is mostly associated with retinol, and its main oxidized derivatives are 11-cis-retinal and ATRA. Other well-known retinoids with clinical significance in skin-protection cosmetics are 3,4-didehydroretinol, 11-cis-retinal, 5,6-epoxyretinoic acid, retro anhydroretinol, 4-oxoretinol, and retinoyl-β-glucoronide (retinylglucoronide) (Figure 2) [3,20,21].
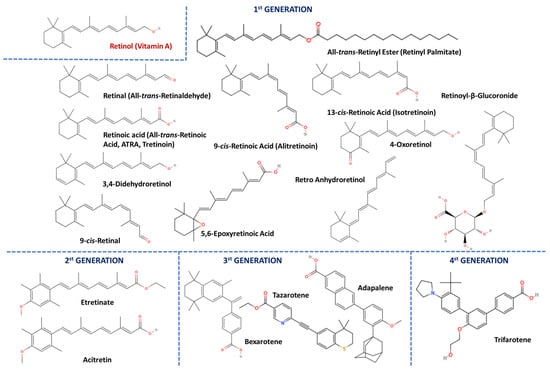
Figure 2.
The structure of vitamin A and retinoids of the 1st, 2nd, 3rd, and 4th generations.
A common vitamin A source derives from its preformed form and is obtained via animal-based food. Major retinol contributors include milk [22], dairy products [23], meat [24], chicken [25], eggs (mainly egg yolks) [3,26], and fish [27], while its general concentration ranges from 20 to 80 μg/100 mL of whole milk (the higher the fat content in the dairy product, the higher the retinol content) [3,20]. Retinol and ATRA are primarily obtained during breastfeeding, since vitamin A is responsible for mammary gland development and lactation, not only for the offspring but also for maternal health [22,28].
Considering vitamin A content in meat and poultry, tens of mg of retinol and its esters have been reported (per 100 g), and the highest content was recorded in pigs’ liver [3,20,29,30]. It must be noted that the amount of vitamin A traced in such foods, is dependent on the amount of β-carotene included in the feed or food supplements the animal is provided with [3]. Vitamin A is also obtained by consuming fish, with predator species such as carnivorous fish (e.g., sharks) and liver oil from various marine species, owning the highest levels of retinol and retinoids [27,31,32]. Egg yolks, especially chicken egg yolks, are stated to contain almost up to 1 mg of vitamin A per 100 mg, in contrast to duck and quail eggs, which comprise it in lower and elevated amounts, respectively [25,33]. Furthermore, retinoids also derive from several dietary groups like animal (e.g., chicken, egg yolks), marine (like salmon), and plant sources (e.g., broccoli, spinach, avocado, sweet potatoes), as well as whole grains, cereal, dry nuts, and other similar sources [34,35].
In the early 20th century (1932), after the discovery of its all-trans-retinol structure, scientists had focused on isolating vitamin A from several natural sources [36,37]. However, specialized analytical methods had to overcome various obstacles, such as the multiple isomeric retinol forms. Traditional chromatographic and spectroscopic methods concerning vitamin A, vitaminoids, and carotenoids were improved over the years. This is apparent from the development of advanced quantitative analysis techniques, including high-performance liquid chromatography (HPLC) [38,39,40,41], high-resolution mass spectrometry (HRMS) [42], tandem mass spectrometry (MS/MS) [38,43,44], gas chromatography (GC) or coupled with mass spectrometry (GC–MS) [45,46], UV/Vis spectrometry [41,47,48], and liquid chromatography coupled with mass spectrometry (LC–MS) [40,44,49,50,51] and qualitative, non-destructive ones, like the nuclear magnetic resonance (NMR) spectroscopy [18,52,53], near-infrared spectroscopy (NIRS) [54,55], solvent extraction [21,40,49,56,57,58], crystallization [59,60], and antioxidant estimation [47,61,62,63,64,65] methods, widely utilized in several related experiments since [20,56]. More specifically, fat-soluble vitamins like vitamin A, according to the U.S. Food and Drug Administration (FDA), are commonly clinically analyzed via nutrient analysis methods such as UV, HPLC, HPLC-UV, and LC–MS/MS [66]. Plus, enzyme-linked immunosorbent assays (ELISA) are also utilized for the detection and quantification of such bioactives, offering mainly simplicity, compactness, and low analysis costs for routine quality control benefits [3,20]. Interestingly, for retinyl palmitate analysis in fortified oils, a rapid, direct HPLC method was developed, ensuring no time-consuming sample preparation [67].
Due to its ester form, vitamin A is highly soluble in organic solvents but practically insoluble in aqueous solutions, similarly to β-carotene. Liver tissue is rich in vitamin A, due to the high retinol amounts discovered mainly in liver oil, which is derived from marine mammals and fish. Nowadays, by exploiting molecular distillation at extremely low pressure, a widely employed technique in the commercial preparation of vitamin A-rich oils, the ester compounds can be effectively isolated from such oils [68]. Alternatively, vitamin A is easily extracted directly using chloroform or a different solvent mixture like hexane and ethanol and then purified by advanced chromatographic methods [57,58]. Saponification with KOH, followed by organic solvent-assisted extraction, is also used for hydrolyzing esters such as vitamin A, carotenoids, triglycerides, and other lipids [40], while at low temperatures retinol and/or its esters may be crystallized by a variety of organic solvents (e.g., methanol, propylene oxide, ethyl formate) [21,40,49,56].
3.2. Vitamin A-Related Bioactive Compounds: Provitamins (Carotenoids)
Carotenoids are vibrant yellowish–orange–red organic pigments mainly isolated from plants, with remarkable antioxidant activity (only 40 are traced in the diet). These metabolites are tetraterpenoids but may also be metabolized to retinol. In carotenoids, two 20-carbon structures with β-ionone rings and a poly-isoprenoid side chain are linked together, tail-to-tail [16]. Carotenoid molecules can be converted into retinol (provitamin A derivatives) due to having at least one unsubstituted β-ionone ring and are β-carotene, α-carotene, and β-cryptoxanthin, while those unable to be transformed by any metabolic pathway to vitamin A but also owning vital activity are xanthophylls like lutein, lycopene, astaxanthin, zeaxanthin, canthaxanthin, fucoxanthin, phytoene, phytofluene, and capsanthin (Figure 3), with oxygen atoms in their containing hydroxyl groups [3,20].
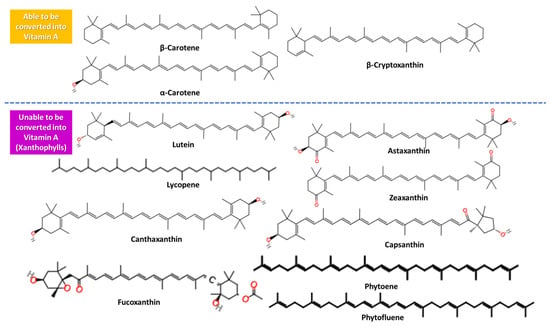
Figure 3.
The structure of carotenoids able and unable to be converted into vitamin A.
Carotenoid content in vegetables, fruits, and plant species is strongly influenced by flesh color, degree of ripeness (fully ripe fruits display higher carotenoid content), method and location of cultivation, post-cultivation modification, storage techniques (sun favors carotenoid loss, while storage in dry places at high temperatures favors their perseverance), intracellular carotenoids’ position, purification method, cooking way, and heat treatment [3,29,30]. Photosynthetic organisms, including plants, herbs, animals, marine organisms, and microorganisms (e.g., bacteria, fungi), may produce carotenoids in different amounts. Red and orange vegetables on a great scale (tomato, red chili pepper, red pepper, carrots, pumpkins, sweet potatoes, etc.), green vegetables (like kale, spinach, broccoli, parsley, lettuce, etc.), mushrooms, fruits such as apricots and mango, herbs like basil, cereals, grains, and spirulina entail high carotenoid levels, with β-carotene being mostly present. α-carotene is similarly to β-carotene obtained via carrots, sweet potatoes, pumpkin, mangoes, broccoli, oranges, and spinach [69], while β-cryptoxanthin, either as a free compound or esterified with fatty acids (lauric, myristic, palmitic), is administered via the same sources as β-carotene and α-carotene (Figure 3) [3,70].
3.3. Indicative Role and Functions of Vitamin A, Its Vitaminoids, and Carotenoids—Vitamin A Deficiency, Hypervitaminosis, and Toxicity
Vitamin A is an essential bioactive metabolite with multiple biological functions in the human body. Initially, it plays a vital role in the cycle of vision primarily in the form of retinal, which partakes in several enzymatic and isomerization reactions. In fact, loss of night vision (night blindness) is associated with vitamin A deficiency [71]. Retinol and ATRA act as signaling molecules for gene expression in vertebrates (i.e., of growth hormones) and are involved in cell growth and differentiation [72]. Retinol is crucial for normal immune system function of both innate and adaptive immunity, while it is an immune booster resulting in antibodies vast response, preserving and reestablishing integration of all mucosal surfaces [73,74]. Moreover, minimal amounts of vitamin A and β-carotene synergistic effects may prevent cancer and scavenge ROS. β-carotene’s anti-cancer and anti-proliferation properties especially delve from its antioxidant profile and conversion into vitamin A [3,75]. Plus, red blood cells require retinoids and vitamin A for proper differentiation, while retinol contributes to spermatogenesis, glycoprotein synthesis, taste and hearing function, and retinoids partake in maintaining normal epithelial homeostasis by promoting the differentiation of keratinocytes into mature epidermal cells [3,10].
Carotenoids, vitamin A, and vitaminoids have emerged as pivotal components in synthesizing nutraceuticals, nutricosmetics, cosmeceuticals, and cosmetics. β-carotene, lutein, and astaxanthin act as potent antioxidants by combating oxidative stress, reducing photoaging, and promoting a healthy complexion. On the other hand, retinoids such as retinol and retinoic acid accelerate skin renewal, reduce hyperpigmentation, and mitigate aging impact. Such vitaminoids extend the scope of cosmetic applications by supporting hydration, skin elasticity, and barrier repair [76]. Their role in the cosmetic field and importance in cosmetic formulations will be further discussed in Section 4.
Intake recommendations on vitamin A differ by age and are typically measured in retinol activity equivalents (RAE). Reportedly, one RAE corresponds to 1 μg of retinol, 12 μg of β-carotene, or 24 μg of α-carotene/β-cryptoxanthin [3,77]. For infants and children, the suggested intake ranges from 400 to 500 RAE; adult males are advised to consume 900 RAE daily, and females, pregnant, and lactating women should consume 700 to 1300 RAE, with lactating women requiring the highest doses. Sufficient vitamin A levels are crucial for proper fetal development and ensuring adequate nutrition for the newborn [3,77].
Vitamin A deficiency impairs vision, and in severe cases, prolonged deficiency may lead to xerophthalmia and even irreversible blindness [71,78]. Such deficiency affects various body systems (inadequate plasma retinol (<0.52 μm) or liver levels (<5–20 μg/g)). The respiratory system displays increased susceptibility to infections due to epithelial changes, skin keratinization, and dryness [3,77], while impaired sperm production and testicular degeneration are observed in the reproductive system [79]. Reduced intestinal goblet cells and alterations in the epithelium lead to poor digestion (gastrointestinal system) [80], while nerve lesions and diminished taste/smell (nervous system) [81], weakened immunity and risk of infection due to malnutrition (immune system) [73], iron metabolism disruption [82], and anemia, particularly in children and pregnant women [83], are only a few indications implying vitamin A deficiency. Either because of a poor diet (primary), malabsorption, or chronic diseases (secondary), hypovitaminosis affects around one-third of children worldwide [3,77]. Vitamin A supplementation [84], biofortification (i.e., transgenic golden rice and fortified samples (e.g., sweet potatoes)) [85], and a balanced diet are effective solutions against hypovitaminosis [3,77].
Hypervitaminosis, otherwise known as vitamin A toxicity, on the contrary, is rare but can occur due to excessive vitamin A or retinoid intake (plasma retinol levels > 2.09 μm). Toxicity plus arises from supplement overuse or vitamin A-rich food overconsumption and may be either acute or chronic. Retinoid acid syndrome, hypertriglyceridemia, teratogenicity, impaired bone health, skin redness, peeling, irritation, and allergic reactions [3,77,86] may follow vitamin A’s toxicity. Plus, excess carotenoid intake can subsequently cause a range of comorbidities, from harmless, easily dissolved carotenoderma (yellow/orange skin) to complications like nephrotic syndrome, cancer, or liver issues [3,25,71,77].
4. Vitamin A, Vitaminoids, and Carotenoids as Nutricosmetic and/or Cosmeceutical and Cosmetic Factors: Absorption, Conversion, Storage, Distribution, Metabolism, and Biochemical Mechanisms of Action
As the human body is unable to produce vitamin A, it is of great importance to obtain it from the diet either in its preformed version (70–90% absorption) or as provitamin A in the form of carotenoids (≤3% absorption), and mainly β-carotene, α-carotene, and β-cryptoxanthin [3,16]. Vitamin A is implicated in several mechanisms of absorption and mainly partakes in oral, skin, parenteral, sublingual, and rectal absorption, which means that additional administration routes, like intramuscular and topical ways besides one’s diet, are also possible. Oral and topical administration pathways of vitamin A are common and will be described in further detail [3,16,77].
4.1. Mechanisms of Absorption, Conversion, Storage-Distribution, and Metabolic Pathways of Vitamin A, Its Vitaminoids, and Carotenoids
4.1.1. Oral Absorption and Metabolism of Vitamin A, Its Vitaminoids, and Carotenoids
Vitamin A is primarily absorbed through the gastrointestinal (GI) tract when consumed orally, while vitamin A and its vitaminoids’ absorption highly differs from that of carotenoids, indicated by the fact that animal-derived retinyl esters are almost completely absorbed, while plant-based carotenoids’ absorption is notably lower [3,87].
As for animal-derived vitamin A and its vitaminoids, predominantly retinyl esters, they reach the intestine, where their metabolization into retinol by a triglyceride lipase or phospholipase B in the intestinal lumen of the GI tract occurs. After being metabolized, retinol and retinoids penetrate into enterocytes along with β-carotene, associate with chylomicrons (ChM), and are finally secreted into the lymphatic system. The uptake of retinol may be initiated either by active transport or by passive diffusion, while fat-soluble retinol’s absorption is increased after consuming fatty meals and micronutrients like zinc, as micelle formation encourages it. Both preformed retinol and retinol occurring from β-carotene transform into retinal, then to retinyl esters, and lastly to retinol, binding to a specific protein, namely cellular retinol-binding protein (CRBP), which is responsible for their intracellular transport and is acquired in two forms: CRBPI (widely expressed) and CRBPII, exclusively expressed in intestinal cells [3,87,88].
Retinol bound to CRBP while in the enterocytes may either be transformed back to retinyl esters bound to ChM and continue its path in the lymphatic system (cascade 1) or be transported directly in the blood systemic circulation via ChM’s assistance, with retinol binding to the retinol-binding protein (Retinol-RBP) (cascade 2). More specifically, following cascade 1, ChM-bound retinyl esters first enter the lymphatic system and then the bloodstream as remnant chylomicrons-retinyl esters (RChM-retinyl esters), where again two different vitamin A pathways are likely to be induced: In the first pathway (cascade 3), RChM-retinyl esters enter the liver, the main retinoid storage body organ with the most vital retinoid-storing tissues/cells. Inside the liver, in the presence of hepatocytes and via the assistance of the RBP, retinyl esters are transformed into retinol and vice versa (“recycling”). Liver-derived retinol and preformed enterocyte-derived retinol are directly secreted into the bloodstream, where they bind to the RBP, transthyretin (thyroid hormone transport function) [89], or later on to other transport proteins like ATRA-bound albumin. Retinols’ transportation in the target tissues is facilitated by the RBP receptor (RBPR), basically the “gate” to the target cell. Vitamin A deficiency is able to be maintained due to stored reserves in the liver (~90%), while circulating retinoids are usually bound to proteins like albumin via the mediation of lipoprotein receptors or stimulation by the retinoic acid 6 receptor (STRA6) [3,87,88].
During the second pathway (cascade 4), RChM-retinyl esters bind directly to the lipoprotein receptor (LR), which is reportedly another “gate” to the target cell, and undergo another conversion to retinol. RBPR and LR-acquired retinol are further oxidized to ATRA, and via the nuclear receptors (NRs), all necessary genetic body functions of vitamin A are initiated (the biochemical vitamin A-related function will be clarified in Section 4.2) [3,16,77,87,88,90]. The cellular and metabolic pathway of orally given vitamin A and its transportation to target cells is depicted in Figure 4.
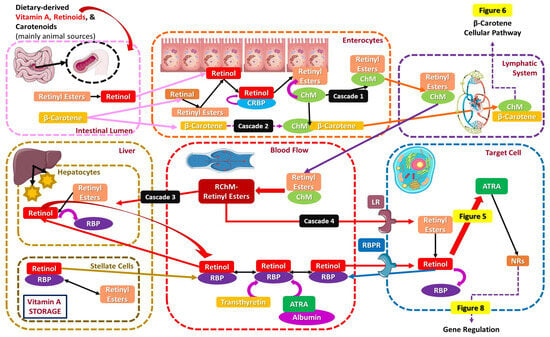
Figure 4.
The cellular pathway of orally given vitamin A and its transportation to target cells.
Retinyl esters, primarily retinyl palmitate, before even entering the enterocytes while in the intestinal lumen, are reversibly converted by esterase into retinol. Retinol, in order to biochemically participate in gene regulation and ROS scavenging, must be previously transformed into ATRA, which interacts with the NRs. Retinol, prior to binding to the CRBP, is similarly metabolized into retinal by retinol dehydrogenases, and then retinal dehydrogenases further oxidize irreversibly retinal into acidic ATRA, the final product of vitamin A oxidation that finally binds to the cellular retinoic acid-binding protein (CRABP). ATRA is not able to be reduced back into retinal and retinol or be stored but can be ultimately isomerized into 9-cis-retinoic acid and 13-cis-retinoic acid (Figure 5) [3,16,77,87,88,90,91].
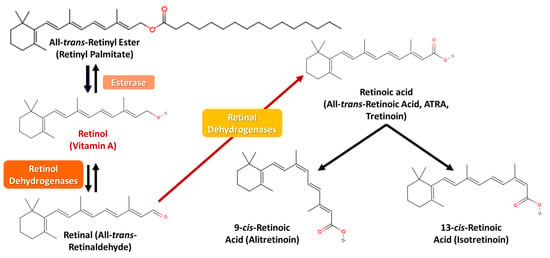
Figure 5.
Vitamin A’s conversion pathway from retinyl palmitate to retinoic acid.
Considering the plant-derived carotenoids, β-carotene is absorbed from the intestinal lumen, predominantly through a passive diffusion cascade or by the mediation of the scavenger receptor class B1 (SCARB1) transporter [92] and the cluster of differentiation 36 (CD36) [93]. Almost one-half of carotenoids are reportedly absorbed unaltered, while the rest are metabolized into retinol, regarding the body’s retinol levels and the diet’s fat content. More specifically, β-carotene, after being absorbed in the intestinal lumen, is transferred in the enterocytes, where either it directly binds to the ChM, surpasses the lymphatic system, and enters the blood flow, or is converted to retinal by β-carotene 15,15′-oxygenase (β-carotene oxygenase 1 (BOC1)) central cleavage (black arrows), which is then either further oxidized into ATRA by aldehyde dehydrogenase 1 family of enzymes (ALDH1s or RALDHS) or reduced to retinol by different forms of alcohol dehydrogenase (ADH) or retinol dehydrogenases (RDH), a process likely to happen in several GI tract locations. ATRA may be further oxidized into a more polar counterpart, namely 4-oxo-retinoic acid (4-oxo-ATRA), by cytochrome P450 (CYP) 26 family enzymes, which is believed to be transcriptionally inactive, while retinol can be further esterified into retinyl esters by the enzyme lecithin retinol acyltransferase (LRAT) [94,95]. ATRA finally, biochemically, partakes in the retinoic acid receptor (RAR) genetic expression [1,3,16,29,77,87,88,91,96]. When β-carotene intake is high, there is only a small conversion rate, and most of its quantity is stored in the adipose tissue and other fat reserves. At this point, it must be clarified that some retinyl esters are metabolized back into retinol, and the remaining are further transported along with the very-low-density (VLDL) and low-density (LDL) lipoproteins (Figure 6 and Figure 7) [3,87,88,97,98].
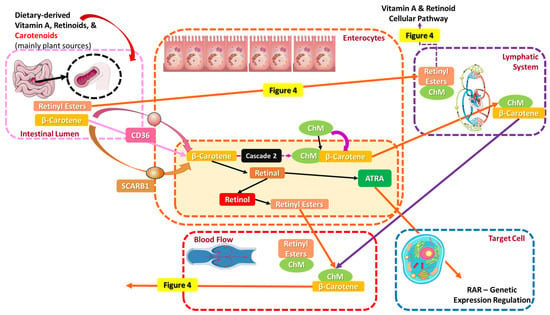
Figure 6.
The cellular pathway of orally given carotenoids and their transportation to target cells.
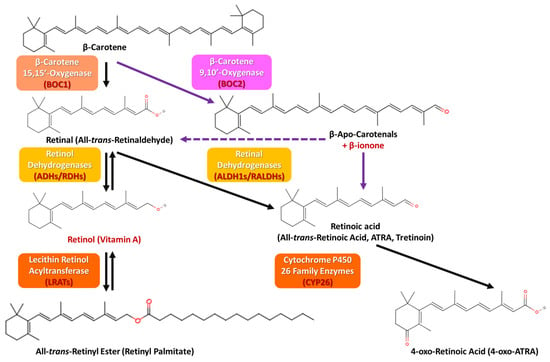
Figure 7.
β-carotene’s conversion and metabolic pathways (the central cleavage is depicted with the black arrows and the eccentric cleavage with the purple arrows).
Apart from the usual cellular pathway depicted in Figure 6, an alternative cleavage procedure occurs with ATRA as the end product, where β-carotene is enzymatically metabolized into β-apo-carotenals (able to ultimately be converted into one molecule of retinaldehyde (purple dashed arrow) and β-ionone by β-carotene 9,10′-oxygenase (or β-carotene oxygenase 2 (BOC2)), eccentric cleavage (purple arrows)) (Figure 7) [98,99]. Although the metabolic cascades are similar for β-carotene, α-carotene, and β-cryptoxanthin, their conversion efficiencies differ due to mainly structural differences, as β-carotene is symmetric and highly efficient (two retinal molecules per molecule), in contrast to α-carotene and β-cryptoxanthin, which are asymmetric and less efficient (1 retinal molecule per molecule) [98,99]. Apart from the main retinoids present in all metabolic pathways concluding in the target cells, namely retinol, retinal, and retinyl esters, ATRA, isotretinoin (“Accutane”), etretinate, and acitretin are also used in oral therapy [3,87,88]. For instance, ATRA has shown great potential towards dermatological conditions like skin cancer, psoriasis, acne, and ichthyosis [100]; isotretinoin is used for treating moderate to severe acne vulgaris [72], while etretinate and acitretin offer UV damage protection and have been suggested as potent therapy for squamous cell carcinoma (SCC) [101]. Isotretinoin, however, may also cause adverse effects such as cell apoptosis by altering the expression of specific genes, an important issue for those suffering from chronic hand eczema, while its isomer, alitretinoin, has not displayed the same impact [3,100]. Excess ATRA is oxidized and slowly evacuates the body via the kidneys or the liver into bile or urine, since retinoids are fat-soluble [3,87,88].
4.1.2. Topical Absorption of Carotenoids, Vitamin A, and Its Vitaminoids
In contrast to oral vitamin A, its vitaminoids, and carotenoid absorption, topically applied vitamin A and its derivatives are absorbed through the skin, targeting the epidermis and dermis. There are several pathways of penetration, involving the transcellular (via keratinized cells of the stratum corneum), intercellular (between cells via lipid bilayers), and appendageal (via hair follicles and sweat glands, which bypass the stratum corneum barrier) routes [102,103]. Retinoids like retinol and tretinoin are often stabilized in formulations with liposomes or nanocarriers for enhanced penetration, due to the fact that skin absorption is boosted by occlusion, hydration, and the presence of penetration enhancers such as ethanol, in order to influence gene transcription, promote collagen synthesis, and improve skin elasticity. The conversion, storage-distribution, and metabolism of vitamin A and its derivatives also differ depending on the absorption route because of the distinct cascades via which orally ingested and topically applied vitamin A compounds enter the body and their subsequent metabolic fates. Skin metabolism is localized, prioritizing direct biochemical action [15,102,103].
Retinol skin-absorbed via any of the above routes is enzymatically converted to retinal and then ATRA in local skin cells, according to the oral route. ATRA directly acts at the site of absorption, binding both to RARs and retinoid X receptors (RXRs) by CRABPs, which are strong modulators of apoptosis, proliferation, differentiation, and cell cycle, influence gene transcription and keratinocyte proliferation, promote collagen synthesis, and improve skin elasticity and barrier function. If ATRA binds to CRABPI, it is transferred to CYP26 to be degraded, while if bound to CRABPII, it is relocated in the nucleus so as to interact with RARs and RXRs, along with several co-activators, and promote transcriptional activity of ATRA target genes [104]. This conversion mainly occurs in the epidermis and dermis, with minimal systemic involvement, and this conversion takes place locally, primarily yielding ATRA for immediate use, in contrast to oral absorption, where this conversion takes place in the intestine and liver [11,100,104]. Meanwhile, carotenoids, including β-carotene, are less commonly utilized topically and are not enzymatically cleaved to vitamin A in the skin [7,19,105]. Retinol, retinoids, and carotenoids are retained locally in the skin cells (and peripheral tissues) and metabolized in situ, while minimum systemic distribution—unless high retinoid concentrations are applied, causing overflow into circulation—occurs (Figure 8) [7,15,19,102,103,106].
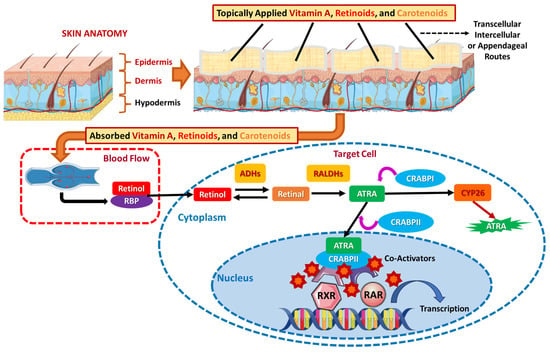
Figure 8.
The cellular pathway of topically administered vitamin A.
4.1.3. Other Absorption Pathways of Vitamin A, Its Vitaminoids, and Carotenoids
Vitamin A can be delivered intramuscularly (IM) as well, with a view to achieving rapid systemic effects, during a parenteral absorption (via injection) pathway [3,21]. IM injections bypass the GI tract, delivering vitamin A directly to the bloodstream, commonly in cases of a malabsorption syndrome or severe deficiency. Once in circulation, retinol binds to the RBP and transthyretin in order to be transported to the target tissues. Liquid formulations provide a convenient way of rapidly addressing vitamin A deficiency, especially when oral supplementation through water or parenteral administration is inefficient, via promoting bioavailability and micronutrient use [3,21].
Vitamin A in sublingual drops (sublingual absorption) bypasses the GI tract by direct absorption into systemic circulation through the sublingual mucosa. Sublingual absorption is remarkably effective for individuals with impaired digestive systems [3,107]. Furthermore, vitamin A suppositories or enemas deliver the nutrient via the rectal mucosa following the rectal absorption pathway, suitable for patients unable to take oral or injectable forms in cases of vitamin A deficiency [3,108].
4.2. Biochemical Mechanisms of Action of Vitamin A, Its Vitaminoids, and Carotenoids: Gene Regulation and Mediation in Thromboinflammatory Pathways
Once metabolized, vitamin A, its vitaminoids, and carotenoids partake in several biochemical pathways and thrombo-inflammatory manifestations, crucial for cellular and physiological functions. Carotenoids and vitamin A derivatives scavenge excessive free radicals during oxidative stress cases and inflammatory manifestations from several genetic, epigenetic, age-, UV-, or dietary-related, photo-oxidative, or concomitant factors. Hence, they provide enhanced antioxidant and anti-inflammatory shielding by affecting the binding, inhibiting the associated signaling, or modulating the metabolism of thrombotic and inflammatory mediators (reduction), reducing inflammatory/oxidative, and increasing anti-inflammatory/antioxidant genes [3,77,88,109].
ATRA, as the end product of the vitamin A, retinoids, and carotenoids’ metabolism, acts as aforementioned, as a ligand via several co-activators and CRABPII binding for NRs, namely RARs that regulate gene expression involved in cell differentiation, proliferation, and apoptosis and RXRs that partner with other receptors (e.g., peroxisome proliferator-activated receptor (PPARs) and VDRs), modulating diverse pathways. Such a pathway of gene expression regulation is vital for embryonic development, immune function, and skin renewal [3,77,87,100]. ATRA is involved in cellular differentiation and apoptosis in epithelial tissues, prevents keratinization disorders, aids in tissue regeneration, and induces apoptosis in aberrant cells, with implications for anti-cancer therapy [3,77,87,100]. Furthermore, ATRA promotes immune homeostasis by regulating T-regulatory cells (T-regs), hence enhancing anti-inflammatory responses and type 1T helper/type 2 T helper (Th1/Th2) balance, by supporting adaptive immunity and affecting pro-inflammatory cytokine formation as well [110]. Lastly, ATRA, when retinol-bound protein 4 (holo-RBP4) is bound, triggers the retinoic acid 6 (STRA6) gene. Once STRA6 is boosted to relocate in the cell membrane, it catalyzes the transport of RBP4’s retinol release. Thus, it sets off tyrosine phosphorylation on the C-terminal tail, which attracts signal transducer and activator of transcription 3 and 5 (STAT 3/5) and Janus kinase 2 (JAK2). Notably, STRA6 partakes in the intrinsic mechanism of p53-mediated apoptosis, occurring after cellular DNA damage or elevated intracellular ROS [111].
Concerning the visual cycle, retinal is a key molecule where it combines with opsin in the retina to form rhodopsin, essential for low-light vision. The cycle involves the conversion of 11-cis-retinal to all-trans-retinal upon light activation and recycling via the retinoid cycle in the cytoplasm of retinal cells, by the aid of several enzymatic reactions, such as retinol dehydrogenase and isomerase enzymes [92]. All-trans-retinal is then reduced by nicotinamide adenine dinucleotide (NADH) to NAD+ and, via ADHs in the liver, is converted to all-trans-retinol. All-trans-retinol is subsequently transformed through isomerases (liver) to 11-cis-retinol, which is then again converted by NAD+ to NADH transformation and ADHs to 11-cis-retinal during the Wald’s visual cycle [112].
As for the antioxidant activity, retinoids and carotenoids like β-carotene scavenge ROS, protecting cells from oxidative stress and damage, supporting cardiovascular (CV) health, skin protection, and immune defense, while also being able to combat oxidative modification of LDLs (oxidized LDLs), thereby mitigating thrombo-inflammatory stimuli and atherogenesis, as well as supporting CV health, through inhibiting the inhibitor of κΒα (IκΒα) pathway [19,113]. Moreover, platelet activating factor (PAF) inhibition is induced by both retinoids and carotenoids, able to counteract the pro-inflammatory effects of PAF, which is implicated in skin inflammation, allergic responses, and vascular changes. PAF via binding to the G-protein-coupled membrane receptors (GPCMRs) and PAF-receptor (PAF-R/PAR signaling), via pro-inflammatory genes and proteins like phospholipase Cβ (PLCβ), phosphatidylinositol 4,5-biphosphate (PIP2), and Gq protein i, γ and β subunits (Gq,i/γ/β) and via partaking in mitogen-activated protein kinase (MAPK), inhibits PAF-cholinetransferase (PAF-CPT) and lysophosphatidylcholine (LPC) acyltransferase (LPCAT) thrombo-inflammation [114,115,116]. Thrombin, eicosanoids (cyclooxygenases (COX)), collagen (type I and II), and adenosine diphosphate (ADP), implicated in several inflammatory cascades, are restrained by carotenoids action against the nuclear factor κ light-chain enhancer of activated B cells (NF-κΒ) and gene expression [105,111,117,118].
Another biochemical pathway retinoids influence is the phosphatidylinositol 3-kinase (PI3K)/protein kinase (Akt) pathway, implicated in cell survival, metabolism, and proliferation. By regulating this cascade, vitamin A, retinoids, and carotenoids contribute to tissue repair (wound healing) and protection against ROS formation and oxidative stress. More specifically, via the kelch-like ECH-associated protein 1 (keap1), an adaptor subunit of cullin 3-based E3 ubiquitin ligase, the activity of the nuclear factor erythroid 2-related factor 2 (Nrf2) that acts as a sensor for oxidative/electrophilic stress via PI3K/Akt is regulated [110,119,120]. All the above biochemical mechanisms of the beneficial action of vitamin A, retinoids, and carotenoids are thoroughly depicted in Figure 9.
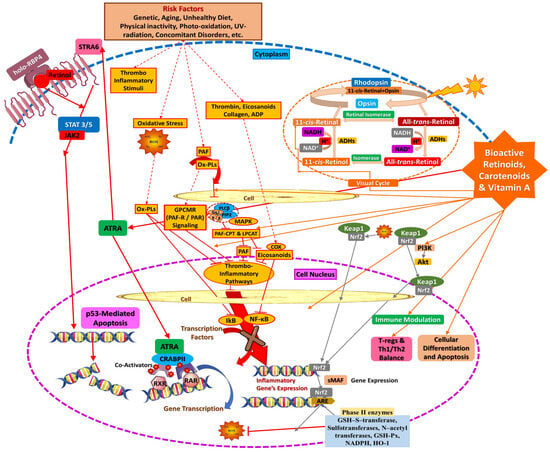
Figure 9.
Biochemical mechanisms of action of vitamin A, its vitaminoids, and carotenoids.
The metabolism of carotenoids, retinoids, and vitamin A prepares and regulates the availability of vitamin A by converting it into active or stored forms, while biochemical cascades describe how these active forms exert their physiological effects. Both processes are interdependent but have distinct purposes in preserving health and cellular function.
5. Vitamin A, Vitaminoids, and Carotenoids’ General Role in Nutricosmetic, Cosmeceutical, and Cosmetic Applications
As aforementioned, both retinoids and carotenoids have a lot of beneficial impact on human health. Regarding carotenoids, more specifically, both their bioactive groups, carotenes and xanthophylls, depending on the presence of oxygen in their structure, own an antioxidant and anti-aging effect, sometimes working synergistically with other antioxidants too. Vitamin A, along with its derivatives, namely retinoid esters and carotenoids, is consumed via dietary supplements or food, providing a variety of sources that could benefit the general growth and maintenance of the immune system. Retinal, for example, a retinoid aldehyde, is responsible for the normal function of the eyes, while retinoic acid acts as a hormone-like growth factor in cells. Other retinoid esters have anti-aging activity, as they are used in the cosmetic field for treating skin’s fine lines and wrinkles [3,16,77]. Deficiency of vitamin A is linked with post-natal growth retardation in children, while in adults it can cause problems for the eyes, immune, or reproductive system [121]. On the other hand, carotenoids, such as zeaxanthin and α/β-carotene, have exhibited bio-functional activity against chronic diseases, like diabetes, cardiovascular diseases (CVDs), cancer, and obesity incidents [122]. They can also be photo-protective, as they are experts in protecting the skin from ROS and lipid peroxyl radicals induced by UV radiation A and B (UVA and UVB, respectively), species that damage cells when incorporated in the DNA, causing mutations and early aging, wrinkles, skin cancer, melanoma, ichthyosis, acne, and other skin-associated issues [123].
Because the human body cannot produce vitamin A de novo, all needed bioactive compounds are obtained following a diet rich with animal- and marine-sourced products but also comprised of fruits and vegetables. Preformed vitamin A (retinol and retinyl esters) is mostly found in animals, while provitamin A carotenoids, including α- and β-carotene, come from colored plants and fruits [69]. In Western countries, a percent bigger than 70% of the daily dose of vitamin A comes from preformed vitamin A in animals, while less than 30% derives from carotenoids in plants, fruits, and vegetables. On the other hand, in developing countries, the opposite prevails [124]. Lately, nutraceuticals, nutricosmetics, and cosmeceuticals have grown an upward interest in providing several bioactive ingredients, like carotenoids and vitamins, as an alternative, healthier way of supplementation instead of drugs for preventing several health conditions, such as skin cancer, or delaying others, like skin aging. The term “nutraceutical” was discovered in 1989 by Dr. Stephen De Felice, as a compound from the words “nutrition” and “pharmaceutical”, while the term “cosmeceutical” originated from Raymond Reed in 1962 but became famous in 1984 from a dermatologist professor named Albert Kligman, in research of retinoic acid as an anti-aging ingredient [125,126]. These categories are placed between nutrition/beauty and pharmaceuticals to provide the bioactive ingredients to the consumers for the cure or prevention of several health issues, with no need for synthetic medications and their potent side effects. Carotenoids in nutraceuticals and/or nutricosmetics provide the same effect as in functional foods, presenting an anti-aging and photo-protective role against UVR in the skin. Interestingly, a supplement mixture of α- and β-carotene as well as lutein has been proven beneficial in photo-protection, while a combination of β-carotene, lutein, and lycopene has been reported as an effective treatment against erythema [125].
However, β-carotene is generally unstable; hence, other vitamin A forms (retinoids) are frequently utilized in cosmetic formulations in the skincare industry. Either applied topically, orally, or systemically, vitamin A, its retinoids, and carotenoids actively participate in protein synthesis, cell division, and cellular metabolism. Normalization of keratinization and plus growth and differentiation control of epithelial cells have been declared as the main advantages of vitamin A (retinol) and its retinoids, including retinyl palmitate, retinyl acetate, retinal, and ATRA, which are widely exploited in the cosmetic field at different concentrations [100,127]. ATRA, otherwise known as tretinoin, is the most bioactive retinoid for modifying skin function, prescribed commonly at high doses, due to the fact that all vitamin A forms metabolize into ATRA, known for reducing inflammation in the sebaceous glands, inhibiting keratosis, and triggering epidermal cell proliferation during psoriasis and/or chronic inflammation-related disorders. ATRA is reported to activate genes that trigger keratinocytes, namely the precursors of immature skin cells, to differentiate into mature epidermal ones [100,127].
Concurrently, other retinoids are also essential counterparts in the nutricosmetics, cosmeceuticals, and cosmetics generation and production department. Retinol reportedly inhibits collagenase, restrains matrix metalloproteinases (MMPs) expression, stimulates glycosaminoglycans’ (GAGS) and collagen type I’s synthesis, and treats skin dyspigmentation, dryness, and anti-wrinkle cases. Finally, retinyl acetate/palmitate and retinal have been recorded to trigger epidermal cell proliferation after their conversion to ATRA as stabilizers in wrinkle treatment. Generally, retinoids downregulate a unique gene, known as cellular communication network factor 1 (CCN1), which is highly expressed in the dermis of photo-aged skin [100,127]. In the following subsections, a more detailed, clinical outcomes-based approach to the main role of vitamin A and its vitaminoids and carotenoids in nutricosmetics, cosmeceuticals, and cosmetics applications is described, while several of these clinical data are included in the comprehensive tables of Section 6.
5.1. Anti-Aging and Photo-Protective Properties of Vitamin A, Retinoids, and Carotenoids
5.1.1. Anti-Aging Effect—Skin Regeneration
Vitamin A and carotenoids have been pointed out to present remarkable anti-aging properties. Skin aging is a result of a plethora of intrinsic and extrinsic factors. Intrinsic reasons are unavoidable (i.e., genetics, the passage of time), but extrinsic factors like UVR exposure, bad diet, stress, or toxins can be avoided. Such factors lead to the breakage of collagen fibers, resulting in early aging [128]. Vitamin A, as an immunomodulatory compound, though, is capable of regulating and proliferating several cells, like skin cells, including keratinocytes and fibroblasts, leading to collagen and elastin synthesis, which is in turn responsible for minimized wrinkles and reversed photo-aging [129]. Retinoids are capable of inhibiting MMPs and modulating gene expression while also regulating the activity of growth factors and cytokines present in complex extracellular matrix protein (ECM) exchange and inflammation [111]. MMPs present in continuous exposure to UVR partake in breaking down the ECM in the dermis, along with collagen and elastin. This situation results in premature photo-aging, with unlikable visual skin spots such as wrinkles and fine lines, dryness, and irregular pigmentation [128]. Retinol and the other retinoids, namely retinal and ATRA, that occur via a sequential conversion (Figure 5), are able to eliminate both intrinsic and extrinsic factors by simultaneously promoting collagen synthesis and preventing collagen breakdown through RARs and RXRs (Figure 8) [111]. Regarding retinoic acid, a study conducted on ATRA confirmed that it can increase types I, II, and III collagen while reorganizing elastic fibers and modulating GAGs of the ECM [117]. Also, ATRA has the capability to enhance epidermis thickness by promoting the growth of epidermal keratinocytes and increasing the formation of endothelial cells and blood vessels in the papillary dermis, all contributing as anti-aging factors for skin protection by visibly decreasing wrinkle formation, skin roughness, and relaxation. As a result, an improvement of the skin’s elasticity, hydration, and resilience refinement, as well as a barrier function enhancement, has been observed with long-term use of vitamin A-based cosmetics [111].
Adding to retinoids beneficial impact, the application of antioxidant and anti-aging substances, like carotenoids, may also aid in the protection of the human skin from environmental factors [130]. The carotenoids present in the human skin are mostly α/β-carotene, zeaxanthin, lycopene, and lutein. In some cases, the synergistic effect of such compounds, along with some vitamins, is highly encouraged as the best way of protecting the skin from aging [123]. For example, in a study where the combination of vitamins E and C with β-carotene was examined, this conjunction of active vitamin forms concluded in an advanced scavenging ability against reactive nitrogen species (RNS), rather than this of single antioxidants [131]. Moreover, a study about the xanthophyll fucoxanthin demonstrated that it can inhibit tyrosinase activity, an enzyme whose hyperactivity leads to hyperpigmentation and skin spots, melanogenesis in melanoma, and UVB-induced skin pigmentation, factors notably contributing to aging [132]. Astaxanthin, additionally, is believed to increase the elasticity factor and improve the skin’s texture while reducing the size of age spots [117]. Furthermore, non-colored carotenoids, namely phytoene and phytofluene, are capable of absorbing UVA and UVB radiation, acting as a photo-protector for the skin, while at the same time lutein and zeaxanthin, also present in the human epidermis, can protect the skin from the blue light and subsequently from early aging [117]. Lycopene, as a very strong antioxidant present mainly in tomatoes, can also act as a natural photo-protective agent towards photo/early aging [133]. The anti-aging activity of different vitamin A forms derives mainly from their antioxidant ability. Besides being photo-protective agents externally in the skin by extensive UVR, carotenoids act as anti-antioxidants as well, by scavenging ROS and covering excited forms of singlet oxygen and triplet-state molecules that could lead to inactivation of vital antioxidant enzymes, lipid peroxidation, and DNA damage, leading to internal protection [117].
Vitamin A deficiency has long been connected to delayed epithelialization and wound healing, which is confirmed by poor wound closure, lower collagen synthesis rates, and lower levels of cross-linking of newly generated collagen. The efficacy of retinoids or carotenoids’ topical application and/or oral consumption appears to be comparable. As previously stated, retinoids function by binding to certain receptors both in the cytoplasm and in the nucleus, which remarkably impact RNA, protein synthesis, lysosome-membrane stability, cell growth, differentiation, and finally division. A widely recognizable sensitivity impact of retinoids is their exceptional capacity to counteract the anti-inflammatory steroids’ inhibitory effects on the healing process, aside from wounds’ contraction. Administering a retinoid therapy partially but significantly reverses the adverse effects of the inflammatory response, tensile strength, and collagen build-up in cutaneous wounds in individuals following steroid treatment [118].
Recent research outcomes supported that topical retinol applied to aged human skin has been pointed out to dramatically increase epidermis thickness by inducing epidermal keratinocyte proliferation, as well as to enhance endothelial cell and blood vessel proliferation in the papillary dermis, by thickening the epidermal layer and leading to the formation of new blood vessels in the dermis [111]. Type I collagen, along with tropoelastin and fibronectin expression, and the formation of collagenous ECM in aged human skin in vivo due to activating dermal fibroblasts, are drastically elevated when topical retinol therapy is applied. Reportedly, vascularity and epidermal thinning are important factors that contribute to skin weakness and restrain rapid wound healing in trauma cases in aged skin. Topical retinol improves the dermal microenvironment by encouraging the growth of vasculature via endothelial cell proliferation in aged human skin, in addition to boosting ECM’s generation. Several reports on age-related decrease in the cutaneous vasculature have been recorded, where topical retinol treatment was confirmed to promote dermal vascularity, which is able to enhance skin blood flow, as well as dermal and epidermal homeostasis [111].
5.1.2. Photo-Protective Effect
Early aging is directly linked with UVR exposure, as aforementioned. UVR generally consists of two categories: UVA (~320–400 nm) and UVB (~290–320 nm), where carotenoids specifically absorb in this spectrum due to their structure that is comprised of conjugated double bonds. Skin damage induced by UVR, in turn, is split into two different sections: (a) acute, including cell necrosis, erythema, and inflammation, and (b) chronic, involving photo-aging with dermatological signs like wrinkles, skin spots, dryness, fine lines, and also the formation of skin cancer. UVA penetrates the deeper dermis, and its main function is to promote early aging by activating ROS, but also inducing cell apoptosis, erythema, and carcinogenesis. UVB is absorbed by keratinocytes in the epidermis and is more erythematogenic than UVA, hence contributing to cancer genesis by interacting with nucleus molecules (DNA and RNA), causing mutation. Phytoene and phytofluene offer unique protection against UV radiation because of their specific absorption properties. These carotenoids are distinct from most of their counterparts that require at least seven conjugated double bonds to exhibit color and absorb light in the visible spectrum, as they possess shorter polyene chains [8,134,135]. Figure 10 explains the results of UVR exposure to aging and demonstrates some selected carotenoids and retinoids that have been extensively studied as a cure for these comorbidities.
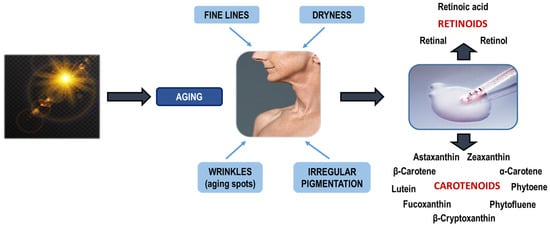
Figure 10.
Sources, bioactivity, and skin protection properties of carotenoids.
An alternative way of treating or preventing skin damage, except by following a diet rich in carotenoids, is by using supplements, orally or topically. Clinical trials conducted in vivo with several carotenoid supplements have revealed photo-protection against the sun’s radiation. More specifically, a study on 11 men and 11 women consuming a carotenoid mixture of α-/β-carotene and lutein, by enhancing the dose to a final 90 mg per day for 8 weeks, revealed a moderate dose-dependent increase in minimal erythema dose (MED), a decrease in lipid peroxidation, and an increase in β-carotene in the serum, but not in the skin. In another similar study, 20 healthy women were administered a moderate dose of β-carotene per day for 10 weeks and then were exposed to natural sun radiation for 13 days, without stopping this administration. Study outcomes supported that an erythema reduction was observed, along with a yellow pigmentation on the skin surface. The predominant study conclusion was that pre-treatment and treatment with carotenoids during sun exposure treatment displayed a protective effect against sunburn by raising the Langerhans cells’ level and minimizing the erythema. Lastly, another similar study with two groups receiving two different supplement mixtures was tested. The first one took a mix of α-/β-carotene, cryptoxanthin, zeaxanthin, and lutein per day, while the other group was given the same mix but with the addition of vitamin E, per day for 12 weeks. After the treatment, both teams revealed yellowish skin and higher concentrations of β-carotene in their serum and skin. The level of erythema was higher in those who took only the mix of carotenoids, while those who were administered the mixture with the addition of vitamin E displayed higher protection against UVR, revealing the possible synergistic effects of carotenoids with other antioxidants [128].
5.2. Antioxidant and Anti-Inflammatory Profile of Vitamin A, Retinoids, and Carotenoids as Immunodefensive Mechanisms
Carotenoids effectively neutralize harmful free radicals able to impair skin cells and contribute to premature aging. Such antioxidant capacity stems from their unique molecular structure, featuring a long chain of conjugated double bonds (polyene chain) that can readily donate electrons to stabilize free radicals; hence, the number of conjugated double bonds determines their antioxidant capacity [19,136,137]. Lycopene with 11 conjugated double bonds owns the highest antioxidant efficiency, followed by α-carotene, β-cryptoxanthin, β-carotene, zeaxanthin, and lutein [19]. Vitamin A and carotenoids act as very effective quenchers of singlet oxygen, both in vivo and in vitro, by reacting with free radicals, breaking them down into active degradation products [16].
Notably, dietary supplementation with carotenoids has been proved to increase the antioxidant status of the skin, resulting in a lower concentration of free radicals generated in the skin after sunlight exposure to a simulator [19,138]. Moreover, carotenoids may also enhance the activity of the skin’s endogenous antioxidant enzymes and have the ability to combat ROS and RNS [134,139]. Topical application of several carotenoids, primarily as an ingredient in cosmetic preparations, increases carotenoid levels in the stratum corneum, the outermost layer of the skin, preventing skin damage caused by free radicals from UV rays and other factors [127,139]. At this point it must be noted that the less unsaturated, colorless carotenoids, namely phytoene and phytofluene, in contrast to the more saturated lycopene, have not displayed as exceptional an antiradical ability, even though they have been proved as great skin-whitening agents [8]. As a result, β-carotene, for instance, has been confirmed as a great antioxidant candidate in sunscreens, with a great role against sun-induced lipid peroxidation [61], while a recent crossover study demonstrated that lutein capsules stabilized by 10% of the antioxidant carnosic acid may protect from photodamage by reducing UVR-modulated gene expression like heme-oxygenase 1 (HO-1), intercellular adhesion molecule 1 (ICAM-1), and matrix metallopeptidase 1 (MMP1) genes [8,140]. Interestingly, oral supplementation with a red paprika product led to notable MED and hydration enhancement and a reduction in skin tanning in skin previously exposed to UV on the back [141].
At the same time, the anti-inflammatory properties of carotenoids stem from their ability to modulate cellular signaling pathways involved in inflammation. Carotenoids can interact with the NF-κΒ pathway, inhibiting its activation and subsequent production of inflammatory mediators. They can also reduce the expression of inflammatory markers such as interleukin 24 (IL-24) in keratinocytes [19,29,113,136]. Oxidative stress, induced by factors like UV radiation and pollution, is a major driver of inflammation in the skin. By valuably scavenging free radicals and quenching singlet oxygen, carotenoids aid in reducing oxidative stress, thereby mitigating the inflammatory cascade [142]. Additionally, some carotenoids, like β-carotene, can be metabolized into retinoids, which are known for their potent anti-inflammatory effects in the skin [8,143]. Retinoids can regulate keratinocyte differentiation, reduce sebum production, and modulate immune responses, contributing to the overall anti-inflammatory benefits of carotenoids in cosmetics. Carotenoid structure allows them to neutralize ROS and absorb UV radiation, offering protection from environmental damage such as pollution and sun exposure, benefits that translate into an enhanced skin tone and a strengthened skin barrier [8,143].
5.3. Hyperpigmentation Improvement Activity of Vitamin A, Retinoids, and Carotenoids
Following the path of this vitamin’s photo-protective and anti-aging profile, retinol has shown great potential towards several disorders and skin-related comorbidities. Hyperpigmentation is characterized by topical darkening of the skin, otherwise known as skin discoloration, resulting from excess melanin production in certain skin areas, which often appears due to excessive exposure to the sun, aging, skin inflammation, melasma, etc., and is usually a worldwide concern of women, as its appearance on the skin is not aesthetically pleasing. ATRA is one of the main adaptable solutions to this problem, while a plethora of unique mechanisms contribute to reducing the pigmentation spots on the skin. ATRA can balance the production of melanin by reducing tyrosinase activity, a valuable enzyme for melanin development and its transportation to keratinocytes. Moreover, it exfoliates pigmented cells, leaving room for fresh skin appearance with less pigmentation. Additionally, ATRA has an anti-inflammatory ability, aiding in alleviating post-inflammation hyperpigmentation spots by encouraging skin cell turnover. Lastly, it contributes to collagen activation, providing better skin texture, fewer spots, and an overall youthful complexion. Other retinoids like retinal, tretinoin, adapalene, and tazarotene have also been proved effective in topical skin treatments [111]. Also, the xanthophyll fucoxanthin can inhibit tyrosinase activity, an enzyme whose over-activity results in hyperpigmentation and skin spots, as mentioned for early aging factors [132].
5.4. Acne Treatment Activity of Vitamin A, Retinoids, and Carotenoids
Acne vulgaris is a very common skin condition, affecting many teenagers and younger individuals, that occurs because of hair follicle clogging under the skin with oil, dead skin, or bacteria and appears on the face, neck, shoulders, upper chest, and back. Reasons triggering the onset of acne include inflammation, immunological causes, hypercolonization of microorganisms, follicular hyper-keratinization, and sebaceous hyperplasia. Other factors stimulating acne may also be habitual, including smoking, bad diet patterns, or hormonal alterations [129]. Several antioxidant and anti-inflammatory sources can be valuable in treating acne vulgaris, providing better skin texture. Interestingly, blackberries, watermelon, aloe vera, and papaya preserve skin-regenerating properties, as they contain many bioactives like the carotenoids β-cryptoxanthin and β-carotene [129]. Furthermore, strong retinoid forms such as retinal, tretinoin, adapalene, and tazarotene are more effective than retinol and retinyl palmitate, providing advanced acne skin treatment [111]. Lastly, naphthalene-carbozylic acid, as another vitamin A derivative, has been applied to the skin and was confirmed as an effective acne treatment solution that reduces inflammation and hyper-keratinization in hair follicles [127].
Only severe acne that has not responded to topical treatment and adequate antibiotics, however, should be treated with isotretinoin. Isotretinoin possesses a strong anti-inflammatory profile, as it lessens the formation of comedones by effectively decreasing hyper-keratinization, sebum generation, and Propionibacterium acnes colonization of the pilosebaceous duct. P. acnes is able to induce the immune system’s inflammatory reaction. Isotretinoin treatment is thus responsible for inhibiting monocyte expression of the toll-like receptor-2 (TLR2), which lowers inflammatory cytokines’ formation in response to P. acnes, with a six-month lasting impact. Adapalene, tazarotene, and tretinoin are topically applied retinoids, authorized for moderate acne treatment as well. In addition to restraining micro-comedones’ development, the precursor lesion of comedones, they also display anti-inflammatory properties. Adapalene and tazarotene, especially, bind selectively only to the β and γ types of RARs, whereas tretinoin binds to all three types. In a clinical trial, where tretinoin 0.1% microsponges were contrasted to tazarotene 0.1% gel, tazarotene was reported to exhibit similar tolerability and better efficacy in treating facial mild-to-moderate acne vulgaris [144]. Concurrently, adapalene gels (0.3%) demonstrated superior tolerability and comparable efficacy over a ten-week period when compared with tretinoin cream 0.05% in a clinical trial evaluating the safety and performance of these products in patients with mild-to-moderate acne [145].
5.5. Psoriasis Treatment with Vitamin A, Retinoids, and Carotenoids
Psoriasis is an inflammation-associated skin condition affecting 2–3% of the global population with symptoms like itchiness, redness, burning feeling, and soreness. It is a long-lasting immune disease that appears in the skin and/or joints, with lesions mostly located on the skin surface of the knees, elbows, or scalp region. People with psoriasis were deficient in vitamins, especially D and A. The synergistic effect of these vitamins may have a better outcome in treating psoriasis than either single-treatment therapy [146]. In vivo and in vitro studies have globally taken place to examine the anti-psoriasis effect of acitretin, apremilast, and tazarotene retinoids. Acitretin reverses both increased cell proliferation and keratinization in psoriasis, resulting in skin thickness, plaque formation, and scaling reduction. Meanwhile, tazarotene is a pro-drug with a significant role in regulating skin damage, psoriasis, and acne. None of the mechanisms of action of the two retinoids, though, has been well illustrated yet [127,146]. ATRA has also exhibited anti-psoriasis activity, while astaxanthin can inhibit the production of the inducible nitric oxide (iNOS), making them possible candidates for the development of anti-psoriasis and other inflammatory skin condition drugs [130,147].
5.6. Skin and Other Cancers’ Treatment Potential of Vitamin A, Retinoids, and Carotenoids
Skin cancer is another result of over-exposure to the sun’s radiation. Photo-protection is once again the solution to avoid this undesirable and mostly irreversible condition, operated either topically or systematically. The main aim of this treatment solution is the achievement of UV light absorption by certain bioactive groups, like phenols, vitamins, and carotenoids. Such antioxidants may prevent UVR by penetrating the skin and causing the formation of ROS, which eventually leads to skin cancer and photo-carcinogenesis, along with other side effects. Both in vivo-animal model and in vitro studies have proved the protective impact of carotenoids against skin cancer. Astaxanthin also inhibited skin cancer and tyrosinase over-activity in a rat model, while a trial among Australian volunteers showed that a diet rich in lutein and zeaxanthin was associated with a decrease of SCC in people with a family background of skin cancer (Figure 11) [8,130].
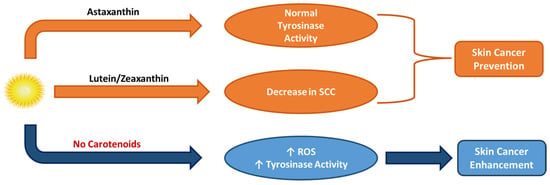
Figure 11.
Activity of astaxanthin, lutein, and zeaxanthin carotenoids in skin cancer prevention.
Briefly, similar studies conducted on other retinoids and carotenoids have implied their notable potential towards several cancer disorders. Meals enriched with β-carotene were effective against esophagus cancer, lutein and zeaxanthin minimized the risk of breast cancer, and lycopene had a lower risk of prostate cancer and heart failure, while retinoid precursors prevent cancer in sunburn reactions [18]. Carotenoid and retinol treatment in human breast, cervix, prostate, colorectal, oral cancer, and leukemia has been confirmed highly effective [16]. A lower incidence of epithelial lung cancer and respiratory diseases (acute lung injury) was also observed in patients receiving a high carotenoid intake that restrains ROS and induces apoptosis in tumor cells [3,123,124].
6. Vitamin A and Its Derivatives of Several Sources: Health-Promoting Skin Benefits for Nutricosmetic, Cosmeceutical, and Cosmetic Applications
Retinoids and carotenoids have multiple benefits and have great clinical importance towards several diseases, disorders, and comorbidities. Lutein as a nutraceutical and nutricosmetic is able to mitigate the likelihood of ocular complications like cataract, retinitis pigmentosa, and age-related macular degeneration (AMD) [123,148]. Regarding hyperhomocysteinemia, coronary artery dyslipidemia, diabetes mellitus (type II mainly), hyperhomocysteinemia, atherosclerosis, metabolic disorders, CVDs, steatohepatitis, and neurodegenerative diseases, carotenoids modulate pro-inflammatory mediators (e.g., cytokines), thereby lowering blood pressure and improving insulin sensitivity [124].
Several health-related bioactive compounds are traced in functional foods, including fruits, vegetables, animal-based, marine-associated, and many microorganism-related products, naturally rich in bioactive compounds like carotenoids (e.g., tomatoes, citrus fruits, eggs, fish). Nutricosmetics, cosmeceuticals, and cosmetics are supplementary or topical formulations generated to deliver such bioactives directly via controlled dosage for health or aesthetic benefits. Carotenoids like β-carotene, α-carotene, β-cryptoxanthin, astaxanthin, zeaxanthin, lutein, lycopene, fucoxanthin, phytoene, and phytofluene are highlighted as key players in skin health [109,149,150]. A vital role of carotenoids is their antioxidant capacity, due to combating oxidative stress by neutralizing ROS and helping cellular damage. Meanwhile, these constituents are also photo-protective against harmful UV radiation, hence reducing photo-dermatoses (against Erythropoietic protoporphyria (EPP), Porphyria cutanea tarda (PCT), premature skin aging-cancer, and polymorphic light eruption (PMLE)) risk, as well as sun-induced erythema, photo-carcinogenesis, photo-aging, pigmentation, and melanogenesis disorders like melasma. Lastly, carotenoids reduce inflammatory responses, benefiting chronic inflammatory skin conditions and improving skin appearance because of their anti-inflammatory and immunomodulatory properties against conditions such as atopic dermatitis (AD) [109,149,150]. The sources of carotenoids and retinoids, their bioactivity, and their potential role towards effective skin protection are shown in Figure 12, while the most common carotenoids along with their sources and bioactive properties are displayed in Table 1, Table 2, Table 3 and Table 4.
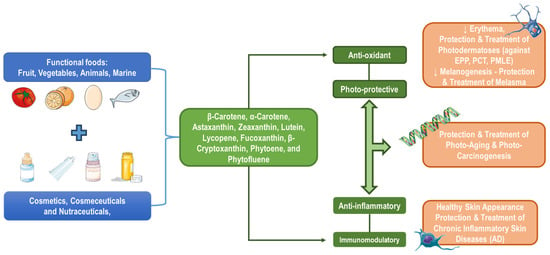
Figure 12.
Indicative sources, bioactivity, and skin protection properties of carotenoids.
At this point, it must be clarified that most animals, marine mammals, plant herbs, and microorganisms mainly produce carotenoids and not retinoids directly. However, our previous knowledge leads to the conclusion that β-carotene and other carotenoids are consumed either orally or topically and then are converted by the human body, following established metabolic pathways, to retinol and retinoids. This explains why only a few clinical trials over mainly the past decade have evaluated animal-, marine-, plant-, herb-, and microorganism-derived retinoid oral supplements or topically applied products [3].
6.1. Vitamin A, Vitaminoids, and Carotenoids of Animal Origin
Animal-sourced carotenoids found, for instance, in hens producing eggs, are namely lutein, zeaxanthin, β-cryptoxanthin, and β-carotene [122,151]. In milk, β-carotene seems to be the main carotenoid, while lutein, zeaxanthin, and β-cryptoxanthin are present in lower amounts [151]. Fried beef for human consumption displays higher bioavailability of preformed vitamin A (retinol) than that of provitamin A (carotenoids) like β-carotene in vegetables, with a range of 74% and 2–65%, respectively, while hen, chicken, or quail eggs also have displayed a notable carotenoid content [152].
Other carriers of carotenoids, which are also used for protecting the human skin, are insects. Insects receive their carotenoids like β-carotene, β-cryptoxanthin, lutein, and zeaxanthin from their food, as they cannot produce them themselves. However, aphids and whiteflies can synthesize carotenoid products de novo [153]. Birds, frogs, lizards, and snails are also carriers of carotenoids, β-carotene, β-cryptoxanthin, lutein, and astaxanthin, giving them their colored skin [153]. Similarly, in humans, β-cryptoxanthin, lutein, and β-carotene, but also α-carotene and zeaxanthin, constitute more than 90% of the total carotenoids, which mostly belong to the skin part of the palms and forehead of humans [130,153,154]. Clinical data mainly from the last decade regarding experiments conducted to prove the benefits of vitamin A, vitaminoids, and carotenoids derived from animal sources (predominantly milk, dairy, and eggs/egg yolks) in many nutricosmetics, cosmeceuticals, and cosmetics applications are demonstrated in Table 1.

Table 1.
Clinical data of the beneficial use of carotenoids, vitamin A, and its vitaminoids of animal origin in nutricosmetics, cosmeceuticals, and cosmetics applications.
6.2. Vitamin A, Vitaminoids, and Carotenoids of Marine Origin
Marine use in the cosmetic field appeared to escalate in the past few years because of their antioxidant, anti-aging, anti-wrinkle, and anti-acne properties. Several carotenoids derive from Halophilic archaea, specifically Halobacterium salinarum and Haloferax mediterranei, and have exhibited a notable anti-aging and antioxidant effect, while other animal products, like sea cucumber extracts, contain vitamins A and B, collagen, gelatin, etc., and have demonstrated a promising wound healing, antioxidant, and antimicrobial impact in treating skin issues, wrinkles, and sunburn [109,149,150]. Some carotenoids produced by halophilic organisms are phytoene, phytofluene, β-carotene, lycopene, derivatives of bacterioruberin, and salinixanthin [163].
Cyanobacteria also contain carotenoids, like astaxanthin and fucoxanthin, and thus, display antioxidant and photo-protective properties by enhancing skin elasticity and promoting collagen synthesis, respectively [109,149,150]. Brown seaweeds (Phaeophyceae), like Laminaria japonica and Laminaria digitate, and diatoms (Bacillariophyta), are also vital sources of fucoxanthin [130,132,153]. Other producers of astaxanthin seem to be the microalgae Haematococcus pluvialis and the fungus Phaffia rhodozyma [131,149,164]. Marine bacteria are also able to synthesize carotenoids, astaxanthin, and zeaxanthin, also showing anti-aging properties, preventing macular degeneration, as confirmed by Mesoflavibacter zeaxanthinifaciens, Zeaxanthinibacter enoshimensis, Muricauda lutaonensis, and several others [23]. Such marine microorganisms, along with lutein and tunaxanthin, are also traced in pink-fleshed fish like tuna and salmon, but also in shrimp, lobster, trout, and other mollusks and crustaceans [109,149,150], while β-carotene is produced as well by the microalgae strains, Dunaliella salina and Dunalielle bardawii [165]. Clinical data about the benefits of vitamin A, vitaminoids, and carotenoids from marine sources (mainly shellfish, algae, sea cucumbers, bacteria, and archaea) in nutricosmetics, cosmeceuticals, and cosmetics applications are included in Table 2.
Table 2.
Clinical data of the beneficial use of carotenoids, vitamin A, and its vitaminoids of marine origin in nutricosmetics, cosmeceuticals, and cosmetics applications.
6.3. Vitamin A, Vitaminoids, and Carotenoids of Plant and Herb Origin
Between animal-, plant-sourced, and herb-derived products, the latter engage a bigger part as suppliers of carotenoids. For humans, carotenoids are obtained through their diet, as they cannot produce them de novo and are responsible for the vibrant yellow, orange, red, and green colors of many fruits and vegetables displayed [136]. Firstly, lycopene is found in red fruits like tomatoes and watermelons, but also in pink grapefruits, apricots, papayas, and guavas [123,164]. α-carotene is a significant component in carrots, red pepper, corn, cloudberry, pumpkin, and coleslaw, and concurrently, β-carotene is present in carrots, apricot, citrus, pineapple, papaya, red pepper, red beans, celery, cucumber, lettuce, blueberry, melon, mustard, and numerous other fruits and vegetables, and β-cryptoxanthin is contained in citrus fruit like tangerine and orange but also sweet red pepper, peach, and papaya. Lutein and zeaxanthin, furthermore, are acquired from yellow fruits but also from leafy green–yellow vegetables, broccoli, corn, lettuce, parsley, etc. Lastly, the uncolored carotenoids phytoene and phytofluene are present in tomato, watermelon, papaya, peach, etc. [123,130,164].
The bioactive profile of carotenoids is mostly the same, presenting predominantly antioxidant, anti-inflammatory, and anti-aging properties. Research, however, has already shifted its focus towards displaying both their individual and synergistic effects in conjunction with compounds of the same bioactive group or other active vitaminoids, like vitamin E or K derivatives [130]. Clinical data regarding the significance of vitamin A, vitaminoids, and carotenoids from plants and herbs in nutricosmetics, cosmeceuticals, and cosmetics formulations are depicted in Table 3.
Table 3.
Clinical data of the beneficial use of carotenoids, vitamin A, and its vitaminoids of plant and herb origin in nutricosmetics, cosmeceuticals, and cosmetics applications.
6.4. Vitamin A, Vitaminoids, and Carotenoids from Microorganism Sources
Microbial-derived sources provide a sustainable, scalable platform for producing carotenoids and retinoids without relying on animal, plant, herb, or marine sources. More specifically, except for microorganisms of animal, plant, or marine origin that have been included in the previous sections, other microbial-associated sources can be cultivated in controlled bioreactors with optimized conditions, yielding high-purity carotenoids and retinoid derivatives for nutricosmetics, cosmeceuticals, and cosmetics applications. Microorganisms, including bacteria, fungi, yeast, and microalgae, represent a cutting-edge solution for meeting increasing demands in the cosmetic industry, particularly for products targeting skin photo-protection and anti-aging [18,149].
Filamentous fungi, especially ascomycetes, generate secondary vitamin A metabolites such as β-carotene, while a heterothallic zygomycetes fungus formulates lycopene with significant coloring, antimicrobial, anti-cancer, antioxidant, anti-proliferative, antiradical, and cytotoxic activity. In Blakeslea trispora fungus, the biosynthesized β-carotene and is utilized widely in biotechnological processes for microbial stimulation and oxidative stress prevention, while other Mucor circinelloides strains and Fusarium sporotrichioides species yield lycopene and β-carotene, have the potential to produce mycotoxins, enhance health, and lower cardiovascular/circulatory disease risk [18,106,213,214]. Moreover, yeast-derived microorganisms like Rhodotorula spp. (e.g., Rhodotorula glutinis) are prolific producers of carotenoids like β-carotene, torulene, and torularhodin, with several antioxidant and photo-protective properties in nutraceuticals and nutricosmetics, while others like Phaffia rhodozyma generate β-carotene and astaxanthin, with vital skin photo-protection and anti-aging properties [48,106,215].
Bacteria producing microorganisms, such as Paracoccus spp., Dietzia natronolimnaea, Croceibacterium aestuarii, Micrococcus luteus, and Gordonia jacobaea, are capable of producing astaxanthin, canthaxanthin, and zeaxanthin and also own antioxidant, anti-inflammatory, and anti-aging properties. Additionally, non-marine microalgae (cultured in controlled systems), including Chlorella zofingiensis and Coelastrella spp., produce β-carotene, astaxanthin, and canthaxanthin under stress conditions and exhibit beneficial skin photo-protection, anti-aging, and antioxidant protection [106]. Finally, retinoid-producing microorganisms like engineered bacteria (e.g., Escherichia coli) that produce retinol and yeast (e.g., Saccharomyces cerevisiae) that convert carotenoids such as β-carotene to retinoids like retinol and retinoic acid in controlled fermentation systems also possess several benefits [106,216,217]. Clinical data regarding the significance of vitamin A, vitaminoids, and carotenoids from microorganisms in nutricosmetics, cosmeceuticals, and cosmetics products are included in Table 4.
Table 4.
Clinical data of the beneficial use of carotenoids, vitamin A, and its vitaminoids from microorganism sources in nutricosmetics, cosmeceuticals, and cosmetics applications.
7. Recent Advances in the Delivery Systems of Vitamin A and Its Derivatives in Nutricosmetics, Cosmeceuticals, and Cosmetics Applications
Vitamin A, retinoids, and carotenoids are incorporated into a variety of formulations for topical, oral, and injectable applications, depending on their intended use in nutricosmetics, cosmeceuticals, and cosmetics. Considering vitamin A and retinoids in topical formulations, creams and serums containing retinol, retinal, and tretinoin are commonly included in anti-aging and acne treatment products as stabilizers against oxidation and as agents for skin penetration enhancement [111,242]. Similarly, carotenoids, including β-carotene, lutein, or lycopene, have been utilized in topical creams and lotions for photo-protection and antioxidant benefits enhancement, which are best achieved when such formulations use liposomes, nanoemulsions, or microencapsulation for stability and better skin delivery [15,243]. Apart from creams and serums, retinol and retinoids have also been utilized in higher concentrations in gels and ointments for conditions like psoriasis and severe acne [244,245], while they have been incorporated in microneedle patches able to deliver them into deeper skin layers [246].
Oral vitamin A and retinoid supplementation is accomplished by the consumption of capsules, soft gels, or tablets that are comprised of retinyl esters or retinol for systemic benefits, including addressing vitamin A deficiency or enhancing skin and eye health [11,247,248]. Furthermore, injectable retinol-based formulations are also preferred during vitamin A deficiency [21,74]. Following vitamin A supplementation, nutraceuticals in the form of capsules or powders contain carotenoids like β-carotene, lutein, zeaxanthin, or lycopene as dietary supplements and nutricosmetics for skin protection, eye health, and antioxidant support [53,249]. Functional foods and beverages infused with carotenoids such as astaxanthin or lycopene for oral consumption have been confirmed as well to be great antioxidants, anti-proliferatives, and anti-aging agents [250,251].
However, the increase in the resilience and complications of several disorders has called for the improvement in the delivery systems of both vitamin A and its derivatives. Advanced delivery systems include formulations tailored to the bioavailability, stability, and specific health benefits of the active compounds, ensuring their efficacy across a range of applications. Retinol, retinoids, and carotenoids have been incorporated in nanocarriers like nanoemulsions and liposomes as nanoscale delivery systems so as to enhance stability, skin absorption, and targeted delivery [15]. Moreover, hydrogels, biodegradable polymers, and emulsions have also been exploited in topical applications for controlled release and sustained effects of retinol, retinoids, and carotenoids [244,245]. Finally, microencapsulation has also been utilized for enclosing and rapidly delivering vitamin A, retinoids, and carotenoids to target cells (Figure 13) [15].
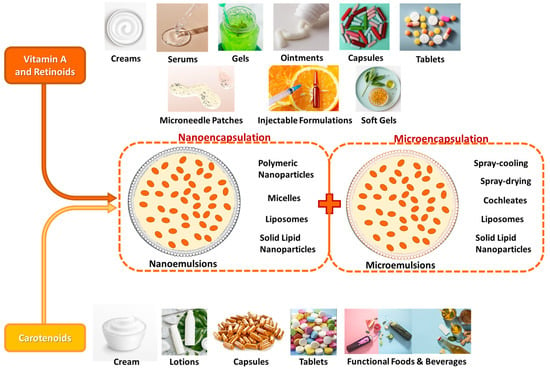
Figure 13.
Different formulations, microencapsulation, and nanoencapsulation for systemic delivery of vitamin A, retinoids, and carotenoids.
Microencapsulation and nanoencapsulation are two encapsulation technologies currently utilized for vitamin transportation, and many attempts towards overcoming their chemical instability have been conducted. The procedure of coating or enclosing tiny solid, liquid, or gas particles in a continuous polymeric layer is microencapsulation. The coated substance, also known as the payload, internal phase, fill, core material, or actives, can be encapsulated alone or in conjunction with additional substances, while the coating material is also referred to as an encapsulating agent, wall material, shell, or carrier and frequently takes the form of combined materials. Since vitamin A is a hydrophobic, fat-soluble compound, it can rapidly break down in the presence of water-based solutions, while it is poorly soluble in aqueous solutions due to its low polarity. Furthermore, it is an extremely sensitive substance that reacts with a variety of other substances, including oxidants, temperature, trace metals, light, heat, and moisture [15,252,253,254]. By selecting the correct encapsulation method, vitamin A can be added to carriers with favorable chemical and physical characteristics to boost its remedial ability, bioavailability, dispersity, controlled release, accurate delivery at the intended spot, and maximum absorbance. Importantly, with a view to achieving the desired encapsulation efficiency, microparticle stability, and essential final product qualities in accordance with its applicability, the right encapsulating agent must be selected, which should not react with the core material, be eco-friendly, and be non-toxic. Spray-cooling, spray-drying, cochleates, coacervation (phase separation), emulsion systems, liposomes, and solid lipid nanoparticles are widely appealing microencapsulation techniques, while the most utilized microencapsulation method for carotenoids is spray-drying technology, with multiple studies globally documenting using a variety of biopolymers as encapsulating materials [244,245,255,256,257]. For instance, Arabic gum has been pointed out as a promising encapsulating agent due to its capacity to generate stable microemulsions of sufficient solubility and low viscosity for carotenoids’ delivery (Figure 13) [257,258].
Nanoencapsulation has also been proven as a highly efficacious approach for improving carotenoid’s chemical stability, thereby safeguarding their bioactivity and stability against potential external influences throughout diverse processing and storage conditions. This method refers to the entrapment of active compounds like vitamin A, its vitaminoids, and carotenoids within nanoscale carriers including liposomes, polymeric nanoparticles, micelles, and solid-lipid nanoparticles for targeted and controlled delivery of bioactives, improved skin penetration, protection from degradation and sun exposure, reduced toxicity and irritation, enhanced stability, antioxidant activity, and bioavailability, as well as effective treatment of retinol deficiency, wound healing, and skin disorders (i.e., acne in topical, oral, or systemic delivery) [15,197,252,259,260]. The most frequently used nanoencapsulation methods are nanoemulsions of vitamin A and carotenoids, namely colloidal dispersions with nanometer-sized droplets (20–200 nm) in a continuous phase, stabilized with surfactants. Nanoemulsions improve the solubility of lipophilic compounds like retinol, enhance their stability, amplify their bioavailability, and promote rapid penetration, while being created via ultrasonication, high-pressure homogenization, and/or micro-fluidization (Figure 13) [15,197,252,259,260].
8. Limitations and Future Perspectives of Vitamin A, Vitaminoids, and Carotenoids in Nutricosmetics, Cosmeceuticals, and Cosmetics Applications
Despite the recognized potential of vitamin A, its vitaminoids, and carotenoids in several scientific fields, certain limitations hinder their widespread application in nutricosmetic, cosmeceutical, and cosmetic formulations. Vitamin A, vitaminoids, and carotenoids are highly sensitive to light, heat, and oxygen; thus, they are prone to oxidative degradation and loss of potency during formulation, storage, and use (photodegradation) [3,136,261]. Due to their chemical instability [262], vitamin A and its derivatives also demonstrate chemical instability that requires the use of advanced encapsulation and stabilizers, which may subsequently increase production costs and manufacturing complexity. Moreover, such compounds are lipophilic, resulting in poor solubility in aqueous systems and limited absorption in the GI, while their low skin penetration is also an important disadvantage [105,127]. Additionally, several adverse effects follow systemic retinoids and carotenoids excessive usage in some cases, counting skin irritation, oxidative stress, elevated cancer risk, hypervitaminosis, carotenemia [123], build-up of tolerance, dermatological issues (e.g., erythema) [100], and concurrently, inconsistent clinical efficacy, questionable bioavailability, safety aspects, and environmental sustainability play a significant role in boundless utilization [53,125,127].
Concerns regarding the lack of robust and recent clinical evidence to support the anti-aging, photo-protective, and antioxidant claims of many retinoids and carotenoids pose another significant limitation. Many strategies have been put out to slow down the aging process and rejuvenate older skin, but as of right now, there are no 100% safe and effective treatment solutions to prevent skin atrophy in older people [242,263]. More specifically, the cellular and molecular processes ATRA uses to prevent aging are currently unknown exactly, while there is a significant lack of studies examining the molecular basis of topical retinol anti-aging benefits in aged human skin, in contrast to ATRA. As a consequence, it has been proven challenging to comprehend the molecular pathways by which retinoids exacerbate aging in human skin, due to an absence of suitable in vivo and in vitro models [111]. Numerous studies evaluating retinoids and carotenoids suffer from methodological limitations like small sample sizes, inappropriate control groups, little to no rigorous scientific backing, and a lack of standardization, which create uncertainty regarding the true efficacy of retinoids currently used in cosmetics [242,263].
The future of vitamin A and its derivatives in cosmetics lies in addressing these limitations and exploring new frontiers. With a view to overcoming all limitations and bettering the clinical experience for patients, many advances, primarily in the delivery systems of vitamin A and vitaminoids, are necessary. More specifically, nanotechnology improvements, including nanoemulsions, liposomes, solid lipid nanoparticles, and micelles, offer promising solutions for developing more stable, bioavailable systems capable of controlled release of such compounds [15,125,127,197,242,253,254]. For instance, retinyl retinoate, retinyl N-formyl aspartamate, and other chemically stable ester derivatives are being thoroughly investigated to enhance resistance to degradation [242]. Biopolymeric encapsulation, antioxidants encapsulation, light-protective packaging, and hybrid delivery systems utilizing hydrogels for encapsulating such bioactives may further optimize targeted delivery and enhance stability for topical and oral applications [244,245]. Furthermore, advances in bioengineered sources, microbial fermentation, genetic engineering of microorganisms, and synthetic biology may enable the production of carotenoids and retinoid-like compounds from non-animal, plant, herb, or marine sources. Hence, sustainable plant-based and microbial-derived agents can offer more sustainable, scalable, and ethical alternatives [1,18,142]. Additionally, combination therapies of vitamin A derivatives with other bioactive compounds, such as antioxidants (e.g., vitamin E, coenzyme Q10 (CoQ10)) or peptides, may enhance synergistic effects in anti-aging and photo-protection formulations [14,26,80,123,243].
Personalized/customized formulations capable of targeting specific skin concerns based on an individual’s profile, more in vivo and in vitro clinical trials, and exploration of other novel applications of vitamin A and its derivatives such as epigenetics [264,265], anti-inflammatory treatment [73,84], and skin microbiome modulation [11,266] are only a few scientific approaches needed to be made prior to these bioactives’ systemic use.
9. Conclusions
Vitamin A and its derivatives, including retinoids and carotenoids, represent a cornerstone in skin health, offering many benefits in nutricosmetic, cosmeceutical, and cosmetic applications. Such bioactives have an unparalleled efficacy in skin rejuvenation, photo-protection, and anti-aging. Retinoids like retinol and mainly retinoic acid enhance collagen synthesis, accelerate epidermal turnover, and repair UV-induced damage by upregulating repair enzymes and inhibiting MMPs. Meanwhile, carotenoids like β-carotene and lycopene exhibit antioxidant properties, scavenge ROS, and mitigate oxidative stress, crucial for preventing photo-aging and inflammation. Topical formulations, including creams, serums, and nanocarrier-based systems, ensure targeted delivery, addressing diverse skin concerns such as hyperpigmentation, acne, and elasticity loss. Moreover, nutraceutical supplementation with β-carotene or astaxanthin during several topical applications offers promising skin-health-related benefits by integrating carotenoids into human skin tissues, boosting systemic antioxidant defenses, and enhancing the skin’s resilience and tolerance against environmental aggressors.
Despite their remarkable benefits for skin health, aging, and other health aspects, vitamin A, vitaminoids, and carotenoids face limitations primarily related to stability, bioavailability, and adverse effects. Advances in nanotechnology, synthetic biology, and personalized approaches offer promising solutions for overcoming these challenges. By focusing on sustainable production, robust clinical validation, enhancing delivery systems, and developing encapsulation technologies, these bioactives hold immense potential to effectively drive the future of the cosmetic field. With ongoing research into novel delivery systems and synergistic formulations, vitamin A derivatives remain pivotal towards advancing skin health and solidifying their role as indispensable agents in nutricosmetics, cosmeceuticals, and cosmetics.
Author Contributions
Conceptualization, A.T.; methodology, A.T.; software, all authors; validation, A.T.; investigation, A.T., T.A. and M.-P.L.; writing—original draft preparation, all authors; writing—review and editing, A.T. and T.A.; visualization, A.T., T.A. and M.-P.L.; supervision, A.T.; project administration, A.T. All authors have read and agreed to the published version of the manuscript.
Funding
This research received no external funding.
Acknowledgments
The authors like to thank the School of Chemistry of the Faculty of Sciences of Democritus University of Thrace for the continuous support.
Conflicts of Interest
The authors declare no conflicts of interest.
References
- Martínez-Cámara, S.; Ibañez, A.; Rubio, S.; Barreiro, C.; Barredo, J.-L. Main Carotenoids Produced by Microorganisms. Encyclopedia 2021, 1, 1223–1245. [Google Scholar] [CrossRef]
- Steenbock, H. White Corn vs. Yellow Corn and a Probable Relation Between the Fat-Soluble Vitamine and Yellow Plant Pigments. Science 1919, 50, 352–353. [Google Scholar] [CrossRef] [PubMed]
- Carazo, A.; Macáková, K.; Matoušová, K.; Krčmová, L.K.; Protti, M.; Mladěnka, P. Vitamin A Update: Forms, Sources, Kinetics, Detection, Function, Deficiency, Therapeutic Use and Toxicity. Nutrients 2021, 13, 1703. [Google Scholar] [CrossRef]
- Shastak, Y.; Pelletier, W. Pet Wellness and Vitamin A: A Narrative Overview. Animals 2024, 14, 1000. [Google Scholar] [CrossRef]
- Baswan, S.M.; Klosner, A.E.; Weir, C.; Salter-Venzon, D.; Gellenbeck, K.W.; Leverett, J.; Krutmann, J. Role of Ingestible Carotenoids in Skin Protection: A Review of Clinical Evidence. Photodermatol. Photoimmunol. Photomed. 2021, 37, 490–504. [Google Scholar] [CrossRef]
- Crupi, P.; Faienza, M.F.; Naeem, M.Y.; Corbo, F.; Clodoveo, M.L.; Muraglia, M. Overview of the Potential Beneficial Effects of Carotenoids on Consumer Health and Well-Being. Antioxidants 2023, 12, 1069. [Google Scholar] [CrossRef]
- Biskanaki, F.; Kalofiri, P.; Tertipi, N.; Sfyri, E.; Andreou, E.; Kefala, V.; Rallis, E. Carotenoids and Dermoaesthetic Benefits: Public Health Implications. Cosmetics 2023, 10, 120. [Google Scholar] [CrossRef]
- Meléndez-Martínez, A.J.; Stinco, C.M.; Mapelli-Brahm, P. Skin Carotenoids in Public Health and Nutricosmetics: The Emerging Roles and Applications of the UV Radiation-Absorbing Colourless Carotenoids Phytoene and Phytofluene. Nutrients 2019, 11, 1093. [Google Scholar] [CrossRef] [PubMed]
- Flieger, J.; Raszewska-Famielec, M.; Radzikowska-Büchner, E.; Flieger, W. Skin Protection by Carotenoid Pigments. Int. J. Mol. Sci. 2024, 25, 1431. [Google Scholar] [CrossRef]
- VanBuren, C.A.; Everts, H.B. Vitamin A in Skin and Hair: An Update. Nutrients 2022, 14, 2952. [Google Scholar] [CrossRef] [PubMed]
- Milani, M.; Colombo, F. Skin Anti-Aging Effect of Oral Vitamin A Supplementation in Combination with Topical Retinoic Acid Treatment in Comparison with Topical Treatment Alone: A Randomized, Prospective, Assessor-Blinded, Parallel Trial. Cosmetics 2023, 10, 144. [Google Scholar] [CrossRef]
- Cho, Y.S.; Kim, H.O.; Woo, S.M.; Lee, D.H. Use of Dexpanthenol for Atopic Dermatitis—Benefits and Recommendations Based on Current Evidence. J. Clin. Med. 2022, 11, 3943. [Google Scholar] [CrossRef] [PubMed]
- Stettler, H.; de Salvo, R.; Olsavszky, R.; Nanu, E.A.; Dumitru, V.; Trapp, S. Performance and Tolerability of a New Topical Dexpanthenol-Containing Emollient Line in Subjects with Dry Skin: Results from Three Randomized Studies. Cosmetics 2021, 8, 18. [Google Scholar] [CrossRef]
- Temova Rakuša, Ž.; Roškar, R. Quality Control of Vitamins A and E and Coenzyme Q10 in Commercial Anti-Ageing Cosmetic Products. Cosmetics 2021, 8, 61. [Google Scholar] [CrossRef]
- Li, Y.; Zhao, Y.; Zhang, H.; Ding, Z.; Han, J. The Application of Natural Carotenoids in Multiple Fields and Their Encapsulation Technology: A Review. Molecules 2024, 29, 967. [Google Scholar] [CrossRef]
- Kim, J.A.; Jang, J.-H.; Lee, S.-Y. An Updated Comprehensive Review on Vitamin A and Carotenoids in Breast Cancer: Mechanisms, Genetics, Assessment, Current Evidence, and Future Clinical Implications. Nutrients 2021, 13, 3162. [Google Scholar] [CrossRef]
- Villar, A.; Vázquez-González, I.; Vicente, F.; Salcedo, G.; González, L.; Botana, A.; Royo, L.J.; Eguinoa, P.; Busqué, J. Study of the Variability in Fatty Acids and Carotenoid Profiles: Laying the Ground for Tank Milk Authentication. Sustainability 2021, 13, 4506. [Google Scholar] [CrossRef]
- Sharma, C.; Kamle, M.; Kumar, P. Microbial-Derived Carotenoids and Their Health Benefits. Microbiol. Res. 2024, 15, 1670–1689. [Google Scholar] [CrossRef]
- Darvin, M.E.; Lademann, J.; von Hagen, J.; Lohan, S.B.; Kolmar, H.; Meinke, M.C.; Jung, S. Carotenoids in Human Skin In Vivo: Antioxidant and Photo-Protectant Role against External and Internal Stressors. Antioxidants 2022, 11, 1451. [Google Scholar] [CrossRef]
- Shastak, Y.; Pelletier, W.; Kuntz, A. Insights into Analytical Precision: Understanding the Factors Influencing Accurate Vitamin A Determination in Various Samples. Analytica 2024, 5, 54–73. [Google Scholar] [CrossRef]
- Shastak, Y.; Pelletier, W. Review of Liquid Vitamin A and E Formulations in Veterinary and Livestock Production: Applications and Perspectives. Vet. Sci. 2024, 11, 421. [Google Scholar] [CrossRef]
- Zhang, H.; Ren, X.; Yang, Z.; Lai, J. Vitamin A Concentration in Human Milk: A Meta-Analysis. Nutrients 2022, 14, 4844. [Google Scholar] [CrossRef] [PubMed]
- Górska-Warsewicz, H.; Rejman, K.; Laskowski, W.; Czeczotko, M. Milk and Dairy Products and Their Nutritional Contribution to the Average Polish Diet. Nutrients 2019, 11, 1771. [Google Scholar] [CrossRef] [PubMed]
- Stadnik, J. Nutritional Value of Meat and Meat Products and Their Role in Human Health. Nutrients 2024, 16, 1446. [Google Scholar] [CrossRef] [PubMed]
- Khan, R.U.; Khan, A.; Naz, S.; Ullah, Q.; Puvača, N.; Laudadio, V.; Mazzei, D.; Seidavi, A.; Ayasan, T.; Tufarelli, V. Pros and Cons of Dietary Vitamin A and Its Precursors in Poultry Health and Production: A Comprehensive Review. Antioxidants 2023, 12, 1131. [Google Scholar] [CrossRef]
- Ionov, I.A.; Katerinich, O.O.; Kuchmistov, V.O.; Anisimova, O.V.; Griffin, D.K.; Romanov, M.N.; Zhukova, I.O. Vitamin E and A Availability in Goose Embryos and Goslings and Improvement of Reproduction Traits Depending on the Starting Temperature Regime of Egg Incubation. Poultry 2023, 2, 305–319. [Google Scholar] [CrossRef]
- Wu, P.; Zhang, L.; Jiang, W.; Liu, Y.; Jiang, J.; Kuang, S.; Li, S.; Tang, L.; Tang, W.; Zhou, X.; et al. Dietary Vitamin A Improved the Flesh Quality of Grass Carp (Ctenopharyngodon Idella) in Relation to the Enhanced Antioxidant Capacity through Nrf2/Keap 1a Signaling Pathway. Antioxidants 2022, 11, 148. [Google Scholar] [CrossRef]
- Cabezuelo, M.T.; Zaragozá, R.; Barber, T.; Viña, J.R. Role of Vitamin A in Mammary Gland Development and Lactation. Nutrients 2020, 12, 80. [Google Scholar] [CrossRef] [PubMed]
- Álvarez, R.; Meléndez-Martínez, A.J.; Vicario, I.M.; Alcalde, M.J. Carotenoid and Vitamin A Contents in Biological Fluids and Tissues of Animals as an Effect of the Diet: A Review. Food Rev. Int. 2015, 31, 319–340. [Google Scholar] [CrossRef]
- Darwish, W.S.; Ikenaka, Y.; Morshdy, A.E.; Eldesoky, K.I.; Nakayama, S.; Mizukawa, H.; Ishizuka, M. β-Carotene and Retinol Contents in the Meat of Herbivorous Ungulates with a Special Reference to Their Public Health Importance. J. Vet. Med. Sci. 2016, 78, 351–354. [Google Scholar] [CrossRef]
- Shastak, Y.; Pelletier, W. Vitamin A in Fish Well-Being: Integrating Immune Strength, Antioxidant Capacity and Growth. Fishes 2024, 9, 330. [Google Scholar] [CrossRef]
- Tarricone, S.; Ragni, M.; Carbonara, C.; Giannico, F.; Bozzo, F.; Petrontino, A.; Caputi Jambrenghi, A.; Colonna, M.A. Growth Performance and Flesh Quality of Sea Bass (Dicentrarchus Labrax) Fed with Diets Containing Olive Oil in Partial Replacement of Fish Oil—With or Without Supplementation with Rosmarinus Officinalis L. Essential Oil. Animals 2024, 14, 3237. [Google Scholar] [CrossRef] [PubMed]
- Dajnowska, A.; Tomaszewska, E.; Świątkiewicz, S.; Arczewska-Włosek, A.; Dobrowolski, P.; Domaradzki, P.; Rudyk, H.; Brezvyn, O.; Muzyka, V.; Kotsyumbas, I.; et al. Yolk Fatty Acid Profile and Amino Acid Composition in Eggs from Hens Supplemented with SS-Hydroxy-ß-Methylbutyrate. Foods 2023, 12, 3733. [Google Scholar] [CrossRef]
- Bjørklund, G.; Shanaida, M.; Lysiuk, R.; Butnariu, M.; Peana, M.; Sarac, I.; Strus, O.; Smetanina, K.; Chirumbolo, S. Natural Compounds and Products from an Anti-Aging Perspective. Molecules 2022, 27, 7084. [Google Scholar] [CrossRef]
- Kawczak, P.; Feszak, I.; Brzeziński, P.; Bączek, T. Structure–Activity Relationships and Therapeutic Applications of Retinoids in View of Potential Benefits from Drug Repurposing Process. Biomedicines 2024, 12, 1059. [Google Scholar] [CrossRef]
- Karrer, P.; Morf, B.; Schöpp, K. Isolation of Vitamin A. Nature 1932, 129, 88. [Google Scholar] [CrossRef]
- Holmes, H.N.; Corbet, R.E. The Isolation of Crystalline Vitamin A1. J. Am. Chem. Soc. 1937, 59, 2042–2047. [Google Scholar] [CrossRef]
- Lizama, C.; Romero-Parra, J.; Andrade, D.; Riveros, F.; Bórquez, J.; Ahmed, S.; Venegas-Salas, L.; Cabalín, C.; Simirgiotis, M.J. Analysis of Carotenoids in Haloarchaea Species from Atacama Saline Lakes by High Resolution UHPLC-Q-Orbitrap-Mass Spectrometry: Antioxidant Potential and Biological Effect on Cell Viability. Antioxidants 2021, 10, 1230. [Google Scholar] [CrossRef] [PubMed]
- Parra-Rivero, O.; Paes de Barros, M.; del Mar Prado, M.; Gil, J.-V.; Hornero-Méndez, D.; Zacarías, L.; Rodrigo, M.J.; Limón, M.C.; Avalos, J. Neurosporaxanthin Overproduction by Fusarium Fujikuroi and Evaluation of Its Antioxidant Properties. Antioxidants 2020, 9, 528. [Google Scholar] [CrossRef] [PubMed]
- Słowik-Borowiec, M.; Głąb, N.; Stach, S.; Szpyrka, E. A Miniaturized Sample Preparation Method for the Determination of Vitamins A and E in Food Products. Molecules 2023, 28, 3449. [Google Scholar] [CrossRef]
- Średnicka-Tober, D.; Kopczyńska, K.; Góralska-Walczak, R.; Hallmann, E.; Barański, M.; Marszałek, K.; Kazimierczak, R. Are Organic Certified Carrots Richer in Health-Promoting Phenolics and Carotenoids than the Conventionally Grown Ones? Molecules 2022, 27, 4184. [Google Scholar] [CrossRef] [PubMed]
- Di Lorenzo, R.; Castaldo, L.; Sessa, R.; Ricci, L.; Vardaro, E.; Izzo, L.; Grosso, M.; Ritieni, A.; Laneri, S. Chemical Profile and Promising Applications of Cucurbita Pepo L. Flowers. Antioxidants 2024, 13, 1476. [Google Scholar] [CrossRef]
- Rojas-Garbanzo, C.; Gleichenhagen, M.; Heller, A.; Esquivel, P.; Schulze-Kaysers, N.; Schieber, A. Carotenoid Profile, Antioxidant Capacity, and Chromoplasts of Pink Guava (Psidium guajava L. Cv. ‘Criolla’) during Fruit Ripening. J. Agric. Food Chem. 2017, 65, 3737–3747. [Google Scholar] [CrossRef]
- Suo, A.; Fan, G.; Wu, C.; Li, T.; Cong, K. Green Extraction of Carotenoids from Apricot Flesh by Ultrasound Assisted Corn Oil Extraction: Optimization, Identification, and Application. Food Chem. 2023, 420, 136096. [Google Scholar] [CrossRef] [PubMed]
- Staropoli, A.; Vassetti, A.; Salvatore, M.M.; Andolfi, A.; Prigigallo, M.I.; Bubici, G.; Scagliola, M.; Salerno, P.; Vinale, F. Improvement of Nutraceutical Value of Parsley Leaves (Petroselinum crispum) upon Field Applications of Beneficial Microorganisms. Horticulturae 2021, 7, 281. [Google Scholar] [CrossRef]
- Hegazy, G.E.; Abu-Serie, M.M.; Abo-Elela, G.M.; Ghozlan, H.; Sabry, S.A.; Soliman, N.A.; Abdel-Fattah, Y.R. In Vitro Dual (Anticancer and Antiviral) Activity of the Carotenoids Produced by Haloalkaliphilic Archaeon Natrialba sp. M6. Sci. Rep. 2020, 10, 5986. [Google Scholar] [CrossRef] [PubMed]
- Thevamirtha, C.; Balasubramaniyam, A.; Srithayalan, S.; Selvakumar, P.M. An Insight into the Antioxidant Activity of the Facial Cream, Solid Soap and Liquid Soap Made Using the Carotenoid Extract of Palmyrah (Borassus flabellifer) Fruit Pulp. Ind. Crops Prod. 2023, 195, 116413. [Google Scholar] [CrossRef]
- Dyaa, A.; Soliman, H.; Abdelrazak, A.; Samra, B.N.; Khojah, E.; Ahmed, A.F.; El-Esawi, M.A.; Elsayed, A. Optimization of Carotenoids Production from Rhodotorula sp. Strain ATL72 for Enhancing Its Biotechnological Applications. J. Fungi 2022, 8, 160. [Google Scholar] [CrossRef] [PubMed]
- Pan, Q.; Shen, M.; Yu, T.; Yang, X.; Li, Q.; Zhao, B.; Zou, J.; Zhang, M. Liquid Chromatography as Candidate Reference Method for the Determination of Vitamins A and E in Human Serum. J. Clin. Lab. Anal. 2020, 34, e23528. [Google Scholar] [CrossRef] [PubMed]
- Liu, H.; Zhang, C.; Zhang, X.; Tan, K.; Zhang, H.; Cheng, D.; Ye, T.; Li, S.; Ma, H.; Zheng, H. A Novel Carotenoids-Producing Marine Bacterium from Noble Scallop Chlamys Nobilis and Antioxidant Activities of Its Carotenoid Compositions. Food Chem. 2020, 320, 126629. [Google Scholar] [CrossRef]
- Rhenals-Montoya, P.; Villamil, L.; Sánchez-Suárez, J.; Díaz, L.; Coy-Barrera, E. Optimized Carotenoid Production and Antioxidant Capacity of Gordonia Hongkongensis. Sci. Prog. 2024, 107, 1–33. [Google Scholar] [CrossRef] [PubMed]
- Mata-Gómez, L.C.; Mapelli-Brahm, P.; Meléndez-Martínez, A.J.; Méndez-Zavala, A.; Morales-Oyervides, L.; Montañez, J. Microbial Carotenoid Synthesis Optimization in Goat Cheese Whey Using the Robust Taguchi Method: A Sustainable Approach to Help Tackle Vitamin A Deficiency. Foods 2023, 12, 658. [Google Scholar] [CrossRef] [PubMed]
- Rodriguez-Amaya, D.B.; Esquivel, P.; Meléndez-Martínez, A.J. Comprehensive Update on Carotenoid Colorants from Plants and Microalgae: Challenges and Advances from Research Laboratories to Industry. Foods 2023, 12, 4080. [Google Scholar] [CrossRef]
- Rosales, A.; Crossa, J.; Cuevas, J.; Cabrera-Soto, L.; Dhliwayo, T.; Ndhlela, T.; Palacios-Rojas, N. Near-Infrared Spectroscopy to Predict Provitamin A Carotenoids Content in Maize. Agronomy 2022, 12, 1027. [Google Scholar] [CrossRef]
- Soulat, J.; Andueza, D.; Graulet, B.; Girard, C.L.; Labonne, C.; Aït-Kaddour, A.; Martin, B.; Ferlay, A. Comparison of the Potential Abilities of Three Spectroscopy Methods: Near-Infrared, Mid-Infrared, and Molecular Fluorescence, to Predict Carotenoid, Vitamin and Fatty Acid Contents in Cow Milk. Foods 2020, 9, 592. [Google Scholar] [CrossRef]
- Beshgetoor, D. Handbook of Vitamins. Am. J. Clin. Nutr. 2002, 75, 955. [Google Scholar] [CrossRef]
- Vállez-Gomis, V.; Carchano-Olcina, S.; Azorín, C.; Benedé, J.L.; Chisvert, A.; Salvador, A. Simultaneous Quantification of Vitamin A and Derivatives in Cosmetic Products by Liquid Chromatography with Ultraviolet Detection. Separations 2022, 9, 40. [Google Scholar] [CrossRef]
- Gradinaru, I.; Ciubotaru, B.I.; Butnaru, M.; Cojocaru, F.D.; Covașă, C.T.; Bibire, T.; Dascalu, M.; Bargan, A.; Cazacu, M.; Zaltariov, M.-F. The Impact of the Addition of Vitamins on a Silicone Lining Material to the Oral Mucosa Tissue—Evaluation of the Biocompatibility, Hydrolytic Stability and Histopathological Effect. Medicina 2023, 59, 1936. [Google Scholar] [CrossRef]
- Bas, T.G. Bioactivity and Bioavailability of Carotenoids Applied in Human Health: Technological Advances and Innovation. Int. J. Mol. Sci. 2024, 25, 7603. [Google Scholar] [CrossRef]
- Jiang, Y.; Ye, J.; Hu, Y.; Zhang, J.; Li, W.; Zhou, X.; Yu, M.; Yu, Y.; Yang, J.; Yang, W.; et al. Extraction and Synthesis of Typical Carotenoids: Lycopene, β-Carotene, and Astaxanthin. Molecules 2024, 29, 4549. [Google Scholar] [CrossRef] [PubMed]
- Freitas, J.V.; Junqueira, H.C.; Martins, W.K.; Baptista, M.S.; Gaspar, L.R. Antioxidant Role on the Protection of Melanocytes against Visible Light-Induced Photodamage. Free. Radic. Biol. Med. 2019, 131, 399–407. [Google Scholar] [CrossRef] [PubMed]
- De Oliveira, G.L.R.; Medeiros, I.; da Cruz Nascimento, S.S.; Viana, R.L.S.; Porto, D.L.; Rocha, H.A.O.; Aragão, C.F.S.; Maciel, B.L.L.; de Assis, C.F.; de Araújo Morais, A.H.; et al. Antioxidant Stability Enhancement of Carotenoid Rich-Extract from Cantaloupe Melon (Cucumis melo L.) Nanoencapsulated in Gelatin under Different Storage Conditions. Food Chem. 2021, 348, 129055. [Google Scholar] [CrossRef] [PubMed]
- Paliwal, C.; Ghosh, T.; Bhayani, K.; Maurya, R.; Mishra, S. Antioxidant, Anti-Nephrolithe Activities and in Vitro Digestibility Studies of Three Different Cyanobacterial Pigment Extracts. Mar. Drugs 2015, 13, 5384–5401. [Google Scholar] [CrossRef] [PubMed]
- Mohana, D.C.; Thippeswamy, S.; Abhishek, R.U. Antioxidant, Antibacterial, and Ultraviolet-Protective Properties of Carotenoids Isolated from Micrococcus Spp. Radiat. Prot. Environ. 2013, 36, 168. [Google Scholar] [CrossRef]
- Giani, M.; Gervasi, L.; Loizzo, M.R.; Martínez-Espinosa, R.M. Carbon Source Influences Antioxidant, Antiglycemic, and Antilipidemic Activities of Haloferax Mediterranei Carotenoid Extracts. Mar. Drugs 2022, 20, 659. [Google Scholar] [CrossRef] [PubMed]
- MAN-000064; QMS Standard Operating Procedure (SOP) Template for Laboratory Operations. Food and Drug Administration Office of Regulatory Affairs, Office of Regulatory Science: Silver Spring, MD, USA, 2022; pp. 1–34.
- Rimkus, G.G.; Schubert, M.; Morgan, D.; Jungjohann, S. Rapid Direct Analysis of Retinyl Palmitate (Vitamin A) in Fortified Vegetable Oils by HPLC-FLD. Food Addit. Contam. Part A 2022, 39, 24–34. [Google Scholar] [CrossRef] [PubMed]
- Han, L.; Zhang, S.; Qi, B.-K.; Li, H.; Xie, F.-Y.; Li, Y. Molecular Distillation-Induced Deacidification of Soybean Oil Isolated by Enzyme-Assisted Aqueous Extraction: Effect of Distillation Parameters. Appl. Sci. 2019, 9, 2123. [Google Scholar] [CrossRef]
- González-Peña, M.A.; Ortega-Regules, A.E.; Anaya de Parrodi, C.; Lozada-Ramírez, J.D. Chemistry, Occurrence, Properties, Applications, and Encapsulation of Carotenoids—A Review. Plants 2023, 12, 313. [Google Scholar] [CrossRef] [PubMed]
- Olmedilla-Alonso, B.; Rodríguez-Rodríguez, E.; Beltrán-de-Miguel, B.; Estévez-Santiago, R. Dietary β-Cryptoxanthin and α-Carotene Have Greater Apparent Bioavailability Than β-Carotene in Subjects from Countries with Different Dietary Patterns. Nutrients 2020, 12, 2639. [Google Scholar] [CrossRef]
- Sajovic, J.; Meglič, A.; Glavač, D.; Markelj, Š.; Hawlina, M.; Fakin, A. The Role of Vitamin A in Retinal Diseases. Int. J. Mol. Sci. 2022, 23, 1014. [Google Scholar] [CrossRef] [PubMed]
- Cardoso-Demartini, A.A.; Boguszewski, C.L.; Boguszewski, M.C.S. Potential Effects of Oral Isotretinoin on Growth Plate and Height. Endocrines 2023, 4, 281–292. [Google Scholar] [CrossRef]
- Amimo, J.O.; Michael, H.; Chepngeno, J.; Raev, S.A.; Saif, L.J.; Vlasova, A.N. Immune Impairment Associated with Vitamin A Deficiency: Insights from Clinical Studies and Animal Model Research. Nutrients 2022, 14, 5038. [Google Scholar] [CrossRef]
- Huang, Z.; Liu, Y.; Qi, G.; Brand, D.; Zheng, S.G. Role of Vitamin A in the Immune System. J. Clin. Med. 2018, 7, 258. [Google Scholar] [CrossRef]
- Nagai, Y.; Ambinder, A.J. The Promise of Retinoids in the Treatment of Cancer: Neither Burnt Out Nor Fading Away. Cancers 2023, 15, 3535. [Google Scholar] [CrossRef] [PubMed]
- Bakac, E.R.; Percin, E.; Gunes-Bayir, A.; Dadak, A. A Narrative Review: The Effect and Importance of Carotenoids on Aging and Aging-Related Diseases. Int. J. Mol. Sci. 2023, 24, 15199. [Google Scholar] [CrossRef]
- O’Connor, C.; Varshosaz, P.; Moise, A.R. Mechanisms of Feedback Regulation of Vitamin A Metabolism. Nutrients 2022, 14, 1312. [Google Scholar] [CrossRef] [PubMed]
- Fukuoka, H.; Yokoi, N.; Sotozono, C. Immunohistochemistry in an Adult Case of Bitot’s Spots Caused by Vitamin A Deficiency. Diagnostics 2023, 13, 3676. [Google Scholar] [CrossRef] [PubMed]
- Zhao, Y.; Deng, S.; Li, C.; Cao, J.; Wu, A.; Chen, M.; Ma, X.; Wu, S.; Lian, Z. The Role of Retinoic Acid in Spermatogenesis and Its Application in Male Reproduction. Cells 2024, 13, 1092. [Google Scholar] [CrossRef]
- Filipe Rosa, L.; Petersen, P.P.; Görtz, L.F.; Stolzer, I.; Kaden-Volynets, V.; Günther, C.; Bischoff, S.C. Vitamin A- and D-Deficient Diets Disrupt Intestinal Antimicrobial Peptide Defense Involving Wnt and STAT5 Signaling Pathways in Mice. Nutrients 2023, 15, 376. [Google Scholar] [CrossRef] [PubMed]
- El Soury, M.; Fornasari, B.E.; Carta, G.; Zen, F.; Haastert-Talini, K.; Ronchi, G. The Role of Dietary Nutrients in Peripheral Nerve Regeneration. Int. J. Mol. Sci. 2021, 22, 7417. [Google Scholar] [CrossRef] [PubMed]
- Gutowska, K.; Formanowicz, D.; Formanowicz, P. Interrelations between Iron and Vitamin A—Studied Using Systems Approach. Int. J. Mol. Sci. 2022, 23, 1189. [Google Scholar] [CrossRef] [PubMed]
- Mugware, A.; Motadi, S.A.; Bere, A.; Mushaphi, L.F. Iron and Vitamin A Status of Children Aged 0 to 36 Months in Thulamela Municipality, Vhembe District, South Africa. Children 2024, 11, 1018. [Google Scholar] [CrossRef]
- Luca, M.M.; Buzatu, R.; Bumbu, B.A. Evaluating the Protective Role of Vitamin A Supplementation in Periodontal Health: A Comprehensive Systematic Review and Meta-Analysis. J. Clin. Med. 2024, 13, 4775. [Google Scholar] [CrossRef] [PubMed]
- Tavares Antunes, P.; das Graças Vaz-Tostes, M.; Tomáz Sant’Ana, C.; Araújo de Faria, R.; Lopes Toledo, R.C.; Brunoro Costa, N.M. Bioavailability of Iron and the Influence of Vitamin a in Biofortified Foods. Agronomy 2019, 9, 777. [Google Scholar] [CrossRef]
- Yee, M.M.F.; Chin, K.-Y.; Ima-Nirwana, S.; Wong, S.K. Vitamin A and Bone Health: A Review on Current Evidence. Molecules 2021, 26, 1757. [Google Scholar] [CrossRef] [PubMed]
- Harrison, E.H. Mechanisms Involved in the Intestinal Absorption of Dietary Vitamin A and Provitamin A Carotenoids. Biochim. Et Biophys. Acta (BBA)—Mol. Cell Biol. Lipids 2012, 1821, 70–77. [Google Scholar] [CrossRef]
- Harrison, E.H. Mechanisms of Transport and Delivery of Vitamin A and Carotenoids to the Retinal Pigment Epithelium. Mol. Nutr. Food Res. 2019, 63, 1801046. [Google Scholar] [CrossRef] [PubMed]
- Vieira, M.; Saraiva, M.J. Transthyretin: A Multifaceted Protein. BioMolecular Concepts 2014, 5, 45–54. [Google Scholar] [CrossRef] [PubMed]
- D’Ambrosio, D.N.; Clugston, R.D.; Blaner, W.S. Vitamin A Metabolism: An Update. Nutrients 2011, 3, 63–103. [Google Scholar] [CrossRef]
- Urbach, J.; Rando, R.R. Isomerization of All-Trans-Retinoic Acid to 9-Cis-Retinoic Acid. Biochem. J. 1994, 299, 459–465. [Google Scholar] [CrossRef]
- Martin Ask, N.; Leung, M.; Radhakrishnan, R.; Lobo, G.P. Vitamin A Transporters in Visual Function: A Mini Review on Membrane Receptors for Dietary Vitamin A Uptake, Storage, and Transport to the Eye. Nutrients 2021, 13, 3987. [Google Scholar] [CrossRef] [PubMed]
- Reboul, E. Absorption of Vitamin A and Carotenoids by the Enterocyte: Focus on Transport Proteins. Nutrients 2013, 5, 3563–3581. [Google Scholar] [CrossRef] [PubMed]
- Shete, V.; Quadro, L. Mammalian Metabolism of β-Carotene: Gaps in Knowledge. Nutrients 2013, 5, 4849–4868. [Google Scholar] [CrossRef]
- Samara, I.; Moula, A.I.; Moulas, A.N.; Katsouras, C.S. The Effect of Retinoids in Vascular Smooth Muscle Cells: From Phenotyping Switching to Proliferation and Migration. Int. J. Mol. Sci. 2024, 25, 10303. [Google Scholar] [CrossRef] [PubMed]
- Zhang, R.; Wang, Y.; Li, R.; Chen, G. Transcriptional Factors Mediating Retinoic Acid Signals in the Control of Energy Metabolism. Int. J. Mol. Sci. 2015, 16, 14210–14244. [Google Scholar] [CrossRef] [PubMed]
- Yu, L.; Wang, Y.; Yu, D.; Zhang, S.; Zheng, F.; Ding, N.; Zhu, L.; Zhu, Q.; Sun, W.; Li, S.; et al. Association between Serum Vitamin A, Blood Lipid Level and Dyslipidemia among Chinese Children and Adolescents. Nutrients 2022, 14, 1444. [Google Scholar] [CrossRef]
- Shin, K.-C.; Seo, M.-J.; Kim, Y.-S.; Yeom, S.-J. Molecular Properties of β-Carotene Oxygenases and Their Potential in Industrial Production of Vitamin A and Its Derivatives. Antioxidants 2022, 11, 1180. [Google Scholar] [CrossRef]
- Harrison, E.H. Carotenoids, β-Apocarotenoids, and Retinoids: The Long and the Short of It. Nutrients 2022, 14, 1411. [Google Scholar] [CrossRef]
- Szymański, Ł.; Skopek, R.; Palusińska, M.; Schenk, T.; Stengel, S.; Lewicki, S.; Kraj, L.; Kamiński, P.; Zelent, A. Retinoic Acid and Its Derivatives in Skin. Cells 2020, 9, 2660. [Google Scholar] [CrossRef] [PubMed]
- Everts, H.B.; Akuailou, E.-N. Retinoids in Cutaneous Squamous Cell Carcinoma. Nutrients 2021, 13, 153. [Google Scholar] [CrossRef]
- Zsikó, S.; Csányi, E.; Kovács, A.; Budai-Szűcs, M.; Gácsi, A.; Berkó, S. Methods to Evaluate Skin Penetration In Vitro. Sci. Pharm. 2019, 87, 19. [Google Scholar] [CrossRef]
- Hasan, M.; Khatun, A.; Kogure, K. Iontophoresis of Biological Macromolecular Drugs. Pharmaceutics 2022, 14, 525. [Google Scholar] [CrossRef] [PubMed]
- Conserva, M.R.; Anelli, L.; Zagaria, A.; Specchia, G.; Albano, F. The Pleiotropic Role of Retinoic Acid/Retinoic Acid Receptors Signaling: From Vitamin A Metabolism to Gene Rearrangements in Acute Promyelocytic Leukemia. Int. J. Mol. Sci. 2019, 20, 2921. [Google Scholar] [CrossRef] [PubMed]
- Zasada, M.; Budzisz, E. Retinoids: Active Molecules Influencing Skin Structure Formation in Cosmetic and Dermatological Treatments. Postep. Dermatol. Alergol. 2019, 36, 392–397. [Google Scholar] [CrossRef]
- Saubenova, M.; Rapoport, A.; Venkatachalam, M.; Dufossé, L.; Yermekbay, Z.; Oleinikova, Y. Production of Carotenoids by Microorganisms. Fermentation 2024, 10, 502. [Google Scholar] [CrossRef]
- Lešić, S.; Ivanišević, Z.; Špiljak, B.; Tomas, M.; Šoštarić, M.; Včev, A. The Impact of Vitamin Deficiencies on Oral Manifestations in Children. Dent. J. 2024, 12, 109. [Google Scholar] [CrossRef]
- Rathi, R.; Sanshita; Kumar, A.; Vishvakarma, V.; Huanbutta, K.; Singh, I.; Sangnim, T. Advancements in Rectal Drug Delivery Systems: Clinical Trials, and Patents Perspective. Pharmaceutics 2022, 14, 2210. [Google Scholar] [CrossRef]
- Cholidis, P.; Kranas, D.; Chira, A.; Galouni, E.A.; Adamantidi, T.; Anastasiadou, C.; Tsoupras, A. Shrimp Lipid Bioactives with Anti-Inflammatory, Antithrombotic, and Antioxidant Health-Promoting Properties for Cardio-Protection. Mar. Drugs 2024, 22, 554. [Google Scholar] [CrossRef]
- Baeza-Morales, A.; Medina-García, M.; Martínez-Peinado, P.; Pascual-García, S.; Pujalte-Satorre, C.; López-Jaén, A.B.; Martínez-Espinosa, R.M.; Sempere-Ortells, J.M. The Antitumour Mechanisms of Carotenoids: A Comprehensive Review. Antioxidants 2024, 13, 1060. [Google Scholar] [CrossRef]
- Quan, T. Human Skin Aging and the Anti-Aging Properties of Retinol. Biomolecules 2023, 13, 1614. [Google Scholar] [CrossRef]
- Andreazzoli, M.; Longoni, B.; Angeloni, D.; Demontis, G.C. Retinoid Synthesis Regulation by Retinal Cells in Health and Disease. Cells 2024, 13, 871. [Google Scholar] [CrossRef] [PubMed]
- Kaulmann, A.; Bohn, T. Carotenoids, Inflammation, and Oxidative Stress--Implications of Cellular Signaling Pathways and Relation to Chronic Disease Prevention. Nutr. Res. 2014, 34, 907–929. [Google Scholar] [CrossRef]
- Tsoupras, A.; Lordan, R.; Zabetakis, I. Inflammation, Not Cholesterol, Is a Cause of Chronic Disease. Nutrients 2018, 10, 604. [Google Scholar] [CrossRef] [PubMed]
- Lordan, R.; Redfern, S.; Tsoupras, A.; Zabetakis, I. Inflammation and Cardiovascular Disease: Are Marine Phospholipids the Answer? Food Funct. 2020, 11, 2861–2885. [Google Scholar] [CrossRef] [PubMed]
- Tsoupras, A.; Adamantidi, T.; Finos, M.A.; Philippopoulos, A.; Detopoulou, P.; Tsopoki, I.; Kynatidou, M.; Demopoulos, C.A. Re-Assessing the Role of Platelet Activating Factor and Its Inflammatory Signaling and Inhibitors in Cancer and Anti-Cancer Strategies. FBL 2024, 29, 345. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Y.; Liu, L.; Yue, L.; Huang, Y.; Wang, B.; Liu, P. Uncovering Key Mechanisms and Intervention Therapies in Aging Skin. Cytokine Growth Factor Rev. 2024, 79, 66–80. [Google Scholar] [CrossRef] [PubMed]
- Polcz, M.E.; Barbul, A. The Role of Vitamin A in Wound Healing. Nutr. Clin. Pract. 2019, 34, 695–700. [Google Scholar] [CrossRef]
- Ahmed, S.A.; Mendonca, P.; Messeha, S.S.; Oriaku, E.T.; Soliman, K.F.A. The Anticancer Effects of Marine Carotenoid Fucoxanthin through Phosphatidylinositol 3-Kinase (PI3K)-AKT Signaling on Triple-Negative Breast Cancer Cells. Molecules 2024, 29, 61. [Google Scholar] [CrossRef]
- Park, H.-A.; Hayden, M.M.; Bannerman, S.; Jansen, J.; Crowe-White, K.M. Anti-Apoptotic Effects of Carotenoids in Neurodegeneration. Molecules 2020, 25, 3453. [Google Scholar] [CrossRef]
- Tozer, S.; O’Mahony, C.; Hannah, J.; O’Brien, J.; Kelly, S.; Kosemund-Meynen, K.; Alexander-White, C. Aggregate Exposure Modelling of Vitamin A from Cosmetic Products, Diet and Food Supplements. Food Chem. Toxicol. 2019, 131, 110549. [Google Scholar] [CrossRef]
- Tsimidou, M.Z.; Mantzouridou, F.T.; Nenadis, N. Chapter Two—Minor Bioactive Lipids. In Advances in Food and Nutrition Research; Gallegos, C., Ruiz-Méndez, M.-V., Eds.; Dietary Lipids: Nutritional and Technological Aspects; Academic Press: Cambridge, MA, USA, 2023; Volume 105, pp. 51–95. [Google Scholar]
- Islam, F.; Khan, J.; Zehravi, M.; Das, R.; Haque, M.A.; Banu, A.; Parwaiz, S.; Nainu, F.; Nafady, M.H.; Shahriar, S.M.S.; et al. Synergistic Effects of Carotenoids: Therapeutic Benefits on Human Health. Process Biochem. 2024, 136, 254–272. [Google Scholar] [CrossRef]
- Bufka, J.; Vaňková, L.; Sýkora, J.; Křížková, V. Exploring Carotenoids: Metabolism, Antioxidants, and Impacts on Human Health. J. Funct. Foods 2024, 118, 106284. [Google Scholar] [CrossRef]
- Puri, V.; Nagpal, M.; Singh, I.; Singh, M.; Dhingra, G.A.; Huanbutta, K.; Dheer, D.; Sharma, A.; Sangnim, T. A Comprehensive Review on Nutraceuticals: Therapy Support and Formulation Challenges. Nutrients 2022, 14, 4637. [Google Scholar] [CrossRef] [PubMed]
- Morganti, P.; Morganti, G.; Gagliardini, A.; Lohani, A. From Cosmetics to Innovative Cosmeceuticals—Non-Woven Tissues as New Biodegradable Carriers. Cosmetics 2021, 8, 65. [Google Scholar] [CrossRef]
- Hoang, H.T.; Moon, J.-Y.; Lee, Y.-C. Natural Antioxidants from Plant Extracts in Skincare Cosmetics: Recent Applications, Challenges and Perspectives. Cosmetics 2021, 8, 106. [Google Scholar] [CrossRef]
- Pérez-Sánchez, A.; Barrajón-Catalán, E.; Herranz-López, M.; Micol, V. Nutraceuticals for Skin Care: A Comprehensive Review of Human Clinical Studies. Nutrients 2018, 10, 403. [Google Scholar] [CrossRef]
- Ahmed, I.A.; Mikail, M.A. Diet and Skin Health: The Good and the Bad. Nutrition 2024, 119, 112350. [Google Scholar] [CrossRef]
- Balić, A.; Mokos, M. Do We Utilize Our Knowledge of the Skin Protective Effects of Carotenoids Enough? Antioxidants 2019, 8, 259. [Google Scholar] [CrossRef] [PubMed]
- Galasso, C.; Corinaldesi, C.; Sansone, C. Carotenoids from Marine Organisms: Biological Functions and Industrial Applications. Antioxidants 2017, 6, 96. [Google Scholar] [CrossRef]
- Peng, J.; Yuan, J.-P.; Wu, C.-F.; Wang, J.-H. Fucoxanthin, a Marine Carotenoid Present in Brown Seaweeds and Diatoms: Metabolism and Bioactivities Relevant to Human Health. Mar. Drugs 2011, 9, 1806–1828. [Google Scholar] [CrossRef]
- Kontaxi, N.-I.; Panoutsopoulou, E.; Ofrydopolou, A.; Tsoupras, A. Anti-Inflammatory Benefits of Grape Pomace and Tomato Bioactives as Ingredients in Sun Oils against UV Radiation for Skin Protection. Appl. Sci. 2024, 14, 6236. [Google Scholar] [CrossRef]
- Michalak, M. Plant-Derived Antioxidants: Significance in Skin Health and the Ageing Process. Int. J. Mol. Sci. 2022, 23, 585. [Google Scholar] [CrossRef]
- Darvin, M.E.; Fluhr, J.W.; Meinke, M.C.; Zastrow, L.; Sterry, W.; Lademann, J. Topical Beta-Carotene Protects against Infra-Red-Light-Induced Free Radicals. Exp. Dermatol. 2011, 20, 125–129. [Google Scholar] [CrossRef] [PubMed]
- Carotenoids: New Applications of “Old” Pigments. Phyton-Int. J. Exp. Bot. 2021, 90, 1041–1062. [CrossRef]
- Baran, M.T.; Miziak, P.; Bonio, K. Characteristics of Carotenoids and Their Use in the Cosmetics Industry. J. Educ. Health Sport 2020, 10, 192–196. [Google Scholar] [CrossRef]
- Meinke, M.C.; Friedrich, A.; Tscherch, K.; Haag, S.F.; Darvin, M.E.; Vollert, H.; Groth, N.; Lademann, J.; Rohn, S. Influence of Dietary Carotenoids on Radical Scavenging Capacity of the Skin and Skin Lipids. Eur. J. Pharm. Biopharm. 2013, 84, 365–373. [Google Scholar] [CrossRef] [PubMed]
- Mayne, S.T.; Cartmel, B.; Scarmo, S.; Jahns, L.; Ermakov, I.V.; Gellermann, W. Resonance Raman Spectroscopic Evaluation of Skin Carotenoids as a Biomarker of Carotenoid Status for Human Studies. Arch. Biochem. Biophys. 2013, 539, 163–170. [Google Scholar] [CrossRef]
- Grether-Beck, S.; Marini, A.; Jaenicke, T.; Stahl, W.; Krutmann, J. Molecular Evidence That Oral Supplementation with Lycopene or Lutein Protects Human Skin against Ultraviolet Radiation: Results from a Double-blinded, Placebo-controlled, Crossover Study. Br. J. Dermatol. 2017, 176, 1231–1240. [Google Scholar] [CrossRef]
- Nishino, A.; Sugimoto, K.; Sambe, H.; Ichihara, T.; Takaha, T.; Kuriki, T. Effects of Dietary Paprika Xanthophylls on Ultraviolet Light-Induced Skin Damage: A Double-Blind Placebo-Controlled Study. J. Oleo Sci. 2018, 67, 863–869. [Google Scholar] [CrossRef]
- De Cássia Dias Mendes-Silva, T.; Fontenele da Silva Andrade, R.; Ootani, M.A.; Mendes, P.V.D.; de Queiroz Cavalcanti de Sá, R.A.; da Silva, M.R.F.; Souza, K.S.; dos Santos Correia, M.T.; de Oliveira, M.B.M. Biotechnological Potential of Carotenoids Produced by Extremophilic Microorganisms and Application Prospects for the Cosmetics Industry. Adv. Microbiol. 2020, 10, 397–410. [Google Scholar] [CrossRef]
- Motamedi, M.; Chehade, A.; Sanghera, R.; Grewal, P. A Clinician’s Guide to Topical Retinoids. J. Cutan. Med. Surg. 2022, 26, 71–78. [Google Scholar] [CrossRef] [PubMed]
- Leyden, J.J.; Tanghetti, E.A.; Miller, B.; Ung, M.; Berson, D.; Lee, J. Once-Daily Tazarotene 0.1 % Gel versus Once-Daily Tretinoin 0.1 % Microsponge Gel for the Treatment of Facial Acne Vulgaris: A Double-Blind Randomized Trial. Cutis 2002, 69, 12–19. [Google Scholar]
- Bagatin, E.; de Sá Gonçalves, H.; Sato, M.; Almeida, L.M.C.; Miot, H.A. Comparable Efficacy of Adapalene 0.3% Gel and Tretinoin 0.05% Cream as Treatment for Cutaneous Photoaging. Eur. J. Dermatol. 2018, 28, 343–350. [Google Scholar] [CrossRef] [PubMed]
- Agnihotri, S.; Kaur, J.; Masand, P.; Anurag; Parihar, V.K.; Sharma, A. Vitamins Strategies for Psoriasis: An Update on Current Scientific Evidence. J. Holist. Integr. Pharm. 2023, 4, 299–309. [Google Scholar] [CrossRef]
- Davinelli, S.; Nielsen, M.E.; Scapagnini, G. Astaxanthin in Skin Health, Repair, and Disease: A Comprehensive Review. Nutrients 2018, 10, 522. [Google Scholar] [CrossRef] [PubMed]
- Sauer, L.; Li, B.; Bernstein, P.S. Ocular Carotenoid Status in Health and Disease. Annu. Rev. Nutr. 2019, 39, 95–120. [Google Scholar] [CrossRef] [PubMed]
- Rotter, A.; Varamogianni-Mamatsi, D.; Zvonar Pobirk, A.; Gosenca Matjaž, M.; Cueto, M.; Díaz-Marrero, A.R.; Jónsdóttir, R.; Sveinsdóttir, K.; Catalá, T.S.; Romano, G.; et al. Marine Cosmetics and the Blue Bioeconomy: From Sourcing to Success Stories. iScience 2024, 27, 111339. [Google Scholar] [CrossRef]
- Papikinou, M.-A.; Pavlidis, K.; Cholidis, P.; Kranas, D.; Adamantidi, T.; Anastasiadou, C.; Tsoupras, A. Marine Fungi Bioactives with Anti-Inflammatory, Antithrombotic and Antioxidant Health-Promoting Properties Against Inflammation-Related Chronic Diseases. Mar. Drugs 2024, 22, 520. [Google Scholar] [CrossRef] [PubMed]
- Meléndez-Martínez, A.J.; Mandić, A.I.; Bantis, F.; Böhm, V.; Borge, G.I.A.; Brnčić, M.; Bysted, A.; Cano, M.P.; Dias, M.G.; Elgersma, A.; et al. A Comprehensive Review on Carotenoids in Foods and Feeds: Status Quo, Applications, Patents, and Research Needs. Crit. Rev. Food Sci. Nutr. 2022, 62, 1999–2049. [Google Scholar] [CrossRef]
- Chungchunlam, S.M.S.; Moughan, P.J. Comparative Bioavailability of Vitamins in Human Foods Sourced from Animals and Plants. Crit. Rev. Food Sci. Nutr. 2024, 64, 11590–11625. [Google Scholar] [CrossRef]
- Maoka, T. Carotenoids as Natural Functional Pigments. J. Nat. Med. 2020, 74, 1–16. [Google Scholar] [CrossRef]
- Zerres, S.; Stahl, W. Carotenoids in Human Skin. Biochim. Biophys. Acta Mol. Cell Biol. Lipids 2020, 1865, 158588. [Google Scholar] [CrossRef]
- Naisi, S.; Bayat, M.; Zahraei Salehi, T.; Rahimian Zarif, B.; Yahyaraeyat, R. Antimicrobial and Anti-Biofilm Effects of Carotenoid Pigment Extracted from Rhodotorula Glutinis Strain on Food-Borne Bacteria. Iran J. Microbiol. 2023, 15, 79–88. [Google Scholar] [CrossRef]
- Larsson, S.C.; Bergkvist, L.; Näslund, I.; Rutegård, J.; Wolk, A. Vitamin A, Retinol, and Carotenoids and the Risk of Gastric Cancer: A Prospective Cohort Study2. Am. J. Clin. Nutr. 2007, 85, 497–503. [Google Scholar] [CrossRef]
- Adeyemi, K.D.; Sabow, A.B.; Ebrahimi, M.; Samsudin, A.A.; Sazili, A.Q. Fatty Acid Composition, Cholesterol and Antioxidant Status of Infraspinatus Muscle, Liver and Kidney of Goats Fed Blend of Palm Oil and Canola Oil. Ital. J. Anim. Sci. 2016, 15, 181–190. [Google Scholar] [CrossRef]
- Erkiert-Polguj, A.; Kazimierska, K.; Kalinowska-Lis, U. Assessment of the Impact of a Cosmetic Product with Sheep Colostrum on Acne Skin. Appl. Sci. 2024, 14, 2199. [Google Scholar] [CrossRef]
- Flegler, A.; Runzheimer, K.; Kombeitz, V.; Mänz, A.T.; Heidler von Heilborn, D.; Etzbach, L.; Schieber, A.; Hölzl, G.; Hüttel, B.; Woehle, C.; et al. Arthrobacter bussei sp. nov., a Pink-Coloured Organism Isolated from Cheese Made of Cow’s Milk. Int. J. Syst. Evol. Microbiol. 2020, 70, 3027–3036. [Google Scholar] [CrossRef] [PubMed]
- Ballesteros, M.N.; Valenzuela, F.; Robles, A.E.; Artalejo, E.; Aguilar, D.; Andersen, C.J.; Valdez, H.; Fernandez, M.L. One Egg per Day Improves Inflammation When Compared to an Oatmeal-Based Breakfast without Increasing Other Cardiometabolic Risk Factors in Diabetic Patients. Nutrients 2015, 7, 3449–3463. [Google Scholar] [CrossRef]
- Ballesteros, M.N.; Cabrera, R.M.; Saucedo, S.; Fernandez, M.L. Two Eggs per Day Increase Plasma Lutein and Zeaxanthin in a Pediatric Population Characterized by Low Intake of Fruits and Vegetables. J. Adv. Med. Med. Res. 2013, 3, 2203–2213. [Google Scholar] [CrossRef]
- Kelly, E.R.; Plat, J.; Haenen, G.R.M.M.; Kijlstra, A.; Berendschot, T.T.J.M. The Effect of Modified Eggs and an Egg-Yolk Based Beverage on Serum Lutein and Zeaxanthin Concentrations and Macular Pigment Optical Density: Results from a Randomized Trial. PLoS ONE 2014, 9, e92659. [Google Scholar] [CrossRef] [PubMed]
- Rodrigo-Baños, M.; Garbayo, I.; Vílchez, C.; Bonete, M.J.; Martínez-Espinosa, R.M. Carotenoids from Haloarchaea and Their Potential in Biotechnology. Mar. Drugs 2015, 13, 5508–5532. [Google Scholar] [CrossRef] [PubMed]
- Gebregziabher, B.S.; Gebremeskel, H.; Debesa, B.; Ayalneh, D.; Mitiku, T.; Wendwessen, T.; Habtemariam, E.; Nur, S.; Getachew, T. Carotenoids: Dietary Sources, Health Functions, Biofortification, Marketing Trend and Affecting Factors—A Review. J. Agric. Food Res. 2023, 14, 100834. [Google Scholar] [CrossRef]
- Miao, Q.; Si, X.; Zhao, Q.; Zhang, H.; Qin, Y.; Tang, C.; Zhang, J. Deposition and Enrichment of Carotenoids in Livestock Products: An Overview. Food Chem. X 2024, 21, 101245. [Google Scholar] [CrossRef] [PubMed]
- Chiu, H.-F.; Liao, J.-Y.; Lu, Y.-Y.; Han, Y.-C.; Shen, Y.-C.; Venkatakrishnan, K.; Golovinskaia, O.; Wang, C.-K. Anti-Proliferative, Anti-Inflammatory and pro-Apoptotic Effects of Dunaliella Salina on Human KB Oral Carcinoma Cells. J. Food Biochem. 2017, 41, e12349. [Google Scholar] [CrossRef]
- Teixé-Roig, J.; Oms-Oliu, G.; Odriozola-Serrano, I.; Martín-Belloso, O. Enhancing in Vivo Retinol Bioavailability by Incorporating β-Carotene from Alga Dunaliella salina into Nanoemulsions Containing Natural-Based Emulsifiers. Food Res. Int. 2023, 164, 112359. [Google Scholar] [CrossRef]
- Havas, F.; Krispin, S.; Cohen, M.; Loing, E.; Farge, M.; Suere, T.; Attia-Vigneau, J. A Dunaliella Salina Extract Counteracts Skin Aging under Intense Solar Irradiation Thanks to Its Antiglycation and Anti-Inflammatory Properties. Mar. Drugs 2022, 20, 104. [Google Scholar] [CrossRef] [PubMed]
- Yang, D.-J.; Lin, J.-T.; Chen, Y.-C.; Liu, S.-C.; Lu, F.-J.; Chang, T.-J.; Wang, M.; Lin, H.-W.; Chang, Y.-Y. Suppressive Effect of Carotenoid Extract of Dunaliella salina Alga on Production of LPS-Stimulated pro-Inflammatory Mediators in RAW264.7 Cells via NF-κB and JNK Inactivation. J. Funct. Foods 2013, 5, 607–615. [Google Scholar] [CrossRef]
- Ito, N.; Seki, S.; Ueda, F. The Protective Role of Astaxanthin for UV-Induced Skin Deterioration in Healthy People—A Randomized, Double-Blind, Placebo-Controlled Trial. Nutrients 2018, 10, 817. [Google Scholar] [CrossRef] [PubMed]
- Rao, A.R.; Sindhuja, H.N.; Dharmesh, S.M.; Sankar, K.U.; Sarada, R.; Ravishankar, G.A. Effective Inhibition of Skin Cancer, Tyrosinase, and Antioxidative Properties by Astaxanthin and Astaxanthin Esters from the Green Alga Haematococcus Pluvialis. J. Agric. Food Chem. 2013, 61, 3842–3851. [Google Scholar] [CrossRef] [PubMed]
- Yoon, H.-S.; Cho, H.H.; Cho, S.; Lee, S.-R.; Shin, M.-H.; Chung, J.H. Supplementing with Dietary Astaxanthin Combined with Collagen Hydrolysate Improves Facial Elasticity and Decreases Matrix Metalloproteinase-1 and -12 Expression: A Comparative Study with Placebo. J. Med. Food 2014, 17, 810–816. [Google Scholar] [CrossRef] [PubMed]
- Hao, Z. Astaxanthin Improves Serum Cytokine Expression and Semen Quality of Diabetes Mellitus KKAy Mice. Chem. Biol. Interact. 2020, 332, 109303. [Google Scholar] [CrossRef] [PubMed]
- Nazarudin, M.F.; Yasin, I.S.M.; Mazli, N.A.I.N.; Saadi, A.R.; Azizee, M.H.S.; Nooraini, M.A.; Saad, N.; Ferdous, U.T.; Fakhrulddin, I.M. Preliminary Screening of Antioxidant and Cytotoxic Potential of Green Seaweed, Halimeda opuntia (Linnaeus) Lamouroux. Saudi J. Biol. Sci. 2022, 29, 2698–2705. [Google Scholar] [CrossRef] [PubMed]
- Shih, P.-H.; Shiue, S.-J.; Chen, C.-N.; Cheng, S.-W.; Lin, H.-Y.; Wu, L.-W.; Wu, M.-S. Fucoidan and Fucoxanthin Attenuate Hepatic Steatosis and Inflammation of NAFLD through Modulation of Leptin/Adiponectin Axis. Mar. Drugs 2021, 19, 148. [Google Scholar] [CrossRef]
- Sun, G.; Xin, T.; Zhang, R.; Liu, C.; Pang, Q. Fucoxanthin Attenuates Behavior Deficits and Neuroinflammatory Response in 1-Methyl-4-Phenyl-1,2,3,6 -Tetrahydropyridine-Induced Parkinson’s Disease in Mice. Pharmacogn. Mag. 2020, 16, 51–56. [Google Scholar] [CrossRef]
- Hitoe, S.; Shimoda, H. Seaweed Fucoxanthin Supplementation Improves Obesity Parameters in Mild Obese Japanese Subjects. Funct. Foods Health Dis. 2017, 7, 246–262. [Google Scholar] [CrossRef]
- Wang, P.-T.; Sudirman, S.; Hsieh, M.-C.; Hu, J.-Y.; Kong, Z.-L. Oral Supplementation of Fucoxanthin-Rich Brown Algae Extract Ameliorates Cisplatin-Induced Testicular Damage in Hamsters. Biomed. Pharmacother. 2020, 125, 109992. [Google Scholar] [CrossRef]
- Wang, J.-Y.; Lee, Y.-J.; Chou, M.-C.; Chang, R.; Chiu, C.-H.; Liang, Y.-J.; Wu, L.-S. Astaxanthin Protects Steroidogenesis from Hydrogen Peroxide-Induced Oxidative Stress in Mouse Leydig Cells. Mar. Drugs 2015, 13, 1375–1388. [Google Scholar] [CrossRef]
- Serra, A.T.; Silva, S.D.; Pleno de Gouveia, L.; Alexandre, A.M.R.C.; Pereira, C.V.; Pereira, A.B.; Partidário, A.C.; Silva, N.E.; Bohn, T.; Gonçalves, V.S.S.; et al. A Single Dose of Marine Chlorella Vulgaris Increases Plasma Concentrations of Lutein, β-Carotene and Zeaxanthin in Healthy Male Volunteers. Antioxidants 2021, 10, 1164. [Google Scholar] [CrossRef]
- Lopes, G.; Clarinha, D.; Vasconcelos, V. Carotenoids from Cyanobacteria: A Biotechnological Approach for the Topical Treatment of Psoriasis. Microorganisms 2020, 8, 302. [Google Scholar] [CrossRef]
- Pagels, F.; Salvaterra, D.; Amaro, H.M.; Lopes, G.; Sousa-Pinto, I.; Vasconcelos, V.; Guedes, A.C. Bioactive Potential of Cyanobium sp. Pigment-Rich Extracts. J. Appl. Phycol. 2020, 32, 3031–3040. [Google Scholar] [CrossRef]
- Morone, J.; Lopes, G.; Preto, M.; Vasconcelos, V.; Martins, R. Exploitation of Filamentous and Picoplanktonic Cyanobacteria for Cosmetic Applications: Potential to Improve Skin Structure and Preserve Dermal Matrix Components. Mar. Drugs 2020, 18, 486. [Google Scholar] [CrossRef] [PubMed]
- Squillaci, G.; Parrella, R.; Carbone, V.; Minasi, P.; La Cara, F.; Morana, A. Carotenoids from the Extreme Halophilic Archaeon Haloterrigena Turkmenica: Identification and Antioxidant Activity. Extremophiles 2017, 21, 933–945. [Google Scholar] [CrossRef]
- Caimi, A.T.; Yasynska, O.; Rivas Rojas, P.C.; Romero, E.L.; Morilla, M.J. Improved Stability and Biological Activity of Bacterioruberin in Nanovesicles. J. Drug Deliv. Sci. Technol. 2022, 77, 103896. [Google Scholar] [CrossRef]
- Sahli, K.; Gomri, M.A.; Esclapez, J.; Gómez-Villegas, P.; Ghennai, O.; Bonete, M.-J.; León, R.; Kharroub, K. Bioprospecting and Characterization of Pigmented Halophilic Archaeal Strains from Algerian Hypersaline Environments with Analysis of Carotenoids Produced by Halorubrum sp. BS2. J. Basic Microbiol. 2020, 60, 624–638. [Google Scholar] [CrossRef]
- Safarpour, A.; Ebrahimi, M.; Shahzadeh Fazeli, S.A.; Amoozegar, M.A. Supernatant Metabolites from Halophilic Archaea to Reduce Tumorigenesis in Prostate Cancer In-Vitro and In-Vivo. Iran J. Pharm. Res. 2019, 18, 241–253. [Google Scholar]
- Von Oppen-Bezalel, L.; Fishbein, D.; Havas, F.; Ben-Chitrit, O.; Khaiat, A. The Photoprotective Effects of a Food Supplement Tomato Powder Rich in Phytoene and Phytofluene, the Colorless Carotenoids, a Preliminary Study. Glob. Dermatol. 2015, 2, 4. [Google Scholar] [CrossRef][Green Version]
- Okonogi, S.; Riangjanapatee, P. Physicochemical Characterization of Lycopene-Loaded Nanostructured Lipid Carrier Formulations for Topical Administration. Int. J. Pharm. 2015, 478, 726–735. [Google Scholar] [CrossRef]
- Groten, K.; Marini, A.; Grether-Beck, S.; Jaenicke, T.; Ibbotson, S.H.; Moseley, H.; Ferguson, J.; Krutmann, J. Tomato Phytonutrients Balance UV Response: Results from a Double-Blind, Randomized, Placebo-Controlled Study. Skin Pharmacol. Physiol. 2019, 32, 101–108. [Google Scholar] [CrossRef]
- Sokoloski, L.; Borges, M.; Bagatin, E. Lycopene Not in Pill, nor in Natura Has Photoprotective Systemic Effect. Arch. Dermatol. Res. 2015, 307, 545–549. [Google Scholar] [CrossRef] [PubMed]
- Armoza, A.; Haim, Y.; Bashiri, A.; Wolak, T.; Paran, E. Tomato Extract and the Carotenoids Lycopene and Lutein Improve Endothelial Function and Attenuate Inflammatory NF-κB Signaling in Endothelial Cells. J. Hypertens. 2013, 31, 521–529, discussion 529. [Google Scholar] [CrossRef]
- Stajčić, S.; Ćetković, G.; Čanadanović-Brunet, J.; Djilas, S.; Mandić, A.; Četojević-Simin, D. Tomato Waste: Carotenoids Content, Antioxidant and Cell Growth Activities. Food Chem. 2015, 172, 225–232. [Google Scholar] [CrossRef]
- Havas, F.; Krispin, S.; Meléndez-Martínez, A.J.; von Oppen-Bezalel, L. Preliminary Data on the Safety of Phytoene- and Phytofluene-Rich Products for Human Use Including Topical Application. J. Toxicol. 2018, 2018, 5475784. [Google Scholar] [CrossRef]
- Vasconcelos, A.G.; Amorim, A.d.G.; dos Santos, R.C.; Souza, J.M.T.; de Souza, L.K.M.; Araújo, T.d.S.; Nicolau, L.A.D.; Carvalho, L.d.L.; de Aquino, P.E.A.; Martins, C.d.S.; et al. Lycopene Rich Extract from Red Guava (Psidium guajava L.) Displays Anti-Inflammatory and Antioxidant Profile by Reducing Suggestive Hallmarks of Acute Inflammatory Response in Mice. Food Res. Int. 2017, 99, 959–968. [Google Scholar] [CrossRef]
- Pons, E.; Alquézar, B.; Rodríguez, A.; Martorell, P.; Genovés, S.; Ramón, D.; Rodrigo, M.J.; Zacarías, L.; Peña, L. Metabolic Engineering of β-Carotene in Orange Fruit Increases Its In Vivo Antioxidant Properties. Plant Biotechnol. J. 2014, 12, 17–27. [Google Scholar] [CrossRef]
- Petito, N.d.L.; Devens, J.M.; Falcão, D.Q.; Dantas, F.M.L.; Passos, T.S.; Araujo, K.G.d.L. Nanoencapsulation of Red Bell Pepper Carotenoids: Comparison of Encapsulating Agents in an Emulsion Based System. Colorants 2022, 1, 132–148. [Google Scholar] [CrossRef]
- Marra, A.; Manousakis, V.; Zervas, G.P.; Koutis, N.; Finos, M.A.; Adamantidi, T.; Panoutsopoulou, E.; Ofrydopoulou, A.; Tsoupras, A. Avocado and Its By-Products as Natural Sources of Valuable Anti-Inflammatory and Antioxidant Bioactives for Functional Foods and Cosmetics with Health-Promoting Properties. Appl. Sci. 2024, 14, 5978. [Google Scholar] [CrossRef]
- Edwards, C.G.; Walk, A.M.; Thompson, S.V.; Reeser, G.E.; Erdman, J.W.; Burd, N.A.; Holscher, H.D.; Khan, N.A. Effects of 12-Week Avocado Consumption on Cognitive Function among Adults with Overweight and Obesity. Int. J. Psychophysiol. 2020, 148, 13–24. [Google Scholar] [CrossRef]
- Villa-Rodriguez, J.A.; Yahia, E.M.; González-León, A.; Ifie, I.; Robles-Zepeda, R.E.; Domínguez-Avila, J.A.; González-Aguilar, G.A. Ripening of ‘Hass’ Avocado Mesocarp Alters Its Phytochemical Profile and the in Vitro Cytotoxic Activity of Its Methanolic Extracts. South Afr. J. Bot. 2020, 128, 1–8. [Google Scholar] [CrossRef]
- Wang, L.; Tao, L.; Hao, L.; Stanley, T.H.; Huang, K.-H.; Lambert, J.D.; Kris-Etherton, P.M. A Moderate-Fat Diet with One Avocado per Day Increases Plasma Antioxidants and Decreases the Oxidation of Small, Dense LDL in Adults with Overweight and Obesity: A Randomized Controlled Trial. J. Nutr. 2020, 150, 276–284. [Google Scholar] [CrossRef]
- Hsu, H.-Y.; Chen, B.-H. A Comparative Study on Inhibition of Breast Cancer Cells and Tumors in Mice by Carotenoid Extract and Nanoemulsion Prepared from Sweet Potato (Ipomoea batatas L.) Peel. Pharmaceutics 2022, 14, 980. [Google Scholar] [CrossRef]
- Kavalappa, Y.P.; Gopal, S.S.; Ponesakki, G. Lutein Inhibits Breast Cancer Cell Growth by Suppressing Antioxidant and Cell Survival Signals and Induces Apoptosis. J. Cell. Physiol. 2021, 236, 1798–1809. [Google Scholar] [CrossRef] [PubMed]
- Borja-Martínez, M.; Lozano-Sánchez, J.; Borrás-Linares, I.; Pedreño, M.A.; Sabater-Jara, A.B. Revalorization of Broccoli By-Products for Cosmetic Uses Using Supercritical Fluid Extraction. Antioxidants 2020, 9, 1195. [Google Scholar] [CrossRef]
- Freitas, A.; Moldão-Martins, M.; Costa, H.S.; Albuquerque, T.G.; Valente, A.; Sanches-Silva, A. Effect of UV-C Radiation on Bioactive Compounds of Pineapple (Ananas comosus L. Merr.) by-Products. J. Sci. Food Agric. 2015, 95, 44–52. [Google Scholar] [CrossRef] [PubMed]
- Górnaś, P.; Rudzińska, M.; Raczyk, M.; Mišina, I.; Soliven, A.; Segliņa, D. Composition of Bioactive Compounds in Kernel Oils Recovered from Sour Cherry (Prunus cerasus L.) by-Products: Impact of the Cultivar on Potential Applications. Ind. Crops Prod. 2016, 82, 44–50. [Google Scholar] [CrossRef]
- Fam, V.W.; Holt, R.R.; Keen, C.L.; Sivamani, R.K.; Hackman, R.M. Prospective Evaluation of Mango Fruit Intake on Facial Wrinkles and Erythema in Postmenopausal Women: A Randomized Clinical Pilot Study. Nutrients 2020, 12, 3381. [Google Scholar] [CrossRef]
- Diprat, A.B.; Silveira Thys, R.C.; Rodrigues, E.; Rech, R. Chlorella sorokiniana: A New Alternative Source of Carotenoids and Proteins for Gluten-Free Bread. LWT 2020, 134, 109974. [Google Scholar] [CrossRef]
- Mudroňová, D.; Karaffová, V.; Koščová, J.; Bartkovský, M.; Marcinčáková, D.; Popelka, P.; Klempová, T.; Čertík, M.; Mačanga, J.; Marcinčák, S. Effect of Fungal Gamma-Linolenic Acid and Beta-Carotene Containing Prefermented Feed on Immunity and Gut of Broiler Chicken. Poult. Sci. 2018, 97, 4211–4218. [Google Scholar] [CrossRef]
- Jimenez-Escobar, M.P.; Pascual-Mathey, L.I.; Beristain, C.I.; Flores-Andrade, E.; Jiménez, M.; Pascual-Pineda, L.A. In Vitro and In Vivo Antioxidant Properties of Paprika Carotenoids Nanoemulsions. LWT 2020, 118, 108694. [Google Scholar] [CrossRef]
- Morse, N.L.; Reid, A.-J.; St-Onge, M. An Open-Label Clinical Trial Assessing the Efficacy and Safety of Bend Skincare Anti-Aging Formula on Minimal Erythema Dose in Skin. Photodermatol. Photoimmunol. Photomed. 2018, 34, 152–161. [Google Scholar] [CrossRef] [PubMed]
- Divya, P.; Puthusseri, B.; Neelwarne, B. Carotenoid Content, Its Stability during Drying and the Antioxidant Activity of Commercial Coriander (Coriandrum sativum L.) Varieties. Food Res. Int. 2012, 45, 342–350. [Google Scholar] [CrossRef]
- Azizi, M.; Zare, D.; Akhavan Sepahy, A.; Azin, M. Evaluating the Effect of Microbial Stimulation and Oxidative Stress on Increasing β-Carotene Production in Blakeslea Trispora. Microbiol. Metab. Biotechnol. 2021, 4, 1–11. [Google Scholar] [CrossRef]
- Naz, T.; Nosheen, S.; Li, S.; Nazir, Y.; Mustafa, K.; Liu, Q.; Garre, V.; Song, Y. Comparative Analysis of β-Carotene Production by Mucor Circinelloides Strains CBS 277.49 and WJ11 under Light and Dark Conditions. Metabolites 2020, 10, 38. [Google Scholar] [CrossRef] [PubMed]
- Ochoa-Viñals, N.; Alonso-Estrada, D.; Pacios-Michelena, S.; García-Cruz, A.; Ramos-González, R.; Faife-Pérez, E.; Michelena-Álvarez, L.G.; Martínez-Hernández, J.L.; Iliná, A. Current Advances in Carotenoid Production by Rhodotorula Sp. Fermentation 2024, 10, 190. [Google Scholar] [CrossRef]
- Li, J.; Bates, E.; Perera, D.S.; Palage, A.M.; Ward, V.C.A. Membrane Engineering for Carotenoid Production in Escherichia coli. SynBio 2024, 2, 349–362. [Google Scholar] [CrossRef]
- Promdonkoy, P.; Watcharawipas, A.; Bubphasawan, S.; Sansatchanon, K.; Suwanakitti, N.; Kocharin, K.; Runguphan, W. Metabolic Engineering of Saccharomyces Cerevisiae for Production of Canthaxanthin, Zeaxanthin, and Astaxanthin. J. Fungi 2024, 10, 433. [Google Scholar] [CrossRef] [PubMed]
- Luo, W.; Xue, C.; Zhao, Y.; Zhang, H.; Rao, Z.; Yu, X. Blakeslea Trispora Photoreceptors: Identification and Functional Analysis. Appl. Environ. Microbiol. 2020, 86, e02962-19. [Google Scholar] [CrossRef] [PubMed]
- Mantzouridou, F.T.; Sferopoulou, E.; Thanou, P. Uncovering the Hidden Potential of Phytoene Production by the Fungus Blakeslea Trispora. Foods 2024, 13, 2882. [Google Scholar] [CrossRef] [PubMed]
- Naz, T.; Nazir, Y.; Nosheen, S.; Ullah, S.; Halim, H.; Fazili, A.B.A.; Li, S.; Mustafa, K.; Mohamed, H.; Yang, W.; et al. Redirecting Metabolic Flux towards the Mevalonate Pathway for Enhanced β-Carotene Production in M. Circinelloides CBS 277.49. BioMed Res. Int. 2020, 2020, 8890269. [Google Scholar] [CrossRef]
- Allahkarami, S.; Sepahi, A.A.; Hosseini, H.; Razavi, M.R. Isolation and Identification of Carotenoid-Producing Rhodotorula sp. from Pinaceae Forest Ecosystems and Optimization of in Vitro Carotenoid Production. Biotechnol. Rep. (Amst) 2021, 32, e00687. [Google Scholar] [CrossRef]
- Li, Z.; Li, C.; Cheng, P.; Yu, G. Rhodotorula mucilaginosa—Alternative Sources of Natural Carotenoids, Lipids, and Enzymes for Industrial Use. Heliyon 2022, 8, e11505. [Google Scholar] [CrossRef]
- Hernández-Almanza, A.; Navarro Macias V., E.; Aguilar, O.; Aguilar, M.; Aguilar, N.C. Carotenoids Extraction from Rhodotorula Glutinis Cells Using Various Techniques: A Comparative Study. Indian J. Exp. Biol. 2024, 55, 479–484. [Google Scholar]
- Liu, C.; Cui, Y.; Pi, F.; Guo, Y.; Cheng, Y.; Qian, H. Torularhodin Ameliorates Oxidative Activity in Vitro and D-Galactose-Induced Liver Injury via the Nrf2/HO-1 Signaling Pathway in Vivo. J. Agric. Food Chem. 2019, 67, 10059–10068. [Google Scholar] [CrossRef] [PubMed]
- Du, C.; Guo, Y.; Cheng, Y.; Han, M.; Zhang, W.; Qian, H. Torulene and Torularhodin, Protects Human Prostate Stromal Cells from Hydrogen Peroxide-Induced Oxidative Stress Damage through the Regulation of Bcl-2/Bax Mediated Apoptosis. Free Radic. Res. 2017, 51, 113–123. [Google Scholar] [CrossRef]
- Eliane, P.C.; Bruna, A.B.; Carolina, D.S.S.; Carlos, A.V.B.; Eliana, B.F.; Janaina, F.D.M.B. Carotenoids from Phaffia Rhodozyma: Antioxidant Activity and Stability of Extracts. Afr. J. Biotechnol. 2015, 14, 1982–1988. [Google Scholar] [CrossRef]
- Bu, X.; Lin, J.; Duan, C.; Koffas, M.A.G.; Yan, G. Dual Regulation of Lipid Droplet-Triacylglycerol Metabolism and ERG9 Expression for Improved β-Carotene Production in Saccharomyces Cerevisiae. Microb. Cell Factories 2022, 21, 3. [Google Scholar] [CrossRef]
- Hwang, C.Y.; Cho, E.-S.; Bae, E.H.; Jung, D.-H.; Seo, M.-J. Carotenoid-Producing Paracoccus aurantius sp. nov., Isolated from the West Coast of Dokdo Island, Republic of Korea. J. Microbiol. Biotechnol. 2024, 34, 2012–2022. [Google Scholar] [CrossRef] [PubMed]
- Hayashi, M.; Ishibashi, T.; Maoka, T. Effect of Astaxanthin-Rich Extract Derived from Paracoccus Carotinifaciens on Cognitive Function in Middle-Aged and Older Individuals. J. Clin. Biochem. Nutr. 2018, 62, 195–205. [Google Scholar] [CrossRef] [PubMed]
- Flegler, A.; Lipski, A. The C50 Carotenoid Bacterioruberin Regulates Membrane Fluidity in Pink-Pigmented Arthrobacter Species. Arch. Microbiol. 2021, 204, 70. [Google Scholar] [CrossRef]
- Hagaggi, N.S.A.; Abdul-Raouf, U.M. Production of Bioactive β-Carotene by the Endophytic Bacterium Citricoccus Parietis AUCs with Multiple in Vitro Biological Potentials. Microb. Cell Factories 2023, 22, 90. [Google Scholar] [CrossRef]
- Jinendiran, S.; Dahms, H.-U.; Dileep Kumar, B.S.; Kumar Ponnusamy, V.; Sivakumar, N. Diapolycopenedioic-Acid-Diglucosyl Ester and Keto-Myxocoxanthin Glucoside Ester: Novel Carotenoids Derived from Exiguobacterium acetylicum S01 and Evaluation of Their Anticancer and Anti-Inflammatory Activities. Bioorganic Chem. 2020, 103, 104149. [Google Scholar] [CrossRef]
- Gharibzahedi, S.M.T.; Razavi, S.H.; Mousavi, M. Carotenoid Production from Hydrolyzed Molasses by Dietzia Natronolimnaea HS-1 Using Batch, Fed-Batch and Continuous Culture. Ann. Microbiol. 2014, 64, 945–953. [Google Scholar] [CrossRef]
- De Carbon, C.B.; Thurotte, A.; Wilson, A.; Perreau, F.; Kirilovsky, D. Biosynthesis of Soluble Carotenoid Holoproteins in Escherichia Coli. Sci. Rep. 2015, 5, 9085. [Google Scholar] [CrossRef] [PubMed]
- Xinrui, D.; Bo, L.; Yihong, B.; Weifeng, L.; Yong, T. Metabolic Engineering of Escherichia Coli for High-Level Production of Violaxanthin. Microb. Cell Factories 2023, 22, 115. [Google Scholar] [CrossRef] [PubMed]
- Choi, B.H.; Hwang, H.J.; Lee, J.E.; Oh, S.H.; Hwang, J.S.; Lee, B.Y.; Lee, P.C. Microbial Production of Retinyl Palmitate and Its Application as a Cosmeceutical. Antioxidants 2020, 9, 1130. [Google Scholar] [CrossRef]
- Gunes, S.; Tamburaci, S.; Dalay, M.C.; Deliloglu Gurhan, I. In Vitro Evaluation of Spirulina Platensis Extract Incorporated Skin Cream with Its Wound Healing and Antioxidant Activities. Pharm. Biol. 2017, 55, 1824–1832. [Google Scholar] [CrossRef] [PubMed]
- Darvin, M.E.; Jung, S.; Schanzer, S.; Richter, H.; Kurth, E.; Thiede, G.; Meinke, M.C.; Lademann, J. Influence of the Systemic Application of Blue–Green Spirulina Platensis Algae on the Cutaneous Carotenoids and Elastic Fibers in Vivo. Cosmetics 2015, 2, 302–312. [Google Scholar] [CrossRef]
- McLoughlin, I.J.; Voss, A.L.; Hale, J.D.F.; Jain, R. Cosmetic Efficacy of the Topical Probiotic Micrococcus Luteus Q24 in Healthy Human Adults. Cosmetics 2024, 11, 122. [Google Scholar] [CrossRef]
- Castro-Puyana, M.; Pérez-Sánchez, A.; Valdés, A.; Ibrahim, O.H.M.; Suarez-Álvarez, S.; Ferragut, J.A.; Micol, V.; Cifuentes, A.; Ibáñez, E.; García-Cañas, V. Pressurized Liquid Extraction of Neochloris oleoabundans for the Recovery of Bioactive Carotenoids with Anti-Proliferative Activity against Human Colon Cancer Cells. Food Res. Int. 2017, 99, 1048–1055. [Google Scholar] [CrossRef] [PubMed]
- Gupte, Y.; Iyer, G.; Desai, S.; Iyer, M.; Sawant, V.; Moramkar, N. Characterization of High Carotenoid Producing Coelastrella Oocystiformis and Its Anti-Cancer Potential. Int. J. Curr. Microbiol. Appl. Sceinces 2024, 4, 527–536. [Google Scholar]
- Milosheska, D.; Roškar, R. Use of Retinoids in Topical Antiaging Treatments: A Focused Review of Clinical Evidence for Conventional and Nanoformulations. Adv. Ther. 2022, 39, 5351. [Google Scholar] [CrossRef] [PubMed]
- De Souza Guedes, L.; Martinez, R.M.; Bou-Chacra, N.A.; Velasco, M.V.R.; Rosado, C.; Baby, A.R. An Overview on Topical Administration of Carotenoids and Coenzyme Q10 Loaded in Lipid Nanoparticles. Antioxidants 2021, 10, 1034. [Google Scholar] [CrossRef] [PubMed]
- Kulawik-Pióro, A.; Miastkowska, M. Polymeric Gels and Their Application in the Treatment of Psoriasis Vulgaris: A Review. Int. J. Mol. Sci. 2021, 22, 5124. [Google Scholar] [CrossRef] [PubMed]
- Elango, J.; Zamora-Ledezma, C.; Negrete-Bolagay, D.; Aza, P.N.D.; Gómez-López, V.M.; López-González, I.; Belén Hernández, A.; De Val, J.E.M.S.; Wu, W. Retinol-Loaded Poly(Vinyl Alcohol)-Based Hydrogels as Suitable Biomaterials with Antimicrobial Properties for the Proliferation of Mesenchymal Stem Cells. Int. J. Mol. Sci. 2022, 23, 15623. [Google Scholar] [CrossRef]
- Hamed, R.; Abu Nahia, B.J.; Alkilani, A.Z.; Al-Adhami, Y.; Obaidat, R. Recent Advances in Microneedling-Assisted Cosmetic Applications. Cosmetics 2024, 11, 51. [Google Scholar] [CrossRef]
- Sinopoli, A.; Caminada, S.; Isonne, C.; Santoro, M.M.; Baccolini, V. What Are the Effects of Vitamin A Oral Supplementation in the Prevention and Management of Viral Infections? A Systematic Review of Randomized Clinical Trials. Nutrients 2022, 14, 4081. [Google Scholar] [CrossRef] [PubMed]
- Al-Mekhlafi, H.M.; Al-Zabedi, E.M.; Al-Maktari, M.T.; Atroosh, W.M.; Al-Delaimy, A.K.; Moktar, N.; Sallam, A.A.; Abdullah, W.A.; Jani, R.; Surin, J. Effects of Vitamin A Supplementation on Iron Status Indices and Iron Deficiency Anaemia: A Randomized Controlled Trial. Nutrients 2014, 6, 190–206. [Google Scholar] [CrossRef]
- Saini, R.K.; Prasad, P.; Lokesh, V.; Shang, X.; Shin, J.; Keum, Y.-S.; Lee, J.-H. Carotenoids: Dietary Sources, Extraction, Encapsulation, Bioavailability, and Health Benefits—A Review of Recent Advancements. Antioxidants 2022, 11, 795. [Google Scholar] [CrossRef] [PubMed]
- Conboy Stephenson, R.; Ross, R.P.; Stanton, C. Carotenoids in Milk and the Potential for Dairy Based Functional Foods. Foods 2021, 10, 1263. [Google Scholar] [CrossRef] [PubMed]
- Roy, S.; Deshmukh, R.K.; Tripathi, S.; Gaikwad, K.K.; Das, S.S.; Sharma, D. Recent Advances in the Carotenoids Added to Food Packaging Films: A Review. Foods 2023, 12, 4011. [Google Scholar] [CrossRef] [PubMed]
- Gomes, C.d.C.; Passos, T.S.; Morais, A.H.A. Vitamin A Status Improvement in Obesity: Findings and Perspectives Using Encapsulation Techniques. Nutrients 2021, 13, 1921. [Google Scholar] [CrossRef] [PubMed]
- Mujica-Álvarez, J.; Gil-Castell, O.; Barra, P.A.; Ribes-Greus, A.; Bustos, R.; Faccini, M.; Matiacevich, S. Encapsulation of Vitamins A and E as Spray-Dried Additives for the Feed Industry. Molecules 2020, 25, 1357. [Google Scholar] [CrossRef] [PubMed]
- Gonçalves, A.; Estevinho, B.N.; Rocha, F. Microencapsulation of Vitamin A: A Review. Trends Food Sci. Technol. 2016, 51, 76–87. [Google Scholar] [CrossRef]
- Drosou, C.; Krokida, M. A Comparative Study of Encapsulation of β-Carotene via Spray-Drying and Freeze-Drying Techniques Using Pullulan and Whey Protein Isolate as Wall Material. Foods 2024, 13, 1933. [Google Scholar] [CrossRef]
- De Lima, P.M.; Dacanal, G.C.; Pinho, L.S.; de Sá, S.H.G.; Thomazini, M.; Favaro-Trindade, C.S. Combination of Spray-Chilling and Spray-Drying Techniques to Protect Carotenoid-Rich Extracts from Pumpkin (Cucurbita Moschata) Byproducts, Aiming at the Production of a Powdered Natural Food Dye. Molecules 2022, 27, 7530. [Google Scholar] [CrossRef]
- Pinho, L.S.; de Lima, P.M.; de Sá, S.H.G.; Chen, D.; Campanella, O.H.; da Costa Rodrigues, C.E.; Favaro-Trindade, C.S. Encapsulation of Rich-Carotenoids Extract from Guaraná (Paullinia Cupana) Byproduct by a Combination of Spray Drying and Spray Chilling. Foods 2022, 11, 2557. [Google Scholar] [CrossRef]
- Łupina, K.; Kowalczyk, D.; Kazimierczak, W. Gum Arabic/Gelatin and Water-Soluble Soy Polysaccharides/Gelatin Blend Films as Carriers of Astaxanthin—A Comparative Study of the Kinetics of Release and Antioxidant Properties. Polymers 2021, 13, 1062. [Google Scholar] [CrossRef] [PubMed]
- Pinna, N.; Blasi, F.; Schoubben, A. Optimization of Solid Lipid Nanoparticles for the Encapsulation of Carotenoids from Cucurbita Moschata Pulp. Eng. Proc. 2023, 37, 5. [Google Scholar] [CrossRef]
- Ortega-Regules, A.E.; Martínez-Thomas, J.A.; Schürenkämper-Carrillo, K.; de Parrodi, C.A.; López-Mena, E.R.; Mejía-Méndez, J.L.; Lozada-Ramírez, J.D. Recent Advances in the Therapeutic Potential of Carotenoids in Preventing and Managing Metabolic Disorders. Plants 2024, 13, 1584. [Google Scholar] [CrossRef]
- Torres-Montilla, S.; Rodriguez-Concepcion, M. Making Extra Room for Carotenoids in Plant Cells: New Opportunities for Biofortification. Prog. Lipid Res. 2021, 84, 101128. [Google Scholar] [CrossRef] [PubMed]
- Zhong, J.; Zhao, N.; Song, Q.; Du, Z.; Shu, P. Topical Retinoids: Novel Derivatives, Nano Lipid-Based Carriers, and Combinations to Improve Chemical Instability and Skin Irritation. J. Cosmet. Dermatol. 2024, 23, 3102–3115. [Google Scholar] [CrossRef] [PubMed]
- Spierings, N.M.K. Evidence for the Efficacy of Over-the-Counter Vitamin A Cosmetic Products in the Improvement of Facial Skin Aging: A Systematic Review. J. Clin. Aesthet. Dermatol. 2021, 14, 33–40. [Google Scholar] [PubMed]
- Martínez-Iglesias, O.; Naidoo, V.; Carrera, I.; Corzo, L.; Cacabelos, R. Natural Bioactive Products as Epigenetic Modulators for Treating Neurodegenerative Disorders. Pharmaceuticals 2023, 16, 216. [Google Scholar] [CrossRef]
- Acevedo, N.; Alashkar Alhamwe, B.; Caraballo, L.; Ding, M.; Ferrante, A.; Garn, H.; Garssen, J.; Hii, C.S.; Irvine, J.; Llinás-Caballero, K.; et al. Perinatal and Early-Life Nutrition, Epigenetics, and Allergy. Nutrients 2021, 13, 724. [Google Scholar] [CrossRef]
- Roche, F.C.; Harris-Tryon, T.A. Illuminating the Role of Vitamin A in Skin Innate Immunity and the Skin Microbiome: A Narrative Review. Nutrients 2021, 13, 302. [Google Scholar] [CrossRef] [PubMed]













Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2025 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).