
The Effect of Germination Duration on the Biochemical Indicators and Functional-Technological Properties of Triticale Grain

 , , , , ,
, , , , ,
Abstract

1. Introduction
2. Materials and Methods
2.1. Materials, Chemicals, and Reagents Used
2.2. Germination of Whole Triticale Grains
2.3. Sample Preparation
2.4. Determination of Biochemical Indicators
2.4.1. Amino Acid Composition
2.4.2. Vitamins
2.4.3. Minerals
2.4.4. Total Phenols and Antioxidant Activity
2.5. Determination of Functional-Technological Properties
2.6. Viscosity
2.7. Statistical Analysis
3. Results
3.1. Biochemical Indicators
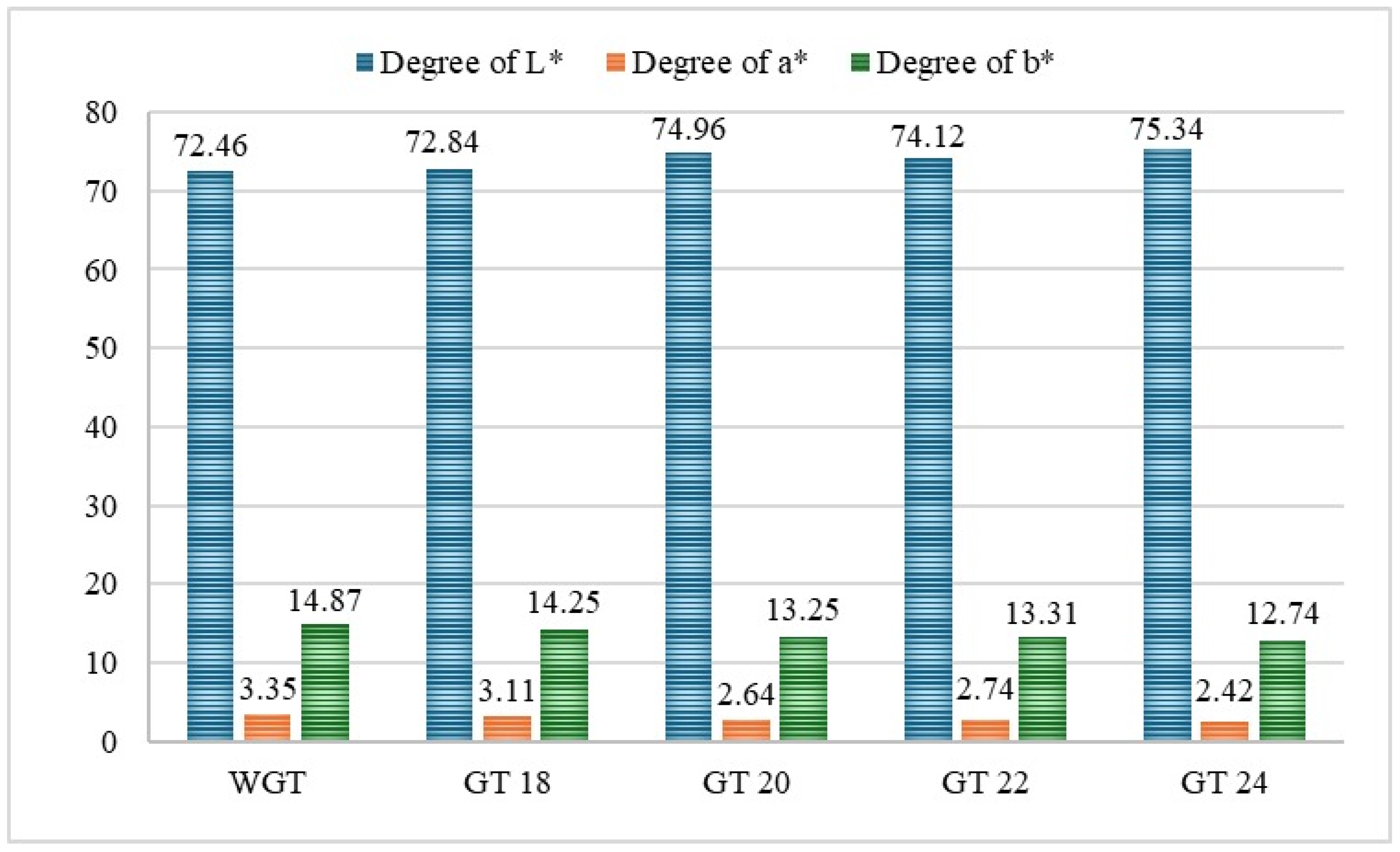
3.1.1. Color Profile
3.1.2. Amino Acid Composition
3.1.3. Vitamins
3.1.4. Minerals
3.1.5. Total Phenolic Content and Antioxidant Activity
3.2. Functional and Technological Indicators
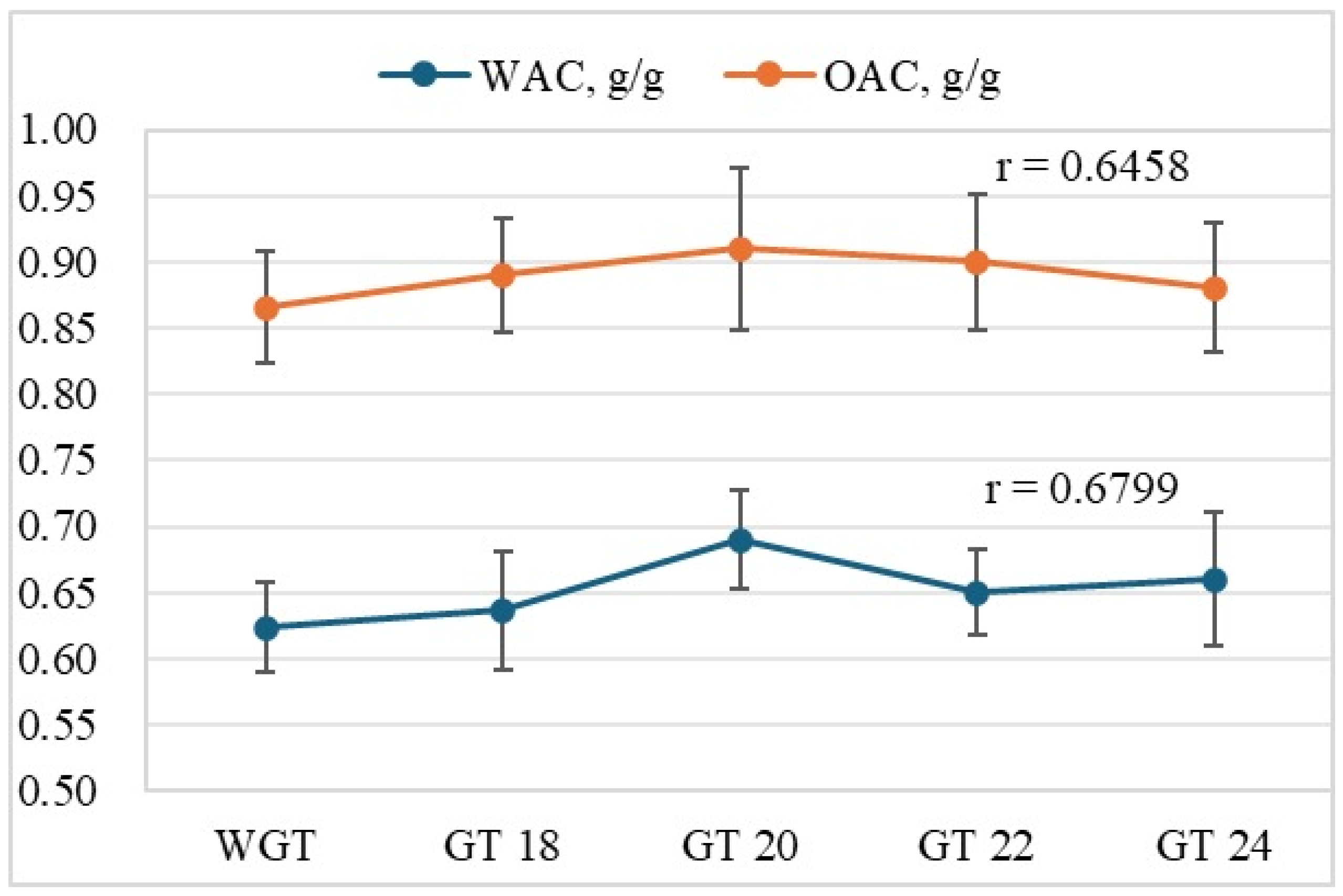
3.2.1. Water and Oil Absorption Capacities
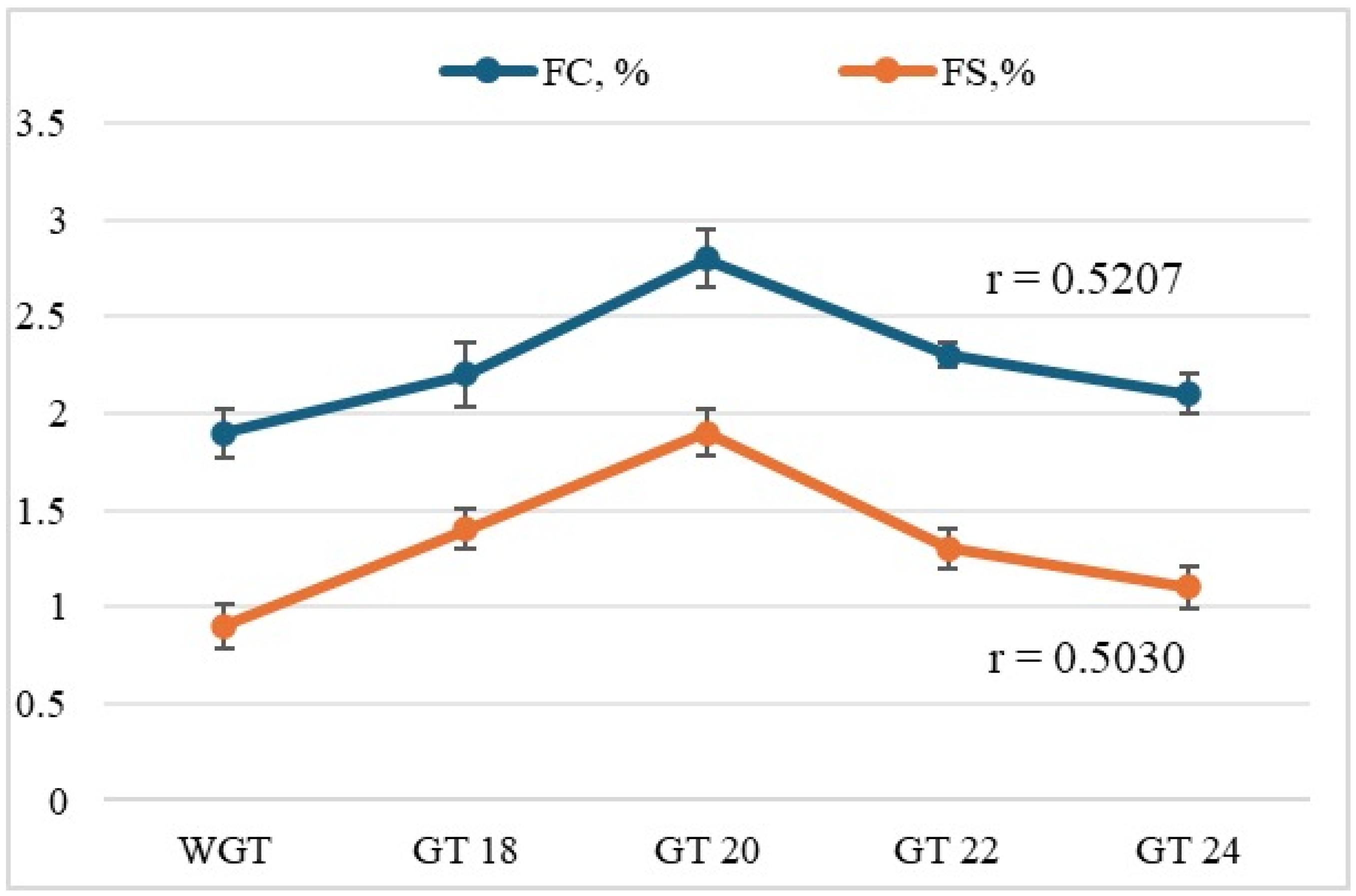
3.2.2. Foam Formation Ability and Foam Stability
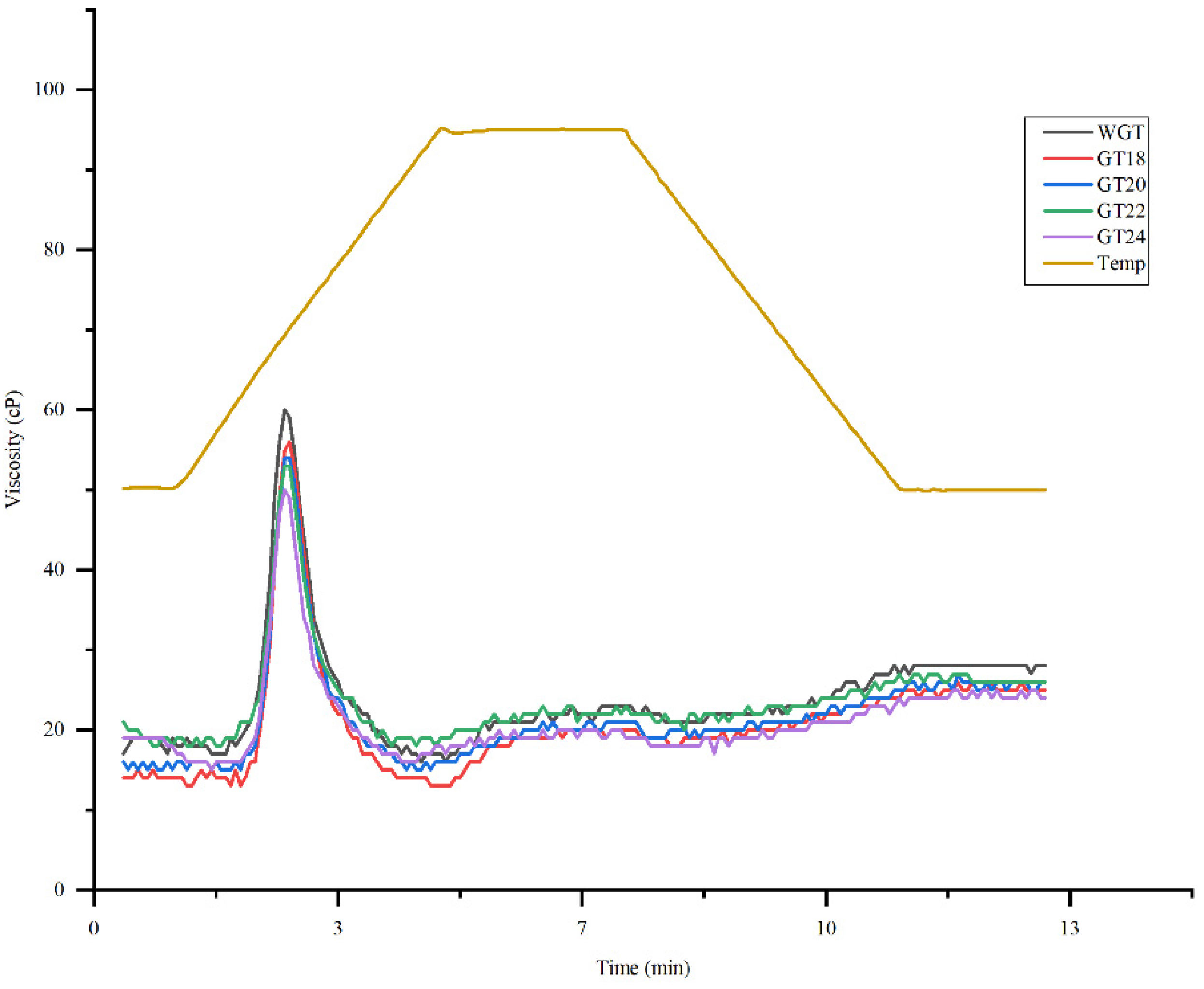
3.3. Determination of Pasting Properties of Whole and Germinated Triticale Grain
4. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Data Availability Statement
Conflicts of Interest
Abbreviations
| MP MN | Measurement Procedure Metrological Norm |
| GT | Germination Time |
| WGT | Whole-Grain Triticale |
| EN | European Norm |
| AACCI | American Association of Cereal Chemists International |
| n/d | Not detected |
References
- McGoverin, C.M.; Snyders, F.; Muller, N.; Botes, W.; Fox, G.; Manley, M. A review of triticale uses and the effect of growth environment on grain quality. J. Sci. Food Agric. 2011, 91, 1155–1165. [Google Scholar] [CrossRef] [PubMed]
- Zhu, F. Triticale: Nutritional composition and food uses. Food Chem. 2018, 241, 468–479. [Google Scholar] [CrossRef] [PubMed]
- Fraś, A.; Gołębiewska, K.; Gołębiewski, D.; Mańkowski, D.R.; Boros, D.; Szecówka, P. Variability in the chemical composition of triticale grain, flour and bread. J. Cereal Sci. 2016, 71, 66–72. [Google Scholar] [CrossRef]
- Camerlengo, F.; Kiszonas, A.M. Genetic factors influencing triticale quality for food. J. Cereal Sci. 2023, 113, 103744. [Google Scholar] [CrossRef]
- Buchholz, M.; Drotleff, A.M.; Ternes, W. Thiamin (vitamin B1) and thiamin phosphate esters in five cereal grains during maturation. J. Cereal Sci. 2012, 56, 109–114. [Google Scholar] [CrossRef]
- Hosseinian, F.; Mazza, G. Triticale bran and straw: Potential new sources of phenolic acids, proanthocyanidins, and lignans. J. Funct. Foods 2009, 1, 57–64. [Google Scholar] [CrossRef]
- Makowska, A.; Szwengiel, A.; Kubiak, P.; Tomaszewska-Gras, J. Characteristics and structure of starch isolated from triticale. Starch-Stärke 2014, 66, 895–902. [Google Scholar] [CrossRef]
- Messina, V.; Cano, J.; Silvio, A.; Pattison, A.L.; Roberts, T.H. Wholegrain triticale sourdough: Effects of triticale: Wheat flour ratio and hydration level on bread quality. Food Sci. Nutr. 2024, 12, 3910–3919. [Google Scholar] [CrossRef]
- Kaszuba, J.; Jaworska, G.; Krochmal-Marczak, B.; Kogut, B.; Kuźniar, P. Effect of bran addition on rheological properties of dough and quality of triticale bread. J. Food Process. Preserv. 2021, 45, e15093. [Google Scholar] [CrossRef]
- Langó, B.; Bóna, L.; Ács, E.; Tömösközi, S. Nutritional features of triticale as affected by genotype, crop year, and location. Acta Aliment. 2017, 46, 238–245. [Google Scholar] [CrossRef]
- Dennett, A.L.; Wilkes, M.A.; Trethowan, R.M. Characteristics of modern triticale quality: The relationship between carbohydrate properties, α-amylase activity, and falling number. Cereal Chem. 2013, 90, 594–600. [Google Scholar] [CrossRef]
- Sibian, M.S.; Saxena, D.C.; Riar, C.S. Effect of germination on chemical, functional and nutritional characteristics of wheat, brown rice and triticale: A comparative study. J. Sci. Food Agric. 2017, 97, 4643–4651. [Google Scholar] [CrossRef]
- Meija, L.; Havensone, G.; Lejnieks, A. Postprandial glycaemic and insulinaemic responses after consumption of activated wheat and triticale grain flakes. J. Nutr. Metab. 2019, 2019, 6594896. [Google Scholar] [CrossRef] [PubMed]
- Piazza, I.; Carnevali, P.; Faccini, N.; Baronchelli, M.; Terzi, V.; Morcia, C.; Ghizzoni, R.; Patrone, V.; Morelli, L.; Cervini, M. Combining native and malted triticale flours in biscuits: Nutritional and technological implications. Foods 2023, 12, 3418. [Google Scholar] [CrossRef]
- Allai, F.M.; Azad, Z.; Gul, K.; Dar, B. Wholegrains: A review on the amino acid profile, mineral content, physicochemical, bioactive composition and health benefits. Int. J. Food Sci. Technol. 2022, 57, 1849–1865. [Google Scholar] [CrossRef]
- Xu, M.; Jin, Z.; Simsek, S.; Hall, C.; Rao, J.; Chen, B. Effect of germination on the chemical composition, thermal, pasting, and moisture sorption properties of flours from chickpea, lentil, and yellow pea. Food Chem. 2019, 295, 579–587. [Google Scholar] [CrossRef] [PubMed]
- Rodríguez-España, M.; Figueroa-Hernández, C.Y.; de Dios Figueroa-Cárdenas, J.; Rayas-Duarte, P.; Hernández-Estrada, Z.J. Effects of germination and lactic acid fermentation on nutritional and rheological properties of sorghum: A graphical review. Curr. Res. Food Sci. 2022, 5, 807–812. [Google Scholar] [CrossRef]
- Lemmens, E.; Moroni, A.V.; Pagand, J.; Heirbaut, P.; Ritala, A.; Karlen, Y.; Lê, K.A.; Van den Broeck, H.C.; Brouns, F.J.; De Brier, N. Impact of cereal seed sprouting on its nutritional and technological properties: A critical review. Compr. Rev. Food Sci. Food Saf. 2019, 18, 305–328. [Google Scholar] [CrossRef] [PubMed]
- Singh, A.; Sharma, S. Bioactive components and functional properties of biologically activated cereal grains: A bibliographic review. Crit. Rev. Food Sci. Nutr. 2017, 57, 3051–3071. [Google Scholar] [CrossRef]
- Wang, T.; He, F.; Chen, G. Improving bioaccessibility and bioavailability of phenolic compounds in cereal grains through processing technologies: A concise review. J. Funct. Foods 2014, 7, 101–111. [Google Scholar] [CrossRef]
- Benincasa, P.; Falcinelli, B.; Lutts, S.; Stagnari, F.; Galieni, A. Sprouted grains: A comprehensive review. Nutrients 2019, 11, 421. [Google Scholar] [CrossRef] [PubMed]
- Sangronis, E.; Machado, C. Influence of germination on the nutritional quality of Phaseolus vulgaris and Cajanus cajan. LWT-Food Sci. Technol. 2007, 40, 116–120. [Google Scholar] [CrossRef]
- Frias, J.; Miranda, M.L.; Doblado, R.; Vidal-Valverde, C. Effect of germination and fermentation on the antioxidant vitamin content and antioxidant capacity of Lupinus albus L. var. Multolupa. Food Chem. 2005, 92, 211–220. [Google Scholar] [CrossRef]
- Ghavidel, R.A.; Prakash, J. The impact of germination and dehulling on nutrients, antinutrients, in vitro iron and calcium bioavailability and in vitro starch and protein digestibility of some legume seeds. LWT-Food Sci. Technol. 2007, 40, 1292–1299. [Google Scholar] [CrossRef]
- Guzmán-Ortiz, F.A.; Castro-Rosas, J.; Gómez-Aldapa, C.A.; Mora-Escobedo, R.; Rojas-León, A.; Rodríguez-Marín, M.L.; Falfán-Cortés, R.N.; Román-Gutiérrez, A.D. Enzyme activity during germination of different cereals: A review. Food Rev. Int. 2019, 35, 177–200. [Google Scholar] [CrossRef]
- Nguyen, B.C.Q.; Shahinozzaman, M.; Tien, N.T.K.; Thach, T.N.; Tawata, S. Effect of sucrose on antioxidant activities and other health-related micronutrients in gamma-aminobutyric acid (GABA)-enriched sprouting Southern Vietnam brown rice. J. Cereal Sci. 2020, 93, 102985. [Google Scholar] [CrossRef]
- Sahai, V.; Kumar, V. Anti-diabetic, hepatoprotective and antioxidant potential of Brassica oleracea sprouts. Biocatal. Agric. Biotechnol. 2020, 25, 101623. [Google Scholar] [CrossRef]
- Nelson, K.; Mathai, M.L.; Ashton, J.F.; Donkor, O.N.; Vasiljevic, T.; Mamilla, R.; Stojanovska, L. Effects of malted and non-malted whole-grain wheat on metabolic and inflammatory biomarkers in overweight/obese adults: A randomised crossover pilot study. Food Chem. 2016, 194, 495–502. [Google Scholar] [CrossRef]
- Kadiri, O. A review on the status of the phenolic compounds and antioxidant capacity of the flour: Effects of cereal processing. Int. J. Food Prop. 2017, 20, S798–S809. [Google Scholar] [CrossRef]
- Chinma, C.E.; Adewuyi, O.; Abu, J.O. Effect of germination on the chemical, functional and pasting properties of flour from brown and yellow varieties of tigernut (Cyperus esculentus). Food Res. Int. 2009, 42, 1004–1009. [Google Scholar] [CrossRef]
- Ding, H.; Fu, T.J.; Smith, M.A. Microbial contamination in sprouts: How effective is seed disinfection treatment? J. Food Sci. Technol. 2013, 78, R495–R501. [Google Scholar] [CrossRef]
- Malik, I.O.M.; Yousif, S.A.; Ali, A.E.; Hamadnalla, H.M. Effect of germination on proximate composition of three grains from Sudan. J. Nutr. Food Sci. 2021, 3, 104–109. [Google Scholar]
- Baranzelli, J.; Kringel, D.H.; Colussi, R.; Paiva, F.F.; Aranha, B.C.; de Miranda, M.Z.; da Rosa Zavareze, E.; Dias, A.R.G. Changes in enzymatic activity, technological quality and gamma-aminobutyric acid (GABA) content of wheat flour as affected by germination. LWT 2018, 90, 483–490. [Google Scholar] [CrossRef]
- Marti, A.; Cardone, G.; Pagani, M.A.; Casiraghi, M.C. Flour from sprouted wheat as a new ingredient in bread-making. LWT 2018, 89, 237–243. [Google Scholar] [CrossRef]
- Dong, L.; Piao, Y.; Zhang, X.; Zhao, C.; Hou, Y.; Shi, Z. Analysis of volatile compounds from a malting process using headspace solid-phase micro-extraction and GC–MS. Food Res. Int. 2013, 51, 783–789. [Google Scholar] [CrossRef]
- Wu, F.; Yang, N.; Chen, H.; Jin, Z.; Xu, X. Effect of germination on flavor volatiles of cooked brown rice. Cereal Chem. 2011, 88, 497–503. [Google Scholar] [CrossRef]
- Waleed, A.-A.; Fadhl, J.A.; Abdullah, A.B.; Al-Adeeb, A.; Mahdi, A.A.; Al-Maqtari, Q.A.; Mushtaq, B.S.; Fan, M.; Li, Y.; Qian, H. Effect of highland barely germination on thermomechanical, rheological, and micro-structural properties of wheat-oat composite flour dough. Food Biosci. 2023, 53, 102521. [Google Scholar]
- Sharma, P.; Gujral, H.S. Antioxidant and polyphenol oxidase activity of germinated barley and its milling fractions. Food Chem. 2010, 120, 673–678. [Google Scholar] [CrossRef]
- Kruma, Z.; Tomsone, L.; Ķince, T.; Galoburda, R.; Senhofa, S.; Sabovics, M.; Straumite, E.; Sturite, I. Effects of germination on total phenolic compounds and radical scavenging activity in hull-less spring cereals and triticale. Agron. Res. 2016, 14, 1372–1383. [Google Scholar]
- AACC Intemational. Approved methods of analysis. In Rheological Behavior of Flour by Farinograph: Constant Flour Weight Procedure; AACC International: St. Paul, MN, USA, 2010. [Google Scholar]
- Alviola, J.N.A.; Monterde, V.G. Physicochemical and Functional Properties of Wheat (Triticum aestivum) and Selected Local Flours in the Philippines. Philipp. J. Sci. 2018, 147, 419–430. [Google Scholar]
- Lohdip, A.; Jikmyan, M. Proximate and amino acid analyses of the stem of Sesamum indicum L. (Black Seed Specie) Pedaliaceae. J. Chem. Soc. Niger. 2019, 44, 65–70. [Google Scholar]
- EN 12822:2014; Foodstuffs—Determination of Vitamin E by High Performance Liquid Chromatography—Measurement of α-, β-, γ- and δ-Tocopherol. Comité Européen de Normalisation (CEN): Brussels, Belgium, 2014.
- EN 14122:2014; Foodstuffs—Determination of Vitamin B1 by High Performance Liquid Chromatography. Comité Européen de Normalisation (CEN): Brussels, Belgium, 2014.
- EN 14152:2014; Foodstuffs—Determination of Vitamin B2 by High Performance Liquid Chromatography. Comité Européen de Normalisation (CEN): Brussels, Belgium, 2014.
- EN 15652:2009; Foodstuffs—Determination of Niacin by HPLC. Comité Européen de Normalisation (CEN): Brussels, Belgium, 2009.
- MP MN 3008-2008; Method for Determining the Mass Fraction of Pantothenic Acid in Specialized Food Products and Dietary Supplements. SI RSPC of Hygiene, MH RB: Minsk, Republic of Belarus, 2008.
- EN 14663:2006; Foodstuffs—Determination of Vitamin B6 (Including Its Glycosylated Forms) by HPLC. Comité Européen de Normalisation (CEN): Brussels, Belgium, 2006.
- MP MN 2146-2004; Method for the Determination of Folic Acid in Fortified Food Products. SI RSPC of Hygiene, MH RB: Minsk, Republic of Belarus, 2004.
- Ragaee, S.; Abdel-Aal, E.-S.M.; Noaman, M. Antioxidant activity and nutrient composition of selected cereals for food use. Food Chem. 2006, 98, 32–38. [Google Scholar] [CrossRef]
- Marchini, M.; Carini, E.; Cataldi, N.; Boukid, F.; Blandino, M.; Ganino, T.; Vittadini, E.; Pellegrini, N. The use of red lentil flour in bakery products: How do particle size and substitution level affect rheological properties of wheat bread dough? LWT 2021, 136, 110299. [Google Scholar] [CrossRef]
- Abd Elmoneim, O.E.; Bernhardt, R. Influence of grain germination on functional properties of sorghum flour. Food Chem. 2010, 121, 387–392. [Google Scholar]
- Coffmann, C.; Garciaj, V. Functional properties and amino acid content of a protein isolate from mung bean flour. Int. J. Food Sci. Technol. 1977, 12, 473–484. [Google Scholar] [CrossRef]
- Liu, S.; Wang, W.; Lu, H.; Shu, Q.; Zhang, Y.; Chen, Q. New perspectives on physiological, biochemical and bioactive components during germination of edible seeds: A review. Trends Food Sci. Technol. 2022, 123, 187–197. [Google Scholar] [CrossRef]
- Singkhornart, S.; Ryu, G.-H.J.P.N.; Science, F. Effect of soaking time and steeping temperature on biochemical properties and γ-aminobutyric acid (GABA) content of germinated wheat and barley. Prev. Nutr. Food Sci. 2011, 16, 67–73. [Google Scholar] [CrossRef]
- Kruma, Z.; Kince, T.; Galoburda, R.; Tomsone, L.; Straumite, E.; Sabovics, M.; Sturite, I.; Kronberga, A. Influence of germination temperature and time on phenolic content and antioxidant properties of cereals. In Proceedings of the Baltic Conference on Food Science and Technology: Conference Proceedings, Jelgava, Latvia, 2–3 May 2019. [Google Scholar]
- Ahmed, W.E.; Ragab, I.; Gadallah, M.G.; Alhomaid, R.M.; Almujaydil, M.S. Effect of sprouting whole wheat grain on the sensory quality, physicochemical properties, and antioxidant activity of cupcakes. Appl. Food Res. 2024, 4, 100412. [Google Scholar] [CrossRef]
- El-Safy, F.; Salem, R.; Mukhtar, E.Y. The impact of soaking and germination on chemical composition, carbohydrate fractions, digestibility, antinutritional factors and minerals content of some legumes and cereals grain seeds. Alex. Sci. Exch. J. 2013, 34, 499–513. [Google Scholar]
- Ongol, M.P.; Niyonzima, E.; Gisanura, I.; Vasanthakaalam, H. Effect of germination and fermentation on nutrients in maize flour. Pak. J. Food Sci. 2013, 23, 183–188. [Google Scholar]
- Erba, D.; Angelino, D.; Marti, A.; Manini, F.; Faoro, F.; Morreale, F.; Pellegrini, N.; Casiraghi, M.C. Effect of sprouting on nutritional quality of pulses. Int. J. Food Sci. Nutr. 2019, 70, 30–40. [Google Scholar] [CrossRef] [PubMed]
- Afify, A.E.-M.M.; El-Beltagi, H.S.; Abd El-Salam, S.M.; Omran, A.A. Protein solubility, digestibility and fractionation after germination of sorghum varieties. PLoS ONE 2012, 7, e31154. [Google Scholar] [CrossRef]
- Tizazu, S.; Urga, K.; Abuye, C.; Retta, N. Improvement of energy and nutrient density of sorghumbased complementary foods using germination. Afr. J. Food Agric. Nutr. Dev. 2010, 10, 2927–2942. [Google Scholar] [CrossRef]
- Oser, B.L. An integrated essential amino acid index for predicting the biological value of proteins. In Protein and Amino Acid Nutrition; Elsevier: Amsterdam, The Netherlands, 1959; pp. 281–295. [Google Scholar]
- Steve, I.O. Influence of germination and fermentation on chemical composition, protein quality and physical properties of wheat flour (Triticum aestivum). J. Cereals Oil Seeds 2012, 3, 35–47. [Google Scholar]
- Lemar, L.E.; Swanson, B.G. Nutritive value of sprouted wheat flour. J. Food Sci. 1976, 41, 719–720. [Google Scholar] [CrossRef]
- Bewley, J.D.; Bradford, K.J.; Hilhorst, H.W.; Nonogaki, H.; Bewley, J.D.; Bradford, K.J.; Hilhorst, H.W.; Nonogaki, H. Mobilization of stored reserves. In Seeds: Physiology of Development, Germination and Dormancy, 3rd ed.; Springer: Berlin/Heidelberg, Germany, 2013; pp. 183–246. [Google Scholar]
- Pal, R.; Bhartiya, A.; Yadav, P.; Kant, L.; Mishra, K.; Aditya, J.; Pattanayak, A. Effect of dehulling, germination and cooking on nutrients, anti-nutrients, fatty acid composition and antioxidant properties in lentil (Lens culinaris). J. Food Sci. Technol. 2017, 54, 909–920. [Google Scholar] [CrossRef]
- Yang, B.; Yin, Y.; Liu, C.; Zhao, Z.; Guo, M. Effect of germination time on the compositional, functional and antioxidant properties of whole wheat malt and its end-use evaluation in cookie-making. Food Chem. 2021, 349, 129125. [Google Scholar] [CrossRef]
- Guardianelli, L.M.; Salinas, M.V.; Puppo, M.C. Chemical and thermal properties of flours from germinated amaranth seeds. J. Food Meas. Charact. 2019, 13, 1078–1088. [Google Scholar] [CrossRef]
- Afify, A.E.-M.M.; El-Beltagi, H.S.; Abd El-Salam, S.M.; Omran, A.A. Effect of soaking, cooking, germination and fermentation processing on proximate analysis and mineral content of three white sorghum varieties (Sorghum bicolor L. Moench). Not. Bot. Horti Agrobot. Cluj-Napoca 2012, 40, 92–98. [Google Scholar]
- Teixeira, C.; Nyman, M.; Andersson, R.; Alminger, M. Effects of variety and steeping conditions on some barley components associated with colonic health. J. Sci. Food Agric. 2016, 96, 4821–4827. [Google Scholar] [CrossRef]
- Donkor, O.; Stojanovska, L.; Ginn, P.; Ashton, J.; Vasiljevic, T. Germinated grains–Sources of bioactive compounds. Food Chem. 2012, 135, 950–959. [Google Scholar] [CrossRef] [PubMed]
- Kavitha, S.; Parimalavalli, R. Effect of processing methods on proximate composition of cereal and legume flours. J. Hum. Nutr. Food Sci. 2014, 2, 1051. [Google Scholar]
- Jan, R.; Saxena, D.; Singh, S. Physico-chemical, textural, sensory and antioxidant characteristics of gluten–Free cookies made from raw and germinated Chenopodium (Chenopodium album) flour. LWT-Food Sci. Technol. 2016, 71, 281–287. [Google Scholar] [CrossRef]
- Kubicka, E.; Grabska, J.; Jedrychowski, L.; Czyz, B. Changes of specific activity of lipase and lipoxygenase during germination of wheat and barley. Int. J. Food Sci. Nutr. 2000, 51, 301. [Google Scholar] [PubMed]
- Poudel, R.; Finnie, S.; Rose, D.J. Effects of wheat kernel germination time and drying temperature on compositional and end-use properties of the resulting whole wheat flour. J. Cereal Sci. 2019, 86, 33–40. [Google Scholar] [CrossRef]
- Kumar, A.; Kaur, A.; Gupta, K.; Gat, Y.; Kumar, V. Assessment of germination time of finger millet for value addition in functional foods. Curr. Sci. 2021, 120, 406–413. [Google Scholar] [CrossRef]
- Singh, A.; Sharma, S.; Singh, B. Effect of germination time and temperature on the functionality and protein solubility of sorghum flour. J. Cereal Sci. 2017, 76, 131–139. [Google Scholar] [CrossRef]
- Gunathunga, C.; Senanayake, S.; Jayasinghe, M.A.; Brennan, C.S.; Truong, T.; Marapana, U.; Chandrapala, J. Germination effects on nutritional quality: A comprehensive review of selected cereals and pulses changes. J. Food Compos. Anal. 2024, 128, 106024. [Google Scholar] [CrossRef]
- Dhillon, B.; Choudhary, G.; Sodhi, N.S. A study on physicochemical, antioxidant and microbial properties of germinated wheat flour and its utilization in breads. J. Food Sci. Technol. 2020, 57, 2800–2808. [Google Scholar] [CrossRef]
- Sharma, S.; Saxena, D.C.; Riar, C.S. Analysing the effect of germination on phenolics, dietary fibres, minerals and γ-amino butyric acid contents of barnyard millet (Echinochloa frumentaceae). Food Biosci. 2016, 13, 60–68. [Google Scholar] [CrossRef]
- Suda, M.; Watanabe, T.; Kobayashi, M.; Matsuda, K. Changes in starch content and related enzyme activities during the growth of germinating soybeans. Agric. Biol. Chem. 1986, 50, 3195–3196. [Google Scholar]
- Enujiugha, V.N.; Badejo, A.A.; Iyiola, S.O.; Oluwamukomi, M.O. Effect of germination on the nutritional and functional properties of African oil bean (Pentaclethra macrophylla Benth) seed flour. J. Food Agric. Environ. 2003, 1, 72–75. [Google Scholar]
- Murungweni, K.T.; Ramashia, S.E.; Mashau, M.E. Effect of malting on physicochemical, antioxidant, and microstructural properties of finger millet (Eleusine coracana) flours. Food Sci. Nutr. 2024, 12, 547–563. [Google Scholar] [CrossRef] [PubMed]
- Nefale, F.E.; Mashau, M.E. Effect of germination period on the physicochemical, functional and sensory properties of finger millet flour and porridge. Asian J. Appl. Sci. 2018, 6, 360–367. [Google Scholar] [CrossRef]
- Perveen, S.; Akhtar, S.; Ismail, T.; Qamar, M.; Saeed, W.; Younis, M.; Esatbeyoglu, T. Comparison of nutritional, antioxidant, physicochemical, and rheological characteristics of whole and sprouted wheat flour. LWT 2024, 209, 116679. [Google Scholar] [CrossRef]
- Wu, Y.V. Lysine content of triticale protein increased by germination. J. Agric. Food Chem. 1982, 30, 820–823. [Google Scholar] [CrossRef]
- Smith, D. The amino acid composition of barley grain protein during development and germination. J. Agric. Sci. 1972, 78, 265–273. [Google Scholar] [CrossRef]
- Li, Q.; Xu, J.-G. Changes in nutritive value and in vitro digestibility of proteins from naked oats during germination. Eur. J. Food Sci. Technol. 2015, 3, 49–57. [Google Scholar] [CrossRef]
- Kamjijam, B.; Bednarz, H.; Suwannaporn, P.; Jom, K.N.; Niehaus, K. Localization of amino acids in germinated rice grain: Gamma-aminobutyric acid and essential amino acids production approach. J. Cereal Sci. 2020, 93, 102958. [Google Scholar] [CrossRef]
- Rutherfurd, S.M.; Bains, K.; Moughan, P.J. Available lysine and digestible amino acid contents of proteinaceous foods of India. Br. J. Nutr. 2012, 108, S59–S68. [Google Scholar] [CrossRef]
- Singh, M.; Rao, D.M.; Pande, S.; Battu, S.; Mahalakshmi, K.; Dutt, K.R.; Ramesh, M. Medicinal uses of L-lysine: Past and future. Int. J. Res. Pharm. Sci. 2011, 2, 637–642. [Google Scholar]
- Sánchez, A.; Rubano, D.A.; Shavlik, G.W.; Hubbard, R.; Horning, M.C. Cholesterolemic effects of the lysine/arginine ratio in rabbits after initial early growth. Arch. Latinoam. Nutr. 1988, 38, 229–238. [Google Scholar]
- Majzoobi, M.; Wang, Z.; Teimouri, S.; Pematilleke, N.; Brennan, C.S.; Farahnaky, A. Unlocking the potential of sprouted cereals, pseudocereals, and pulses in combating malnutrition. Foods 2023, 12, 3901. [Google Scholar] [CrossRef] [PubMed]
- Kim, H.Y.; Hwang, I.G.; Kim, T.M.; Woo, K.S.; Park, D.S.; Kim, J.H.; Kim, D.J.; Lee, J.; Lee, Y.R.; Jeong, H.S. Chemical and functional components in different parts of rough rice (Oryza sativa L.) before and after germination. Food Chem. 2012, 134, 288–293. [Google Scholar] [CrossRef]
- Žilić, S.; Delić, N.; Basić, Z.; Ignjatović-Micić, D.; Janković, M.; Vančetović, J. Effects of alkaline cooking and sprouting on bioactive compounds, their bioavailability and relation to antioxidant capacity of maize flour. J. Food Nutr. Res. 2015, 54, 155. [Google Scholar]
- Okorie, S.; Ekwe, C. The comparative analysis of sprouted legume and cereal based composite diet. J. Appl. Biotechnol. Bioeng. 2017, 4, 554–561. [Google Scholar]
- Chaves-López, C.; Rossi, C.; Maggio, F.; Paparella, A.; Serio, A. Changes occurring in spontaneous maize fermentation: An overview. Fermentation 2020, 6, 36. [Google Scholar] [CrossRef]
- Rico, D.; Peñas, E.; García, M.d.C.; Martínez-Villaluenga, C.; Rai, D.K.; Birsan, R.I.; Frias, J.; Martín-Diana, A.B. Sprouted barley flour as a nutritious and functional ingredient. Foods 2020, 9, 296. [Google Scholar] [CrossRef]
- Kariluoto, S.; Liukkonen, K.-H.; Myllymäki, O.; Vahteristo, L.; Kaukovirta-Norja, A.; Piironen, V. Effect of germination and thermal treatments on folates in rye. J. Agric. Food Chem. 2006, 54, 9522–9528. [Google Scholar] [CrossRef]
- Plaza, L.; de Ancos, B.; Cano, P.M. Nutritional and health-related compounds in sprouts and seeds of soybean (Glycine max), wheat (Triticum aestivum. L) and alfalfa (Medicago sativa) treated by a new drying method. Eur. Food Res. Technol. 2003, 216, 138–144. [Google Scholar] [CrossRef]
- Malleshi, N.; Klopfenstein, C. Nutrient composition, amino acid and vitamin contents of malted sorghum, pearl millet, finger millet and their rootlets. Int. J. Food Sci. Nutr. 1998, 49, 415–422. [Google Scholar] [CrossRef]
- Hucker, B.; Wakeling, L.; Vriesekoop, F. Investigations into the thiamine and riboflavin content of malt and the effects of malting and roasting on their final content. J. Cereal Sci. 2012, 56, 300–306. [Google Scholar] [CrossRef]
- Moongngarm, A.; Saetung, N. Comparison of chemical compositions and bioactive compounds of germinated rough rice and brown rice. Food Chem. 2010, 122, 782–788. [Google Scholar] [CrossRef]
- Ferreira, C.D.; Bubolz, V.K.; da Silva, J.; Dittgen, C.L.; Ziegler, V.; de Oliveira Raphaelli, C.; de Oliveira, M. Changes in the chemical composition and bioactive compounds of chickpea (Cicer arietinum L.) fortified by germination. LWT 2019, 111, 363–369. [Google Scholar] [CrossRef]
- Steve Ijarotimi, O.; Ruth Esho, T. Comparison of nutritional composition and anti-nutrient status of fermented, germinated and roasted bambara groundnut seeds (vigna subterranea). Food Chem. 2009, 111, 376–386. [Google Scholar] [CrossRef]
- Afify, A.E.-M.M.; El-Beltagi, H.S.; Abd El-Salam, S.M.; Omran, A.A. Bioavailability of iron, zinc, phytate and phytase activity during soaking and germination of white sorghum varieties. PLoS ONE 2011, 6, e25512. [Google Scholar] [CrossRef]
- Hübner, F.; O’Neil, T.; Cashman, K.D.; Arendt, E.K. The influence of germination conditions on beta-glucan, dietary fibre and phytate during the germination of oats and barley. Eur. Food Res. Technol. 2010, 231, 27–35. [Google Scholar] [CrossRef]
- Egli, I.; Davidsson, L.; Juillerat, M.; Barclay, D.; Hurrell, R. The influence of soaking and germination on the phytase activity and phytic acid content of grains and seeds potentially useful for complementary feedin. J. Food Sci. 2002, 67, 3484–3488. [Google Scholar] [CrossRef]
- Mostafa, A.M.; Yassin, E.; Eisa, Y.; Abou El-Enien, N.F. Effect of infestation with acaridid mite, Tyrophagus putrescentiae (Schrank) on germination rate of maize grains. Egypt. Acad. J. Biol. Sci. A Entomol. 2013, 6, 81–85. [Google Scholar] [CrossRef]
- Rosique-Esteban, N.; Guasch-Ferré, M.; Hernández-Alonso, P.; Salas-Salvadó, J. Dietary magnesium and cardiovascular disease: A review with emphasis in epidemiological studies. Nutrients 2018, 10, 168. [Google Scholar] [CrossRef]
- Sadawarte, S.; Pawar, V.; Sawate, A.; Thorat, P.; Shere, P.; Surendar, J. Effect of germination on vitamin and mineral content of horse gram and green gram malt. IJCS 2018, 6, 1761–1764. [Google Scholar]
- Agidew, M.G.; Dubale, A.A.; Atlabachew, M.; Abebe, W. Fatty acid composition, total phenolic contents and antioxidant activity of white and black sesame seed varieties from different localities of Ethiopia. Chem. Biol. Technol. Agric. 2021, 8, 14. [Google Scholar] [CrossRef]
- Zeb, A. Concept, mechanism, and applications of phenolic antioxidants in foods. J. Food Biochem. 2020, 44, e13394. [Google Scholar] [CrossRef]
- Tian, S.; Nakamura, K.; Kayahara, H. Analysis of phenolic compounds in white rice, brown rice, and germinated brown rice. J. Agric. Food Chem. 2004, 52, 4808–4813. [Google Scholar] [CrossRef] [PubMed]
- Lemus, C.; Angelis, A.; Halabalaki, M.; Skaltsounis, A.L. γ-Oryzanol: An attractive bioactive component from rice bran. In Wheat and Rice in Disease Prevention and Health; Elsevier: Amsterdam, The Netherlands, 2014; pp. 409–430. [Google Scholar]
- Gawlik-Dziki, U.; Dziki, D.; Nowak, R.; Świeca, M.; Olech, M.; Pietrzak, W. Influence of sprouting and elicitation on phenolic acids profile and antioxidant activity of wheat seedlings. J. Cereal Sci. 2016, 70, 221–228. [Google Scholar] [CrossRef]
- Gao, Y.; Guo, X.; Liu, Y.; Zhang, M.; Zhang, R.; Abbasi, A.M.; You, L.; Li, T.; Liu, R.H. Comparative assessment of phytochemical profile, antioxidant capacity and anti-proliferative activity in different varieties of brown rice (Oryza sativa L.). LWT 2018, 96, 19–25. [Google Scholar] [CrossRef]
- Gernah, D.; Ariahu, C.; Ingbian, E. Effects of malting and lactic fermentation on some chemical and functional properties of maize (Zea mays). Am. J. Food Technol. 2011, 6, 404–412. [Google Scholar] [CrossRef]
- Adedeji, O.; Oyinloye, O.; Ocheme, O. Effects of germination time on the functional properties of maize flour and the degree of gelatinization of its cookies. Afr. J. Food Sci. 2014, 8, 42–47. [Google Scholar] [CrossRef]
- Akaerue, B.I.; Onwuka, G.I. The functional properties of the dehulled and undehulled mung bean (Vigna radiata (l.) Wilczek) flours as influenced by processing treatments. J. Agric. Vet. Sci. 2010, 2, 1–28. [Google Scholar]
- Offia Olua, B.; Onwuzuruike, U.; Nwankpa, M. The effect of different processing treatments on the proximate composition and functional properties of maize-mung bean composite flours. J. Food Stab. 2020, 3, 12–25. [Google Scholar] [CrossRef]
- Siddiqua, A.; Ali, M.; Ahmed, S. Functional properties of germinated and non-germinated cereals: A comparative study. Bangladesh J. Sci. Ind. Res. 2019, 54, 383–390. [Google Scholar] [CrossRef]
- Hussain, I.; Uddin, M.B. Optimization effect of germination on functional properties of wheat flour by response surface methodology. Int. Res. J. Plant Sci. 2012, 3, 31–37. [Google Scholar]
- Hung, S.; Zayas, J. Protein solubility, water retention, and fat binding of corn germ protein flour compared with milk proteins. J. Food Sci. 1992, 57, 372–376. [Google Scholar] [CrossRef]
- Wang, X.-H.; Tai, Z.-J.; Song, X.-J.; Li, Z.-J.; Zhang, D.-J. Effects of germination on the structure, functional properties, and in vitro digestibility of a black bean (Glycine max (L.) Merr.) protein isolate. Foods 2024, 13, 488. [Google Scholar] [CrossRef]
- Ocheme, O.; Adedeji, O.; Lawal, G.; Zakari, U. Effect of germination on functional properties and degree of starch gelatinization of sorghum flour. J. Food Res. 2015, 4, 159. [Google Scholar]
- Eltayeb, A.; Ali, A.O.; Abou-Arab, A.A.; Abu-Salem, F.M. Chemical composition and functional properties of flour and protein isolate extracted from Bambara groundnut (Vigna subterranean). Afr. J. Food Sci. 2011, 5, 82–90. [Google Scholar]
- Pork, B.; Yoon, K.Y. Functional properties of enzymatic hydrolysate and peptide fractions from perilla seed meal protein. Pol. J. Food Nutr. Sci. 2019, 69, 119–127. [Google Scholar] [CrossRef]
- Foegeding, E.A.; Luck, P.; Davis, J.P. Factors determining the physical properties of protein foams. Food Hydrocoll. 2006, 20, 284–292. [Google Scholar] [CrossRef]
- Huang, J.; Wang, Z.; Fan, L.; Ma, S. A review of wheat starch analyses: Methods, techniques, structure and function. Int. J. Biol. Macromol. 2022, 203, 130–142. [Google Scholar] [CrossRef]
- Cardone, G.; Grassi, S.; Scipioni, A.; Marti, A. Bread-making performance of durum wheat as affected by sprouting. LWT 2020, 134, 110021. [Google Scholar] [CrossRef]


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Treatment, h | Moisture, % | Protein, % | Fat, % | Ash, % | Carbohydrates, Including Dietary Fiber |
|---|---|---|---|---|---|
| WGT | 6.42 ± 0.06 a | 15.58 ± 0.44 ab | 1.77 ± 0.11 c | 1.45 ± 0.07 c | 74.46 ± 0.25 b |
| GT18 | 7.91 ± 0.04 b | 15.65 ± 0.14 abc | 1.64 ± 0.09 bc | 1.41 ± 0.06 c | 73.36 ± 0.41 a |
| GT20 | 7.25 ± 0.32 b | 16.27 ± 0.24 c | 1.62 ± 0.03 abc | 1.40 ± 0.14 a | 73.44 ± 0.08 a |
| GT22 | 7.59 ± 0.27 b | 16.12 ± 0.36 bc | 1.58 ± 0.05 ab | 1.39 ± 0.03 b | 73.26 ± 0.33 a |
| GT24 | 7.64 ± 0.19 b | 15.07 ± 0.06 a | 1.56 ± 0.03 a | 1.42 ± 0.03 c | 73.18 ± 0.29 b |
| Germination Time, % | Arginine | Lysine | Tyrosine | Phenylala-Nine | Histidine | Leucine and Isoleucine | Methionine | Valin | Proline | Threonine | Serin | Alanine | Glycine |
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| WGT | 0.86 ± 0.05 ab | 0.49 ± 0.07 a | 0.43 ± 0.07 a | 0.75 ± 0.08 a | 0.45 ± 0.09 a | 0.85 ± 0.07 ab | 0.28 ± 0.06 a | 0.70 ± 0.07 a | 1.85 ± 0.07 b | 0.59 ± 0.07 a | 0.93 ± 0.06 ab | 0.64 ± 0.06 a | 0.75 ± 0.05 ab |
| GT18 | 0.82 ± 0.08 ab | 0.44 ± 0.07 a | 0.43 ± 0.04 a | 0.73 ± 0.07 a | 0.38 ± 0.03 a | 0.83 ± 0.08 ab | 0.30 ± 0.09 a | 0.66 ± 0.07 a | 1.84 ± 0.05 b | 0.60 ± 0.05 a | 0.90 ± 0.05 ab | 0.63 ± 0.07 a | 0.70 ± 0.06 ab |
| GT20 | 0.95 ± 0.08 b | 0.53 ± 0.09 a | 0.46 ± 0.06 a | 0.81 ± 0.07 a | 0.46 ± 0.05 a | 0.98 ± 0.08 b | 0.34 ± 0.05 a | 0.76 ± 0.01 a | 2.08 ± 0.07 c | 0.70 ± 0.05 a | 1.03 ± 0.07 b | 0.74 ± 0.08 a | 0.82 ± 0.07 b |
| GT22 | 0.87 ± 0.05 ab | 0.53 ± 0.06 a | 0.44 ± 0.08 a | 0.77 ± 0.09 a | 0.40 ± 0.06 a | 0.87 ± 0.06 ab | 0.29 ± 0.05 a | 0.73 ± 0.06 a | 1.89 ± 0.07 a | 0.68 ± 0.09 a | 0.96 ± 0.05 ab | 0.73 ± 0.07 a | 0.79 ± 0.07 ab |
| GT24 | 0.74 ± 0.07 a | 0.42 ± 0.08 a | 0.40 ± 0.06 a | 0.67 ± 0.06 a | 0.35 ± 0.05 a | 0.77 ± 0.08 a | 0.28 ± 0.07 a | 0.62 ± 0.09 a | 1.75 ± 0.07 b | 0.57 ± 0.06 a | 0.87 ± 0.07 a | 0.60 ± 0.07 a | 0.65 ± 0.07 a |
| Germination Time, h | Vitamin Content, mg/kg | ||||||
|---|---|---|---|---|---|---|---|
| B1 | B2 | B3 | B5 | B6 | B9 | E | |
| WGT | n/d | n/d | 0.029 ± 0.002 a | n/d | 0.06 ± 0.06 a | 0.19 ± 0.01 a | 2.98 ± 0.29 b |
| GT 18 | n/d | n/d | 0.34 ± 0.03 c | n/d | n/d | 1.66 ± 0.16 b | 1.84 ± 0.18 a |
| GT 20 | n/d | n/d | 0.19 ± 0.01 b | n/d | 0.045 ± 0.004 a | 0.04 ± 0.004 a | 2.01 ± 0.20 a |
| GT 22 | 0.17 ± 0.01 c | 0.48 ± 0.04 a | 0.52 ± 0.05 d | n/d | n/d | 0.06 ± 0.006 a | 2.03 ± 0.20 a |
| GT 24 | 0.11 ± 0.01 b | 2.51 ± 0.25 b | 0.48 ± 0.04 d | n/d | n/d | 0.09 ± 0.009 a | 2.28 ± 0.22 a |
| Germination Time, h | Dynamics of Total Phenolic Content (TPC) During Germination: TPC, mg/100 g | Dynamics of DPPH Radical-Scavenging Activity, DPPH Activity During Germination: DPPH, mg/100 g |
|---|---|---|
| WGT | 42 ± 0.283 a | 380 ± 0.425 a |
| GT18 | 49 ± 0.424 b | 457 ± 0.290 b |
| GT20 | 51 ± 0.560 b | 458 ± 0.324 c |
| GT22 | 52 ± 0.490 c | 458 ± 0.291 c |
| GT24 | 54 ± 0.286 c | 459 ± 0.191 d |
| Germination Time | Peak Viscosity | Trough | Breakdown Viscosity | Final Viscosity | Setback Viscosity |
|---|---|---|---|---|---|
| WGT | 60.00 ± 3.17 b | 21.00 ± 2.09 a | 39.00 ± 1.47 c | 28.00 ± 1.99 a | 7.00 ± 0.86 a |
| GT 18 | 56.00 ± 1.89 ab | 18.00 ± 2.53 a | 38.00 ± 1.82 bc | 25.00 ± 2.14 a | 7.00 ± 0.14 a |
| GT 20 | 54.00 ± 2.87 ab | 19.00 ± 1.04 a | 35.00 ± 2.33 ab | 26.00 ± 1.17 a | 7.00 ± 0.37 a |
| GT 22 | 53.00 ± 3.27 ab | 20.00 ± 2.70 a | 33.00 ± 1.30 a | 26.00 ± 1.94 a | 6.00 ± 1.05 a |
| GT 24 | 50.00 ± 2.77 a | 17.00 ± 1.39 a | 33.00 ± 1.59 a | 24.00 ± 1.39 a | 7.00 ± 0.33 a |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2025 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Ospankulova, G.; Temirova, I.; Toimbayeva, D.; Saduakhasova, S.; Tazhina, S.; Khamitova, D.; Muratkhan, M.; Aldiyeva, A.; Zhumalin, A. The Effect of Germination Duration on the Biochemical Indicators and Functional-Technological Properties of Triticale Grain. Appl. Sci. 2025, 15, 5596. https://doi.org/10.3390/app15105596
Ospankulova G, Temirova I, Toimbayeva D, Saduakhasova S, Tazhina S, Khamitova D, Muratkhan M, Aldiyeva A, Zhumalin A. The Effect of Germination Duration on the Biochemical Indicators and Functional-Technological Properties of Triticale Grain. Applied Sciences. 2025; 15(10):5596. https://doi.org/10.3390/app15105596
Chicago/Turabian StyleOspankulova, Gulnazym, Indira Temirova, Dana Toimbayeva, Saule Saduakhasova, Sayagul Tazhina, Dina Khamitova, Marat Muratkhan, Akmaral Aldiyeva, and Aibek Zhumalin. 2025. "The Effect of Germination Duration on the Biochemical Indicators and Functional-Technological Properties of Triticale Grain" Applied Sciences 15, no. 10: 5596. https://doi.org/10.3390/app15105596
APA StyleOspankulova, G., Temirova, I., Toimbayeva, D., Saduakhasova, S., Tazhina, S., Khamitova, D., Muratkhan, M., Aldiyeva, A., & Zhumalin, A. (2025). The Effect of Germination Duration on the Biochemical Indicators and Functional-Technological Properties of Triticale Grain. Applied Sciences, 15(10), 5596. https://doi.org/10.3390/app15105596