Abstract
Fatty acids (FAs) are the major structural component of erythrocyte membranes. Diet and physical exercise directly influence their incorporation and function. Endurance runners engage in high volumes of weekly aerobic training, alternating between low-intensity and high-intensity sessions. The aim of the study was to assess and compare the erythrocyte FA profile in a group of high-level male endurance runners (EG) with a control group of non-athlete subjects (CG). This observational study was conducted on 85 subjects, 63 high-level male endurance runners (23 ± 3 years; height: 1.76 ± 0.05) and 22 subjects who did not engage in regular physical exercise (21 ± 0.5 years; height: 1.68 ± 0.39). Runners had at least five years of training experience, and all of them were participants in national and international tournaments. FAs determination was performed using gas chromatography. Higher percentages of Palmitic Acid (PA), Stearic Acid (SA), Oleic Acid (OA), Calendic Acid (CA), Eicosapentaenoic Acid (EPA) and Docosapentaenoic Acid (DPA), and lower percentages of Docosahexaenoic Acid (DHA) were found in the EG compared to the CG. High-level endurance runners exhibit altered erythrocyte FA profiles with low percentages of omega-3 index (ω-3 index) and DHA, which may affect erythrocyte membrane function as well as their performance.
1. Introduction
Endurance runners engage in regular training sessions characterized by predominance of moderate and low-intensity sessions alternated with high-intensity ones [1]. Fatty acids (FAs) are an important source of energy during exercise, and their oxidation increases during low and moderate intensity [2]. Nutrition plays a crucial role in the recovery and adaptation processes of these athletes [3]. For years, runners have restricted lipid intake due to concerns about its accumulation as ectopic fat and potential negative impact on performance [4].
Lipids are macronutrients with essential functions in the body. They act as an energy source, hormone precursors, and participate in immune function [5]. FAs are essential components of erythrocyte membranes, and it is known that the composition of FAs ingested in the diet is related to those stored in the membrane [6]. Changes in the composition of erythrocyte membrane FAs can modify their properties and function [7]. It has been reported that dietary FA intake is reflected in plasma within days but takes months to be incorporated into erythrocytes [8]. Erythrocytes have a lifespan of 120 days and are considered an ideal matrix for evaluating long-term FA intake [9].
Polyunsaturated fatty acids (PUFAs) are essential FAs that must be ingested through the diet and have significant benefits for human health [10]. The fluidity and flexibility of the cell membrane are primarily determined by the presence of PUFAs in the lipid bilayers [11]. Omega-3 (ω-3) and Omega-6 (ω-6) families have different functions and compete for the same enzymatic pathways and incorporation into the cell membrane [12]. Proinflammatory eicosanoids are derived from arachidonic acid (AA), an ω-6 FA that in high concentrations contributes to thrombus formation, atheroma, and increased inflammation [13]. However, the intake of ω-3 FAs such as docosahexaenoic acid (DHA) and eicosapentaenoic acid (EPA) increases the concentrations of eicosanoids with vasodilator, platelet aggregation inhibitory, and anti-inflammatory properties [14]. In athletes, various studies have reported that ω-3 FAs optimize recovery processes and contribute to improving the immune system [15].
Physical exercise is another factor that can modify the FAs of erythrocyte membranes. It is known that physical exercise increases the amount of reactive oxygen species (ROS) and free radicals [16]. Since erythrocyte membranes contain a large amount of PUFAs and are exposed to high levels of oxygen, they are susceptible to lipid peroxidation and potential muscle damage [17]. It has been demonstrated that moderate and low-intensity aerobic exercise favors vasodilation and an improvement in immune and anti-inflammatory response [18]. However, high-intensity training increases the production of superoxide radicals in lipid bilayers, leading to increased inflammation and the possibility of oxidative stress and damage to erythrocytes [19]. Additionally, the high intake of ω-6 from current diets is another factor contributing to an increased ω-6/ω-3 inflammatory ratio, which relates to the proportion of ω-6 and ω-3 FAs [12].
Different studies have reported the erythrocyte profile of different sports specialties such as football and basketball [20], water polo [21], rugby [22], and hockey [23]. These sports are characterized by highly competitive activity and the performance of high-intensity actions and training [24]. These athletes showed altered profiles compared to the control population, with insufficient ω-3 values as a consequence of chronic training and inadequate FA intake. However, endurance runners have lower competitive density during the season and different physiological demands. Runners undergo high training volumes, with 70–80% of the completed kilometers below the second ventilatory threshold (VT2) and a reduced percentage above VT2 [25]. Regular aerobic training modifies the lipidomic profile of the erythrocyte membranes [26]. Changes in the membrane FAs composition affect their functions, flexibility, and fluidity, in order to improve the efficiency of oxygen delivery [18]. Few studies have evaluated the FA profile in erythrocytes of high-level endurance runners. In a study by Kamada et al. [27], they reported higher percentages of PUFAs in the erythrocyte membrane of Japanese endurance runners compared to control subjects. However, the sample size was only six runners, and the study was conducted with Oriental athletes who have a different diet from the Mediterranean one. Therefore, the aim of this study was to assess the FA profile in erythrocytes in a group of 63 high-level endurance runners who lived in the same area as a result of chronic training and compare with a group of healthy students who did not perform regular physical exercise.
2. Materials and Methods
2.1. Participants
Eighty-five male subjects who lived in the same geographical area participated in the present study. The participants were divided into two groups: 63 high-level male endurance runners (EG: 23 ± 3 years) who competed at the national/international races and a 22-male control group (CG: 21.23 ± 0.49 years) composed of healthy students.
Before the research, all participants signed a written informed consent. The study protocol was approved by the Ethics Committee of the University of Extremadura (52/2012) and carried out according to the 1975 Declaration of Helsinki, revised at the World Medical Assembly of Fortaleza (Brazil) in 2012 for research with human beings.
Inclusion criteria in EG were: males who compete in national and international endurance races with at least 5 years of experience and perform more than 5 weekly training sessions. CG inclusion criteria were that they did not perform regular physical exercise. The exclusion criteria were the same in both groups: have followed a specific diet, took FAs supplements, medication, or herbal products in the previous 3 months.
2.2. Design
An observational study was carried out in a specific cohort (high-level endurance runners) in the Physiology Laboratory of Sports Science in Extremadura University, Caceres, Spain, for four years (2012–2016).
2.3. Nutritional Assessment
The nutritional evaluation was carried out following the same methodology used in previous studies by our research group [28]. All participants completed a 3-day diet record on the provided nutritional questionnaire, including one weekend day and two weekdays; each participant weighed and indicated the amount in grams of each food consumed. The dietary intake of the athletes was obtained using a food composition table [29]. Table 1 shows the nutritional intake of lipids and FAs in EG and CG.

Table 1.
Nutritional intake of lipids and FAs in runners and controls.
2.4. Anthropometric Measures
Anthropometric assessments were performed always at the same time, between 9 and 10 a.m. after an overnight fast and utilizing the same methodology of previous studies [30]. The subjects refrained from intense exercise in the previous 72 h to the assessments. All measurements were performed by an experienced, trained, and qualified tester. Body weight (Seca 769, Hamburg, Germany) and height (Seca 220, Hamburg, Germany) were measured to the nearest 0.1 kg and 0.1 cm, following the manufacturer’s guidelines. Six skinfolds (subscapular, triceps, supraspinal, abdominal, thigh, and calf) were measured using a skinfold caliper (Holtain, Crosswell, UK) with a precision of 0.1 mm. Three measurements were made for each fold using the average value in mm following the International Society of Anthropometry and Kinanthropometry (ISAK) Protocol. Fat and muscle weight were calculated following the instructions of the ISAK [31]. Table 2 shows Anthropometric characteristics in the EG and CG.
Table 2.
Anthropometric characteristics in the runners and controls.
2.5. Sample Collection
After anthropometric assessments, 10 mL of blood was extracted from the antecubital vein and placed in tubes with Ethylene Diamine Tetra Acetic Acid (EDTA). Samples were immediately centrifuged for 10 min at 3000 rpm. Erythrocytes were washed with 0.9% sodium chloride (NaCl) three times. Finally, erythrocytes were sited in sterile tubes and stored at −80 °C until use.
2.6. Analytical Determination
This methodology section draws on approaches previously utilized by our group for fatty acid (FA) analysis, as detailed in an earlier study [32]. The concentrations of FAs in erythrocytes were measured using a method established by Lepage and Roy [33]. We employed an HP-5890 Series II gas chromatograph equipped with a Flame Ionization Detector (FID). The analytical column was a BP × 70 capillary columns, 50 m in length, and 0.22 mm in internal diameter, with a film thickness of 0.25 µm, sourced from Cromlab in Barcelona, Spain. The oven’s initial temperature was set at 170 °C, maintained for 15.0 min, then ramped up to 190 °C over 15.0 min at a rate of 3 °C/min, and subsequently to 245 °C at the same rate. The final hold time at 245 °C lasted 30.0 min. Helium gas was used as the carrier at a flow rate of 1.0 mL/min. The injector operated in splitless mode at 300 °C, with a purge flow of 6 mL/min initiated 0.5 min post-injection. The FID temperature was set at 250 °C.
FA identification involved comparing the retention times of the methyl esters of the FAs under study to those of a reference FA pattern, using the same chromatographic conditions and adjusting for the internal standard. Heptadecanoic acid was selected as the internal standard due to its chemical similarity to the target analytes and its effective separation within the chromatogram, avoiding interference with other sample peaks. FA quantification was expressed as a relative percentage of the total FAs, with over 97% of the GC peaks accurately identified against known standards.
2.7. Lipid Profile of the Erythrocyte Membranes
Ten FAs were analyzed: palmitic acid (PA) (C16:0) and stearic acid (SA) (C18:0); oleic acid (OA) (C18:1), alpha linoleic acid (ALA) (C18:3.3), docosapentaeonic acid (DPA) (C22:5), EPA (C20:5) and DHA (C22:6), linoleic acid (LA) (C18:2.6), calendic acid (CA) (C18:3.6) and AA (C20:4). Based on these FAs, different indexes were calculated: (% SFAs/% MUFAs) index related to membrane fluidity or saturation index (SI); ω-3 index (ω-3 IND) (DHA + EPA), inflammatory risk index (%ω-6)/(%ω-3). Additionally, the enzymatic indexes of elongase (C18:0/C16:0), desaturase ω-3 (C22:6/C22:5), and delta 9 (C18:1/C18:0) enzymes were calculated. Optimal value intervals for each of the 10 FAs were based on the literature [9]. After obtaining the percentages of fatty acids, indexes were calculated between the FAs according to Nespolo [34].
2.8. Statistical Analysis
Statistical analysis was conducted using IBM SPSS Statistics software version 21.0 (IBM Co., Armonk, NY, USA). The findings are presented as x ± s, where x represents the mean values and s denotes the standard deviation. Prior to analysis, all variables were assessed for normal distribution using the Shapiro–Wilk test, and Levene’s test was employed to assess homogeneity of variances. Mann–Whitney’s U test was utilized for comparing values between groups. The effect size was calculated [35]. Effect size values of 0.2, 0.4, and 0.8 were considered small, moderate, and large, respectively [36]. A p < 0.05 was considered statistically significant.
3. Results
Table 3 shows the erythrocyte profile of the different FAs in erythrocytes in the EG and CG. Significantly higher percentages (p < 0.05) of EPA (1.26 ± 0.70 vs. 0.73 ± 0.28), DPA (1.98 ± 0.76 vs. 1.39 ± 0.45), and PA (24.52 ± 2.08 vs. 18.34 ± 0.85), and very significant percentages (p < 0.01) of SA (20.22 ± 1.78 vs. 16.24 ± 0.92) and OA (17.39 ± 1.21 vs. 11.93 ± 0.59) are observed in the EG compared to the CG. Furthermore, lower percentages (p < 0.05) of DHA (4.26 ± 1.38 vs. 5.08 ± 1.44) were found in the EG in relation to the CG. There were no significant changes in LA, ALA, AA, ω-3 index, and ω-6/ω-3 ratio.
Table 3.
Difference in erythrocyte membrane fatty acids (%) in endurance runners and the control group.
Table 4 shows total FAs and the activity of different enzymes. Higher percentages (p < 0.05) of SFA (44.04 ± 3.33 vs. 42.06 ± 1.29), MUFA (17.21 ± 1.20 vs. 11.93 ± 0.59), and desaturase 9 (1.22 ± 0.85 vs.0.70 ± 0.06) are observed in EG compared to CG. Lower percentages (p < 0.05) of desaturase ω-3 (2.15 ± 0.97 vs. 3.65 ± 1.12) in EG compared to CG were also reported. There were no differences in PUFA, saturation index, and elongase.
Table 4.
Difference in erythrocyte membrane in total fatty acids (%) and enzymes in endurance runners and controls.
4. Discussion
The aim of our study was to determine the erythrocyte profile of a group of high-level endurance runners and observe the differences compared to a control group of individuals with low physical activity. Physical exercise and nutrition are two variables that modify the composition and function of the erythrocyte membrane [11]. Different FA compositions have been documented in the erythrocyte membrane in athletes from various sports as a result of chronic training, the relationship between the type of sport and the specificity of its training, and the characteristics of its competitions [37].
PUFAs are part of the erythrocyte membrane and regulate its fluidity and plasticity [38]. The function of ω-3 and ω-6 PUFAs in the body differs as both compete for the same enzymatic pathways and their incorporation into the cell membrane. In the ω-6 family, AA is a precursor of proinflammatory eicosanoids, while in the ω-3 family, EPA and DHA promote the production of resolvins and protectins with anti-inflammatory effects [39]. There were no significant differences in the concentrations of different ω-6 PUFAs between EG and CG. However, both groups reported slightly elevated AA levels compared to reference indices. These slightly elevated AA values seem to be a consequence of an ω-3 PUFA deficiency. Various studies have reported elevated AA levels in groups of athletes from different specialties and control groups with low ω-3 concentrations [20,21,37]. In our study, both ER and CG presented a deficiency in the ω-3 index with values significantly below 8%. As mentioned earlier, ω-3 and ω-6 PUFAs compete for the same enzymatic pathways for their incorporation into the cell membrane [12]. In a previous study, Kawataba et al. [40] found a negative relationship between AA concentrations and EPA and DHA in erythrocytes. Therefore, an ω-3 deficiency leads to increased AA incorporation into the erythrocyte. Ultra-processed foods intake, ready to consume and hyperpalatable, has increased in the last years and is associated with poorer diet quality [41]. Hook et al. [23] indicated that athletes were not consuming the recommended amount of ω-3 in their diet. Oily fish (salmon, sardines, shrimp…) are rich sources of ω-3. The main barriers athletes encounter in consuming oily fish are its smell and taste.
EPA and DHA are essential ω-3 PUFAs in the lipid bilayers of erythrocyte membranes [42]. In our study, significantly higher EPA levels were observed in EG compared to CG. EPA has been documented as an essential FA that helps improve cardiovascular health by reducing blood triglycerides and blood pressure [43]. EPA has also been reported to improve the fluidity and flexibility of the cell membrane, enhancing oxygen diffusion and nutrient delivery to the muscle during physical exercise [44]. These benefits are essential in endurance runners who need to optimize their cardiovascular system for optimal performance.
The EG group reported significantly lower DHA levels than the CG. Additionally, the runners had lower values compared to reference indices. DHA deficits have been documented in athletes from different specialties [20,21,22,37,45]. Physical exercise increases the amount of free radicals and reactive oxygen species (ROS) [16]. This increase in ROS leads to an increase in inflammatory cytokines such as TNF-alpha, IL1, IL6, and IL10 that must be balanced by anti-inflammatory systems [46]. DHA has been documented as an FA that reduces lipid peroxidation and erythrocyte damage in a group of athletes as a result of improved superoxide dismutase activity [6]. In this sense, lower DHA concentrations in runners would be a consequence of increased utilization to balance the excess ROS and free radicals produced by training. In this same vein, Allaire et al. [47] observed in a previous study that DHA is a more effective ω-3 PUFA than EPA in modulating inflammatory states.
There were also significantly higher concentrations of DPA in runners compared to CG. DPA plays a fundamental role in resolving inflammatory processes and regulating the immune system [48]. It has recently been reported that the specialized proresolving mediator RvD5 n3DPA can be biosynthesized from DPA when elevated concentrations are available, helping to reduce inflammation [49].
EG reported higher SFAs erythrocyte concentrations of total SFA, PA, and SA than CG. Skeletal muscle uses FAs mainly along with triglycerides as fuel during aerobic exercise [10]. It is known that aerobic training improves insulin sensitivity and FA oxidation [43]. Additionally, it has been documented that the body prioritizes the oxidation of PUFAs as an energy fuel over SFAs during aerobic exercise [50]. Consequently, an increased incorporation of SFAs into membrane erythrocyte occurs as a result of the higher PUFAs oxidation during training sessions in EG. Similarly to our study, higher percentages of SA and PA were reported in endurance runners and athletes of different sports compared to CG [27,37,51]. In this line, a previous study found a positive correlation between SA concentrations with training aerobic volume [52]. Increased incorporation of SA and PA into the erythrocyte membrane have been demonstrated to enhance stiffness while concurrently reducing permeability [26]. This adaptation may be a toll to pay, resulting in enhanced membrane stability and a reduced peroxidation index.
EG showed higher percentages of total MUFA and OA than the CG. The concentrations reported in our runners are similar to those of other groups of athletes [20,21,37]. OA has been reported to reduce the risk of heart disease as it has cardioprotective effects [53]. Additionally, OA has been reported to replace PUFAs in erythrocyte membranes to provide more stability and protect them against free radicals induced by training [54]. Regarding OA, higher percentages of CA were reported in the EG compared to the CG. CA is a PUFA found in vegetables that can be synthesized from OA by the action of fatty acid desaturation (FAD) enzymes as CoFADX-1 and CoFADX-2. Therefore, a greater synthesis of CA occurred in the runners as a result of the higher erythrocyte percentage of OA. Although the current literature on the potential functions of CA is scarce, it has been reported that this FA could exert anti-inflammatory actions [55]
Finally, EG reported significantly higher activity of desaturase 9 compared to the CG, indicating a greater conversion of MUFA from SFA such as PA and SA [56]. Additionally, there was also lower activity of ω-3 desaturase in the EG group, indicating reduced conversion towards long-chain ω-3 fatty acids such as EPA and DHA. Figure 1 shows the main modifications in FAs in erythrocytes in high-level endurance runners due to chronic training.
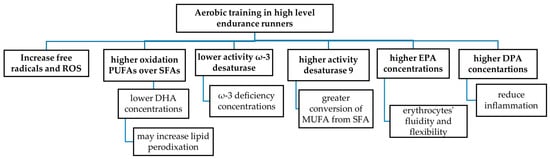
Figure 1.
Fatty acids profile in erythrocytes in high-level endurance runners.
Limitations of the study include the estimative nature of dietary intake, based on a 3-day questionnaire from the week prior to sample collection. Data about endurance runners’ training in the previous months could not be collected. On the other hand, athletes care for their nutrition and usually maintain a suitable and stable diet during the season.
5. Conclusions
Chronic training can modify the FA erythrocyte profile in high-level endurance runners. Runners reported insufficient concentrations of ω-3 index due to a DHA deficiency. There is a greater incorporation of SFAs such as PA and SA, as well as OA in the erythrocyte membrane that can provide more stability to the erythrocytes.
6. Practical Applications
High-level endurance runners should pay special attention to ω-3 intake to assure dietary recommendations in order to avoid deficiency. Runners should incorporate into their diet rich sources of ω-3 such as oily fish (salmon, shrimp, crab, tuna…).
Author Contributions
M.M.-M. designed the study; data were collected and analyzed by F.J.A.-V., G.B.V., I.B.S., F.J.G.P. and V.T.-R.; F.J.A.-V., G.B.V. and M.M.-M. undertook data interpretation and manuscript preparation. Conceptualization, M.M.-M.; methodology, formal analysis, M.M.-M.; investigation, I.B.S., F.J.G.P. and V.T.-R.; writing—original draft preparation, F.J.A.-V. and G.B.V.; writing—review and editing, F.J.A.-V., V.T.-R. and G.B.V. All authors have read and agreed to the published version of the manuscript.
Funding
This research received no external funding.
Institutional Review Board Statement
The study protocol was approved by the Ethics Committee of the University of Extremadura (code: 52/2012; date 9 October 2012) and carried out according to the 1975 Declaration of Helsinki, revised at the World Medical Assembly of Fortaleza (Brazil) in 2013 for research with human beings.
Informed Consent Statement
Written informed consent was obtained from all the participants.
Data Availability Statement
The raw data supporting the conclusions of this article will be made available by the authors on request.
Conflicts of Interest
The authors declare no conflicts of interest.
References
- Enoksen, E.; Tjelta, A.R.; Tjelta, L.I. Distribution of Training Volume and Intensity of Elite Male and Female Track and Marathon Runners. Int. J. Sports Sci. Coach. 2011, 6, 273–293. [Google Scholar] [CrossRef]
- Chycki, J.; Zajac, A.; Michalczyk, M.; Maszczyk, A.; Langfort, J. Hormonal and Metabolic Substrate Status in Response to Exercise in Men of Different Phenotype. Endocr. Connect. 2019, 8, 814–821. [Google Scholar] [CrossRef] [PubMed]
- Beermann, B.L.; Lee, D.G.; Almstedt, H.C.; McCormack, W.P. Nutritional Intake and Energy Availability of Collegiate Distance Runners. J. Am. Coll. Nutr. 2020, 39, 747–755. [Google Scholar] [CrossRef] [PubMed]
- Arrese, A.L.; Ostariz, E.S.; Ostáriz, E.S. Skinfold Thicknesses Associated with Distance Running Performance in Highly Trained Runners. J. Sports Sci. 2006, 24, 69–76. [Google Scholar] [CrossRef] [PubMed]
- Li, T.; Ruan, D.G.; Lin, Z.M.; Liu, T.Y.; Wang, K.; Xu, X.Y.; Duan, R. Endurance Training Counteracts the High-Fat Diet-Induced Profiling Changes of ω-3 Polyunsaturated Fatty Acids in Skeletal Muscle of Middle-Aged Rats. Front. Physiol. 2019, 10, 971. [Google Scholar] [CrossRef] [PubMed]
- Martorell, M.; Pons, V.; Domingo, J.C.; Capó, X.; Sureda, A.; Drobnic, F.; Tur, J.A.; Pons, A. Erythrocytes and Skeletal Muscle Unsaturated and Omega-6 Fatty Acids Are Positively Correlated after Caloric Restriction and Exercise. Ann. Nutr. Metab. 2018, 72, 126–133. [Google Scholar] [CrossRef] [PubMed]
- Jaworska, M.; Siatkowski, S.; Żebrowska, A. The Effects of Omega-3 Fatty Acid Supplementation on the Lipid Profile and Cardiovascular Markers Following Downhill Running in Long-Distance Runners. J. Hum. Kinet. 2023, 89, 123–138. [Google Scholar] [CrossRef] [PubMed]
- Browning, L.M.; Walker, C.G.; Mander, A.P.; West, A.L.; Madden, J.; Gambell, J.M.; Young, S.; Wang, L.; Jebb, S.A.; Calder, P.C. Incorporation of Eicosapentaenoic and Docosahexaenoic Acids into Lipid Pools When given as Supplements Providing Doses Equivalent to Typical Intakes of Oily Fish. Am. J. Clin. Nutr. 2012, 96, 748. [Google Scholar] [CrossRef] [PubMed]
- Ferreri, C.; Chatgilialoglu, C. Role of Fatty Acid-Based Functional Lipidomics in the Development of Molecular Diagnostic Tools. Expert. Rev. Mol. Diagn. 2012, 12, 767–780. [Google Scholar] [CrossRef]
- Philpott, J.D.; Bootsma, N.J.; Rodriguez-Sanchez, N.; Hamilton, D.L.; Mackinlay, E.; Dick, J.; Mettler, S.; Galloway, S.D.R.; Tipton, K.D.; Witard, O.C. Influence of Fish Oil-Derived n-3 Fatty Acid Supplementation on Changes in Body Composition and Muscle Strength during Short-Term Weight Loss in Resistance-Trained Men. Front. Nutr. 2019, 6, 102. [Google Scholar] [CrossRef]
- Nikolaidis, M.G.; Mougios, V. Effects of Exercise on the Fatty-Acid Composition of Blood and Tissue Lipids. Sports Med. 2004, 34, 1051–1076. [Google Scholar] [CrossRef]
- Simopoulos, A.P. The Importance of the Ratio of Omega-6/Omega-3 Essential Fatty Acids. Biomed. Pharmacother. 2002, 56, 365–379. [Google Scholar] [CrossRef]
- Smyth, M.; Lunken, G.; Jacobson, K. Insights into Inflammatory Bowel Disease and Effects of Dietary Fatty Acid Intake with a Focus on Polyunsaturated Fatty Acids Using Preclinical Models. J. Can. Assoc. Gastroenterol. 2023, 7, 104–114. [Google Scholar] [CrossRef] [PubMed]
- Simopoulos, A.P. Omega-3 Fatty Acids and Athletics. Curr. Sports Med. Rep. 2007, 6, 230–236. [Google Scholar] [CrossRef]
- Tomczyk, M.; Heileson, J.L.; Babiarz, M.; Calder, P.C. Athletes Can Benefit from Increased Intake of EPA and DHA—Evaluating the Evidence. Nutrients 2023, 15, 4925. [Google Scholar] [CrossRef] [PubMed]
- He, F.; Li, J.; Liu, Z.W.; Chuang, C.C.; Yang, W.G.; Zuo, L. Redox Mechanism of Reactive Oxygen Species in Exercise. Front. Physiol. 2016, 7, 486. [Google Scholar] [CrossRef]
- Martorell, M.; Capó, X.; Bibiloni, M.M.; Sureda, A.; Mestre-Alfaro, A.; Batle, J.M.; Llompart, I.; Tur, J.A.; Pons, A. Docosahexaenoic Acid Supplementation Promotes Erythrocyte Antioxidant Defense and Reduces Protein Nitrosative Damage in Male Athletes. Lipids 2015, 50, 131–148. [Google Scholar] [CrossRef]
- Paraiso, L.F.; Gonçalves-E-Oliveira, A.F.M.; Cunha, L.M.; De Almeida Neto, O.P.; Pacheco, A.G.; Araújo, K.B.G.; Da Silva Garrote-Filho, M.; Neto, M.B.; Penha-Silva, N. Effects of Acute and Chronic Exercise on the Osmotic Stability of Erythrocyte Membrane of Competitive Swimmers. PLoS ONE 2017, 12, e0171318. [Google Scholar] [CrossRef]
- Cipryan, L. IL-6, Antioxidant Capacity and Muscle Damage Markers Following High-Intensity Interval Training Protocols. J. Hum. Kinet. 2017, 56, 139–148. [Google Scholar] [CrossRef]
- Tepsic, J.; Vucic, V.; Arsic, A.; Blazencic-Mladenovic, V.; Mazic, S.; Glibetic, M. Plasma and Erythrocyte Phospholipid Fatty Acid Profile in Professional Basketball and Football Players. Eur. J. Appl. Physiol. 2009, 107, 359–365. [Google Scholar] [CrossRef]
- Arsić, A.; Vučić, V.; Tepšić, J.; Mazić, S.; Djelić, M.; Glibetić, M. Altered Plasma and Erythrocyte Phospholipid Fatty Acid Profile in Elite Female Water Polo and Football Players. Appl. Physiol. Nutr. Metab. 2012, 37, 40–47. [Google Scholar] [CrossRef] [PubMed]
- Armstrong, A.; Anzalone, A.J.; Pethick, W.; Murray, H.; Dahlquist, D.T.; Askow, A.T.; Heileson, J.L.; Hillyer, L.M.; Ma, D.W.L.; Oliver, J.M. An Evaluation of Omega-3 Status and Intake in Canadian Elite Rugby 7s Players. Nutrients 2021, 13, 3777. [Google Scholar] [CrossRef] [PubMed]
- Hooks, M.P.; Madigan, S.M.; Woodside, J.V.; Nugent, A.P. Dietary Intake, Biological Status, and Barriers towards Omega-3 Intake in Elite Level (Tier 4), Female Athletes: Pilot Study. Nutrients 2023, 15, 2821. [Google Scholar] [CrossRef] [PubMed]
- Peña, N.; Amézaga, J.; Marrugat, G.; Landaluce, A.; Viar, T.; Arce, J.; Larruskain, J.; Lekue, J.; Ferreri, C.; Ordovás, J.M.; et al. Competitive Season Effects on Polyunsaturated Fatty Acid Content in Erythrocyte Membranes of Female Football Players. J. Int. Soc. Sports Nutr. 2023, 20, 2245386. [Google Scholar] [CrossRef] [PubMed]
- Stöggl, T.L.; Sperlich, B. The Training Intensity Distribution among Well-Trained and Elite Endurance Athletes. Front. Physiol. 2015, 6, 295. [Google Scholar] [CrossRef] [PubMed]
- Marini, M.; Abruzzo, P.M.; Bolotta, A.; Veicsteinas, A.; Ferreri, C. Aerobic Training Affects Fatty Acid Composition of Erythrocyte Membranes. Lipids Health Dis. 2011, 10, 188. [Google Scholar] [CrossRef] [PubMed]
- Kamada, T.; Tokuda, S.; Aozaki, S.I.; Otsuji, S. Higher Levels of Erythrocyte Membrane Fluidity in Sprinters and Long-Distance Runners. J. Appl. Physiol. 1993, 74, 354–358. [Google Scholar] [CrossRef] [PubMed]
- Maynar, M.; Llerena, F.; Grijota, F.J.; Alves, J.; Robles, M.C.; Bartolomé, I.; Muñoz, D. Serum Concentration of Several Trace Metals and Physical Training. J. Int. Soc. Sports Nutr. 2017, 14, 19. [Google Scholar] [CrossRef] [PubMed]
- Moreiras, O.; Carbajal, A.; Cabrera, L.; Cuadrado, C. Tablas de Composicion de Alimentos: Guia de Prácticas; Pirámide: Madrid, Spain, 2016; ISBN 978-84-368-3623-3. [Google Scholar]
- Alves, J.; Barrientos, G.; Toro, V.; Sánchez, E.; Muñoz, D.; Maynar, M. Changes in Anthropometric and Performance Parameters in High-Level Endurance Athletes during a Sports Season. Int. J. Environ. Res. Public. Health 2021, 18, 2782. [Google Scholar] [CrossRef]
- Stewart, A.; Marfell-Jones, M.; Olds, T.; de Ridder, H. International Society for Advancement of Kinanthropometry. In International Standards for Anthropometric Assessment; International Society for the Advancement of Kinanthropometry: Lower Hutt, New Zealand, 2011; ISBN 0620362073/9780620362078. [Google Scholar]
- Muñoz Marín, D.; Barrientos, G.; Alves, J.; Grijota, F.J.; Robles, M.C.; May Nar, M. Oxidative Stress, Lipid Peroxidation Indexes and Antioxidant Vitamins in Long and Middle Distance Athletes during a Sport Season. J. Sports Med. Phys. Fit. 2018, 58, 1713–1719. [Google Scholar] [CrossRef]
- Lepage, G.; Roy, C.C. Direct Transesterification of All Classes of Lipids in a One-Step Reaction. J. Lipid Res. 1986, 27, 114–120. [Google Scholar] [CrossRef] [PubMed]
- Nespolo, M. Free Radicals in Biology and Medicine. Fifth Edition. Oxford University Press, 2015. Pp 905(Paperback, ISBN 9780198717485), (Hardback, ISBN 9780198717478). Acta Crystallogr. D Struct. Biol. 2017, 73, 384–385. [Google Scholar] [CrossRef]
- Tomczak, M.; Tomczak, E. The Need to Report Effect Size Estimates Revisited. An Overview of Some Recommended Measures of Effect Size; Akademia Wychowania Fizycznego w Poznaniu: Poznań, Poland, 2014; Volume 1. [Google Scholar]
- Cohen, J. Statistical Power Analysis for the Behavioral Sciences; Routledge Academic: New York, NY, USA, 1988. [Google Scholar]
- Tepsic, J.; Vucic, V.; Arsic, A.; Mazic, S.; Djelic, M.; Glibetic, M. Unfavourable Plasma and Erythrocyte Phospholipid Fatty Acid Profile in Elite Amateur Boxers. Eur. J. Sport. Sci. 2013, 13, 414–421. [Google Scholar] [CrossRef] [PubMed]
- Riediger, N.D.; Othman, R.A.; Suh, M.; Moghadasian, M.H. A Systemic Review of the Roles of N-3 Fatty Acids in Health and Disease. J. Am. Diet. Assoc. 2009, 109, 668–679. [Google Scholar] [CrossRef]
- Shei, R.J.; Lindley, M.R.; Mickleborough, T.D. Omega-3 Polyunsaturated Fatty Acids in the Optimization of Physical Performance. Mil. Med. 2014, 179, 144–156. [Google Scholar] [CrossRef] [PubMed]
- Kawabata, T.; Hirota, S.; Hirayama, T.; Adachi, N.; Kaneko, Y.; Iwama, N.; Kamachi, K.; Araki, E.; Kawashima, H.; Kiso, Y. Associations between Dietary N-6 and n-3 Fatty Acids and Arachidonic Acid Compositions in Plasma and Erythrocytes in Young and Elderly Japanese Volunteers. Lipids Health Dis. 2011, 10, 138. [Google Scholar] [CrossRef] [PubMed]
- Gómez-Donoso, C.; Sánchez-Villegas, A.; Martínez-González, M.A.; Gea, A.; Mendonça, R.D.D.; Lahortiga-Ramos, F.; Bes-Rastrollo, M. Ultra-Processed Food Consumption and the Incidence of Depression in a Mediterranean Cohort: The SUN Project. Eur. J. Nutr. 2020, 59, 1093–1103. [Google Scholar] [CrossRef] [PubMed]
- Gammone, M.A.; Riccioni, G.; Parrinello, G.; D’orazio, N. Omega-3 Polyunsaturated Fatty Acids: Benefits and Endpoints in Sport. Nutrients 2019, 11, 46. [Google Scholar] [CrossRef] [PubMed]
- Davinelli, S.; Intrieri, M.; Corbi, G.; Scapagnini, G. Metabolic Indices of Polyunsaturated Fatty Acids: Current Evidence, Research Controversies, and Clinical Utility. Crit. Rev. Food Sci. Nutr. 2021, 61, 259–274. [Google Scholar] [CrossRef]
- Ayee, M.A.A.; Bunker, B.C.; De Groot, J.L. Membrane Modulatory Effects of Omega-3 Fatty Acids: Analysis of Molecular Level Interactions, 1st ed.; Elsevier Inc.: Amsterdam, The Netherlands, 2020; Volume 86, ISBN 9780128210215. [Google Scholar]
- Von Schacky, C.; Kemper, M.; Haslbauer, R.; Halle, M. Low Omega-3 Index in 106 German Elite Winter Endurance Athletes: A Pilot Study. Int. J. Sport. Nutr. Exerc. Metab. 2014, 24, 559–564. [Google Scholar] [CrossRef]
- Thielecke, F.; Blannin, A. Omega-3 Fatty Acids for Sport Performance—Are They Equally Beneficial for Athletes and Amateurs? A Narrative Review. Nutrients 2020, 12, 3712. [Google Scholar] [CrossRef] [PubMed]
- Allaire, J.; Couture, P.; Leclerc, M.; Charest, A.; Marin, J.; Lépine, M.C.; Talbot, D.; Tchernof, A.; Lamarche, B. A Randomized, Crossover, Head-to-Head Comparison of Eicosapentaenoic Acid and Docosahexaenoic Acid Supplementation to Reduce Inflammation Markers in Men and Women: The Comparing EPA to DHA (ComparED) Study. Am. J. Clin. Nutr. 2016, 104, 280–287. [Google Scholar] [CrossRef] [PubMed]
- Dalli, J.; Colas, R.A.; Serhan, C.N. Novel N-3 Immunoresolvents: Structures and Actions. Sci. Rep. 2013, 3, 1940. [Google Scholar] [CrossRef] [PubMed]
- Ervik, K.; Reinertsen, A.F.; Koenis, D.S.; Dalli, J.; Hansen, T.V. Stereoselective Synthesis, Pro-Resolution, and Anti-Inflammatory Actions of RvD5 n-3 DPA. J. Nat. Prod. 2023, 86, 2546–2553. [Google Scholar] [CrossRef] [PubMed]
- Ney, J.G.; Koury, J.C.; Azeredo, V.B.; Casimiro-Lopes, G.; Trugo, N.M.F.; Torres, A.G. Associations of N-6 and n-3 Polyunsaturated Fatty Acids and Tocopherols with Proxies of Membrane Stability and Subcutaneous Fat Sites in Male Elite Swimmers. Nutr. Res. 2009, 29, 623–630. [Google Scholar] [CrossRef] [PubMed]
- Andersson, A.; Sjödin, A.; Olsson, R.; Vessby, B.; Hedman, A.; Olsson, R.; Vessby, B. Fatty Acid Profile of Skeletal Muscle Phospholipids in Trained and Untrained Young Men. Am. J. Physiol. Endocrinol. Metab. 1998, 279, 744–751. [Google Scholar] [CrossRef]
- Thomas, T.R.; Londeree, B.R.; Gerhardt, K.O.; Gehrke, C.W. Fatty Acid Profile and Cholesterol in Skeletal Muscle of Trained and Untrained Men. J. Appl. Physiol. Respir. Environ. Exerc. Physiol. 1977, 43, 709–713. [Google Scholar] [CrossRef] [PubMed]
- Høstmark, A.T.; Haug, A. Percentages of Oleic Acid and Arachidonic Acid Are Inversely Related in Phospholipids of Human Sera. Lipids Health Dis. 2013, 12, 106. [Google Scholar] [CrossRef]
- Pamplona, R.; Portero-Otin, M.; Sanz, A.; Requena, J.; Barja, G. Modification of the Longevity-Related Degree of Fatty Acid Unsaturation Modulates Oxidative Damage to Proteins and Mitochondrial DNA in Liver and Brain. Exp. Gerontol. 2004, 39, 725–733. [Google Scholar] [CrossRef]
- Yasui, Y.; Hosokawa, M.; Kohno, H.; Tanaka, T.; Miyashita, K. Growth inhibition and apoptosis induction by all-trans-conjugated linolenic acids on human colon cancer cells. Anticancer Res. 2006, 26, 1855–1860. [Google Scholar] [PubMed]
- Carta, G.; Murru, E.; Banni, S.; Manca, C. Palmitic Acid: Physiological Role, Metabolism and Nutritional Implications. Front. Physiol. 2017, 8, 902. [Google Scholar] [CrossRef] [PubMed]
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).