
Marine Bioactives and Their Application in the Food Industry: A Review




Abstract
:1. Introduction
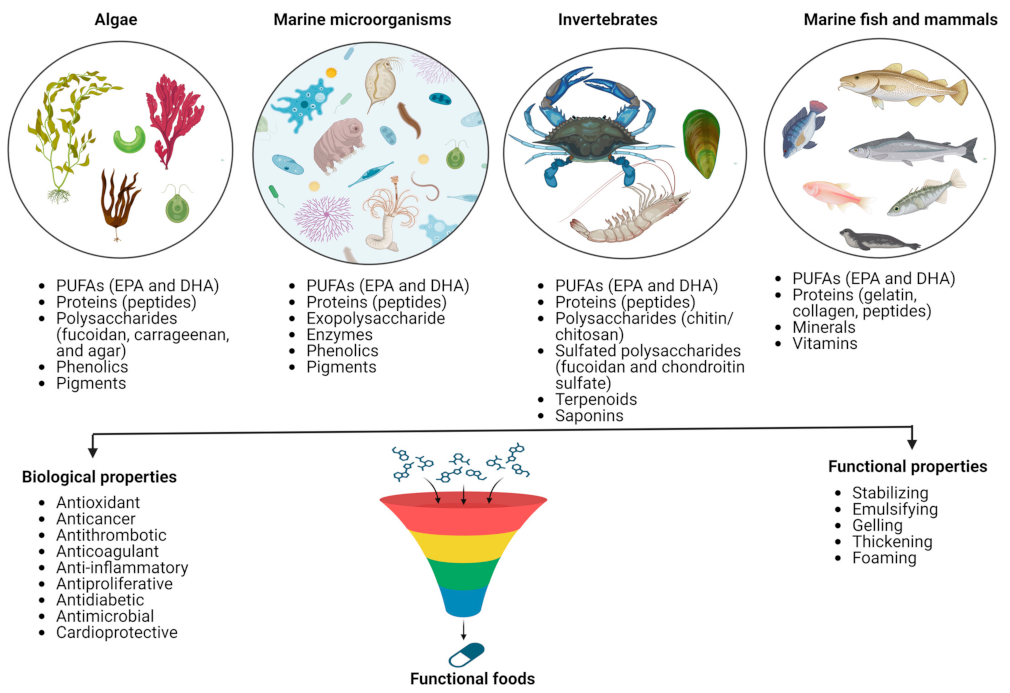
2. Marine Sources of Bioactive Molecules
2.1. Marine Microorganisms
2.1.1. Bioactive Polysaccharides from Marine Microbiota
Marine Bacteria
Marine Cyanobacteria
2.1.2. Proteins and Bioactive Peptides from Microbiota

{kind=link}
{kind=link}
{kind=link}
| Type of Enzymes | Responsible Microorganism | Applications | References |
|---|---|---|---|
| Protease | Aureobasidium pullulans, Bacillus mojavensis, Psychrobacter, Clostridium, Rhizopus, Penicillium, and Aspergillus | Pharmaceutical (digestive drugs and anti-inflammatory drugs), detergent, and leather industries | [34,36] |
| Lipase | Penicillium Oxalicum, Aspergillus flavus, Moraxella, Candida intermedia, Pichia guilliermondii, Lodderomyces elongisporus, Candida parapsilosis, Candia rugosa, Candida quercitrusa, and Yarrowia lipolytica | Detergents, cosmetic production, paper production, and food flavoring | [33,34,37] |
| Chitinase and chitosanase | Aspergillus, Penicillium, Rhizopus Myxobacter, Sporocytophaga, Bacillus, Enterobacter, Flavobacterium, Arthrobacter, Vibrio parahaemolyticus, S Vibrio fluvialis, Vibrio mimicus, Vibrio alginolyticus, Listonella anguillarum, Klebsiella, Pseudomonas, Serratia, Chromobacterium, Clostridium, and Aeromonas hydrophila | Pharmaceutical, functional food, and cosmetic sectors | [33,38,39] |
| Agarases | Pseudomonas galatica, Cytophaga, Alteromonas, Bacillus, Vibrio, Pseudoalteromonas, Streptomyces, and Pseudomonas | Food industry (beverages, bread, cookies, and some low-calorie food) | [33,34,40] |
| Cellulase | Cytophaga, Cellulomonas, Vibrio, and Clostridium, Nocardia, Streptomyces, Trichoderma, Aspergillus, Fusarium, Chaetomium, Phoma, Sporotrichum, and Penicillium | Cotton and linen product processing, bio-textile auxiliaries, and bio-fertilizer processing | [33,34,38,41] |
| Xylanases | Aspergillus niger and Penicillium chrysogenum | Paper and pulp industries and beverage clarification | [33,42] |
2.1.3. Fatty Acids from Microbiota
2.2. Marine Algae
2.2.1. Macroalgae
Macroalgae Polysaccharides
Macroalgae Proteins, Peptides, and Amino Acids
Macroalgae Lipids and Fatty Acids
2.2.2. Microalgae
2.2.3. Other Biomolecules from Marine Algae
2.3. Marine Invertebrates
2.3.1. Sponges
2.3.2. Mollusk
2.3.3. Echinoderms
2.3.4. Crustaceans
2.4. Marine Fish
2.4.1. Fish Lipids
2.4.2. Fish Proteins
Development of Fish Protein Hydrolysates/Peptides
2.4.3. Other Compounds/Elements Derived from Marine Fish
3. Marine Bioactive Compounds and the Food Industry
3.1. Bioactive Marine Polysaccharides
3.2. Bioactive Marine Peptides
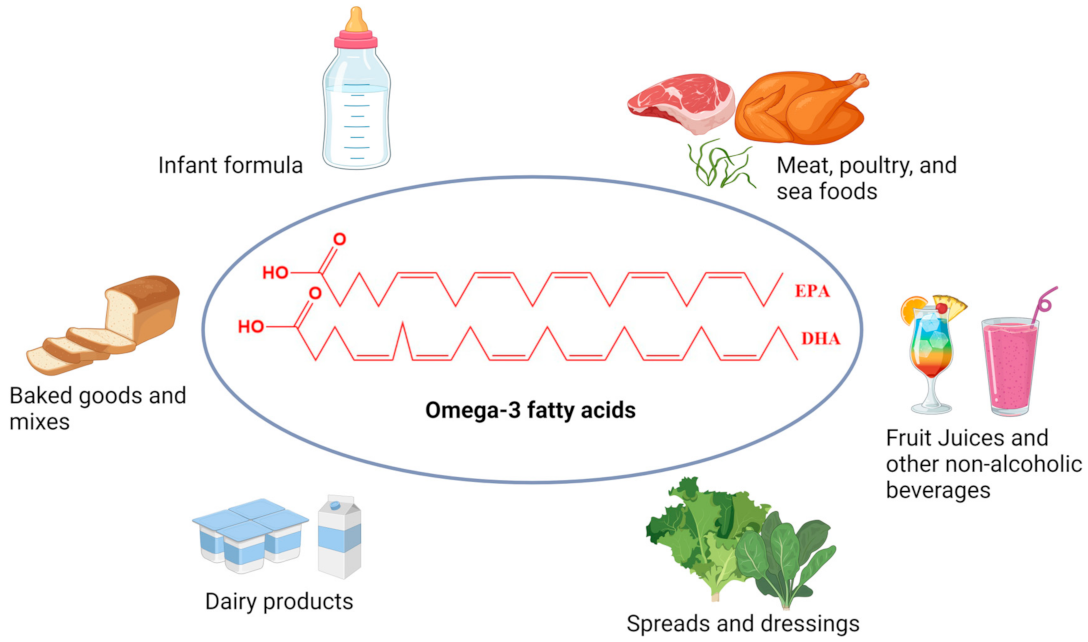
3.3. Bioactive Marine Polyunsaturated Fatty Acids
4. Challenges and Future Trends
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Shahidi, F.; Ambigaipalan, P. Novel functional food ingredients from marine sources. Curr. Opin. Food Sci. 2015, 2, 123–129. [Google Scholar] [CrossRef]
- Huang, I.; Pietrasiak, N.; Gobler, C.J.; Johansen, J.R.; Burkholder, J.M.; D’Antonio, S.; Zimba, P.V. Diversity of bioactive compound content across 71 genera of marine, freshwater, and terrestrial cyanobacteria. Harmful Algae 2021, 109, 102116. [Google Scholar] [CrossRef] [PubMed]
- Hosseini, S.F.; Rezaei, M.; McClements, D.J. Bioactive functional ingredients from aquatic origin: A review of recent progress in marine-derived nutraceuticals. Crit. Rev. Food Sci. Nutr. 2020, 62, 1242–1269. [Google Scholar] [CrossRef] [PubMed]
- Suleria, H.A.R.; Osborne, S.A.; Masci, P.P.; Gobé, G.C. Marine-based nutraceuticals: An innovative trend in the food and supplement industries. Mar. Drugs 2015, 13, 6336–6351. [Google Scholar] [CrossRef]
- Jo, C.; Khan, F.F.; Khan, M.I.; Iqbal, J. Marine bioactive peptides: Types, structures, and physiological functions. Food Rev. Int. 2016, 33, 44–61. [Google Scholar] [CrossRef]
- Ali, A.; Wei, S.; Liu, Z.; Fan, X.; Sun, Q.; Xia, Q.; Liu, S.; Hao, J.; Deng, C. Non-thermal processing technologies for the recovery of bioactive compounds from marine by-products. LWT Food Sci. Technol. 2021, 147, 111549. [Google Scholar] [CrossRef]
- Hossain, A.; Dave, D.; Shahidi, F. Northern sea cucumber (Cucumaria frondosa): A potential candidate for functional food, nutraceutical, and pharmaceutical sector. Mar. Drugs 2020, 18, 274. [Google Scholar] [CrossRef] [PubMed]
- Ghosh, S.; Sarkar, T.; Pati, S.; Kari, Z.A.; Edinur, H.A.; Chakraborty, R. Novel bioactive compounds from marine sources as a tool for functional food development. Front. Mar. Sci. 2022, 9, 832957. [Google Scholar] [CrossRef]
- Lobine, D.; Rengasamy, K.R.R.; Mahomoodally, M.F. Functional foods and bioactive ingredients harnessed from the ocean: Current status and future perspectives. Crit. Rev. Food Sci. Nutr. 2021, 62, 5794–5823. [Google Scholar] [CrossRef]
- Shahidi, F.; Santhiravel, S. Novel marine bioactives: Application in functional foods, nutraceuticals, and pharmaceuticals. J. Food Bioact. 2022, 19, 4–96. [Google Scholar] [CrossRef]
- Ameen, F.; AlNadhari, S.; Al-Homaidan, A.A. Marine microorganisms as an untapped source of bioactive compounds. Saudi J. Biol. Sci. 2021, 28, 224–231. [Google Scholar] [CrossRef]
- Satheesh, S.; Ba-akdah, M.A.; Al-Sofyani, A.A. Natural antifouling compound production by microbes associated with marine microorganisms—A review. Electron. J. Biotechnol. 2016, 21, 26–35. [Google Scholar] [CrossRef]
- Van Doan, H.; Prakash, P.S.; Hoseinifar, S.H.; Ringø, E.; El-Haroun, E.; Faggio, C.; Olsen, R.E.; Tran, H.Q.; Stejskal, V.; Abdel-Latif, H.M.R.; et al. Marine-derived products as functional feed additives in aquaculture: A review. Aquac. Rep. 2023, 31, 101679. [Google Scholar] [CrossRef]
- Ibrahim, S.R.M.; Mohamed, S.G.A.; Alsaadi, B.H.; Althubyani, M.M.; Awari, Z.I.; Hussein, H.S.; Aljohani, A.A.; Albasri, J.F.; Faraj, S.A.; Mohamed, G.A. Secondary metabolites, biological activities, and industrial and biotechnological importance of Aspergillus sydowii. Mar. Drugs 2023, 21, 441. [Google Scholar] [CrossRef]
- Casillo, A.; Lanzetta, R.; Parrilli, M.; Corsaro, M.M. Exopolysaccharides from marine and marine extremophilic bacteria: Structures, properties, ecological roles and applications. Mar. Drugs 2018, 16, 1–34. [Google Scholar] [CrossRef]
- Qi, M.; Zheng, C.; Wu, W.; Yu, G.; Wang, P. Exopolysaccharides from marine microbes: Source, structure and application. Mar. Drugs 2022, 20, 512. [Google Scholar] [CrossRef]
- Ju, H.; Yu, C.J.; Liu, W.; Li, H.-H.; Fu, Z.; Wu, Y.; Gong, P.-X.; Li, H. Polysaccharides from marine resources exhibit great potential in the treatment of tumor: A review. Carbohydr. Polym. Technol. Appl. 2023, 5, 100308. [Google Scholar] [CrossRef]
- Sardari, R.R.R.; Kulcinskaja, E.; Ron, E.Y.C.; Björnsdóttir, S.H.; Friðjónsson, Ó.H.; Hreggviðsson, G.Ó.; Karlsson, E.N. Evaluation of the production of exopolysaccharides by two strains of the thermophilic bacterium Rhodothermus marinus. Carbohydr. Polym. 2017, 156, 1–8. [Google Scholar] [CrossRef]
- Majee, S.B.; Avlani, D.; Biswas, G.R. Pharmacological, pharmaceutical, cosmetic and diagnostic applications of sulfated polysaccharides from marine algae and bacteria. Afr. J. Pharm. Pharmacol. 2017, 11, 68–77. [Google Scholar]
- Waghmode, S.; Suryavanshi, M.V.; Sharma, D.; Satpute, S.K. Planococcus species—An imminent resource to explore biosurfactant and bioactive metabolites for industrial applications. Front. Bioeng. Biotechnol. 2020, 8, 996. [Google Scholar] [CrossRef]
- Bramhachari, P.V.; Nagaraju, G.P. Extracellular polysaccharide production by bacteria as a mechanism of toxic heavy metal biosorption and biosequestration in the marine environment. In Marine Pollution and Microbial Remediation; Springer: Singapore, 2016; pp. 67–85. [Google Scholar]
- Nadzir, M.M.; Nurhayati, R.W.; Idris, F.N.; Nguyen, M.H. Biomedical applications of bacterial exopolysaccharides: A review. Polymers 2021, 13, 530. [Google Scholar] [CrossRef]
- Dewapriya, P.; Kim, S. Marine microorganisms: An emerging avenue in modern nutraceuticals and functional foods. Food Res. Int. 2014, 56, 115–125. [Google Scholar] [CrossRef]
- Zhang, J.; Xiao, Y.; Wang, H.; Zhang, H.; Zhao, J.; Liu, W. Lactic acid bacteria-derived exopolysaccharide: Formation, immunomodulatory ability, health effects, and structure-function relationship. Microbiol. Res. 2023, 274, 127432. [Google Scholar] [CrossRef] [PubMed]
- Jagannathan, S.V.; Manemann, E.M.; Rowe, S.E.; Callender, M.C.; Soto, W. Marine actinomycetes, new sources of biotechnological products. Mar. Drugs 2021, 19, 365. [Google Scholar] [CrossRef] [PubMed]
- El-Seedi, H.R.; El-Mallah, M.F.; Yosri, N.; Alajlani, M.; Zhao, C.; Mehmood, M.A.; Du, M.; Ullah, H.; Daglia, M.; Guo, Z.; et al. Review of marine Cyanobacteria and the aspects related to their roles: Chemical, biological properties, nitrogen fixation and climate change. Mar. Drugs 2023, 21, 439. [Google Scholar] [CrossRef] [PubMed]
- Franco-Morgado, M.; Amador-Espejo, G.G.; Pérez-Cortes, M.; Gutiérrez-Uribe, J.A. Microalgae and cyanobacteria polysaccharides: Important link for nutrient recycling and revalorization of agro-industrial wastewater. Appl. Food Res. 2023, 3, 100296. [Google Scholar] [CrossRef]
- Singh, S.R.; Kant, C.; Yadav, R.K.; Reddy, Y.P.; Abraham, G. Cyanobacterial exopolysaccharides: Composition, biosynthesis, and biotechnological applications. In Cyanobacteria: From Basic Science to Applications; Academic Press: Cambridge, MA, USA, 2019; pp. 347–358. [Google Scholar]
- Akan, G.; Oner, E.T. Gel properties of microbial polysaccharides. In Polysaccharides of Microbial Origin; Oliveira, J., Radhouani, H., Reis, R.L., Eds.; Springer: Cham, Switzerland, 2021. [Google Scholar]
- Laroche, C. Exopolysaccharides from microalgae and cyanobacteria: Diversity of strains, production strategies, and applications. Mar. Drugs 2022, 20, 336. [Google Scholar] [CrossRef]
- Cunha, S.A.; Pintado, M. Bioactive peptides derived from marine sources: Biological and functional properties. Trends Food Sci. Technol. 2022, 119, 348–370. [Google Scholar] [CrossRef]
- Lee, S.Y.; Hur, S.J. Antihypertensive peptides from animal products, marine organisms, and plants. Food Chem. 2017, 228, 506–517. [Google Scholar] [CrossRef]
- Rao, T.E.; Imchen, M.; Kumavath, R. Marine Enzymes. Marine enzymes: Production and applications for human health. In Advances in Food and Nutrition Research; Academic Press: Cambridge, MA, USA, 2017; pp. 149–163. [Google Scholar]
- Parte, S.; Sirisha, V.L.; D’Souza, J.S. Biotechnological applications of marine enzymes from algae, bacteria, fungi, and sponges. In Advances in Food and Nutrition Research; Academic Press: Cambridge, MA, USA, 2017; pp. 75–106. [Google Scholar]
- Hayes, M.; Aluko, R.E.; Aurino, E.; Mora, L. Generation of bioactive peptides from Porphyridium sp. and assessment of their potential for use in the prevention of hypertension, inflammation and pain. Mar. Drugs 2023, 21, 422. [Google Scholar] [CrossRef]
- Birolli, W.G.; Lima, R.N.; Porto, A.L.M. Applications of marine-derived microorganisms and their enzymes in biocatalysis and biotransformation, the underexplored potentials. Front. Microbiol. 2019, 10, 1453. [Google Scholar] [CrossRef] [PubMed]
- Byreddy, A.R.; Rao, N.M.; Barrow, C.J.; Puri, M. Tween 80 influences the production of intracellular lipase by Schizochytrium S31 in a stirred tank reactor. Process Biochem. 2017, 53, 30–35. [Google Scholar] [CrossRef]
- Bibi, F.; Faheem, M.; Azhar, E.I.; Yasir, M.; Alvi, S.; Kamal, M.A.; Ullah, I.; Naseer, M.I. Bacteria from marine sponges: A source of new drugs. Curr. Drug Metab. 2017, 18, 11–15. [Google Scholar] [CrossRef]
- Sivaperumal, P.; Kamala, K.; Rajaram, R. Bioremediation of industrial waste through enzyme producing marine microorganisms. In Advances in Food and Nutrition Research; Academic Press: Cambridge, MA, USA, 2017; pp. 165–179. [Google Scholar]
- Park, S.H.; Lee, C.-R.; Hong, S.-K. Implications of agar and agarase in industrial applications of sustainable marine biomass. Appl. Microbiol. Biotechnol. 2020, 104, 2815–2832. [Google Scholar] [CrossRef] [PubMed]
- John, J.A.; Samuel, M.S.; Govarthan, M.; Ethiraj, S.A. A comprehensive review on strategic study of cellulase producing marine actinobacteria for biofuel applications. Environ. Res. 2022, 214, 114018. [Google Scholar] [CrossRef]
- Qeshmi, F.I.; Homaei, A.; Fernandes, P.; Hemmati, R.; Dijkstra, B.W.; Khajeh, K. Xylanases from marine microorganisms: A brief overview on scope, sources, features and potential applications. Biochim. Biophys. Acta Proteins Proteom. 2020, 1868, 140312. [Google Scholar] [CrossRef] [PubMed]
- Shah, A.M.; Yang, W.; Mohamed, H.; Zhang, Y.; Song, Y. Microbes: A hidden treasure of polyunsaturated fatty acids. Front. Nutr. 2022, 9, 827837. [Google Scholar] [CrossRef]
- AlFadhly, N.K.Z.; Alhelfi, N.; Altemimi, A.B.; Verma, D.K.; Cacciola, F.; Narayanankutty, A. Trends and technological advancements in the possible food applications of Spirulina and their health benefits: A review. Molecules 2022, 27, 5584. [Google Scholar] [CrossRef]
- Remize, M.; Brunel, Y.; Silva, J.; Berthon, J.; Filaire, E. Microalgae n-3 PUFAs production and use in food and feed industries. Mar. Drugs 2021, 19, 113. [Google Scholar] [CrossRef]
- Bellou, S.; Triantaphyllidou, I.-E.; Aggeli, D.; Elazzazy, A.M.; Baeshen, M.N.; Aggelis, G. Microbial oils as food additives: Recent approaches for improving microbial oil production and its polyunsaturated fatty acid content. Curr. Opin. Biotechnol. 2016, 37, 24–35. [Google Scholar] [CrossRef]
- Burja, A.M.; Radianingtyas, H. Nutraceuticals and functional foods from marine microbes: An introduction to a diverse group of natural products isolated from marine macroalgae, microalgae, bacteria, fungi, and cyanobacteria. In Marine Nutraceuticals and Functional Foods; Barrow, C., Shahidi, F., Eds.; CRC Press: Boca Raton, FL, USA, 2007; pp. 367–403. [Google Scholar]
- Jovanović, S.; Dietrich, D.; Becker, J.; Kohlstedt, M.; Wittmann, C. Microbial production of polyunsaturated fatty acids—High-value ingredients for aquafeed, superfoods, and pharmaceuticals. Curr. Opin. Biotechnol. 2021, 69, 199–211. [Google Scholar] [CrossRef] [PubMed]
- Rioux, L.; Beaulieu, L.; Turgeon, S.L. Seaweeds: A traditional ingredients for new gastronomic sensation. Food Hydrocoll. 2017, 68, 255–265. [Google Scholar] [CrossRef]
- Pole, R.P.P.; Khan, M.F.; Wesley, S.G. Occurrence of 210Po in marine macroalgae inhabiting a coastal nuclear zone, southeast coast of India. J. Environ. Radioact. 2017, 169, 122–130. [Google Scholar] [CrossRef] [PubMed]
- Freitas, A.C.; Pereira, L.; Rodrigues, D.; Carvalho, A.P.; Panteleitchouk, T.S.L.; Gomes, A.; Duarte, A.C. Marine functional foods. In Handbook of Marine Biotechnology; Springer: Berlin/Heidelberg, Germany, 2015; pp. 969–994. [Google Scholar]
- Imchen, T.; Singh, K.S. Marine algae colorants: Antioxidant, anti-diabetic properties and applications in food industry. Algal Res. 2023, 69, 102898. [Google Scholar] [CrossRef]
- Ovando, C.A.; De Carvalho, J.C.; De Melo Pereira, G.V.; Jacques, P.; Soccol, V.T.; Soccol, C.R. Functional properties and health benefits of bioactive peptides derived from Spirulina: A review. Food Rev. Int. 2016, 34, 34–51. [Google Scholar] [CrossRef]
- Kratzer, R.; Murkovic, M. Food ingredients and nutraceuticals from microalgae: Main product classes and biotechnological production. Foods 2021, 10, 1626. [Google Scholar] [CrossRef]
- Senadheera, T.R.L.; Dave, D. Marine algae—Ocean based emerging biomass for functional food ingredients. J. Ocean Technol. 2021, 16, 1–10. [Google Scholar]
- Mensah, E.O.; Kanwugu, O.N.; Panda, P.K.; Adadi, P. Marine fucoidans: Structural, extraction, biological activities and their applications in the food industry. Food Hydrocoll. 2023, 142, 108784. [Google Scholar] [CrossRef]
- Harsha, M.E.; Madhusudan, S.; Baskaran, R. The sea lettuce Ulva sensu lato: Future food with health-promoting bioactives. Algal Res. 2023, 71, 103069. [Google Scholar]
- Salleh, S.N.; Fairus, A.A.H.; Zahary, M.N.; Raj, N.B.; Jalil, A.M.M. Unravelling the effects of soluble dietary fibre supplementation on energy intake and perceived satiety in healthy adults: Evidence from systematic review and meta-analysis of randomised-controlled trials. Foods 2019, 8, 1–22. [Google Scholar] [CrossRef]
- El-Beltagi, H.S.; Mohamed, A.A.; Mohamed, H.I.; Ramadan, K.M.A.; Barqawi, A.A.; Mansour, A.T. Phytochemical and potential properties of seaweeds and their recent applications: A review. Mar. Drugs 2022, 20, 342. [Google Scholar] [CrossRef] [PubMed]
- Hossain, A.; Dave, D.; Shahidi, F. Antioxidant potential of sea cucumbers and their beneficial effects on human health. Mar. Drugs 2022, 20, 521. [Google Scholar] [CrossRef] [PubMed]
- Kadam, S.U.; O’Donnell, C.P.; Dilip, K.; Hossain, M.B.; Burgess, C.M.; Walsh, D.; Tiwari, B.K. Laminarin from Irish brown seaweeds Ascophyllum nodosum and Laminaria hyperborea: Ultrasound assisted extraction, characterization and bioactivity. Mar. Drugs 2015, 13, 4270–4280. [Google Scholar] [CrossRef] [PubMed]
- Cardoso, M.J.; Costa, R.R.; Mano, J.F. Marine origin polysaccharides in drug delivery systems. Mar. Drugs 2016, 14, 34. [Google Scholar] [CrossRef]
- Cunha, L.; Grenha, A. Sulfated seaweed polysaccharides as multifunctional materials in drug delivery applications. Mar. Drugs 2016, 14, 42. [Google Scholar] [CrossRef]
- Shahidi, F.; Rahman, M.J. Bioactives in seaweeds, algae, and fungi and their role in health promotion. J. Food Bioact. 2018, 2, 58–81. [Google Scholar] [CrossRef]
- Kulshreshtha, G.; Burlot, A.-S.; Marty, C.; Critchley, A.T.; Hafting, J.T.; Bedoux, G.; Bourgougnon, N.; Prithiviraj, B. Enzyme assisted extraction of bioactive material from Chondrus crispus and Codium fragile and its effect on Herpes simplex virus (HSV-1). Mar. Drugs 2015, 13, 558–580. [Google Scholar] [CrossRef]
- Panggabean, J.A.; Adiguna, S.P.; Rahmawati, S.I.; Ahmadi, P.; Zainuddin, E.N.; Bayu, A.; Putra, M.Y. Antiviral activities of algal-based sulfated polysaccharides. Molecules 2022, 27, 1178. [Google Scholar] [CrossRef]
- Van Weelden, G.; Bobinski, M.; Okła, K.; Van Weelden, W.J.; Romano, A.; Pijnenborg, J.M.A. Fucoidan structure and activity in relation to anti-cancer mechanisms. Mar. Drugs 2019, 17, 32. [Google Scholar] [CrossRef]
- Lin, Y.; Qi, X.; Liu, H.-J.; Xue, K.; Xu, S.; Tian, Z. The anti-cancer effects of fucoidan: A review of both in vivo and in vitro investigations. Cancer Cell Int. 2020, 20, 154. [Google Scholar] [CrossRef]
- Riccio, G.; Lauritano, C. Microalgae with immunomodulatory activities. Mar. Drugs 2019, 18, 2. [Google Scholar] [CrossRef] [PubMed]
- Thiviya, P.; Gamage, A.; Gama-Arachchige, N.S.; Merah, O.; Madhujith, T. Seaweeds as a source of functional proteins. Phycology 2022, 2, 216–243. [Google Scholar] [CrossRef]
- Ganesan, A.R.; Tiwari, U.; Rajauria, G. Seaweed nutraceuticals and their therapeutic role in disease prevention. Food Sci. Hum. Wellness 2019, 8, 252–263. [Google Scholar] [CrossRef]
- Quitério, E.; Soares, C.; Ferraz, R.; Delerue-Matos, C.; Grosso, C. Marine health-promoting compounds: Recent trends for their characterization and human applications. Foods 2021, 10, 3100. [Google Scholar] [CrossRef] [PubMed]
- Saide, A.; Martínez, K.A.; Ianora, A.; Lauritano, C. Unlocking the health potential of microalgae as sustainable sources of bioactive compounds. Int. J. Mol. Sci. 2021, 22, 4383. [Google Scholar] [CrossRef] [PubMed]
- Dagnino-Leone, J.; Figueroa, C.P.; Castañeda, M.L.; Youlton, A.D.; Vallejos-Almirall, A.; Agurto-Muñoz, A.; Pérez, J.P.; Agurto-Muñoz, C. Phycobiliproteins: Structural aspects, functional characteristics, and biotechnological perspectives. Comput. Struct. Biotechnol. J. 2022, 20, 1506–1527. [Google Scholar] [CrossRef]
- De Arruda, M.C.S.; Da Silva, M.R.O.B.; Cavalcanti, V.L.R.; Brandao, R.M.P.C.; De Araújo Viana Marques, D.; De Lima, L.R.A.; Porto, A.L.F.; Bezerra, R.P. Antitumor lectins from algae: A systematic review. Algal Res. 2023, 70, 102962. [Google Scholar] [CrossRef]
- Maliki, I.M.; Misson, M.; Teoh, P.L.; Rodrigues, K.F.; Yong, W.T.L. Production of lectins from marine algae: Current status, challenges, and opportunities for non-destructive extraction. Mar. Drugs 2022, 20, 102. [Google Scholar] [CrossRef]
- Bleakley, S.; Hayes, M. Algal proteins: Extraction, application, and challenges concerning production. Foods 2017, 6, 33. [Google Scholar] [CrossRef]
- Harwood, J.L. Algae: Critical sources of very long-chain polyunsaturated fatty acids. Biomolecules 2019, 9, 708. [Google Scholar] [CrossRef]
- Pradhan, B.; Nayak, R.; Patra, S.; Jit, B.P.; Ragusa, A.; Jena, M. Bioactive metabolites from marine algae as potent pharmacophores against oxidative stress-associated human diseases: A comprehensive review. Molecules 2020, 26, 37. [Google Scholar] [CrossRef] [PubMed]
- Schmid, M.; Guihéneuf, F.; Stengel, D.B. Evaluation of food grade solvents for lipid extraction and impact of storage temperature on fatty acid composition of edible seaweeds Laminaria digitata (Phaeophyceae) and Palmaria palmata (Rhodophyta). Food Chem. 2016, 208, 161–168. [Google Scholar] [CrossRef] [PubMed]
- Cheng, P.; Li, Y.; Wang, C.; Guo, J.; Zhou, C.; Zhang, R.; Ma, Y.; Ma, X.; Wang, L.; Cheng, Y.; et al. Integrated marine microalgae biorefineries for improved bioactive compounds: A review. Sci. Total Environ. 2022, 817, 152895. [Google Scholar] [CrossRef] [PubMed]
- Zhou, L.; Li, K.; Duan, X.; Hill, D.R.; Barrow, C.J.; Dunshea, F.R.; Martin, G.M.; Suleria, H.A.R. Bioactive compounds in microalgae and their potential health benefits. Food Biosci. 2022, 49, 101932. [Google Scholar] [CrossRef]
- Nezafatian, E.; Farhadian, O.; Yegdaneh, A.; Safavi, M.; Daneshvar, E.; Bhatnagar, A. Enhanced production of bioactive compounds from marine microalgae Tetraselmis tetrathele under salinity and light stresses: A two-stage cultivation strategy. Bioresour. Technol. 2023, 376, 128899. [Google Scholar] [CrossRef]
- Mekinić, I.G.; Šimat, V.; Rathod, N.B.; Hamed, I.; Čagalj, M. Algal carotenoids: Chemistry, sources, and application. Foods 2023, 12, 2768. [Google Scholar] [CrossRef]
- Hamed, I.; Özoğul, F.; Özoğul, Y.; Regenstein, J.M. Marine bioactive compounds and their health benefits: A review. Compr. Rev. Food Sci. Food Saf. 2015, 14, 446–465. [Google Scholar] [CrossRef]
- Martins, T.; Barros, A.; Rosa, E.; Antunes, L. Enhancing health benefits through chlorophylls and chlorophyll-rich agro-food: A comprehensive review. Molecules 2023, 28, 5344. [Google Scholar] [CrossRef]
- Catarino, M.D.; Amarante, S.J.; Mateus, N.; Silva, A.M.S.; Cardoso, S.M. Brown algae phlorotannins: A marine alternative to break the oxidative stress, inflammation and cancer network. Foods 2021, 10, 1478. [Google Scholar] [CrossRef]
- Circuncisão, A.R.; Catarino, M.D.; Cardoso, S.M.; Silva, A.M.S. Minerals from macroalgae origin: Health benefits and risks for consumers. Mar. Drugs 2018, 16, 400. [Google Scholar] [CrossRef]
- Das, D.; Arulkumar, A.; Paramasivam, S.; López-Santamarina, A.; Del Carmen Mondragón, A.; Miranda, J.M. Phytochemical constituents, antimicrobial properties and bioactivity of marine red seaweed (Kappaphycus alvarezii) and seagrass (Cymodocea serrulata). Foods 2023, 12, 2811. [Google Scholar] [CrossRef] [PubMed]
- Arora, N.; Philippidis, G.P. The prospects of algae-derived vitamins and their precursors for sustainable cosmeceuticals. Processes 2023, 11, 587. [Google Scholar] [CrossRef]
- Datta, D.; Talapatra, S.N.; Swarnakar, S. Bioactive compounds from marine invertebrates for potential medicines—An overview. Int. Lett. Nat. Sci. 2015, 34, 42–61. [Google Scholar] [CrossRef]
- Hossain, A.; Yeo, J.; Dave, D.; Shahidi, F. Phenolic compounds and antioxidant capacity of sea cucumber (Cucumaria frondosa) processing discards as affected by high-pressure processing (HPP). Antioxidants 2022, 11, 337. [Google Scholar] [CrossRef]
- Romano, G.; Almeida, M.; Coelho, A.V.; Cutignano, A.; Gonçalves, L.G.; Hansen, E.; Khnykin, D.; Mass, T.; Ramšak, A.; Rocha, M.; et al. Bioactive natural products and biomaterials from marine invertebrates: From basic research to innovative applications. Mar. Drugs 2022, 20, 219. [Google Scholar] [CrossRef]
- Liu, L.; Zheng, Y.-Y.; Shao, C.; Wang, C. Metabolites from marine invertebrates and their symbiotic microorganisms: Molecular diversity discovery, mining, and application. Mar. Life Sci. Technol. 2019, 1, 60–94. [Google Scholar] [CrossRef]
- Ko, S.; Jang, J.Y.; Ye, B.-R.; Kim, M.-S.; Choi, I.; Park, W.; Heo, S.; Jung, W. Purification and molecular docking study of angiotensin I-converting enzyme (ACE) inhibitory peptides from hydrolysates of marine sponge Stylotella aurantium. Process Biochem. 2017, 54, 180–187. [Google Scholar] [CrossRef]
- Pailee, P.; Mahidol, C.; Ruchirawat, S.; Prachyawarakorn, V. Sterols from Thai marine sponge Petrosia (Strongylophora) sp. and their cytotoxicity. Mar. Drugs 2017, 15, 54. [Google Scholar] [CrossRef]
- Krishnan, K.; Keerthi, T.R. Comparative evaluation of bioactivities of two marine sponges, Zygomycale parishii and Callyspongia diffusa from Southwest coast of India. Br. J. Pharm. Res. 2016, 11, 1–10. [Google Scholar] [CrossRef]
- Varijakzhan, D.; Loh, J.-Y.; Yap, W.-S.; Yusoff, K.; Seboussi, R.; Lim, S.H.E.; Lai, K.-S.; Chong, C.-M. Bioactive compounds from marine sponges: Fundamentals and applications. Mar. Drugs 2021, 19, 246. [Google Scholar] [CrossRef]
- Xu, J.; Wang, M.; Liu, Z.; Zhang, W.; Ma, J.; Li, G.; Li, P. Terpenoids from the sponge Sarcotragus sp. collected in the South China Sea. J. Nat. Prod. 2023, 86, 330–339. [Google Scholar] [CrossRef] [PubMed]
- Nicácio, K.J.; Ióca, L.P.; Fróes, A.M.; Leomil, L.; Appolinario, L.; Thompson, C.C.; Thompson, F.L.; Ferreira, A.G.; Williams, D.E.; Andersen, R.J.; et al. Cultures of the marine bacterium Pseudovibrio denitrificans Ab134 produce bromotyrosine-derived alkaloids previously only isolated from marine sponges. J. Nat. Prod. 2017, 80, 235–240. [Google Scholar] [CrossRef] [PubMed]
- Jayaprakash, R.; Perera, C.O. Partial purification and characterization of bioactive peptides from cooked New Zealand green-lipped mussel (Perna canaliculus) protein hydrolyzates. Foods 2020, 9, 879. [Google Scholar] [CrossRef] [PubMed]
- Naik, A.S.; Hayes, M. Bioprocessing of mussel by-products for value added ingredients. Trends Food Sci. Technol. 2019, 92, 111–121. [Google Scholar] [CrossRef]
- Stonik, V.A.; Kicha, A.A.; Malyarenko, T.V.; Ivanchina, N.V. Asterosaponins: Structures, taxonomic distribution, biogenesis and biological activities. Mar. Drugs 2020, 18, 584. [Google Scholar] [CrossRef]
- Hossain, A.; Senadheera, T.R.L.; Dave, D.; Shahidi, F. Phenolic profiles of Atlantic sea cucumber (Cucumaria frondosa) tentacles and their biological properties. Food Res. Int. 2023, 163, 112262. [Google Scholar] [CrossRef]
- Senadheera, T.R.L.; Hossain, A.; Dave, D.; Shahidi, F. Antioxidant and ACE-inhibitory activity of protein hydrolysates produced from Atlantic sea cucumber. Molecules 2023, 28, 5263. [Google Scholar] [CrossRef]
- Hossain, A.; Dave, D.; Shahidi, F. Sulfated polysaccharides in sea cucumbers and their biological properties: A review. Int. J. Biol. Macromol. 2023, 253, 127329. [Google Scholar] [CrossRef]
- Sridhar, S.C.J.; Deo, S.C.R. Marine and other aquatic dermatoses. Indian J. Dermatol. 2017, 62, 66–78. [Google Scholar] [CrossRef]
- Afzali, M.; Baharara, J.; Shahrokhabadi, K.; Amini, E. Evaluation of the Cytotoxic Effect of the Brittle Star (Ophiocoma Erinaceus) Dichloromethane Extract and Doxorubicin on EL4 Cell Line. Iran J. Pharm. Res. 2017, 16, 216–226. [Google Scholar]
- Zhukova, N.V. Fatty acids of echinoderms: Diversity, current applications and future opportunities. Mar. Drugs 2022, 21, 21. [Google Scholar] [CrossRef] [PubMed]
- Senadheera, T.R.L.; Hossain, A.; Dave, D.; Shahidi, F. In silico analysis of bioactive peptides produced from underutilized sea cucumber by-products—A bioinformatics approach. Mar. Drugs 2022, 20, 610. [Google Scholar] [CrossRef] [PubMed]
- Senadheera, T.R.L.; Hossain, A.; Dave, D.; Shahidi, F. Functional and physiochemical properties of protein isolates from different body parts of North Atlantic sea cucumber (Cucumaria frondosa). Food Biosci. 2023, 52, 102511. [Google Scholar] [CrossRef]
- Hossain, A.; Dave, D.; Shahidi, F. Effect of high-pressure processing (HPP) on phenolics of North Atlantic sea cucumber (Cucumaria frondosa). J. Agric. Food Chem. 2022, 70, 3489–3501. [Google Scholar] [CrossRef]
- Marmouzi, I.; Tamsouri, N.; Hamdani, M.E.; Attar, A.E.; Kharbach, M.; Alami, R.; Jemli, M.E.; Cherrah, Y.; Ebada, S.S.; Faouzi, M.E.A. Pharmacological and chemical properties of some marine echinoderms. Rev. Bras. Farmacogn. 2018, 28, 575–581. [Google Scholar] [CrossRef]
- Ambigaipalan, P.; Shahidi, F. Bioactive peptides from shrimp shell processing discards: Antioxidant and biological activities. J. Funct. Foods 2017, 34, 7–17. [Google Scholar] [CrossRef]
- Šimat, V.; Rathod, N.B.; Čagalj, M.; Hamed, I.; Mekinić, I.G. Astaxanthin from crustaceans and their byproducts: A bioactive metabolite candidate for therapeutic application. Mar. Drugs 2022, 20, 206. [Google Scholar] [CrossRef]
- Hamed, I.; Özoğul, F.; Regenstein, J.M. Industrial applications of crustacean byproducts (chitin, chitosan, and chitooligosaccharides): A review. Trends Food Sci. Technol. 2016, 48, 40–50. [Google Scholar] [CrossRef]
- Amado, I.R.; González, M.P.; Murado, M.A.; Vázquez, J.A. Shrimp wastewater as a source of astaxanthin and bioactive peptides. J. Chem. Technol. Biotechnol. 2016, 91, 793–805. [Google Scholar] [CrossRef]
- Pangestuti, R.; Kim, S. Bioactive peptide of marine origin for the prevention and treatment of non-communicable diseases. Mar. Drugs 2017, 15, 67. [Google Scholar] [CrossRef]
- Da Silva, C.P.; Bezerra, R.S.; Santos, A.C.O.D.; Messias, J.B.; De Castro, C.R.O.B.; Carvalho, L.B. Biological value of shrimp protein hydrolysate by-product produced by autolysis. LWT Food Sci. Technol. 2017, 80, 456–461. [Google Scholar] [CrossRef]
- De Los Ángeles Pereira, N.; Fangio, M.F.; Rodríguez, Y.E.; Bonadero, M.C.; Harán, N.S.; Fernández-Gimenez, A.V. Characterization of liquid protein hydrolysates shrimp industry waste: Analysis of antioxidant and microbiological activity, and shelf life of final product. J. Food Process. Preserv. 2021, 46, e15526. [Google Scholar]
- Balzano, M.; Pacetti, D.; Lucci, P.; Fiorini, D.; Frega, N.G. Bioactive fatty acids in mantis shrimp, crab and caramote prawn: Their content and distribution among the main lipid classes. J. Food Compos. Anal. 2017, 59, 88–94. [Google Scholar] [CrossRef]
- Mezzomo, N.; Ferreira, S.R.S. Carotenoids functionality, sources, and processing by supercritical technology: A review. J. Chem. 2016, 2016, 3164312. [Google Scholar] [CrossRef]
- Maoka, T. Carotenoids in marine animals. Mar. Drugs 2011, 9, 278–293. [Google Scholar] [CrossRef]
- Pattanaik, S.S.; Sawant, P.B.; Xavier, K.A.M.; Dube, K.; Srivastava, P.P.; Dhanabalan, V.; Chadha, N.K. Characterization of carotenoprotein from different shrimp shell waste for possible use as supplementary nutritive feed ingredient in animal diets. Aquaculture 2020, 515, 734594. [Google Scholar] [CrossRef]
- Fernández-Gimenez, A.V.; Pereira, N.D.L.Á.; Sarasa, M.V. Liposoluble vitamins in crustacean feed: Metabolic and Histological Responses. Indian J. Exp. Biol. 2016, 54, 297–308. [Google Scholar]
- Mao, X.; Guo, N.; Sun, J.; Xue, C. Comprehensive utilization of shrimp waste based on biotechnological methods: A review. J. Clean. Prod. 2017, 143, 814–823. [Google Scholar] [CrossRef]
- Amiri, H.; Shábanpour, B.; Pourashouri, P.; Kashiri, M. Encapsulation of marine bioactive compounds using liposome technique: Evaluation of physicochemical properties and oxidative stability during storage. Food Struct. 2023, 35, 100308. [Google Scholar] [CrossRef]
- Shahidi, F.; Hossain, A. Preservation of aquatic food using edible films and coatings containing essential oils: A review. Crit. Rev. Food Sci. Nutr. 2020, 62, 66–105. [Google Scholar] [CrossRef]
- FAO. 2022. Available online: https://www.fao.org/newsroom/detail/record-fisheries-aquaculture-production-contributes-food-security-290622/en (accessed on 1 October 2023).
- Colletti, A.; Cravotto, G.; Citi, V.; Martelli, A.; Testai, L.; Cicero, A.F.G. Advances in technologies for highly active omega-3 fatty acids from krill oil: Clinical applications. Mar. Drugs 2021, 19, 306. [Google Scholar] [CrossRef]
- Xie, D.; Jin, J.; Sun, J.; Liang, L.; Wang, X.; Zhang, W.; Wang, X.; Jin, Q. Comparison of solvents for extraction of krill oil from krill meal: Lipid yield, phospholipids content, fatty acids composition and minor components. Food Chem. 2017, 233, 434–441. [Google Scholar] [CrossRef]
- Hill, S.E.; Krishnamurthy, R.G.; Hossain, A.; Shahidi, F. Cooking Oils, Salad Oils, and Dressings. In Bailey’s Industrial Oil and Fat Products, 7th ed.; Shahidi, F., Ed.; John Wiley & Sons, Ltd.: Hoboken, NJ, USA, 2020; pp. 1–33. [Google Scholar]
- Zhang, H.; Zhao, H.; Zhang, Y.; Shen, Y.; Su, H.; Jin, J.; Jin, Q.; Wang, X. Characterization of positional distribution of fatty acids and triacylglycerol molecular compositions of marine fish oils rich in omega-3 polyunsaturated fatty acids. Biomed Res. Int. 2018, 2018, 3529682. [Google Scholar] [CrossRef]
- Kaur, G.; Guo, X.; Sinclair, A.J. Short update on docosapentaenoic acid: A bioactive long-chain n-3 fatty acid. Curr. Opin. Clin. Nutr. Metab. Care 2016, 19, 88–91. [Google Scholar] [CrossRef] [PubMed]
- Sala-Vila, A.; Guasch-Ferré, M.; Hu, F.B.; Sánchez-Taínta, A.; Bulló, M.; Serra-Mir, M.; López-Sabater, C.; Sorlí, J.V.; Arós, F.; Fiol, M.; et al. Dietary α-Linolenic Acid, Marine ω-3 Fatty Acids, and Mortality in a Population with High Fish Consumption: Findings from the PREvención con DIeta MEDiterránea (PREDIMED) Study. J. Am. Heart Assoc. 2016, 5, e002543. [Google Scholar] [CrossRef]
- Halim, N.R.A.; Yusof, H.M.; Sarbon, N.M. Functional and bioactive properties of fish protein hydolysates and peptides: A comprehensive review. Trends Food Sci. Technol. 2016, 51, 24–33. [Google Scholar] [CrossRef]
- Villamil, O.; Váquiro, H.A.; Solanilla, J.F. Fish viscera protein hydrolysates: Production, potential applications and functional and bioactive properties. Food Chem. 2017, 224, 160–171. [Google Scholar] [CrossRef]
- Martínez-Álvarez, Ó.; Chamorro, S.; Brenes, A. Protein hydrolysates from animal processing by-products as a source of bioactive molecules with interest in animal feeding: A review. Food Res. Int. 2015, 73, 204–212. [Google Scholar] [CrossRef]
- Hou, Y.; Wu, Z.; Dai, Z.; Wang, G.; Wu, G. Protein hydrolysates in animal nutrition: Industrial production, bioactive peptides, and functional significance. J. Anim. Sci. Biotechnol. 2017, 8, 209–232. [Google Scholar] [CrossRef] [PubMed]
- Yemışken, E.; Jiménez-Rosado, M.; Pérez-Puyana, V.; Sancar, S.; Bektaş, S.; Yıldız, T.; Eryılmaz, L.; Romero, A. Alternative sources of marine bioactive compounds from the Black Sea: Isolation and characterization of fish skin collagen from Neogobius melanostomus (Pallas 1814) (Perciformes: Gobiidae). Reg. Stud. Mar. Sci. 2023, 60, 102887. [Google Scholar]
- Yu, D.; Cui, S.; Chen, L.; Zheng, S.; Zhao, D.; Yin, X.; Yang, F.; Chen, J. Marine-derived bioactive peptides self-assembled multifunctional materials: Antioxidant and wound healing. Antioxidants 2023, 12, 1190. [Google Scholar] [CrossRef] [PubMed]
- Petrova, I.; Tolstorebrov, I.; Eikevik, T.M. Production of fish protein hydrolysates step by step: Technological aspects, equipment used, major energy costs and methods of their minimizing. Int. Aquat. Res. 2018, 10, 223–241. [Google Scholar] [CrossRef]
- Fernandes, P. Enzymes in fish and seafood processing. Front. Bioeng. Biotechnol. 2016, 4, 59. [Google Scholar] [CrossRef] [PubMed]
- Abdelhedi, O.; Salem, A.; Nasri, R.; Nasri, M.; Jridi, M. Food applications of bioactive marine gelatin films. Curr. Opin. Food Sci. 2022, 43, 206–215. [Google Scholar] [CrossRef]
- Wells, M.L.; Potin, P.; Craigie, J.S.; Raven, J.A.; Merchant, S.; Helliwell, K.E.; Smith, A.G.; Camire, M.E.; Brawley, S.H. Algae as nutritional and functional food sources: Revisiting our understanding. J. Appl. Phycol. 2016, 29, 949–982. [Google Scholar] [CrossRef]
- Dinakarkumar, Y.; Krishnamoorthy, S.; Margavelu, G.; Ramakrishnan, G.; Masi, C. Production and characterization of fish protein hydrolysate: Effective utilization of trawl by-catch. Food Chem. Adv. 2022, 1, 100138. [Google Scholar] [CrossRef]
- Shahidi, F.; Hossain, A. Role of lipids in food flavor generation. Molecules 2022, 27, 5014. [Google Scholar] [CrossRef] [PubMed]
- Ciriminna, R.; Meneguzzo, F.; Delisi, R.; Pagliaro, M. Enhancing and improving the extraction of omega-3 from fish oil. Sustain. Chem. Pharm. 2017, 5, 54–59. [Google Scholar] [CrossRef]
- Shahidi, F.; Hossain, A. Bioactives in spices, and spice oleoresins: Phytochemicals and their beneficial effects in food preservation and health promotion. J. Food Bioact. 2018, 3, 8–75. [Google Scholar] [CrossRef]
- Sun, H.; Zhang, Y.; Shen, Y.; Zhu, Y.; Wang, H.; Xu, Z. Inhibitory effects of red wine on lipid oxidation in fish oil emulsion and angiogenesis in zebrafish embryo. J. Food Sci. 2017, 82, 781–786. [Google Scholar] [CrossRef] [PubMed]
- Ghorbanzade, T.; Jafari, S.M.; Akhavan, S.; Hadavi, R. Nano-encapsulation of fish oil in nano-liposomes and its application in fortification of yogurt. Food Chem. 2017, 216, 146–152. [Google Scholar] [CrossRef] [PubMed]
- Hosseini, S.F.; Ramezanzade, L.; McClements, D.J. Recent advances in nanoencapsulation of hydrophobic marine bioactives: Bioavailability, safety, and sensory attributes of nano-fortified functional foods. Trends Food Sci. Technol. 2021, 109, 322–339. [Google Scholar] [CrossRef]

| Bacteria Species | Strains Producing EPSs | Characteristics of Exopolysaccharide | Functions | References |
|---|---|---|---|---|
| Bacillus | Bacillus B3-15, Bacillus B3-72, Bacillus licheniformis, and Bacillus thermoantaticus | Different types of EPSs, consisting of tetra saccharide repeating units or structurally different EPSs consisting of trisaccharide repeating unit with a mannopyranoside configuration | Antiviral and immunomodulatory effects | [15,16,18] |
| Halomonas | Halomonas sp. TG39 | Heterogenous polymer | Emulsifying activity | [19] |
| Planococcus | Planococcus maitriensis Anita I | Contained carbohydrate, protein, uronic acid, and sulfate | Emulsifying activity | [20] |
| Enterobacter | Enterobacter cloacae | Acidic polysaccharide which contained high amount of uronic acid, fucose, and sulfate | Emulsifying activity | [19] |
| Alteromonas | Alteromonus sp. 1545 | Contained glucose, galactose, gluronic acid, galacturonic acid, and 4,6-0-(1-carboxyethilidine)-galactose | Antiviral activity | [21] |
| Biological Activity | Sulfated Polysaccharides | Health Benefits | Species | References |
|---|---|---|---|---|
| Antioxidant | Fucoidan and laminaran | Showed potential to prevent cancer, neurodegenerative, diabetes mellitus, and inflammatory diseases | Sargassum fulvellum, Fucus vesiculosus, and Laminaria japonica | [56,61] |
| Anticoagulant | Fucoidans and carrageenan | Enhanced the inhibition of thrombin and showed anticoagulant activity | Codium fragile, Codium cylindricum, and Monostroma nitidum | [56,62,63] |
| Antiallergic | Fucoidans | Reduced IgE production in both peripheral blood mononuclear cells from healthy donors and atopic dermatitis patients | Undaria pinnatifida, P. binghamiae, Ishige okamurae, Hizikia fusiformis, Meristotheca papulosa, Porphyra yezoensis, S. thunbergia, and E. cava | [19,64] |
| Antiviral | Fucoidan, carrageenan, galactan sulphate, and xylomannan sulphate | Inhibited virus adsorption to cells by competing with virus binding to cells interacting synergistically with the target cell to block the entry of virus novel therapeutic candidates for Herpes simplex virus (HSV-1) and human immunodeficiency virus type (HIV) | Codium fragile, Caulerpa racemose, Sargassum horneri, Sargassum patens, Undaria pinnatifida, Cryptonemia crenulate, Nemalion helminthoides, Chondrus crispus, and Gelidiumcartilagineum | [65,66] |
| Anticancer activity | Fucoidans | Inhibited tumor cell proliferation and tumor cell adhesion to various substrates | Ecklonia cava and Porphyra yezoensis | [67,68] |
| Immunomodulating activity | Carrageenan | Stimulated macrophages modulation and have promising anti-inflammatory activities | Porphyra yezoensis, Gracilaria verrucosa, and Ulva rigida | [62,69] |
| Bioactive Compound | Sources | Biological Activity | References |
|---|---|---|---|
| Peptides | Discodermia sp., Geodia sp., Halichondria, Cylindrata, Theonella sp., Microscleroderma sp., Haliclona nigra, Sidonops microspinosa, Cliona vastifica, Clathria sp., and Clathria sp. | Anticancer, antitumor, antiproliferative, antibacterial, antifungal, and immunosuppressive properties | [5,94,95] |
| Sterols | Halichondria mooriei, Toxadocia zumi, Dysidea arenaria, Hyrtios eubamma, and Disidea pallescens | Antimicrobial properties | [94,96] |
| Phenols | Halichondria panacea, Zygomycale parishii, and Callyspongia diffusa | Antimicrobial and antioxidant properties | [97,98] |
| Terpenoids | Spongia officinalis, Luffarella variabilis, Cacospongia scalaris, Phyllospongia foliascens, Sarcotragus sp., and Prianos sp. | Anticancer, anti-inflammatory, antifungal, antibiotic, and Na, K-ATPase inhibitory activities | [94,98,99] |
| Alkaloids | Stylissa carteri, Hyrtios erecta, Monanchora sp., Corticium sp., Stylissa massa, and Halichondria okadai | Anti-inflammatory and antiviral properties | [93,100] |
| Bioactive Compounds | Echinoderms | Biological Activity | References |
|---|---|---|---|
| Triterpene glycosides/saponins | Bohadschia marmorata, Achlionice Violaecuspidata, Pentacta quadrangularis, Staurocucumis liouvillei, Pearsonothuria graeffei, Eupentacta fraudatrix, Cucumaria frondosa, Holothuria nobilis, Holothuria scabra, and Apostichopus japonicus | Antitumour, anticancer, antiviral, and antifungal activities | [10,91] |
| Chondroitin Sulphate | Holothuria Mexicana, Stichopus japonicas, Holothuria nobilis, Cucumaria frondosa, and Cucumaria japonica | Anticoagulant, antitumor, antiviral antithrombotic, and antioxidant properties | [10,60,106] |
| Steroids | Evasterias retifera, Evasterias echinosoma, Hippasteria kurilensis, Strongylocentrotus nudus, Asterias amurensis, and Craspidaster hesperus | Antitumor, anticancer, immunomodulatory, antibiofouling, antimicrobial, and neuritogenic properties | [107,108] |
| Lipids and fatty acids | Bohadschia argus, Cucumaria frondosa, Acaudina molpadioides, Athyonidium chilensis, Strongylocentrotus nudus, Protoreaster nodosus, Paracentrotus lividus, Strongylocentrotus purpuratus, Lytechinus variegatus, Holothuria eduli, and Holothuria scabra | Anti-inflammatory, antidiabetic, and cholesterol-lowering effects | [10,109] |
| Proteins, amino acids, and peptides | Cucumaria frondosa and Echinus esculentus | Antioxidant, antimicrobial, antihypertensive, and DNA and LDL cholesterol oxidation inhibitory activities | [105,110,111] |
| Phenolic acids and flavonoids | Cucumaria frondosa, Luidia sarsi, Astropecten irregularis, and Ophiura albida | Antioxidant, α-glucosidase, and DNA and LDL cholesterol oxidation inhibitory activities | [104,112,113] |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Senadheera, T.R.L.; Hossain, A.; Shahidi, F. Marine Bioactives and Their Application in the Food Industry: A Review. Appl. Sci. 2023, 13, 12088. https://doi.org/10.3390/app132112088
Senadheera TRL, Hossain A, Shahidi F. Marine Bioactives and Their Application in the Food Industry: A Review. Applied Sciences. 2023; 13(21):12088. https://doi.org/10.3390/app132112088
Chicago/Turabian StyleSenadheera, Tharindu R. L., Abul Hossain, and Fereidoon Shahidi. 2023. "Marine Bioactives and Their Application in the Food Industry: A Review" Applied Sciences 13, no. 21: 12088. https://doi.org/10.3390/app132112088
APA StyleSenadheera, T. R. L., Hossain, A., & Shahidi, F. (2023). Marine Bioactives and Their Application in the Food Industry: A Review. Applied Sciences, 13(21), 12088. https://doi.org/10.3390/app132112088