Abstract
Plinia cauliflora has been associated with numerous ethnobotanical applications. In this study, we uncovered that the fruit extract of P. cauliflora, obtained using 50% ethanol, possesses inhibition activity against the Klebsiella pneumoniae single-stranded DNA-binding protein (KpSSB). SSB plays a critical role in cell survival, making it an attractive target for the development of anti-infective drugs. The inhibition activity against KpSSB by the P. cauliflora extract demonstrated an IC50 value of 73 ± 8 μg/mL. By using gas chromatography–mass spectrometry, the chemical content of this extract was tentatively determined. The top 15 compounds (>0.7%) were as follows: 5-hydroxymethylfurfural, 2,3-dihydro-3,5-dihydroxy-6-methyl-4h-pyran-4-one, 2,5-diformylfuran, furfural, ace-tic acid, citraconic anhydride, formic acid, ethyl 4-hydroxy-3-methylbut-2-enoate, furfuryl alcohol, furyl hydroxymethyl ketone, 3-acetyl-3-hydroxyoxolane-2-one, 2,3-dihydro-5-hydroxy-2-(hydroxymethyl)-4H-pyran-4-one, 2(3H)-furanone, dihy-dro-4-hydroxy-, hydroxyacetone, and 1-hydroxybut-3-en-2-one. To analyze the possible binding modes, the three most abundant compounds were then subjected to docking analysis. We also investigated whether the P. cauliflora extract exhibited any cytotoxic and antiproliferative effects on the survival of B16F10 melanoma cells. Additionally, we found that the extract of P. cauliflora could inhibit the migration and induce apoptosis of B16F10 cells. The results of this study collectively suggest that P. cauliflora holds potential pharmacological benefits, warranting further exploration for therapeutic applications.
1. Introduction
Plants have been characterized as a rich source of pharmaceutical products with therapeutic or prophylactic properties, which are associated with antioxidant, anti-inflammatory, antimicrobial, and antiproliferative activities [1,2,3,4,5]. The strategy of exploring medicinal plants as a source of therapeutic compounds has demonstrated its utility and productivity for potential pharmacological uses [4]. One notable advantage of utilizing natural extracts in therapeutic applications lies in their ability to exert multi-targeted modes of action [6]. Several active constituents derived from plant extracts, including vincristine, vinblastine, and paclitaxel, have already shown promise as effective anticancer drugs [7,8]. Thus, it is worth determining new beneficial properties of plant extracts for various therapeutic purposes.
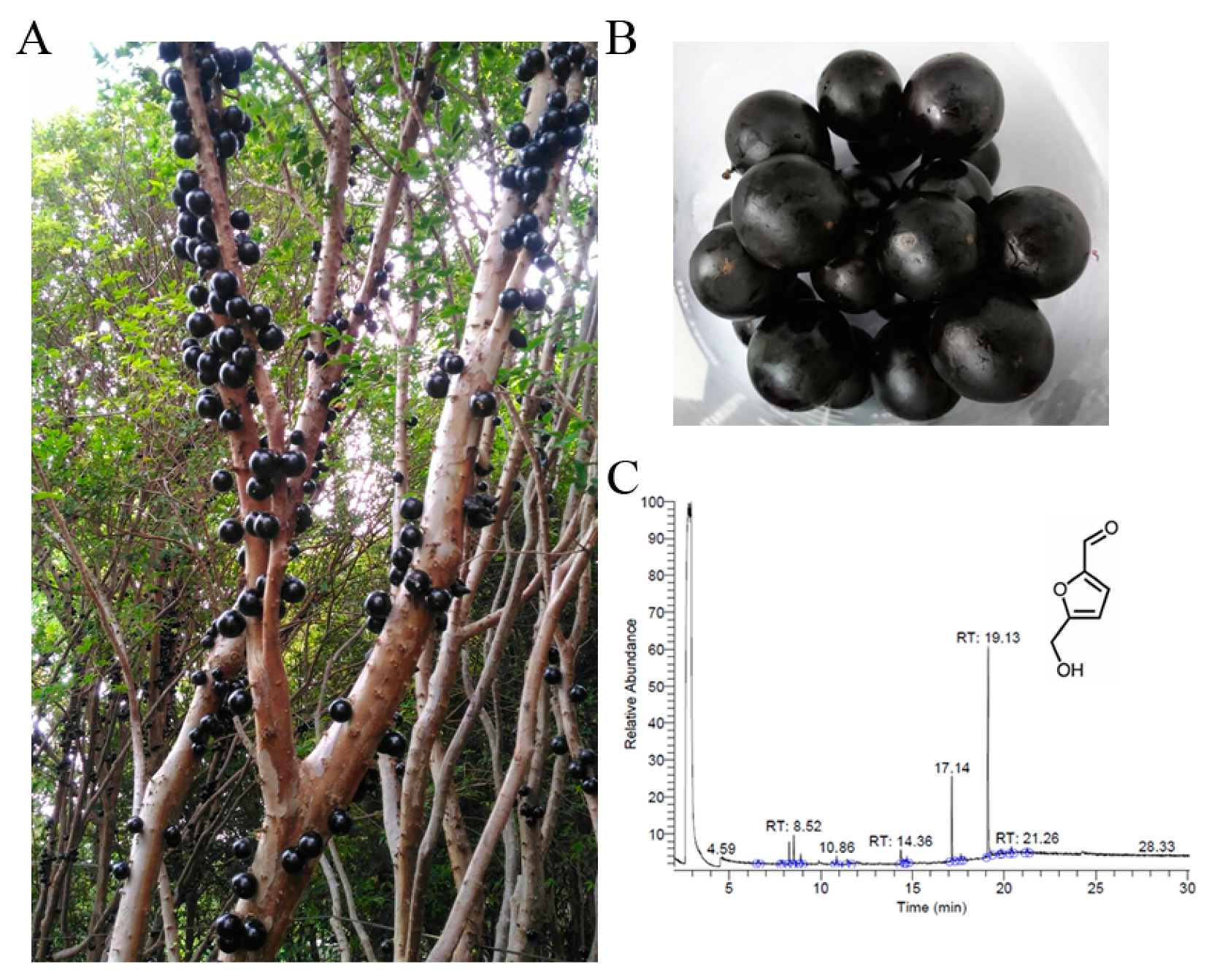
Jaboticaba is the edible fruit of the jabuticabeira (Plinia cauliflora; also known as Myrciaria cauliflora previously) [9]. The fruit of P. cauliflora (jaboticaba) grows directly on the trunk of the tree (Figure 1A). The P. cauliflora fruit is a sweet, thick, and sticky white pulp covered in a dark purple-black outer skin [9,10]. The fruits of P. cauliflora are used as food and contain carbohydrates, minerals, amino acids, and vitamins. The fruits of P. cauliflora are eaten raw or used to make jellies, jams, juices, or wine. This superfruit [11] also contains polyphenols, flavonoids, anthocyanins, carotenoids, and gallotannins [12,13]. P. cauliflora has long been employed for the treatment of diarrhea, labyrinthitis, skin irritations, flu, genitourinary problems, and asthma [10]. Recently, the potential anticarcinogenic and antiproliferative effects of this fruit on several cancer cell lines such as breast [14] and lung cancer cells [15] are reported. Antibacterial activities of the fruit extracts of P. cauliflora were also demonstrated [16,17]. Therefore, determining the molecular targets and understanding the reaction mechanisms are of considerable interest for further biotechnological and pharmacological uses. In this study, the fruit extract of P. cauliflora was used to test for the suppression of B16F10 melanoma cells and the inhibition of the DNA-binding activity of the recombinant single-stranded DNA-binding protein (SSB), an essential DNA replication protein in bacteria [18].
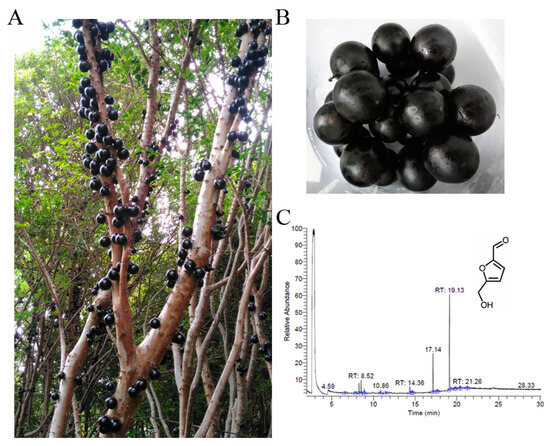
Figure 1.
Preparation and analysis of the fruit extract from Plinia cauliflora. (A) P. cauliflora, commonly used in traditional Brazilian medicine, boasts various ethnobotanical applications. The fruit, also known as jaboticaba, grows directly on the tree trunk. (B) Jaboticaba is the edible fruit of P. cauliflora. (C) GC chromatogram. The content was analyzed by GC–MS. By matching the generated spectra with mass spectral libraries, compounds were tentatively identified. The most abundant compound detected in this extract was 5-hydroxymethylfurfural, with a retention time of 19.13 min.
SSB is essential for all aspects of DNA metabolic processes, including replication, repair, recombination, and replication restart in bacteria [19,20,21,22,23,24]. SSB binds tightly and cooperatively to single-stranded DNA (ssDNA), regardless of sequence [18]. SSB is known to form interactions with numerous DNA metabolism proteins, creating what is referred to as the SSB interactome [25]. Given its crucial roles, SSB presents itself as a potential target for antibacterial drug development [26,27,28]. In eukaryotes, replication protein A (RPA) serves as the equivalent of bacterial SSB [29]. Although RPA shares similar functions with SSB, it differs in terms of structure and numerous other functions [30]. While SSB functions as a homotetramer(s) [24], human RPA (RPA1, RPA2, and RPA3) operates as a heterotrimer [30]. Additionally, RPA binds to its partner proteins through mechanisms that are different from those of SSB [20,30]. Accordingly, pharmacologically inhibiting SSB could be a suitable approach for targeting bacterial pathogens.
As recognized by the Infectious Disease Society of America (IDSA), a faction of antibiotic-resistant bacteria, namely Enterococcus faecium, Staphylococcus aureus, Klebsiella pneumoniae, Acinetobacter baumannii, Pseudomonas aeruginosa, and Enterobacter spp., acronymically dubbed the ESKAPE pathogens, can effectively “escape” the biocidal action of antibiotics [31]. These pathogens cause a number of lethal diseases and antimicrobial resistance (AMR) creates significant challenges in treating bacterial infections in both animals and humans [32,33]. As described by IDSA, these opportunistic ESKAPE pathogens are significantly associated with a high degree of morbidity, mortality, and increased costs. The rise and spread of ESKAPE pathogens with AMR pose a serious threat to global health. K. pneumoniae are one of the dangerous ESKAPE organisms and are a major cause of hospital- and community-acquired infections, including pneumonia, liver abscesses, and sepsis [34,35]. Given that the antibacterial activities of the fruit extracts of P. cauliflora were established [36,37], the molecular targets in K. pneumonia inhibited by extracts of P. cauliflora should be explored. Thus, we assessed here whether the fruit extract of P. cauliflora had activity against K. pneumonia SSB (KpSSB). The pharmacological inhibition of P. cauliflora extract against KpSSB may be useful in suppressing the DNA replication and decreasing the virulence of K. pneumonia.
Considering that DNA replication is an essential process for cell survival [38], DNA replication proteins should be suitable targets for drug development. In this study, we identified that KpSSB, an essential DNA replication protein in K. pneumonia, could be inhibited by the fruit extract of P. cauliflora obtained using 50% ethanol (Figure 1B). Given that human RPA [30] and SSB [20] are different in structure and function, bacterial SSB should be a prime target in antibiotic development. By using gas chromatography–mass spectrometry (GC–MS), the chemical content of this extract was determined. We have recently solved the crystal structure of KpSSB, and thus, the three most abundant compounds in the extract of P. cauliflora were further selected for docking analysis of KpSSB to realize the possible inhibition modes. Given that many natural products have anti-skin-cancer activities [39,40], we also investigated whether the extract of P. cauliflora exhibited any antiproliferative effects on the growth of B16F10 melanoma cells. Additionally, we found that the extract of P. cauliflora could induce apoptosis and inhibit the migration of B16F10 cells. The results of this study collectively suggest that P. cauliflora holds potential pharmacological benefits, warranting further exploration for therapeutic applications.
2. Materials and Methods
2.1. Materials
All solvents and chemicals used were of the highest grade, commercially obtained from Sigma-Aldrich (Saint Louis, MO, USA). The E. coli strain BL21(DE3) pLysS (Novagen, Worcestershire, UK) was used for protein expression. The cell line B16F10 melanoma, obtained from the Food Industry Research and Development Institute, Hsinchu, Taiwan, was maintained as a monolayer culture in Dulbecco’s modified Eagle medium (GibcoTM; Thermo Fisher Scientific, Waltham, MA, USA) supplemented with 10% fetal bovine serum (FBS), 100 unit/mL penicillin, and 100 μg/mL streptomycin. Cells were incubated at 37 °C in a 95% air and 5% CO2 incubator.
2.2. Expression and Isolation of the Recombinant Protein
The gene encoding KpSSB was cloned into pET21b, which incorporates a 6xHis tag at the protein’s N-terminus. The resulting plasmid, referred to as pET21b-KpSSB, was introduced into E. coli BL21 (DE3) cells. These transformed cells were cultivated to an OD600 of 0.9 in LB medium supplemented with 250 μg/mL ampicillin, under vigorous shaking at 37 °C. Recombinant KpSSB was then induced by the addition of 1 mM isopropyl thiogalactopyranoside for 9 h at 25 °C. The induced cells were harvested and resuspended in Buffer A (20 mM Tris–HCl, 5 mM imidazole, and 0.5 M NaCl, pH 7.9), followed by disruption through sonication. The recombinant protein was purified from the soluble supernatant using Ni2+-affinity chromatography (HisTrap HP; GE Healthcare Bio-Sciences, Piscataway, NJ, USA). Elution was performed with Buffer B (20 mM Tris–HCl, 250 mM imidazole, and 0.5 M NaCl, pH 7.9), and the purified protein was subsequently dialyzed against a dialysis buffer (20 mM HEPES and 100 mM NaCl, pH 7.0; Buffer C). Protein purity was assessed by electrophoresis on a 12% SDS-PAGE gel.
2.3. Electrophoretic Mobility Shift Analysis (EMSA)
For analysis of KpSSB binding, a biotinylated deoxythymidine (dT) homopolymer dT30 was employed as a standard assay. The ssDNA dT30 was biotinylated at its 5′ terminal. Incubation of this labeled ssDNA (30 fmol/μL) was with purified KpSSB at various concentrations (0, 20, 39, 78, 156, 312, 625, 1250, 2500, and 5000 nM). The electrophoretic mobility shift analysis (EMSA) was conducted using the LightShift Chemiluminescent EMSA Kit. Briefly, KpSSB and the DNA substrate were incubated together for 60 min at 37 °C in 40 mM Tris–HCl (pH 7.5) and 50 mM NaCl. After adding a dye mixture, an 8% native polyacrylamide gel was used. The reaction samples were analyzed at 100 V for 1 h in TBE running buffer. The protein–DNA complexes were electroblotted onto a positively charged nylon membrane (GE, Boston, MA, USA). A UV-light instrument equipped with 312 nm bulbs was used for a cross-linking reaction. After a 10-min exposure, the transferred DNA was then cross-linked to the nylon membrane. The streptavidin–horseradish peroxidase conjugate and chemiluminescent substrate (Pierce Biotechnology, Waltham, MA, USA) were used for detecting the DNA. The [Protein]50 was estimated based on the concentration of the protein that bound 50% of the input DNA.
2.4. Inhibition Assay
KpSSB at a concentration of 625 nM was subjected to incubation with both dT30 and the extract of P. cauliflora at various concentrations (0, 7.8, 15.6, 31.2, 62.5, 125, 250, 500, and 1000 μg/mL). After EMSA, a titration curve was constructed based on the experimental data. The concentration of the P. cauliflora extract required achieving 50% inhibition (IC50) of KpSSB activity that was directly determined from the graphical analysis.
2.5. Plant Material and Fruit Harvest
Fresh fruits of P. cauliflora (M. cauliflora O. Berg) were procured from a private farm located in Baoshan Township, Hsinchu County, Taiwan (24.7493° N, 121.0213° E). The ripe fruits (around 10 kg) were collected. The collected fruits were cleaned to remove impurities and inspected for damage, alterations, and signs of immaturity or overripeness. Of these collected fruits, 78 fruits of P. cauliflora (1000 g) were selected for further extraction. It took one day from the moment of harvesting the raw material to the point of conducting the analyses and obtaining the extract. The raw material was transported under controlled conditions at 4 °C.
2.6. Extract Preparation
The collected ripe fruits were then dried, pulverized into a fine powder, and subjected to extraction using 50% ethanol to explore their pharmacological potential. From 1000 g of these fruits, 27 g of dry powder were obtained after the lyophilization drying process (BTP-9ES, Benchtop Pro, Wolflabs, Pocklington, UK). In a liquid nitrogen atmosphere, a grinding mill (HR7629, Philips, Amsterdam, The Netherlands) was used for preparing a fine powder. The extraction process (one single extraction) involved placing plant powder (1 g) into a conical flask, adding the extraction solvent (100 mL), shaking the mixture on an orbital shaker for 5 h, and filtering the resulting extract using a 0.45 μm filter. The solvent (50% ethanol) in this extract was then removed via a hot air circulation oven at 40 °C. The P. cauliflora fruit extract was preserved at −80 °C until further use. To create a stock solution (20 mg/mL), the extract powder was dissolved in 20% DMSO. The stock solution was further diluted with the supplemented culture medium to the required assay concentrations for the anticancer cell assays. Incubation of B16F10 cells with the culture medium containing 0.2% DMSO served as a control group. In the case of EMSA, the stock solution was diluted with Buffer C to achieve the designated assay concentrations. Incubation of KpSSB with Buffer C plus 1% DMSO served as a control group for the EMSA experiments.
2.7. GC–MS Analysis
A Thermo Scientific TRACE 1300 Gas Chromatograph was used with a Thermo Scientific ISQ Single Quadrupole Mass Spectrometer system for GC–MS analysis. The conditions were: a Rtx-WAX column, 30 m × 0.25 mm i.d. × 0.25 μm film; the carrier gas, helium (a flow rate of 1 mL/min); the initial oven temperature, 50 °C (and increased to 250 °C at a rate of 10 °C/min); the compounds discharged from the column, a quadrupole mass detector with ions generated through the electron ionization method; and the mass range scanned, 29–650 amu. The relative mass fraction of each chemical component was determined using the peak area normalization method. By comparing the generated spectra with the NIST 2011 and Wiley 10th edition mass spectral libraries, compounds were tentatively identified.
2.8. Agar Well Diffusion Assay
The antibacterial activities of the P. cauliflora extract were assessed using an agar well diffusion assay [41]. A 0.1 McFarland standard suspension was prepared with the pathogens Escherichia coli, Klebsiella pneumoniae, Pseudomonas aeruginosa, and Staphylococcus aureus. These bacterial suspensions were then inoculated into sterile Petri dishes. The plates contained 60 mL of Muller–Hinton agar. The P. cauliflora extract was dissolved in 30% DMSO, and 10 mg of the extract was carefully transferred onto the agar plate. The plates were subsequently incubated at 37 °C for 12 h. The size (diameter) of the inhibition zone serves as an indicator of the antibacterial activity, with larger zones indicating higher potency of the extract. For comparison, ampicillin and 30% DMSO served as the positive and negative controls, respectively. The reported values show the mean standard deviation of at least three independent experiments.
2.9. Total Phenolic Content and Flavonoid Content
The total phenolic content (TPC) was assessed using the Folin–Ciocalteu method [42]. The developed blue color’s absorbance was measured at 750 nm using a UV/VIS spectrophotometer (Hitachi U 3300, Hitachi High-Technologies, Tokyo, Japan). The obtained result was compared to the gallic acid standard curve and expressed as mg of gallic acid equivalent/g dry weight of plant. To determine the flavonoid content, the aluminum chloride colorimetric method was used [43]. The absorbance of both the extracts and standard solutions was measured at 510 nm with the same spectrophotometer. The results are expressed as mg of quercetin equivalent/g dry weight of plant. The individual standard of gallic acid and quercetin used was 0–500 μg/mL. Values show the mean standard deviation of at least three independent experiments.
2.10. Antioxidant Activity Analysis
We determined the antioxidant ability of the P. cauliflora extract using the 1,1-diphenyl-2-picrylhydrazyl (DPPH) radical scavenging assay [44]. The absorbance of the reaction was measured at 517 nm. DPPH free radical scavenging activity was calculated using the formula: %Radical scavenging activity = (Control OD − Sample OD)/Control OD × 100. Values show the mean standard deviation of at least three independent experiments. In comparison, L-ascorbic acid as a positive control had an IC50 value of 28.6 ± 0.2 μg/mL.
2.11. Trypan Blue Cytotoxicity Assay
We evaluated cell death using the trypan blue cytotoxicity assay [45]. In this assay, B16F10 cells (1 × 104) were prepared to incubate with the extract of P. cauliflora in a 100 μL volume. After 24 h, the cytotoxic activity of the extract was estimated via trypan blue dye.
2.12. Chromatin Condensation Assay
We examined apoptosis in cancer cells using the Hoechst 33342 staining method [46]. B16F10 cells were seeded in 96-well plates at a density of 5 × 103 cells per well. The cells were allowed to adhere for 16 h. These resultant cells were then incubated with the P. cauliflora extract for 24 h. Subsequently, the cells were washed with PBS and stained with Hoechst dye (1 μg/mL) in the dark for 10 min. ImageXpress Pico (Molecular Devices, Silicon Valley, CA, USA) was used to capture images of the stained cells. Image acquisition was carried out on the DAPI filter cubes. The obtained images were analyzed using the CellReporterXpress Version 2 software.
2.13. Clonogenic Formation Assay
We evaluated the inhibition of B16F10 cell proliferation using a clonogenic formation assay [47]. In this assay, B16F10 cells were seeded at a density of 1 × 103 cells per well in 6-well plates. These cells were then allowed to incubate overnight for attachment. Following the treatment with the P. cauliflora extract for a period of 5–7 days, the cells were washed with PBS, and the formed colonies were fixed with methanol. After staining with 0.5% crystal violet for 20 min, the number of colonies was counted under a light microscope to determine the extent of proliferation inhibition.
2.14. Wound-Healing Assay
We investigated the inhibition of B16F10 cell migration using the wound-healing assay [48]. The wound-healing assay is a well-established in vitro technique for assessing collective cell migration in two dimensions. In this assay, a cell-free area is generated within a confluent cell monolayer by removing the cells from the designated area. The presence of the cell-free area prompts the remaining cells to migrate and close the gap. Initially, B16F10 cells were seeded and grown in a serum-reduced medium for 6 h. Afterward, a linear wound was created across the well using a pipette tip. Subsequently, the cells were washed twice with the serum-reduced medium. After treatment with the P. cauliflora extract for 24 h, their migration ability was assessed.
2.15. MOE-Dock Analysis
The binding analysis was carried out using MOE-Dock. The binding capacities in comparison to each other were calculated. The structure of KpSSB was used for docking analysis. Any water molecules present in this crystal structure were removed using MOE. To ensure accuracy, a 3D protonation step followed by energy minimization was applied to add hydrogen atoms to the protein structure. The highest-ranked conformations were generated. The binding modes were predicted and visualized directly through the MOE-Dock tool.
2.16. Biorad Protein (Bradford) Assay
The dye reagent was prepared by diluting 1-part Dye Reagent Concentrate with 4 parts deionized water and filtering it through a 0.25 μm filter to remove any particulates. A bovine serum albumin (BSA) protein standard was prepared in concentrations ranging from 0.1 to 0.5 mg/mL and assayed in duplicate. For each standard and sample solution, 10 μL was pipetted into separate wells of a microtiter plate, and 200 μL of the prepared diluted dye reagent was added. The solutions were mixed by pipetting up and down using a multichannel pipette. The plate was then incubated at room temperature for at least 5 min before measuring the absorbance at 595 nm.
2.17. Statistical Analysis
Data were created at least 3 times and expressed as mean ± standard deviation. One-way ANOVA was used to measure significant differences between means. The statistical significance was defined as a p-value < 0.01. These analyses were performed using SPSS software version 26.0 for Windows (IBM, Chicago, IL, USA).
3. Results
3.1. Inhibition of KpSSB by the Extract of Plinia cauliflora
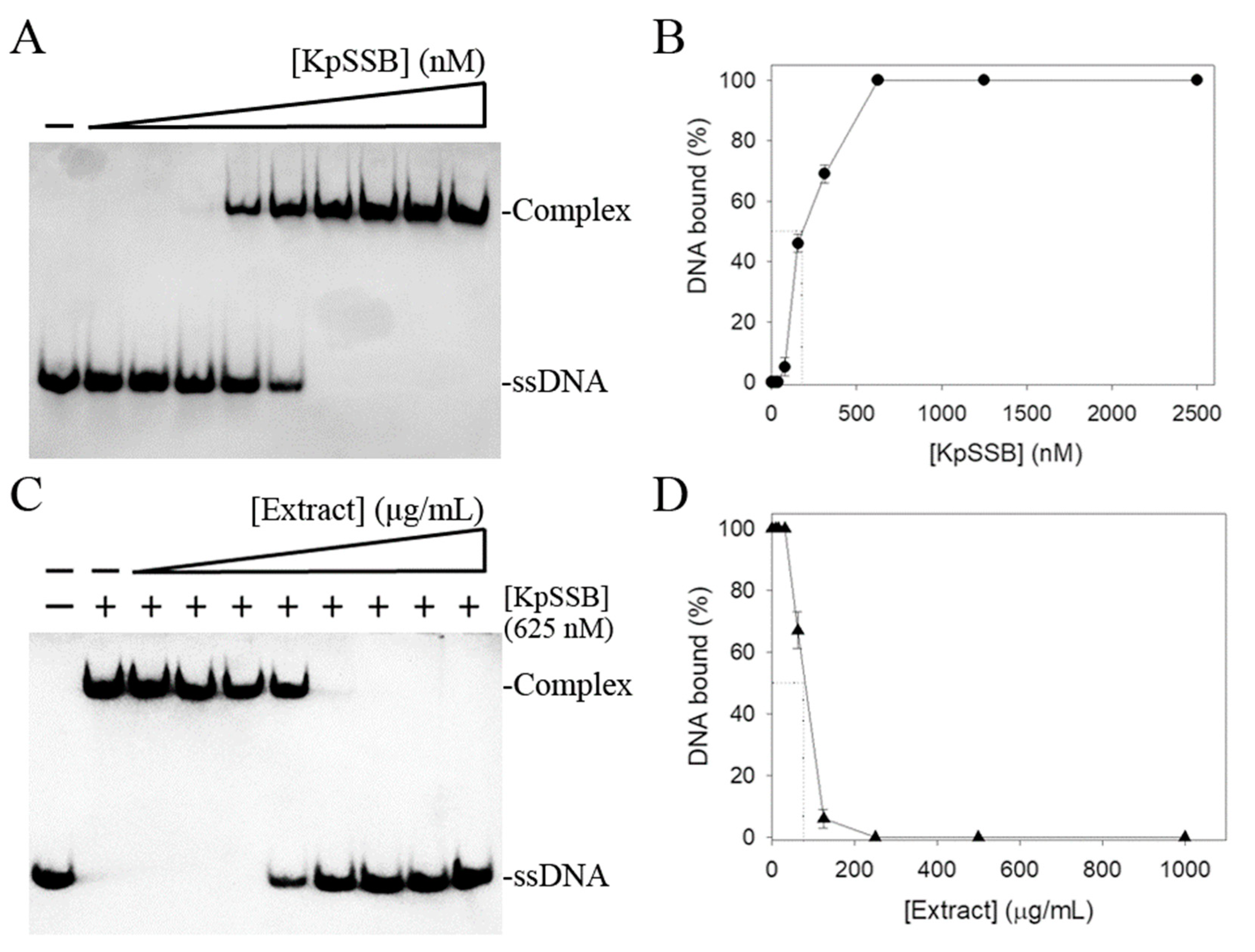
Due to the importance in DNA replication, repair, and recombination, the inhibition of SSB protein by the fruit extract of P. cauliflora as a potential antimicrobial therapy was investigated. KpSSB was used in this demonstration. This extract of P. cauliflora was obtained by 50% ethanol (Figure 1). Recombinant KpSSB was purified by Ni2+-affinity chromatography. To quantify the binding affinity, we conducted EMSA to study the interaction between KpSSB and ssDNA at various protein concentrations. A biotinylated deoxythymidine (dT) homopolymer dT30 was used as a standard assay for the binding analysis of KpSSB. Through a streptavidin–horseradish peroxidase conjugate for detecting the biotin-labeled ssDNA and the complex, incubation of dT30 with KpSSB produced a band shift (Figure 2A). Input dT30 incubated with KpSSB of 625 nM was completely bound (Figure 2A). Through the titration curve, the binding constant ([Protein]50) was calculated to be 175 ± 12 nM (Figure 2B). The extract of P. cauliflora was used to inhibit KpSSB activity (Figure 2C). In this inhibition assay, the extract at concentrations of 7.8–1000 μg/mL was used. The EMSA result showed that the ssDNA binding activity of KpSSB (625 nM) was inhibited by this extract at concentrations of 62.5–1000 μg/mL (Figure 2D). At a concentration of 250 μg/mL, this extract could completely inhibit the binding of KpSSB to dT30. Through the titration curve, the extract of P. cauliflora exhibited an inhibition capacity against KpSSB with an IC50 value of 73 ± 8 μg/mL (Figure 2D). The tuber extract from Sinningia bullata obtained by 50% ethanol was also used for investigating the anti-KpSSB properties; however, the extract of S. bullata (1000 μg/mL) did not influence the binding of KpSSB. Accordingly, a certain compound in the extract of P. cauliflora, alone or in combination, could be a potential anti-KpSSB agent.
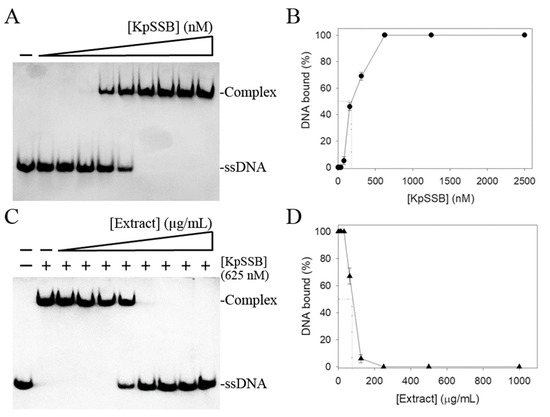
Figure 2.
Inhibition of KpSSB by the fruit extract of P. cauliflora. (A) ssDNA binding of KpSSB. KpSSB (0, 20, 39, 78, 156, 312, 625, 1250, 2500, and 5000 nM) was incubated with dT30. A streptavidin–horseradish peroxidase conjugate was used for detecting this ssDNA and the complex. (B) ssDNA-binding ability of KpSSB. The [Protein]50 of KpSSB was quantified based on the protein concentrations. The errors are standard deviation determined at 3 measurements. (C) Inhibition of the ssDNA-binding activity of KpSSB by the extract of P. cauliflora. KpSSB (625 nM) was incubated with this extract (0, 7.8, 15.6, 31.2, 62.5, 125, 250, 500, and 1000 μg/mL) and the binding activity was analyzed. (D) Inhibition ability of the extract of P. cauliflora. IC50 for KpSSB was determined through the titration curve. The errors are standard deviation determined at 3 measurements.
3.2. Inhibition of KpSSB by Rutin
The fruit extract of P. cauliflora contains several bioactive natural products, including kaempferol, myricetin, quercetin, and rutin [9,49]. To investigate their inhibitory effects on KpSSB, these commercially available chemicals, which were dissolved in 10% dimethyl sulfoxide (DMSO) at concentrations ranging from 0 to 300 μM, were individually included in the binding assay. Using EMSA, we evaluated the impact of each compound on the dT30 binding activity of KpSSB. The results revealed that rutin exhibited an inhibition with an IC50 value of 293 ± 16 μM. On the other hand, kaempferol, myricetin, and quercetin at a concentration of 300 μM did not affect the binding of KpSSB to dT30. As a result, we have identified rutin as a new inhibitor of KpSSB. Our current research involves obtaining crystals of the KpSSB–rutin complex to conduct a more comprehensive analysis of their interactions.
3.3. Antibacterial Activity of the Extract of P. cauliflora
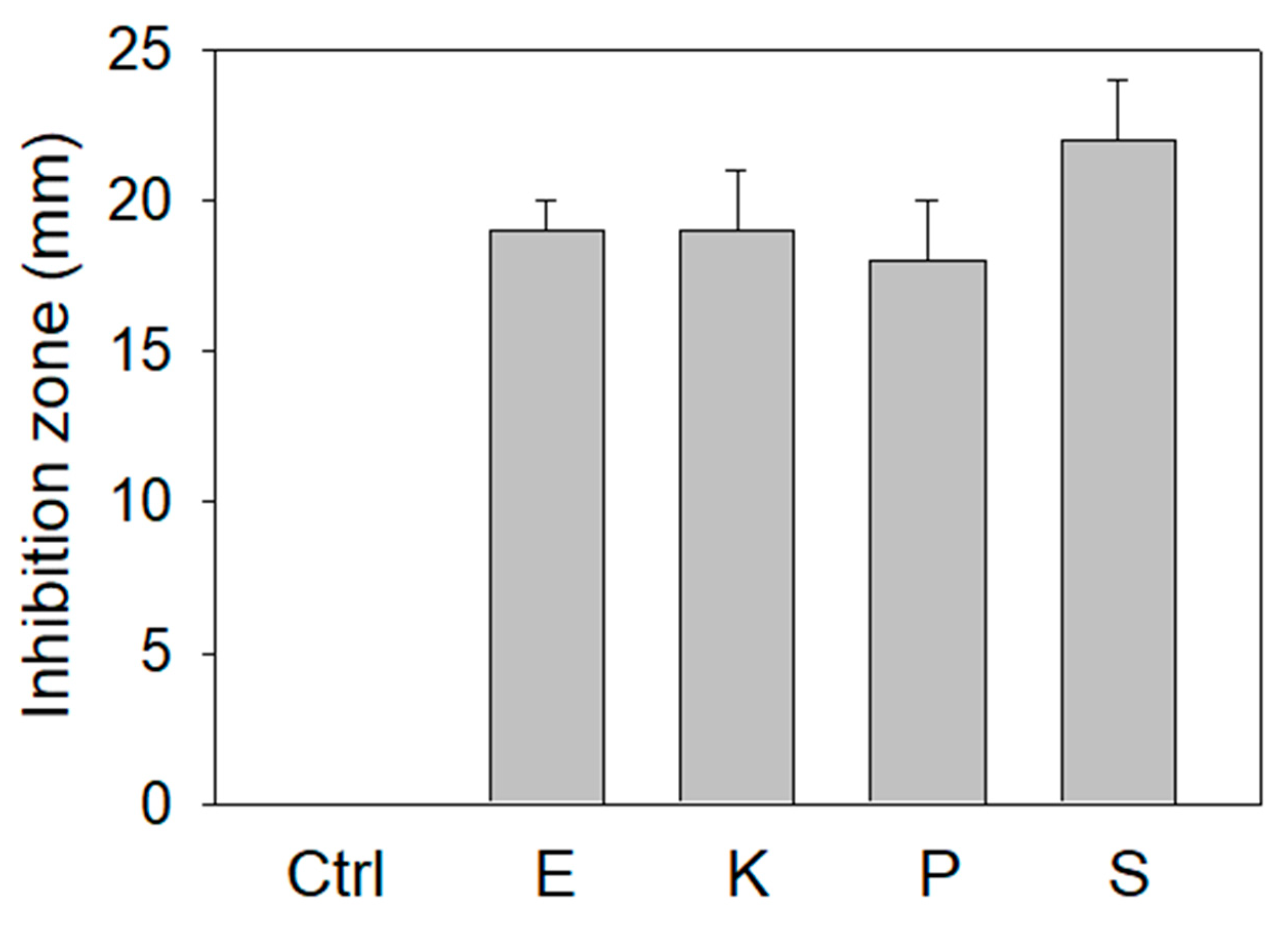
The extract of P. cauliflora could inhibit SSB (Figure 2), the essential protein for bacterial DNA replication. We further investigated the antibacterial activities of this extract using the agar well diffusion method (Figure 3). The zone of inhibition is a circular area around the spot of the extract in which the bacteria colonies do not grow. This extract (10 mg) exhibited antibacterial activities against the human pathogens Escherichia coli, Klebsiella pneumoniae, Pseudomonas aeruginosa, and Staphylococcus aureus used for this analysis and produced the inhibition zones of 19 ± 1, 19 ± 2, 18 ± 2, and 22 ± 2 mm, respectively. Among these bacteria, the extract of P. cauliflora had the highest antibacterial activity against S. aureus. Thus, the extract of P. cauliflora was also effective for antipathogen chemotherapies. This extract may inhibit bacterial DNA replication, resulting in antibacterial activities.
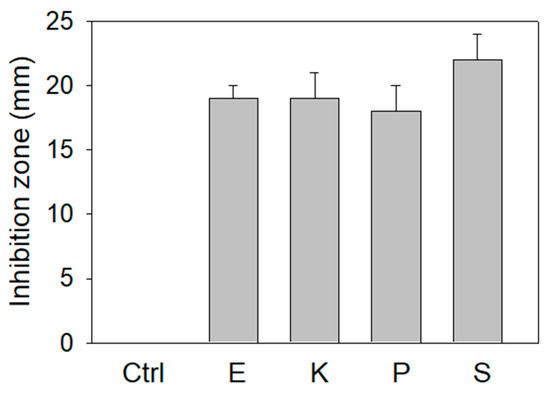
Figure 3.
Agar well diffusion assay. The antibacterial capacities of the extract of P. cauliflora (10 mg) were quantified by the zone of inhibition. The zone of inhibition is a circular area around the spot of the extract in which the bacteria colonies do not grow. This extract exhibited antibacterial activities against human pathogens Escherichia coli (E), Klebsiella pneumoniae (K), Pseudomonas aeruginosa (P), and Staphylococcus aureus (S) used for this analysis and produced the inhibition zones of 19 ± 1, 19 ± 2, 18 ± 2, and 22 ± 2 mm, respectively. DMSO at 30% was used as a negative control.
3.4. TPC, TFC, and Antioxidant Ability of P. cauliflora
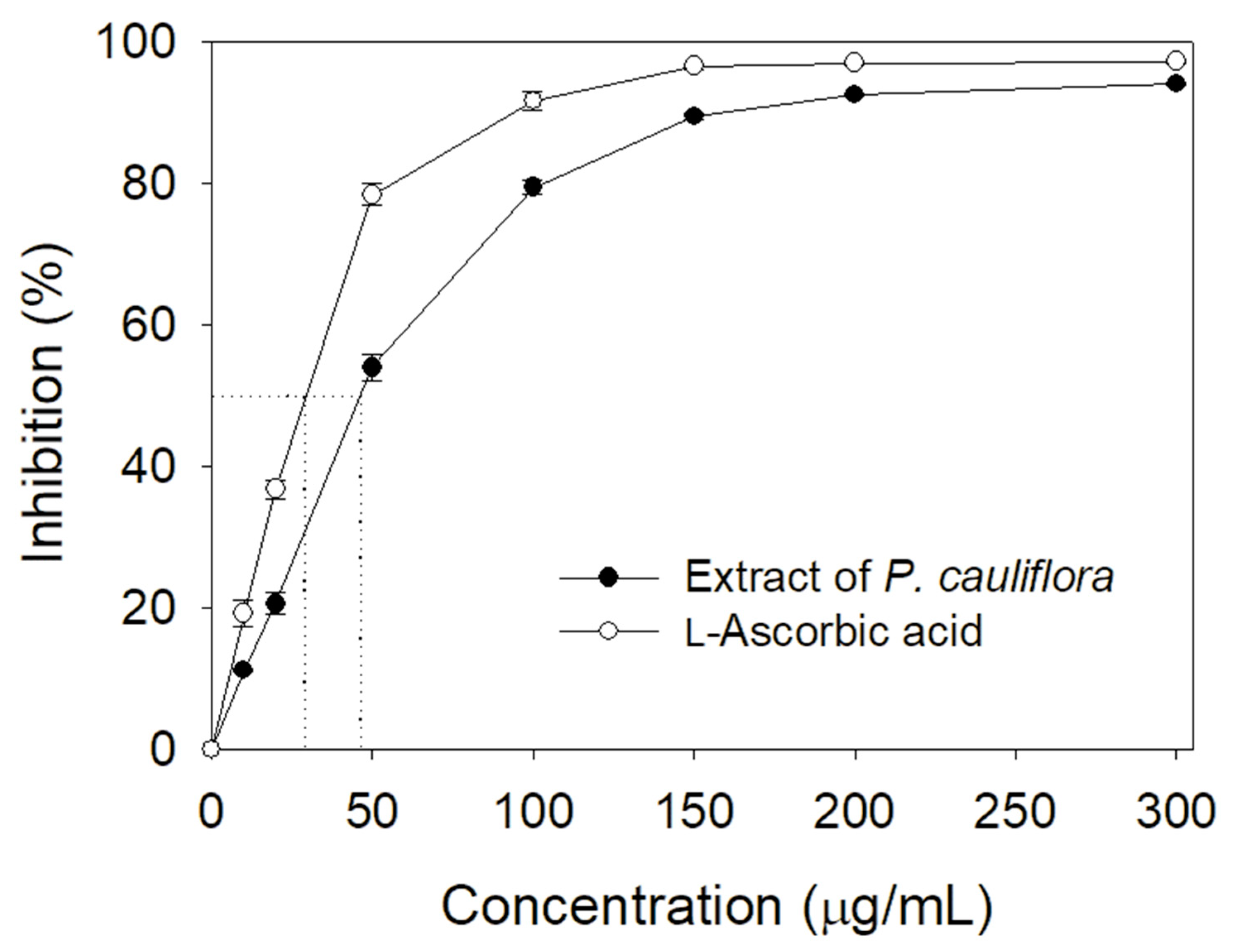
Many polyphenols possessing antioxidant activity can be developed as drug candidates [1,3]. Accordingly, we determined the total phenolic content (TPC), total flavonoid content (TFC), and the antioxidant activity of the extract of P. cauliflora. By using the modified Folin–Ciocalteu method [42] and aluminum chloride colorimetric method [43], the TPC and TFC of this extract were 113.2 ± 1.8 mg gallic acid/g and 102.8 ± 1.2 mg quercetin/g, respectively. By using a 1,1-diphenyl-2-picrylhydrazyl (DPPH) radical scavenging assay, the antioxidant ability of this extract was evaluated and quantified by IC50 value (Figure 4). Through the DPPH titration curve, the extract of P. cauliflora had an IC50 value of 46.5 ± 0.5 μg/mL. In comparison, L-ascorbic acid as a positive control had an IC50 value of 28.6 ± 0.2 μg/mL.
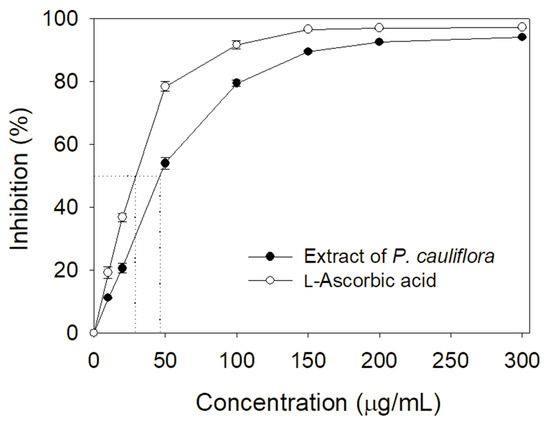
Figure 4.
Antioxidant ability of the extract of P. cauliflora. Through the DPPH titration curve, the extract of P. cauliflora had an IC50 value of 46.5 ± 0.5 μg/mL. L-Ascorbic acid as a positive control had an IC50 value of 28.6 ± 0.2 μg/mL. The errors are standard deviation determined at 3 measurements.
3.5. GC–MS Analysis
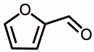
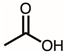
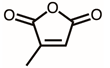
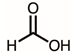
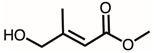
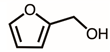
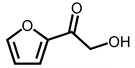
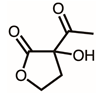
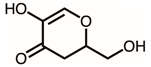
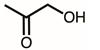
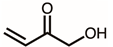
Through GC–MS analysis, the most abundant compounds present in this extract were tentatively determined (Table 1). The top 15 compounds (>0.7%) were as follows: 5-hydroxymethylfurfural, 2,3-dihydro-3,5-dihydroxy-6-methyl-4h-pyran-4-one, 2,5-diformylfuran, furfural, acetic acid, citraconic anhydride, formic acid, ethyl 4-hydroxy-3-methylbut-2-enoate, furfuryl alcohol, furyl hydroxymethyl ketone, 3-acetyl-3-hydroxyoxolane-2-one, 2,3-dihydro-5-hydroxy-2-(hydroxymethyl)-4H-pyran-4-one, 2(3H)-furanone, dihydro-4-hydroxy-, hydroxyacetone, and 1-hydroxybut-3-en-2-one. Possibly, certain compounds in this extract were capable of inhibiting KpSSB and thus attractive inhibitors.

Table 1.
Compounds tentatively identified by the GC–MS analysis.
3.6. Cytotoxic Effect of the Extract of P. cauliflora
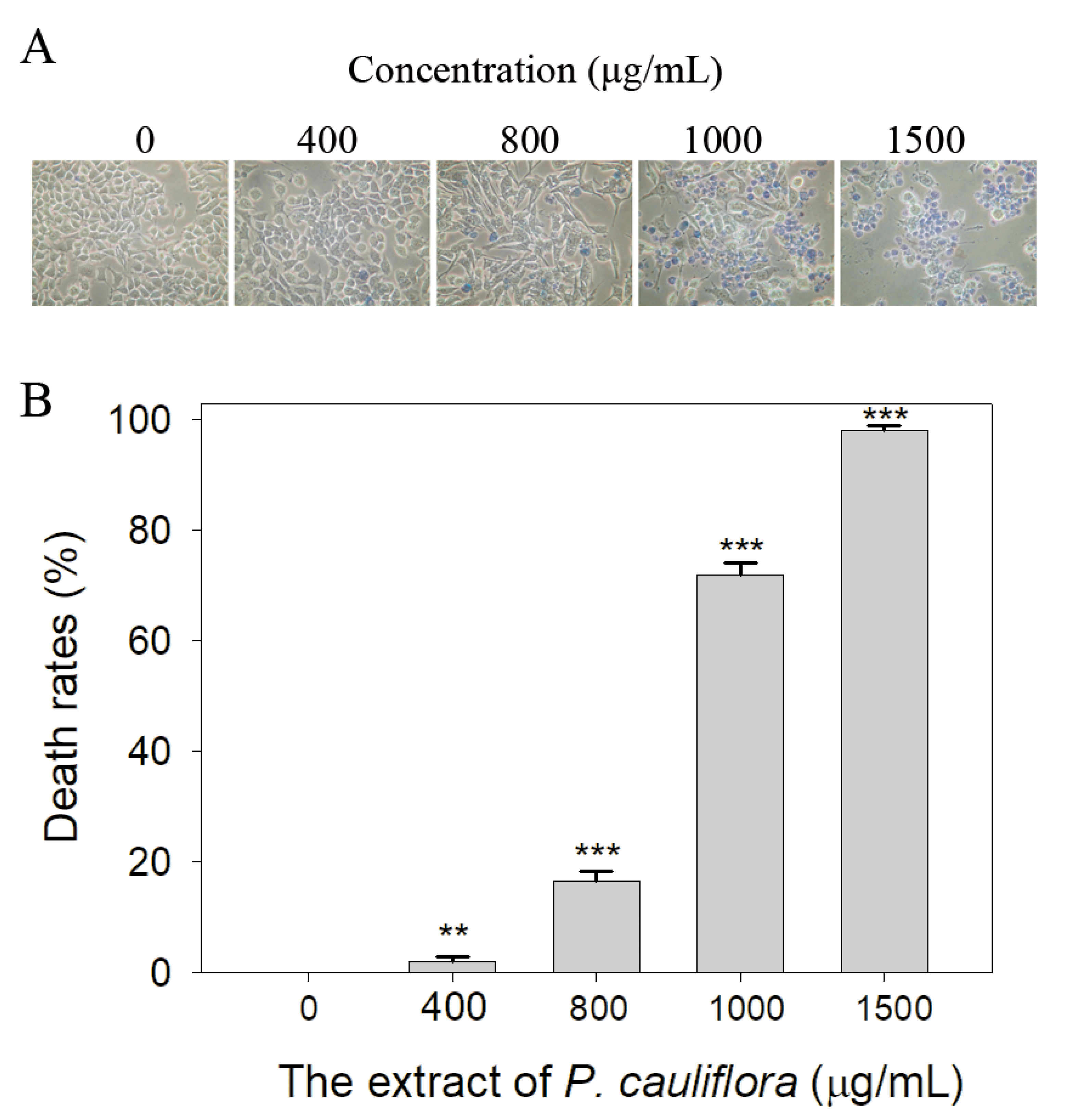
Cancer remains a highly life-threatening disease on a global scale, and its mortality rates have been steadily increasing, making it a major contributor to human mortality [50,51]. Many natural products have demonstrated anticancer properties, especially concerning skin cancers [52]. Accordingly, we investigated the cytotoxic effect of the extract of P. cauliflora. B16F10 melanoma cells and a trypan blue staining assay (Figure 5) were used in this investigation. The B16F10 cell monolayers in 96-well microtitration plates were exposed to the extract at different concentrations per well. To achieve the indicated assay concentrations, the extract stock (20 mg/mL) was diluted using the culture medium. Incubation of B16F10 cells with the resulting extract solutions or the culture medium with 0.2% DMSO served as the treatment and control groups, respectively. Upon incubation, the extract at concentrations of 400, 800, 1000, and 1500 μg/mL caused cell death in B16F10 cells at rates of 2%, 17%, 72%, and 98%, respectively. Incubation of B16F10 cells with the extract (1500 μg/mL) resulted in nearly complete cell death. These findings indicate that the P. cauliflora extract exhibits anti-SSB activity and possesses anti-cancer potential, making it a promising candidate for further medical applications.
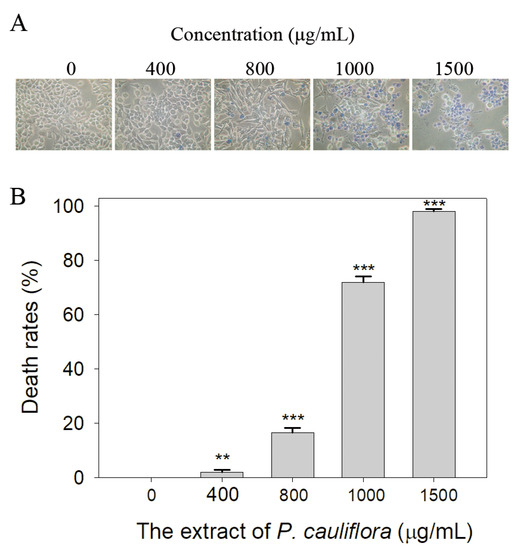
Figure 5.
Cytotoxic effect of the extract of P. cauliflora. (A) The cytotoxic effect of the extract of P. cauliflora was estimated using trypan blue staining assay. The direct identification of live (unstained) and dead (blue) B16F10 cells was allowed by using this assay. The incubation of B16F10 cells with the extract at a concentration of 1500 μg/mL resulted in nearly complete cell death. (B) The death rates of B16F10 cells. Incubation with 400, 800, 1000, and 1500 μg/mL of the extract of P. cauliflora caused the deaths at rates of 2, 17, 72, and 98%, respectively. B16F10 cells were incubated with the culture medium with 0.2% DMSO as a control group. ** p < 0.01 and *** p < 0.001 compared with the control group.
3.7. The Extract of P. cauliflora Induced Apoptosis of B16F10 Cells
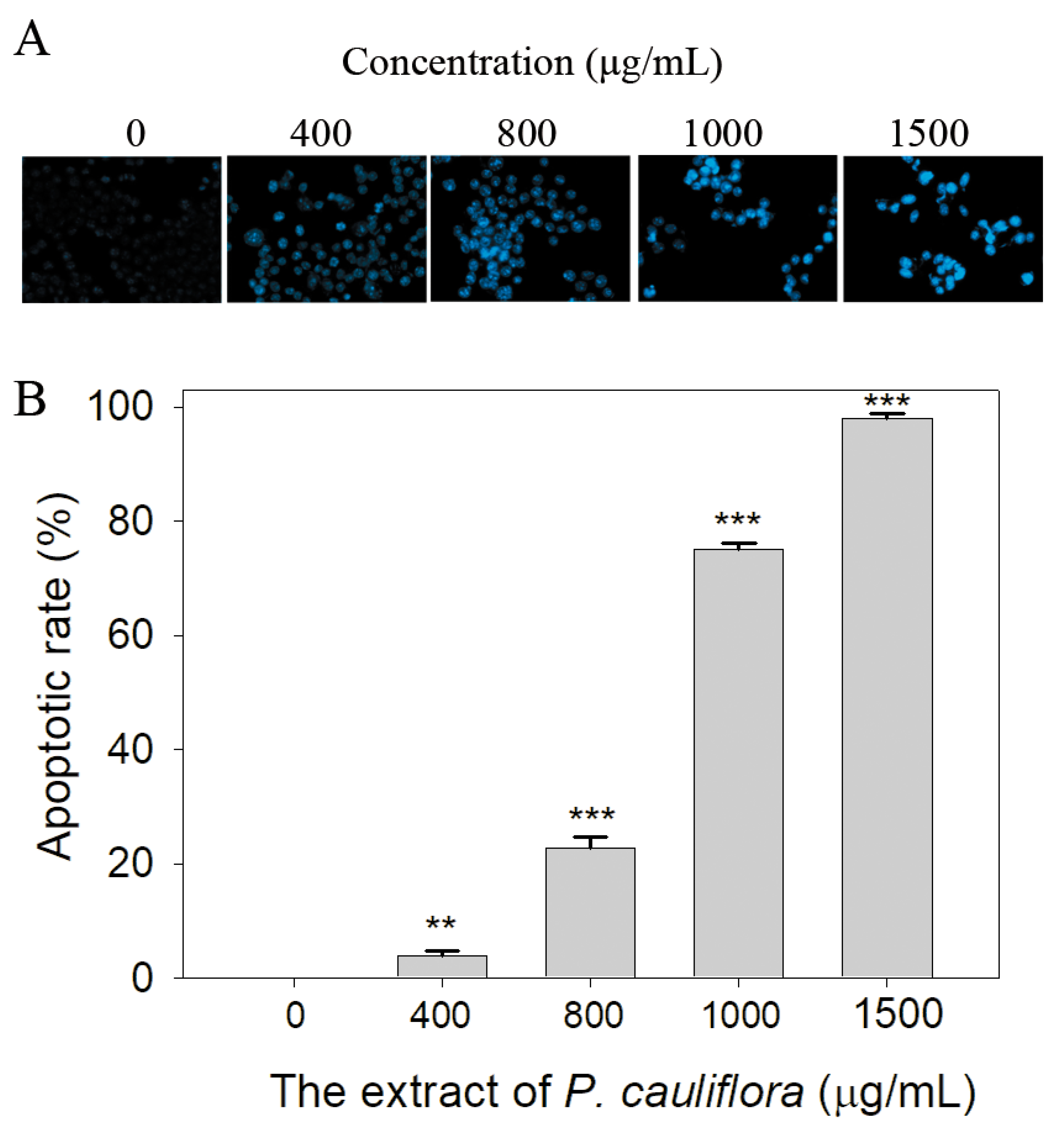
SSB plays a crucial role in various cellular processes, including DNA damage repair, maintenance of genome stability, and the regulation of cell apoptosis [19,53]. Given that the extract of P. cauliflora had anti-SSB properties, we investigated whether this extract could induce apoptosis of B16F10 cells using the Hoechst staining assay. Based on the results, we found that the extract of P. cauliflora could induce apoptosis of B16F10 cells (Figure 6A). The incubation of B16F10 cells with 400, 800, 1000, and 1500 μg/mL of this extract caused 4, 23, 75, and 98% DNA fragmentation (Figure 6B).
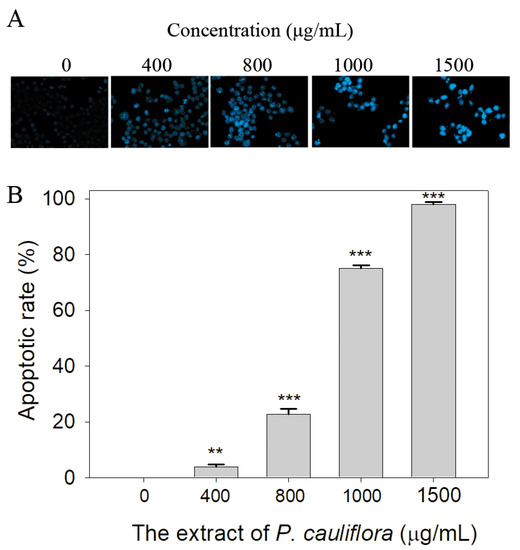
Figure 6.
The extract of P. cauliflora induced apoptosis of the cells. (A) The apoptotic rate in B16F10 cells was estimated using the Hoechst staining assay. Based on our results, the extract of P. cauliflora could induce dose-dependent apoptosis of B16F10 cells. (B) The apoptotic rate. Incubation of B16F10 cells with 400, 800, 1000, and 1500 μg/mL of this extract caused 4, 23, 75, and 98% DNA fragmentation. The errors are the standard deviation as determined over 3 measurements. B16F10 cells were incubated with the culture medium with 0.2% DMSO as a control group. ** p < 0.01 and *** p < 0.001 compared with the control group.
3.8. The Extract of P. cauliflora Inhibited the Migration of B16F10 Cells
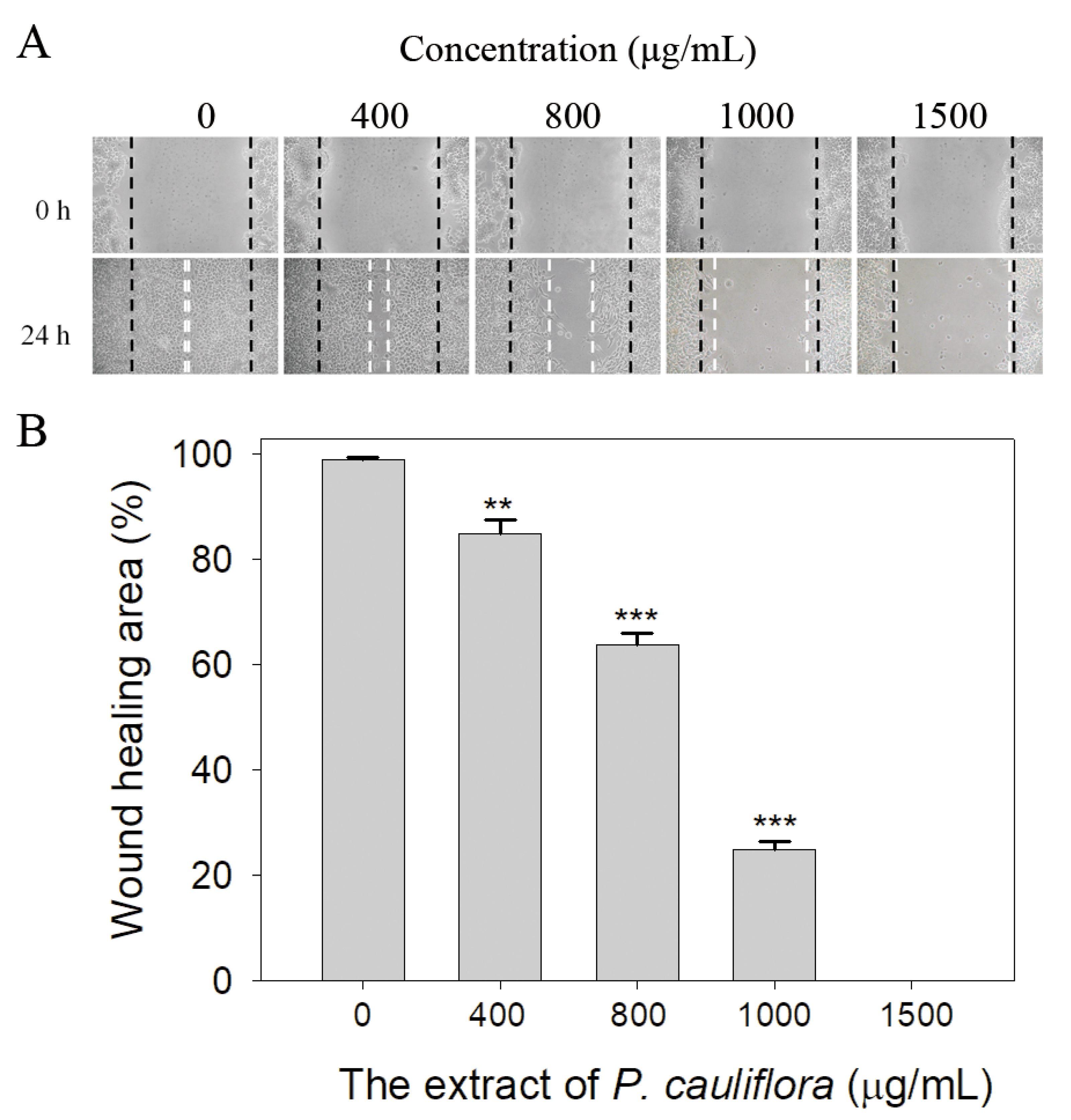
We also observed the extract of P. cauliflora’s ability to inhibit the cell migration using the wound healing assay (Figure 7). The wound healing assay is a well-established in vitro technique for assessing collective cell migration in two dimensions (Figure 7A). In this assay, a cell-free area is generated within a confluent cell monolayer by removing the cells from the designated area. The presence of the cell-free area prompts the remaining cells to migrate and close the gap. We observed that after 24 h of treatment, the P. cauliflora extract at concentrations of 400, 800, 1000, and 1500 μg/mL reduced B16F10 cell migration by 15%, 36%, 75%, and 100%, respectively (Figure 7B). These findings indicate that the P. cauliflora extract is capable of effectively inhibiting the migration of B16F10 cells.
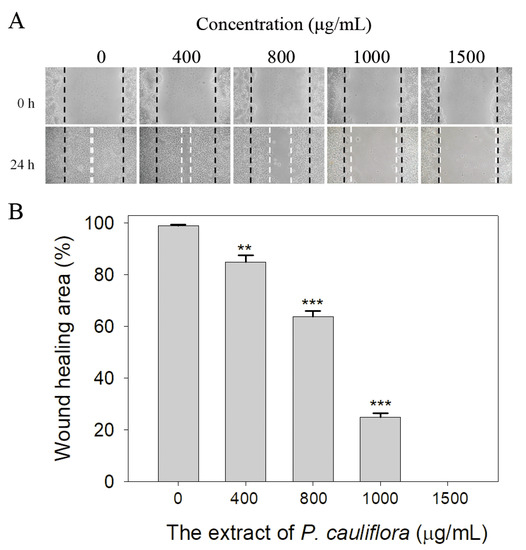
Figure 7.
The extract of P. cauliflora inhibited the cell migration. (A) B16F10 cell migration was analyzed after a 24-h incubation with the P. cauliflora extract using the wound healing assay. The dashed lines indicate the wound edges. The P. cauliflora extract (1500 μg/mL) completely inhibited B16F10 cell migration. (B) The wound healing area was quantified, and the results showed that after 24 h of incubation, the P. cauliflora extract at concentrations of 400, 800, 1000, and 1500 μg/mL reduced B16F10 cell migration by 15%, 36%, 75%, and 100%, respectively. The error bars represent the standard deviation, calculated from three independent measurements. As a control group, B16F10 cells were incubated with the culture medium containing 0.2% DMSO. ** p < 0.01 and *** p < 0.001 compared with the control group.
3.9. The Extract of P. cauliflora Inhibited the Proliferation of B16F10 Cells
Through the clonogenic assay, we observed that the P. cauliflora extract effectively inhibited the cell proliferation (Figure 8A). The clonogenic assay is a well-established in vitro cell survival test, which assesses the capacity for a single cell to form a colony. When pre-treated with the P. cauliflora extract at concentrations of 400, 800, 1000, and 1500 μg/mL, the colony formation of B16F10 cells were suppressed by 8%, 29%, 69%, and 96%, respectively (Figure 8B). These results indicate that the P. cauliflora extract has a significant dose-dependent effect in suppressing the growth and proliferation of B16F10 cells.
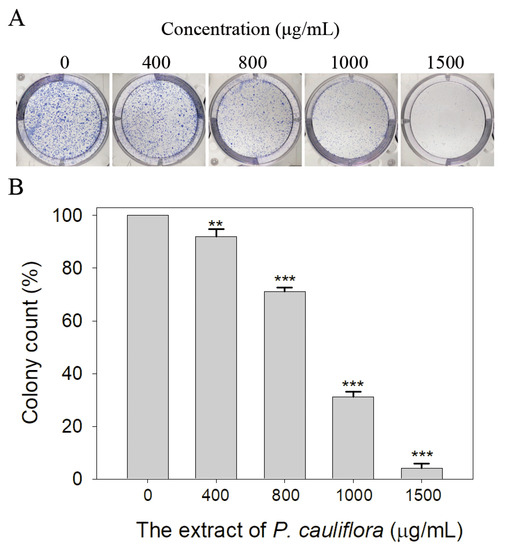
Figure 8.
The extract of P. cauliflora inhibited the cell proliferation. (A) Estimation of the proliferation of B16F10 cells using the clonogenic assay. The P. cauliflora extract (1500 μg/mL) almost completely suppressed the colony formation. (B) The quantified results of the proliferation of B16F10 cells are presented. The P. cauliflora extract at concentrations of 400, 800, 1000, and 1500 μg/mL significantly inhibited the proliferation of B16F10 cells by 8%, 29%, 69%, and 96%, respectively. The error bars represent the standard deviation from 3 independent measurements. As a control group, B16F10 cells were incubated with the culture medium containing 0.2% DMSO. ** p < 0.01 and *** p < 0.001 compared with the control group.
3.10. Molecular Docking
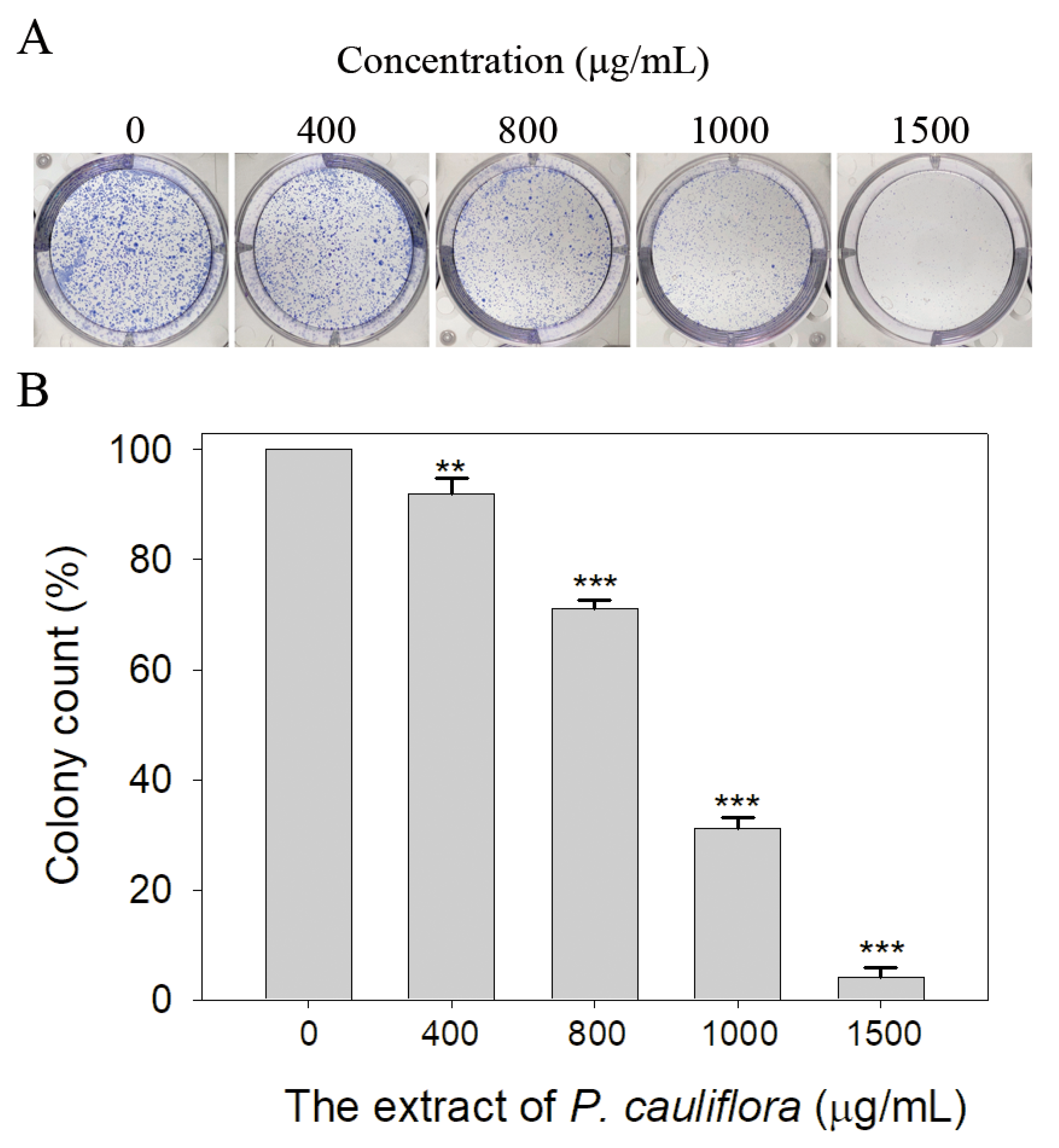
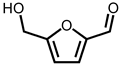
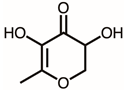
Due to the high inhibitory capability, the compounds abundant in the extract of P. cauliflora were analyzed to determine the possible binding/inhibition modes via the docking studies. Based on the GC–MS analysis (Table 1), the top 3 compounds (>6.0%), namely 5-hydroxymethylfurfural, 2,3-dihydro-3,5-dihydroxy-6-methyl-4h-pyran-4-one, and 2,5-diformylfuran, were used for docking analysis. The binding mode of these compounds to KpSSB was elucidated using the MOE Dock tool (Figure 9). Previously, we solved the crystal structure of KpSSB. The structure of KpSSB (Figure 9A) was therefore used for this docking analysis. KpSSB is a homotetramer. Given that the ssDNA-complexed structure of KpSSB is unavailable, the structure of P. aeruginosa SSB (PaSSB) bound by ssDNA dT25 (PDB ID 6IRQ) was used for analysis (Figure 9B). Bound ssDNA in PaSSB was found to adopt an S-shaped conformation (Figure 9B). Considering their structural similarity, the ssDNA-binding mode between KpSSB and PaSSB may be similar (Figure 9C). According to the docking results (Figure 9C), 5-hydroxymethylfurfural (Figure 9D), 2,3-dihydro-3,5-dihydroxy-6-methyl-4h-pyran-4-one (Figure 9E), and 2,5-diformylfuran (Figure 9F) occupy the ssDNA-binding sites of KpSSB. Thus, these compounds may influence the ssDNA-binding activity of KpSSB by this inhibition mechanism. The binding energies of these compounds were also calculated by MOE (Table 2). The predicted binding affinities for these KpSSB complexes were calculated for all potential binding geometries. Their docking scores revealed the following order for binding affinity (the S score): 5-hydroxymethylfurfural > 2,5-diformylfuran > 2,3-dihydro-3,5-dihydroxy-6-methyl-4h-pyran-4-one. The compound 5-hydroxymethylfurfural may possess the greatest binding affinity for KpSSB among the selected compounds, as indicated by the highest S score (−4.3646). However, this conjecture requires further experimental and structural investigations to be confirmed.
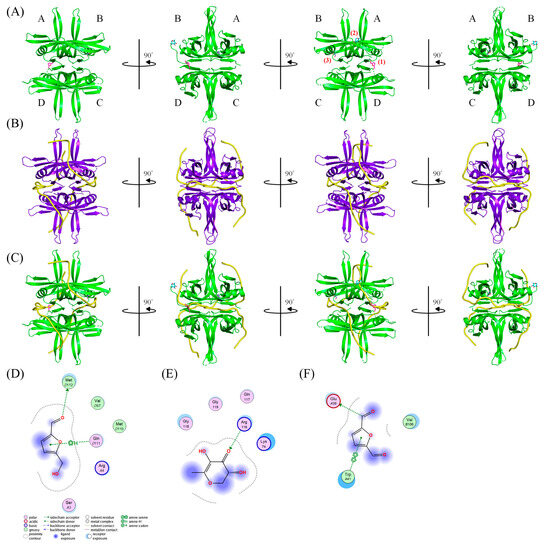
Figure 9.
The docking studies. (A) Crystal structure of KpSSB. KpSSB is a homotetramer with subunits A, B, C, and D. The binding mode of (1) 5-hydroxymethylfurfural (light magenta), (2) 2,3-dihydro-3,5-dihydroxy-6-methyl-4h-pyran-4-one (cyan), and (3) 2,5-diformylfuran (slate) to KpSSB was elucidated using the MOE Dock tool. (B) The structure of PaSSB bound by ssDNA dT25. The complexed crystal structure of a PaSSB tetramer contains two ssDNA molecules (yellow). Given that the ssDNA-complexed structure of KpSSB is unavailable, the complexed structure of PaSSB was used for analyzing the ssDNA-binding mode of KpSSB. (C) The superimposed structures. Superimposing the ssDNA bound by PaSSB and the docked compounds bound by KpSSB indicates that these three compounds are likely to be situated at the sites of KpSSB for ssDNA binding. The binding modes of (D) 5-hydroxymethylfurfural, (E) 2,3-dihydro-3,5-dihydroxy-6-methyl-4h-pyran-4-one, and (F) 2,5-diformylfuran to KpSSB are shown. The interaction analysis is depicted through two-dimensional diagrams.
Table 2.
Molecular docking analysis.
4. Discussion
P. cauliflora has been associated with numerous ethnobotanical applications in treating various conditions such as diarrhea, labyrinthitis, skin irritations, flu, genitourinary problems, and asthma [10]. The commercial potential of P. cauliflora is significant, as its fruit is highly appreciated for both fresh consumption and for its use in making jelly, juice, ice cream, fermented beverages, and liqueurs [10]. This fruit is considered a functional food due to its richness in anthocyanins, exhibiting antioxidant and anti-inflammatory properties, making it sought after by the pharmaceutical industry for its nutraceutical characteristics. In our study, we investigated the extract of P. cauliflora obtained using 50% ethanol and found that it has the capability to inhibit the activity of KpSSB (Figure 2), an essential DNA replication protein in K. pneumoniae. K. pneumoniae is classified as one of the dangerous ESKAPE organisms and can effectively “escape” the biocidal action of antibiotics [31]. Given its association with life-threatening infections, it is important to combat this pathogen in some effective ways [34,35]. SSB plays a vital role in cellular survival, making it a potential target for pharmacological inhibition to combat bacterial pathogens. Inhibiting DNA replication and repair has a long-standing history as an antimicrobial strategy in antibiotic development. Notably, antibiotics such as quinolones and aminocoumarins were successfully developed for clinical applications by targeting bacterial DNA gyrase and topoisomerase IV [54,55]. As plants have been a rich source of pharmaceutical products with antibacterial properties, it is worthwhile to explore different plant extracts and continue the search for inhibitors against SSBs. The findings from our study provide promising insights into the potential therapeutic applications of P. cauliflora and its extract in combating bacterial pathogens by targeting essential DNA replication proteins.
According to the GC–MS analysis (Table 1), the top 15 compounds (>0.7%) in the fruit extract of P. cauliflora were tentatively identified as follows: 5-hydroxymethylfurfural, 2,3-dihydro-3,5-dihydroxy-6-methyl-4h-pyran-4-one, 2,5-diformylfuran, furfural, acetic acid, citraconic anhydride, formic acid, ethyl 4-hydroxy-3-methylbut-2-enoate, furfuryl alcohol, furyl hydroxymethyl ketone, 3-acetyl-3-hydroxyoxolane-2-one, 2,3-dihydro-5-hydroxy-2-(hydroxymethyl)-4H-pyran-4-one, 2(3H)-furanone, dihydro-4-hydroxy-, hydroxyacetone, and 1-hydroxybut-3-en-2-one. This extract may contain compounds that could potentially act as KpSSB inhibitors. However, it is essential to note that other compounds, such as catechin, epicatechin, quercetin, and rutin, have also been identified in various extracts from P. cauliflora [9]. In our study, we observed that rutin exhibited an inhibitory effect on the DNA-binding activity of KpSSB with an IC50 value of 293 ± 16 μM. Rutin, also known as vitamin P, is abundantly found in various vegetables and fruits and is known to possess antibacterial, anti-inflammatory, antitumor, and antiviral properties. Its anticancer potential has been associated with the regulation of multiple signaling pathways, including NF-κB, PI3K-Akt-mTOR, Nrf2, ERK, JNK, and p38 MAPK [56]. Given the inhibition capacity against the DNA replication protein SSB, it is worth continuing the search for inhibitors against SSB and for finding possible DNA-binding targets in cancer cells.
SSB typically recognizes ssDNA [57,58] through a highly conserved oligonucleotide/oligosaccharide-binding (OB) fold. The OB fold structure consists of a five-stranded β-barrel capped by an α-helix [59]. SSB is comprised of two domains: an N-terminal domain for ssDNA-binding and oligomerization (SSBn) and a C-terminal domain for protein–protein interaction (SSBc). To date, the structure of SSBc has not been observed. Recently, the structure of the six additional residues in SSBc is revealed by our crystal structure of KpSSB. These residues, 114-GGRQGG-119 in SSBc, can interact with SSBn but are released when SSBn binds to ssDNA [60]. Thus, this motif in SSB may be a switch in regulating the ssDNA binding [60]. In our study, the docking results indicated that R116 in KpSSB plays a crucial role in forming hydrogen bonds and interacting with the potential inhibitor 2,3-dihydro-3,5-dihydroxy-6-methyl-4h-pyran-4-one (Figure 8E). Consequently, this compound may act as an inhibitor, potentially affecting the regulatory switch for SSBn or ssDNA binding. Given the relationship between the GGRQ motif and the ssDNA binding sites, further investigations are needed to understand whether this motif also impacts the SSB35/SSB65 distribution [24]. We are currently working on obtaining crystals of the KpSSB-inhibitor complex to conduct a more comprehensive analysis of these interactions.
P. cauliflora extracts have been previously recognized for their anti-proliferative activities and potential anti-carcinogenic effects on various cancer cell lines, including breast [14] and lung cancer cells [15]. The peel extracts exhibited inhibitory effects against Bacillus subtilis, S. aureus, P. aeruginosa, and E. coli bacteria [61]. In alignment with these findings, our study revealed the antibacterial activities (Figure 3), as well as the cytotoxic (Figure 5) and antiproliferative effects (Figure 8) of the P. cauliflora extract on the B16F10 cells. Additionally, we also found that the extract of P. cauliflora could inhibit the cell migration and induce apoptosis. These preliminary results suggest that the extract of P. cauliflora holds promise as a potential natural alternative for melanoma cancer, warranting further exploration for therapeutic applications.
By using a DPPH assay, the IC50 of an ethanol extract of P. cauliflora seeds and peel had been reported to be 27 [62] and 37 μg/mL [63], respectively. In this study, the fruit extract obtained using 50% ethanol had an IC50 value of 46.5 ± 0.5 μg/mL (Figure 4). Regardless of the cost of alcohol, ethanol could be a better solvent for extractions. In conclusion, this study represents the first identification that the fruit extract from P. cauliflora has the anti-KpSSB property. Rutin is a new inhibitor against KpSSB. Furthermore, the cytotoxic activities against melanoma cells were also investigated. Through GC–MS analysis, the top 15 compounds in this extract were detected, providing insights into the potential active components and their potential synergistic effects in these biological activities.
Author Contributions
E.-S.L. performed the experiments; E.-S.L. and C.-Y.H. analyzed the data; E.-S.L. and C.-Y.H. contributed to the study design and manuscript writing. All authors reviewed the results, contributed to the data interpretation, and approved the final version of the manuscript. All authors have read and agreed to the published version of the manuscript.
Funding
This research was supported by a grant from the National Science and Technology Council, Taiwan (NSTC 111-2622-E-025-002 to E.-S.L.).
Institutional Review Board Statement
Not applicable.
Informed Consent Statement
Not applicable.
Data Availability Statement
Not applicable.
Conflicts of Interest
The authors declare no conflict of interest.
References
- Roszkowski, S. Application of Polyphenols and Flavonoids in Oncological Therapy. Molecules 2023, 28, 4080. [Google Scholar] [CrossRef] [PubMed]
- Lobiuc, A.; Pavăl, N.E.; Mangalagiu, I.I.; Gheorghiță, R.; Teliban, G.C.; Amăriucăi-Mantu, D.; Stoleru, V. Future Antimicrobials: Natural and Functionalized Phenolics. Molecules 2023, 28, 1114. [Google Scholar] [CrossRef] [PubMed]
- Jakobušić Brala, C.; Karković Marković, A.; Kugić, A.; Torić, J.; Barbarić, M. Combination Chemotherapy with Selected Polyphenols in Preclinical and Clinical Studies-An Update Overview. Molecules 2023, 28, 3746. [Google Scholar] [CrossRef] [PubMed]
- Atanasov, A.G.; Zotchev, S.B.; Dirsch, V.M.; Supuran, C.T. Natural products in drug discovery: Advances and opportunities. Nat. Rev. Drug Discov. 2021, 20, 200–216. [Google Scholar] [CrossRef] [PubMed]
- WHO. Traditional medicine growing needs and potential. WHO Policy Perspect. Med. 2002, 2, 1–6. [Google Scholar]
- Rufino-Palomares, E.E.; Pérez-Jiménez, A.; García-Salguero, L.; Mokhtari, K.; Reyes-Zurita, F.J.; Peragón-Sánchez, J.; Lupiáñez, J.A. Nutraceutical Role of Polyphenols and Triterpenes Present in the Extracts of Fruits and Leaves of Olea europaea as Antioxidants, Anti-Infectives and Anticancer Agents on Healthy Growth. Molecules 2022, 27, 2341. [Google Scholar] [CrossRef]
- Khan, A.W.; Farooq, M.; Haseeb, M.; Choi, S. Role of Plant-Derived Active Constituents in Cancer Treatment and Their Mechanisms of Action. Cells 2022, 11, 1326. [Google Scholar] [CrossRef] [PubMed]
- Gligorov, J.; Richard, S. Breast cancer: Weekly paclitaxel—Still preferred first-line taxane for mBC. Nat. Rev. Clin. Oncol. 2015, 12, 508–509. [Google Scholar] [CrossRef]
- da Veiga Correia, V.T.; da Silva, P.R.; Ribeiro, C.M.S.; Ramos, A.; Mazzinghy, A.; Silva, V.D.M.; Júnior, A.H.O.; Nunes, B.V.; Vieira, A.L.S.; Ribeiro, L.V.; et al. An Integrative Review on the Main Flavonoids Found in Some Species of the Myrtaceae Family: Phytochemical Characterization, Health Benefits and Development of Products. Plants 2022, 11, 2796. [Google Scholar] [CrossRef]
- Gasparotto Junior, A.; de Souza, P.; Lívero, F. Plinia cauliflora (Mart.) Kausel: A comprehensive ethnopharmacological review of a genuinely Brazilian species. J. Ethnopharmacol. 2019, 245, 112169. [Google Scholar] [CrossRef]
- Chang, S.K.; Alasalvar, C.; Shahidi, F. Superfruits: Phytochemicals, antioxidant efficacies, and health effects—A comprehensive review. Crit. Rev. Food Sci. Nutr. 2019, 59, 1580–1604. [Google Scholar] [CrossRef]
- Inada, K.O.P.; Leite, I.B.; Martins, A.B.N.; Fialho, E.; Tomás-Barberán, F.A.; Perrone, D.; Monteiro, M. Jaboticaba berry: A comprehensive review on its polyphenol composition, health effects, metabolism, and the development of food products. Food Res. Int. 2021, 147, 110518. [Google Scholar] [CrossRef]
- Wu, S.B.; Dastmalchi, K.; Long, C.; Kennelly, E.J. Metabolite profiling of jaboticaba (Myrciaria cauliflora) and other dark-colored fruit juices. J. Agric. Food Chem. 2012, 60, 7513–7525. [Google Scholar] [CrossRef]
- Reguengo, L.M.; Nascimento, R.P.D.; Machado, A.; Marostica Junior, M.R. Signaling pathways and the potential anticarcinogenic effect of native Brazilian fruits on breast cancer. Food Res. Int. 2022, 155, 111117. [Google Scholar] [CrossRef]
- Machado, A.; Alves, M.D.R.; Nascimento, R.P.D.; Reguengo, L.M.; Marostica Junior, M.R. Antiproliferative effects and main molecular mechanisms of Brazilian native fruits and their by-products on lung cancer. Food Res. Int. 2022, 162, 111953. [Google Scholar] [CrossRef]
- Filho, A.V.; Avila, L.B.; Lacorte, D.H.; Martiny, T.R.; Rosseto, V.; Moraes, C.C.; Dotto, G.L.; Carreno, N.L.V.; da Rosa, G.S. Brazilian Agroindustrial Wastes as a Potential Resource of Bioative Compounds and Their Antimicrobial and Antioxidant Activities. Molecules 2022, 27, 6876. [Google Scholar] [CrossRef]
- Avila, L.B.; Barreto, E.R.C.; Souza, P.K.; Silva, B.Z.; Martiny, T.R.; Moraes, C.C.; Morais, M.M.; Raghavan, V.; Rosa, G.S.D. Carrageenan-Based Films Incorporated with Jaboticaba Peel Extract: An Innovative Material for Active Food Packaging. Molecules 2020, 25, 5563. [Google Scholar] [CrossRef] [PubMed]
- Meyer, R.R.; Laine, P.S. The single-stranded DNA-binding protein of Escherichia coli. Microbiol. Rev. 1990, 54, 342–380. [Google Scholar] [CrossRef] [PubMed]
- Bianco, P.R. The Biochemical Mechanism of Fork Regression in Prokaryotes and Eukaryotes-A Single Molecule Comparison. Int. J. Mol. Sci. 2022, 23, 8613. [Google Scholar] [CrossRef] [PubMed]
- Antony, E.; Lohman, T.M. Dynamics of E. coli single stranded DNA binding (SSB) protein-DNA complexes. Semin. Cell Dev. Biol. 2019, 86, 102–111. [Google Scholar] [CrossRef] [PubMed]
- Shereda, R.D.; Kozlov, A.G.; Lohman, T.M.; Cox, M.M.; Keck, J.L. SSB as an organizer/mobilizer of genome maintenance complexes. Crit. Rev. Biochem. Mol. Biol. 2008, 43, 289–318. [Google Scholar] [CrossRef] [PubMed]
- Heller, R.C.; Marians, K.J. Replisome assembly and the direct restart of stalled replication forks. Nat. Rev. Mol. Cell Biol. 2006, 7, 932–943. [Google Scholar] [CrossRef] [PubMed]
- Marians, K.J. PriA-directed replication fork restart in Escherichia coli. Trends Biochem. Sci. 2000, 25, 185–189. [Google Scholar] [CrossRef] [PubMed]
- Lohman, T.M.; Ferrari, M.E. Escherichia coli single-stranded DNA-binding protein: Multiple DNA-binding modes and cooperativities. Annu. Rev. Biochem. 1994, 63, 527–570. [Google Scholar] [CrossRef]
- Bianco, P.R. The mechanism of action of the SSB interactome reveals it is the first OB-fold family of genome guardians in prokaryotes. Protein Sci. 2021, 30, 1757–1775. [Google Scholar] [CrossRef]
- Voter, A.F.; Killoran, M.P.; Ananiev, G.E.; Wildman, S.A.; Hoffmann, F.M.; Keck, J.L. A High-Throughput Screening Strategy to Identify Inhibitors of SSB Protein-Protein Interactions in an Academic Screening Facility. SLAS Discov. 2017, 23, 94–101. [Google Scholar] [CrossRef]
- Glanzer, J.G.; Endres, J.L.; Byrne, B.M.; Liu, S.; Bayles, K.W.; Oakley, G.G. Identification of inhibitors for single-stranded DNA-binding proteins in eubacteria. J. Antimicrob. Chemother. 2016, 71, 3432–3440. [Google Scholar] [CrossRef]
- Marceau, A.H.; Bernstein, D.A.; Walsh, B.W.; Shapiro, W.; Simmons, L.A.; Keck, J.L. Protein interactions in genome maintenance as novel antibacterial targets. PLoS ONE 2013, 8, e58765. [Google Scholar] [CrossRef]
- Iftode, C.; Daniely, Y.; Borowiec, J.A. Replication protein A (RPA): The eukaryotic SSB. Crit. Rev. Biochem. Mol. Biol. 1999, 34, 141–180. [Google Scholar] [CrossRef]
- Byrne, B.M.; Oakley, G.G. Replication protein A, the laxative that keeps DNA regular: The importance of RPA phosphorylation in maintaining genome stability. Semin. Cell Dev. Biol. 2019, 86, 112–120. [Google Scholar] [CrossRef]
- Tommasi, R.; Brown, D.G.; Walkup, G.K.; Manchester, J.I.; Miller, A.A. ESKAPEing the labyrinth of antibacterial discovery. Nat. Rev. Drug Discov. 2015, 14, 529–542. [Google Scholar] [CrossRef]
- Darby, E.M.; Trampari, E.; Siasat, P.; Gaya, M.S.; Alav, I.; Webber, M.A.; Blair, J.M.A. Molecular mechanisms of antibiotic resistance revisited. Nat. Rev. Microbiol. 2023, 21, 280–295. [Google Scholar] [CrossRef] [PubMed]
- Du Toit, A. The cost of resistance. Nat. Rev. Microbiol. 2022, 20, 317. [Google Scholar] [CrossRef] [PubMed]
- Dong, N.; Yang, X.; Chan, E.W.; Zhang, R.; Chen, S. Klebsiella species: Taxonomy, hypervirulence and multidrug resistance. eBioMedicine 2022, 79, 103998. [Google Scholar] [CrossRef]
- Arato, V.; Raso, M.M.; Gasperini, G.; Berlanda Scorza, F.; Micoli, F. Prophylaxis and Treatment against Klebsiella pneumoniae: Current Insights on This Emerging Anti-Microbial Resistant Global Threat. Int. J. Mol. Sci. 2021, 22, 4042. [Google Scholar] [CrossRef] [PubMed]
- Franzolin, M.R.; Courrol, D.D.S.; Silva, F.R.O.; Courrol, L.C. Antimicrobial Activity of Silver and Gold Nanoparticles Prepared by Photoreduction Process with Leaves and Fruit Extracts of Plinia cauliflora and Punica granatum. Molecules 2022, 27, 6860. [Google Scholar] [CrossRef]
- Machado, G.H.A.; Marques, T.R.; de Carvalho, T.C.L.; Duarte, A.C.; de Oliveira, F.C.; Gonçalves, M.C.; Piccoli, R.H.; Corrêa, A.D. Antibacterial activity and in vivo wound healing potential of phenolic extracts from jaboticaba skin. Chem. Biol. Drug Des. 2018, 92, 1333–1343. [Google Scholar] [CrossRef]
- Mott, M.L.; Berger, J.M. DNA replication initiation: Mechanisms and regulation in bacteria. Nat. Rev. Microbiol. 2007, 5, 343–354. [Google Scholar] [CrossRef]
- Nižnanský, Ľ.; Osinová, D.; Kuruc, R.; Hengerics Szabó, A.; Szórádová, A.; Masár, M.; Nižnanská, Ž. Natural Taxanes: From Plant Composition to Human Pharmacology and Toxicity. Int. J. Mol. Sci. 2022, 23, 15619. [Google Scholar] [CrossRef]
- Merecz-Sadowska, A.; Sitarek, P.; Kowalczyk, T.; Zajdel, K.; Kucharska, E.; Zajdel, R. The Modulation of Melanogenesis in B16 Cells Upon Treatment with Plant Extracts and Isolated Plant Compounds. Molecules 2022, 27, 4360. [Google Scholar] [CrossRef]
- Balouiri, M.; Sadiki, M.; Ibnsouda, S.K. Methods for in vitro evaluating antimicrobial activity: A review. J. Pharm. Anal. 2016, 6, 71–79. [Google Scholar] [CrossRef] [PubMed]
- Singleton, V.L.; Orthofer, R.; Lamuela-Raventos, R.M. Analysis of total phenols and other oxidation substrates and antioxidants by means of folin-ciocalteu reagent. Methods Enzymol. 1999, 299, 152–178. [Google Scholar]
- Chang, C.C.; Yang, M.H.; Wen, H.M.; Chern, J.C. Estimation of total flavonoid content in propolis by two complementary colorimetric methods. J. Food Drug Anal. 2002, 10, 178–182. [Google Scholar]
- Brand-Williams, W.; Cuvelier, M.E.; Berset, C. Use of a free radical method to evaluate antioxidant activity. LWT Food Sci. Technol. 1995, 28, 25–30. [Google Scholar] [CrossRef]
- Strober, W. Trypan blue exclusion test of cell viability. Curr. Protoc. Immunol. 2001, 21, A-3B. [Google Scholar] [CrossRef]
- Larsson, R.; Nygren, P. A rapid fluorometric method for semiautomated determination of cytotoxicity and cellular proliferation of human tumor cell lines in microculture. Anticancer Res. 1989, 9, 1111–1119. [Google Scholar]
- Chen, M.H.; Yang, W.L.; Lin, K.T.; Liu, C.H.; Liu, Y.W.; Huang, K.W.; Chang, P.M.; Lai, J.M.; Hsu, C.N.; Chao, K.M.; et al. Gene expression-based chemical genomics identifies potential therapeutic drugs in hepatocellular carcinoma. PLoS ONE 2011, 6, e27186. [Google Scholar] [CrossRef]
- Liang, C.C.; Park, A.Y.; Guan, J.L. In vitro scratch assay: A convenient and inexpensive method for analysis of cell migration in vitro. Nat. Protoc. 2007, 2, 329–333. [Google Scholar] [CrossRef]
- Reynertson, K.A.; Yang, H.; Jiang, B.; Basile, M.J.; Kennelly, E.J. Quantitative analysis of antiradical phenolic constituents from fourteen edible Myrtaceae fruits. Food Chem. 2008, 109, 883–890. [Google Scholar] [CrossRef]
- Dahiya, D.; Nigam, P.S. Biotherapy Using Probiotics as Therapeutic Agents to Restore the Gut Microbiota to Relieve Gastrointestinal Tract Inflammation, IBD, IBS and Prevent Induction of Cancer. Int. J. Mol. Sci. 2023, 24, 5748. [Google Scholar] [CrossRef]
- Choi, J.R.; Kozalak, G.; di Bari, I.; Babar, Q.; Niknam, Z.; Rasmi, Y.; Yong, K.W. In Vitro Human Cancer Models for Biomedical Applications. Cancers 2022, 14, 2284. [Google Scholar] [CrossRef] [PubMed]
- Dzialo, M.; Mierziak, J.; Korzun, U.; Preisner, M.; Szopa, J.; Kulma, A. The Potential of Plant Phenolics in Prevention and Therapy of Skin Disorders. Int. J. Mol. Sci. 2016, 17, 160. [Google Scholar] [CrossRef] [PubMed]
- Xu, L.; Halma, M.T.J.; Wuite, G.J.L. Unravelling How Single-Stranded DNA Binding Protein Coordinates DNA Metabolism Using Single-Molecule Approaches. Int. J. Mol. Sci. 2023, 24, 2806. [Google Scholar] [CrossRef] [PubMed]
- Sugino, A.; Peebles, C.L.; Kreuzer, K.N.; Cozzarelli, N.R. Mechanism of action of nalidixic acid: Purification of Escherichia coli nalA gene product and its relationship to DNA gyrase and a novel nicking-closing enzyme. Proc. Natl. Acad. Sci. USA 1977, 74, 4767–4771. [Google Scholar] [CrossRef]
- Gellert, M.; O’Dea, M.H.; Itoh, T.; Tomizawa, J. Novobiocin and coumermycin inhibit DNA supercoiling catalyzed by DNA gyrase. Proc. Natl. Acad. Sci. USA 1976, 73, 4474–4478. [Google Scholar] [CrossRef]
- Pandey, P.; Khan, F.; Qari, H.A.; Oves, M. Rutin (Bioflavonoid) as Cell Signaling Pathway Modulator: Prospects in Treatment and Chemoprevention. Pharmaceuticals 2021, 14, 1069. [Google Scholar] [CrossRef]
- Richard, D.J.; Bolderson, E.; Cubeddu, L.; Wadsworth, R.I.; Savage, K.; Sharma, G.G.; Nicolette, M.L.; Tsvetanov, S.; McIlwraith, M.J.; Pandita, R.K.; et al. Single-stranded DNA-binding protein hSSB1 is critical for genomic stability. Nature 2008, 453, 677–681. [Google Scholar] [CrossRef]
- Raghunathan, S.; Kozlov, A.G.; Lohman, T.M.; Waksman, G. Structure of the DNA binding domain of E. coli SSB bound to ssDNA. Nat. Struct. Biol. 2000, 7, 648–652. [Google Scholar] [CrossRef]
- Dickey, T.H.; Altschuler, S.E.; Wuttke, D.S. Single-stranded DNA-binding proteins: Multiple domains for multiple functions. Structure 2013, 21, 1074–1084. [Google Scholar] [CrossRef]
- Su, X.C.; Wang, Y.; Yagi, H.; Shishmarev, D.; Mason, C.E.; Smith, P.J.; Vandevenne, M.; Dixon, N.E.; Otting, G. Bound or free: Interaction of the C-terminal domain of Escherichia coli single-stranded DNA-binding protein (SSB) with the tetrameric core of SSB. Biochemistry 2014, 53, 1925–1934. [Google Scholar] [CrossRef]
- Pinc, M.M.; Dalmagro, M.; da Cruz Alves Pereira, E.; Donadel, G.; Thomaz, R.T.; da Silva, C.; Macruz, P.D.; Jacomassi, E.; Gasparotto Junior, A.; Hoscheid, J.; et al. Extraction Methods, Chemical Characterization, and In Vitro Biological Activities of Plinia cauliflora (Mart.) Kausel Peels. Pharmaceuticals 2023, 16, 1173. [Google Scholar] [CrossRef] [PubMed]
- Wang, W.H.; Tyan, Y.C.; Chen, Z.S.; Lin, C.G.; Yang, M.H.; Yuan, S.S.; Tsai, W.C. Evaluation of the antioxidant activity and antiproliferative effect of the jaboticaba (Myrciaria cauliflora) seed extracts in oral carcinoma cells. Biomed. Res. Int. 2014, 2014, 185946. [Google Scholar] [CrossRef]
- Mendonça de Assis, P.; Cypriano Dutra, R.; Amarante, C.B.D.; Afonso Miranda Chaves, M.D.G.; Moreira, C.P.S.; Brandão, M.A.F.; Raposo, N.R.B. Plinia cauliflora (Mart.) Kausel: Toxicological assays, biological activities, and elemental analysis of organic compounds. Nat. Prod. Res. 2021, 35, 1727–1731. [Google Scholar] [CrossRef] [PubMed]


Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).