
High-Performance Microcomputing Tomography of Chick Embryo in the Early Stages of Embryogenesis

 ,
,  , and
, and 
Abstract
:1. Introduction
2. Materials and Methods
2.1. Chemicals
2.2. Embryo Preparation
2.3. Stain
2.4. MicroCT Imaging Systems
2.5. Histological Preparation
2.6. Statistical Data Processing
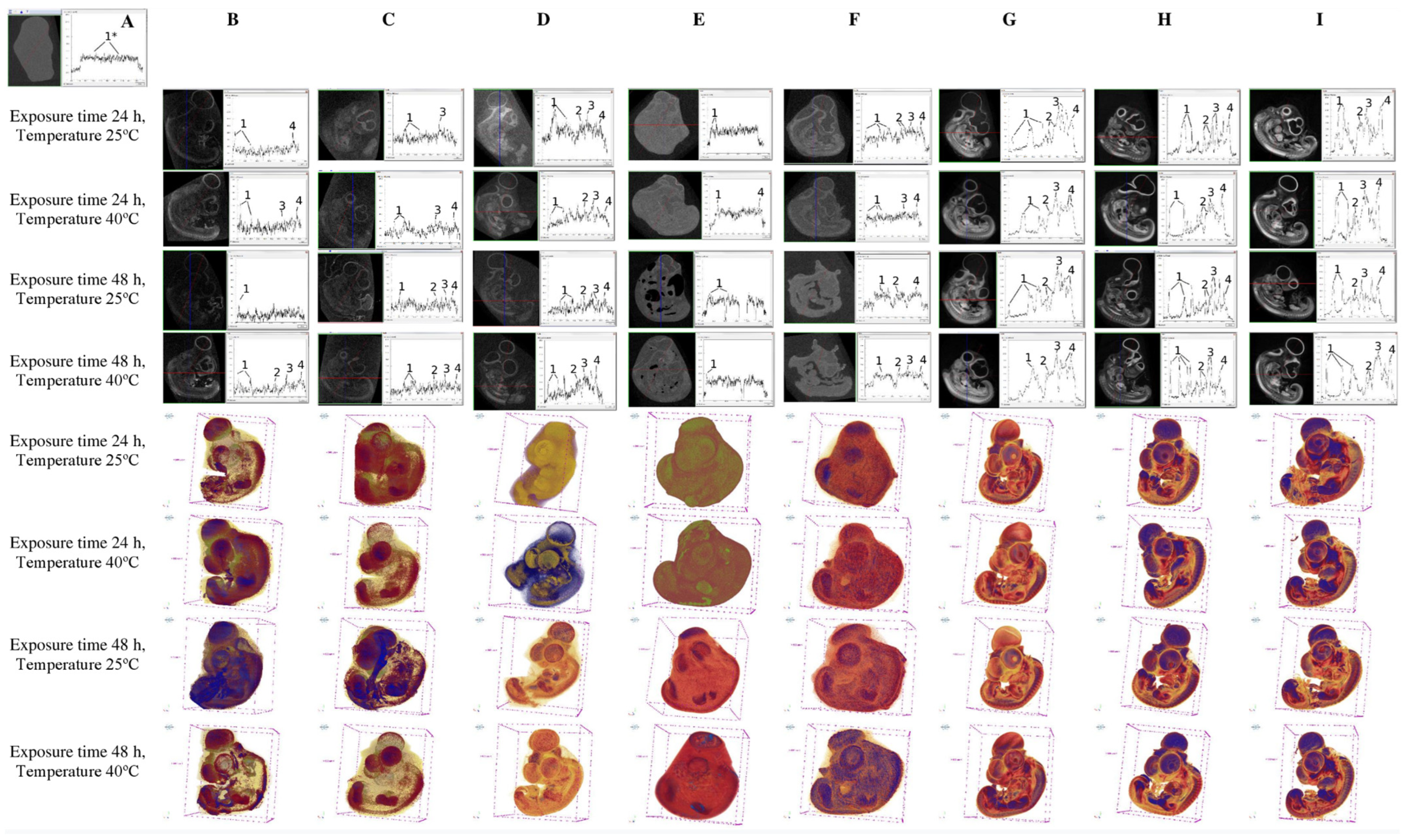
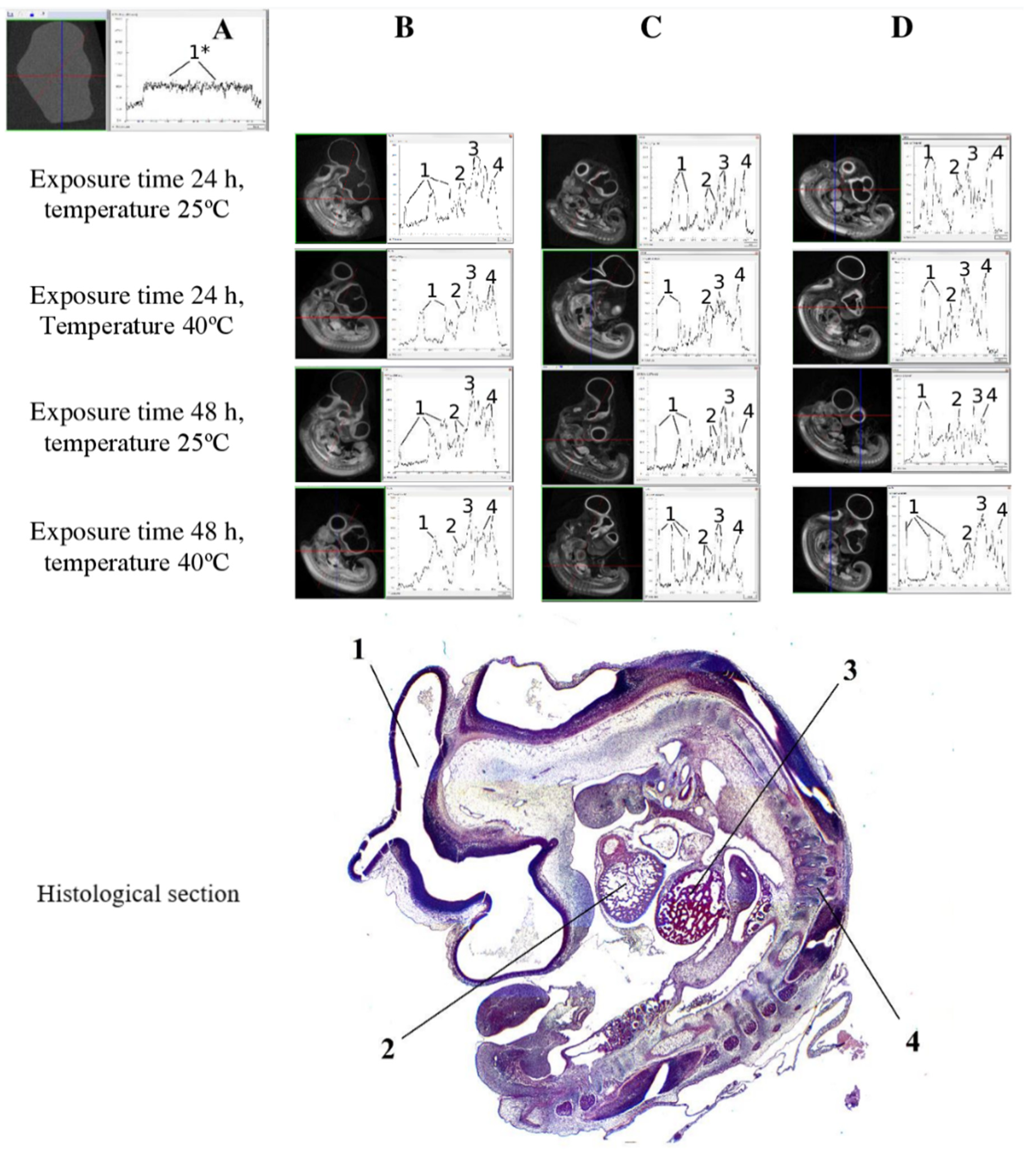
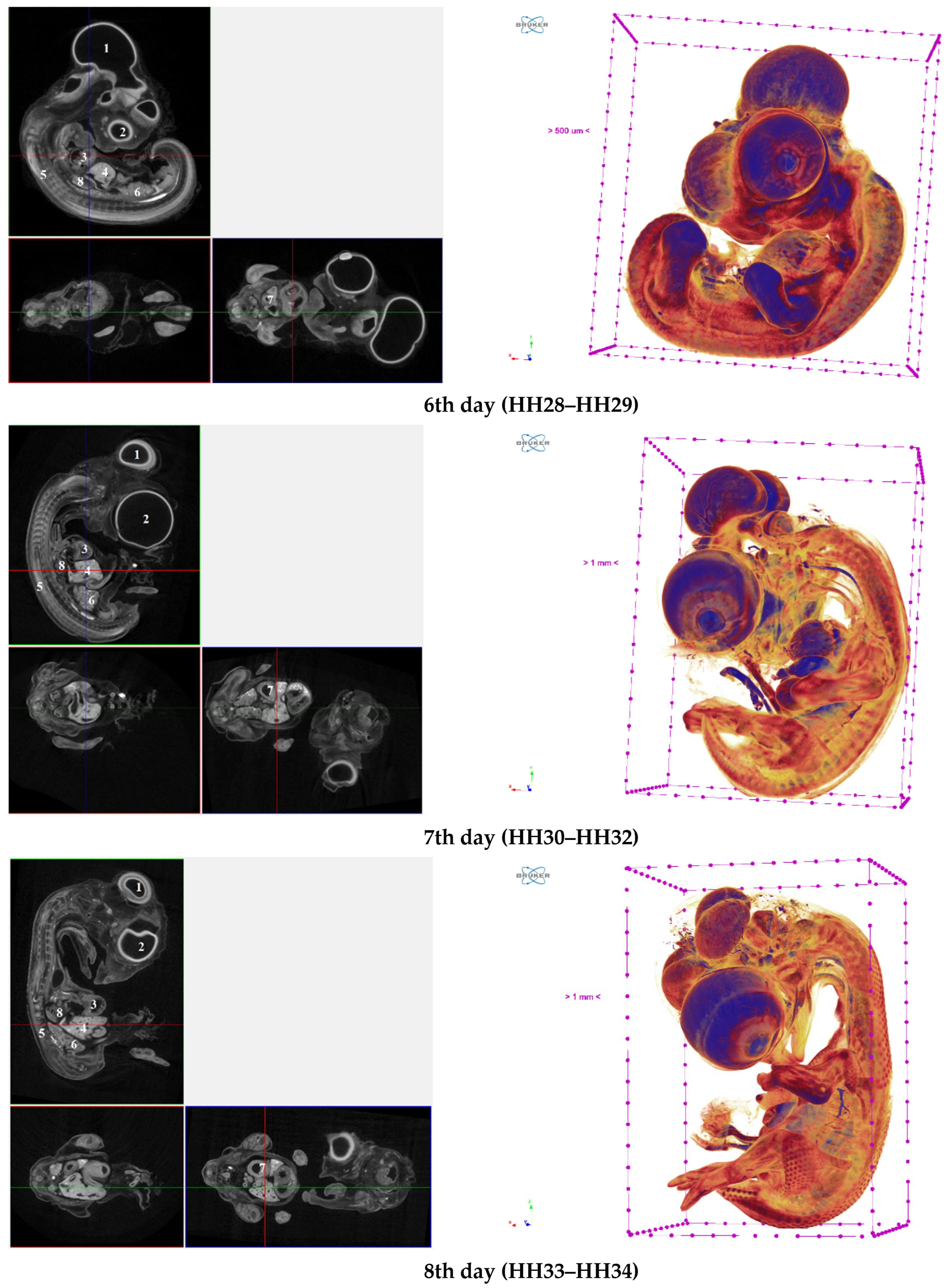
3. Results
4. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Butler, K.S.; Brinker, C.J.; Leong, H.S. Bridging the In Vitro to In Vivo Gap: Using the Chick Embryo Model to Accelerate Nanoparticle Validation and Qualification for In Vivo Studies. ACS Nano 2022, 16, 19626–19650. [Google Scholar] [CrossRef] [PubMed]
- Ghimire, S.; Zhang, X.; Zhang, J.; Wu, C. Use of Chicken Embryo Model in Toxicity Studies of Endocrine-Disrupting Chemicals and Nanoparticles. Chem. Res. Toxicol. 2022, 35, 550–568. [Google Scholar] [CrossRef] [PubMed]
- Ribatti, D.; Annese, T. Chick embryo in experimental embryology and more. Pathol.-Res. Pract. 2023, 245, 154478. [Google Scholar] [CrossRef] [PubMed]
- Pilkington, V.; Pepperrell, T.; Hill, A. A review of the safety of favipiravir—A potential treatment in the COVID-19 pandemic? J. Virus Erad. 2020, 6, 45–51. [Google Scholar] [CrossRef] [PubMed]
- Fernandes, C.H.; Sabongi, R.G.; Santos, J.B.G.D. COVID-19 and upper limb anomalies in newborns: A reason for concern? Acta Ortop. Bras. 2022, 30, e252308. [Google Scholar] [CrossRef]
- Sommerfeld, S.; Mundim, A.V.; Silva, R.R.; Queiroz, J.S.; Rios, M.P.; Notário, F.O.; Medeiros Ronchi, A.A.; Beletti, M.E.; Franco, R.R.; Espindola, F.S.; et al. Physiological Changes in Chicken Embryos Inoculated with Drugs and Viruses Highlight the Need for More Standardization of This Animal Model. Animals 2022, 12, 1156. [Google Scholar] [CrossRef]
- Choi, S.; Kim, K.H.; Kim, S.K.; Wang, K.C.; Lee, J.Y. Three-dimensional visualization of secondary neurulation in chick embryos using microCT. Dev. Dyn. 2022, 251, 885–896. [Google Scholar] [CrossRef]
- Scully, D.M.; Larina, I.V. Mouse embryo phenotyping with optical coherence tomography. Front. Cell Dev. Biol. 2022, 10, 1000237. [Google Scholar] [CrossRef]
- Ban, S.; Cho, N.H.; Min, E.; Bae, J.K.; Ahn, Y.; Shin, S.; Park, S.; Lee, Y.; Jung, W. Label-Free Optical Projection Tomography for Quantitative Three-Dimensional Anatomy of Mouse Embryo. J. Biophotonics 2019, 12, e201800481. [Google Scholar] [CrossRef]
- Bevilacqua, C.; Gomez, J.M.; Fiúza, U.-M.; Chan, C.J.; Wang, L.; Hambura, S.; Eguren, M.; Ellenberg, J.; Diz-Muñoz, A.; Leptin, M.; et al. High-resolution line-scan Brillouin microscopy for live imaging of mechanical properties during embryo development. Nat. Methods 2022, 20, 755–760. [Google Scholar] [CrossRef]
- Chen, L.; Wang, Z.; Fu, X.; Wang, S.; Feng, Y.; Coudyzer, W.; Wu, S.; Zhang, H.; Chai, Z.; Li, Y.; et al. Dynamic 3D morphology of chick embryos and allantois depicted nondestructively by 3.0T clinical magnetic resonance imaging. Poult. Sci. 2023, 102, 102902. [Google Scholar] [CrossRef]
- Griffin, C.T.; Botelho, J.F.; Hanson, M.; Fabbri, M.; Smith-Paredes, D.; Carney, R.M.; Norell, M.A.; Egawa, S.; Gatesy, S.M.; Rowe, T.B.; et al. The developing bird pelvis passes through ancestral dinosaurian conditions. Nature 2022, 608, 346–352. [Google Scholar] [CrossRef]
- Raguin, E.; Weinkamer, R.; Schmitt, C.; Curcuraci, L.; Fratzl, P. Logistics of Bone Mineralization in the Chick Embryo Studied by 3D Cryo FIB-SEM Imaging. Adv. Sci. 2023, 10, 2301231. [Google Scholar] [CrossRef]
- Handschuh, S.; Okada, C.T.C.; Walter, I.; Aurich, C.; Glösmann, M. An optimized workflow for microCT imaging of formalin-fixed and paraffin-embedded (FFPE) early equine embryos. Anat. Histol. Embryol. 2022, 51, 611–623. [Google Scholar] [CrossRef]
- Matula, J.; Polakova, V.; Šalplachta, J.; Tesařová, M.; Zikmund, T.; Kaucká, M.; Adameyko, I.; Kaiser, J. Resolving complex cartilage structures in developmental biology via deep learning-based automatic segmentation of X-ray computed microtomography images. Sci. Rep. 2022, 12, 8728. [Google Scholar] [CrossRef]
- Osterwalder, M.; Bolten, J.S.; Rodgers, G.; Humbel, M.; Schulz, G.; Tanner, C.; Huwyler, J.; Müller, B. Hard X-ray microtomography of Zebrafish larvae. Int. Conf. X-Ray Lasers 2021, 2020, 308–322. [Google Scholar] [CrossRef]
- Metscher, B.D. A Simple Nuclear Contrast Staining Method for MicroCT-Based 3D Histology Using Lead(II) Acetate. bioRxiv 2020, 238, 1036–1041. [Google Scholar] [CrossRef]
- Schoborg, T.A.; Smith, S.L.; Smith, L.N.; Morris, H.D.; Rusan, N.M. Micro-computed tomography as a platform for exploring Drosophila development. Development 2019, 146, 23. [Google Scholar] [CrossRef]
- Metscher, B.D. MicroCT for comparative morphology: Simple staining methods allow high-contrast 3D imaging of diverse non-mineralized animal tissues. BMC Physiol. 2009, 9, 11. [Google Scholar] [CrossRef]
- Silva, J.M.d.S.e.; Zanette, I.; Noël, P.B.; Cardoso, M.B.; Kimm, M.A.; Pfeiffer, F. Three-dimensional non-destructive soft-tissue visualization with X-ray staining micro-tomography. Sci. Rep. 2015, 5, 14088. [Google Scholar] [CrossRef]
- Orhan, K. Micro-Computed Tomography (Micro-CT) in Medicine and Engineering; Springer: Berlin/Heidelberg, Germany, 2020. [Google Scholar]
- Astanina, E.; Petrillo, S.; Genova, T.; Mussano, F.; Bussolino, F. E15.5 Mouse Embryo Micro-CT Using a Bruker Skyscan 1172 Micro-CT. Bio Protoc. 2023, 13, e4662. [Google Scholar] [CrossRef]
- Handschuh, S.; Glösmann, M. Mouse embryo phenotyping using X-ray microCT. Front. Cell Dev. Biol. 2022, 10, 949184. [Google Scholar] [CrossRef]
- Ermakova, O.; Orsini, T.; Gambadoro, A.; Chiani, F.; Tocchini-Valentini, G.P. Three-dimensional microCT imaging of murine embryonic development from immediate post-implantation to organogenesis: Application for phenotyping analysis of early embryonic lethality in mutant animals. Mamm. Genome 2017, 29, 245–259. [Google Scholar] [CrossRef]
- Dullin, C.; Ufartes, R.; Larsson, E.; Martin, S.; Lazzarini, M.; Tromba, G.; Missbach-Guentner, J.; Pinkert-Leetsch, D.; Katschinski, D.M.; Alves, F. μCT of ex-vivo stained mouse hearts and embryos enables a precise match between 3D virtual histology, classical histology and immunochemistry. PLoS ONE 2017, 12, e0170597. [Google Scholar] [CrossRef]
- Kim, J.S.; Min, J.; Recknagel, A.K.; Riccio, M.L.; Butcher, J.T. Quantitative Three-Dimensional Analysis of Embryonic Chick Morphogenesis via Microcomputed Tomography. Anat. Rec. Adv. Integr. Anat. Evol. Biol. 2011, 294, 1–10. [Google Scholar] [CrossRef]
- Lesciotto, K.M.; Motch Perrine, S.M.; Kawasaki, M.; Stecko, T.; Ryan, T.M.; Kawasaki, K.; Richtsmeier, J.T. Phosphotungstic acid-enhanced microCT: Optimized protocols for embryonic and early postnatal mice. Dev. Dyn. 2019, 249, 573–585. [Google Scholar] [CrossRef]
- Bjørnstad, S.; Austdal, L.P.E.; Roald, B.; Glover, J.C.; Paulsen, R.E. Cracking the Egg: Potential of the Developing Chicken as a Model System for Nonclinical Safety Studies of Pharmaceuticals. J. Pharmacol. Exp. Ther. 2015, 355, 386–396. [Google Scholar] [CrossRef]
- Underwood, W.; Anthony, R. AVMA Guidelines for the Euthanasia of Animals: 2020 Edition. Available online: https://www.avma.org/sites/default/files/2020-02/Guidelines-on-Euthanasia-2020.pdf (accessed on 19 August 2023).
- Kraus, N.; Placzek, F.; Metscher, B.D. OCT Meets micro-CT: A Subject-Specific Correlative Multimodal Imaging Workflow for Early Chick Heart Development Modeling. J. Cardiovasc. Dev. Dis. 2022, 9, 379. [Google Scholar] [CrossRef]
- Descamps, E.; Sochacka, A.; Kegel, B.D.; Loo, D.V.; Hoorebeke, L.V.; Adriaens, D. Soft tissue discrimination with contrast agents using micro-CT scanning. Belg. J. Zool. 2020, 144, 101–113. [Google Scholar] [CrossRef]
- Busse, M.; Müller, M.; Kimm, M.A.; Ferstl, S.; Allner, S.; Achterhold, K.; Herzen, J.; Pfeiffer, F. 3D Imaging of Soft-Tissue Samples Using an X-ray Specific Staining Method and Nanoscopic Computed Tomography. J. Vis. Exp. JoVE 2019, 152, e60251. [Google Scholar] [CrossRef]
- Rzhepakovsky, I.V.; Anusha Siddiqui, S.; Avanesyan, S.; Benlidayi, M.E.; Dhingra, K.; Dolgalev, A.A.; Enukashvily, N.I.; Fritsch, T.; Heinz, V.; Kochergin, S.; et al. Anti-arthritic effect of chicken embryo tissue hydrolyzate against adjuvant arthritis in rats (X-ray microtomographic and histopathological analysis). Food Sci. Nutr. 2021, 9, 5648–5669. [Google Scholar] [CrossRef] [PubMed]
- Nagdalian, A.A.; Rzhepakovsky, I.V.; Siddiqui, S.A.; Piskov, S.I.; Oboturova, N.P.; Timchenko, L.D.; Lodygin, A.D.; Blinov, A.V.; Ibrahim, S.A. Analysis of the content of mechanically separated poultry meat in sausage using computing microtomography. J. Food Compos. Anal. 2021, 100, 103918. [Google Scholar] [CrossRef]
- Tahara, R.; Larsson, H.C.E. Quantitative analysis of microscopic X-ray computed tomography imaging: Japanese quail embryonic soft tissues with iodine staining. J. Anat. 2013, 223, 297–310. [Google Scholar] [CrossRef] [PubMed]
- Dawood, Y.; Hagoort, J.; Siadari, B.A.; Ruijter, J.M.; Gunst, Q.D.; Lobé, N.H.J.; Strijkers, G.J.; de Bakker, B.S.; van den Hoff, M.J.B. Reducing soft-tissue shrinkage artefacts caused by staining with Lugol’s solution. Sci. Rep. 2021, 11, 19781. [Google Scholar] [CrossRef] [PubMed]
- Heimel, P.; Swiadek, N.; Slezak, P.; Kerbl, M.; Schneider, C.; Nürnberger, S.; Redl, H.; Teuschl, A.H.; Hercher, D. Iodine-Enhanced Micro-CT Imaging of Soft Tissue on the Example of Peripheral Nerve Regeneration. Contrast Media Mol. Imaging 2018, 2019, 7483745. [Google Scholar] [CrossRef] [PubMed]
- Zhou, Z.; Chen, Z.; Shan, J.; Ma, W.; Li, L.; Zu, J.; Xu, J. Monitoring brain development of chick embryos in vivo using 3.0 T MRI: Subdivision volume change and preliminary structural quantification using DTI. BMC Dev. Biol. 2015, 15, 29. [Google Scholar] [CrossRef]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| # | Stain | Exposure Time, h | Temperature, °C | Visualized Volume Relative to no Stain, mm3 (%) |
|---|---|---|---|---|
| 1 | No stain (70% ethanol) | - | - | 196.0 ± 4.0 (100) |
| 2 | Silver proteinate, 1% (70% ethanol) | 24 | 25 | 147.3 ± 3.1 (75.2) |
| 3 | Silver proteinate, 1% (70% ethanol) | 24 | 40 | 91.8 ± 3.9 (48.8) |
| 4 | Silver proteinate, 1% (70% ethanol) | 48 | 25 | 160.0 ± 4.5 (61.2) |
| 5 | Silver proteinate, 1% (70% ethanol) | 48 | 40 | 84.3 ± 3.1 (43.0) |
| 6 | Eosin, 1% (70% ethanol) | 24 | 25 | 90.1 ± 3.2 (46.0) |
| 7 | Eosin, 1% (70% ethanol) | 24 | 40 | 93.7 ± 3.5 (47.8) |
| 8 | Eosin, 1% (70% ethanol) | 48 | 25 | 104.7 ± 4.2 (53.4) |
| 9 | Eosin, 1% (70% ethanol) | 48 | 40 | 81.2 ± 2.9 (41.4) |
| 10 | I2KI, 1% (70% ethanol) | 24 | 25 | 106.6 ± 4.1 (54.4) |
| 11 | I2KI, 1% (70% ethanol) | 24 | 40 | 92.8 ± 3.8 (47.4) |
| 12 | I2KI, 1% (70% ethanol) | 48 | 25 | 98.0 ± 3.6 (50.0) |
| 13 | I2KI, 1% (70% ethanol) | 48 | 40 | 101.3 ± 4.1 (51.7) |
| 14 | Silver proteinate, 5% (70% ethanol) | 24 | 25 | 128.1 ± 5.0 (65.4) |
| 15 | Silver proteinate, 5% (70% ethanol) | 24 | 40 | 109.4 ± 4.9 (55.8) |
| 16 | Silver proteinate, 5% (70% ethanol) | 48 | 25 | 166.2 ± 6.1 (84.8) |
| 17 | Silver proteinate, 5% (70% ethanol) | 48 | 40 | 213.3 ± 7.2 (108.8) |
| 18 | Eosin, 5% (70% ethanol) | 24 | 25 | 66.0 ± 3.3 (33.7) |
| 19 | Eosin, 5% (70% ethanol) | 24 | 40 | 53.8 ± 3.1 (27.5) |
| 20 | Eosin, 5% (70% ethanol) | 48 | 25 | 34.7 ± 2.1 (17.7) |
| 21 | Eosin, 5% (70% ethanol) | 48 | 40 | 32.1 ± 2.8 (16.4) |
| 22 | I2KI, 3% (70% ethanol) | 24 | 25 | 55.3 ± 3.1 (28,2) |
| 23 | I2KI, 3% (70% ethanol) | 24 | 40 | 47.0 ± 2.2 (24,0) |
| 24 | I2KI, 3% (70% ethanol) | 48 | 25 | 51.6 ± 2.1 (26.3) |
| 25 | I2KI, 3% (70% ethanol) | 48 | 40 | 45.0 ± 2.5 (23.0) |
| 26 | PMA, 1% (70% ethanol) | 24 | 25 | 146.0 ± 4.1 (74.5) |
| 27 | PMA, 1% (70% ethanol) | 24 | 40 | 152.0 ± 5.6 (77.6) |
| 28 | PMA, 1% (70% ethanol) | 48 | 25 | 98.0 ± 3.1 (50.0) |
| 29 | PMA, 1% (70% ethanol) | 48 | 40 | 116.3 ± 6.2 (59.3) |
| 30 | PTA, 1% (70% ethanol) | 24 | 25 | 105.2 ± 6.5 (53.7) |
| 31 | PTA, 1% (70% ethanol) | 24 | 40 | 105.3 ± 5.1 (53.7) |
| 32 | PTA, 1% (70% ethanol) | 48 | 25 | 92.4 ± 4.1 (47.1) |
| 33 | PTA, 1% (70% ethanol) | 48 | 40 | 92.0 ± 3.2 (46.9) |
| Stain | Exposure Time, h | Tempe-Rature, °C | Filter | X-ray Voltage, kV | X-ray Current, μA | Scans Averaged | Voxel Size (μm) | Ring Artifact Correction | Beam Hardening Correction (%) | Minimum for CS to Image Conversion | Maximum for CS to Image Conversion |
|---|---|---|---|---|---|---|---|---|---|---|---|
| No stain (70% ethanol) | - | - | Al 0.2 mm | 45 | 550 | 3 | 8.87 | 10 | 20 | 0.001 | 0.03 |
| Silver proteinate, 1% (70% ethanol) | 24; 48 | 25; 40 | no filter; Al 0.2 mm | 40; 45 | 550; 600 | 3 | 8.87 | 10 | 0 | 0.006–0.01 | 0.02–0.03 |
| Eosin, 1% (70% ethanol) | 24; 48 | 25; 40 | no filter | 40 | 600 | 3 | 8.87 | 10 | 0 | 0.01 | 0.03 |
| I2KI, 1% (70% ethanol) | 24; 48 | 25; 40 | Al 0.2 mm; Al 0.5 mm | 45; 50 | 500; 550 | 3 | 8.87 | 10 | 30 | 0.002–0.005 | 0.017–0.025 |
| Silver proteinate, 5% (70% ethanol) | 24; 48 | 25; 40 | no filter; Al 0.2 mm | 40; 45 | 550; 600 | 3 | 8.87 | 10; 20 | 21 | 0.002–0.004 | 0.025–0.035 |
| Eosin, 5% (70% ethanol) | 24; 48 | 25; 40 | Al 0.2 mm | 45 | 550 | 3 | 8.87 | 10 | 21 | 0.002–0.003 | 0.025 |
| I2KI, 3% (70% ethanol) | 24; 48 | 25; 40 | Al 0.5 mm; Al 1 mm | 50; 65 | 380; 500 | 3 | 8.87 | 10 | 41 | 0.002–0.003 | 0.06–0.09 |
| PMA, 1% (70% ethanol) | 24; 48 | 25; 40 | Al 0.5 mm; Al 1 mm | 50; 65 | 380; 500 | 3 | 8.87 | 10 | 41 | 0.001–0.002 | 0.07–0.09 |
| PTA, 1% (70% ethanol) | 24; 48 | 25; 40 | Al 1 mm | 65 | 380 | 3 | 8.87 | 10 | 41 | 0.001 | 0.075–0.1 |
| Stain, Exposure Time, Temperature | X-ray Density, HU | ||||
|---|---|---|---|---|---|
| The Chick Embryo | Brain | Heart | Liver | Sclerotome (Neural Canal) | |
| I2KI (3%), 24 h, 25 °C | 1089.1 ± 50 | 1526.4 ± 75 | 3503.4 ± 129 | 6972.4 ± 264 | 2991.6 ± 140 |
| I2KI (3%), 24 h, 40 °C | 839.6 ± 47 | 957.3 ± 43 | 3152.4 ± 166 | 6979.3 ± 348 | 3347.9 ± 273 |
| I2KI (3%), 48 h, 25 °C | 2845.1 ± 235 | 3307.6 ± 201 | 7228.8 ± 301 | 15,525.0 ± 699 | 8181.1 ± 465 |
| I2KI (3%), 48 h, 40 °C | 2483.8 ± 178 | 2084.4 ± 156 | 6570.1 ± 435 | 13,575.0 ± 621 | 7785.9 ± 289 |
| PMA (1%), 24 h, 25 °C | 384.9 ± 29 | 759.8 ± 56 | 1629.0 ± 91 | 3603.6 ± 188 | 1680.4 ± 110 |
| PMA (1%), 24 h, 40 °C | 635.1 ± 42 | 1349.2 ± 72 | 2120.9 ± 99 | 4565.9 ± 245 | 2080.9 ± 130 |
| PMA (1%), 48 h, 25 °C | 472.8 ± 29 | 810.5 ± 44 | 1825.1 ± 102 | 4138.9 ± 203 | 1907.0 ± 98 |
| PMA (1%), 48 h, 40 °C | 828.7 ± 37 | 1544.0 ± 128 | 2953.4 ± 144 | 5685.5 ± 276 | 2344.3 ± 175 |
| PTA (1%), 24 h, 25 °C | 1652.7 ± 95 | 1962.4 ± 108 | 3992.9 ± 264 | 8030.0 ± 301 | 4716.8 ± 234 |
| PTA (1%), 24 h, 40 °C | 2065.5 ± 116 | 2395.9 ± 148 | 5714.7 ± 176 | 11,255.0 ± 559 | 5744.5 ± 212 |
| PTA (1%), 48 h, 25 °C | 1790.0 ± 102 | 2258.2 ± 190 | 4026.8 ± 134 | 8611.1 ± 301 | 5653.6 ± 267 |
| PTA (1%), 48 h, 40 °C | 2362.3 ± 157 | 2626.2 ± 114 | 6185.8 ± 312 | 10,816.0 ± 489 | 5716.2 ± 229 |
| Structures | X-ray Density, HU | ||||
|---|---|---|---|---|---|
| 4 Days (HH22–HH24) | 5 Days (HH25–HH27) | 6 Days (HH28–HH29) | 7 Days (HH30–HH32) | 8 Days (HH33–HH34) | |
| The chick embryo | 3115.9 ± 111 | 2065.5 ± 116 | 2447.7 ± 125 | 1517.2 ± 84 | 1807.9 ± 103 |
| Brain | 5284.4 ± 202 | 2395.9 ± 148 | 2752.9 ± 112 | 2393.7 ± 135 | 3238.3 ± 193 |
| Eye (left) | 7514,4 ± 257 | 3780.2 ± 180 | 3255.0 ± 177 | 1693.4 ± 95 | 1897.2 ± 98 |
| Heart | 4923.2 ± 226 | 5714.7 ± 176 | 5401.1 ± 245 | 3513.8 ± 164 | 3563.9 ± 198 |
| Liver | 9244.0 ± 378 | 11,255.0 ± 559 | 12,155.0 ± 589 | 5763.6 ± 243 | 6154.7 ± 302 |
| Sclerotome (spine and neural canal) | 6978.4 ± 320 | 5744.5 ± 212 | 6339.1 ± 369 | 2658.6 ± 139 | 2775.2 ± 173 |
| Mesonephros (left) | 10,091.0 ± 461 | 8785.3 ± 342 | 9659.7 ± 401 | 4302.4 ± 157 | 4382.3 ± 189 |
| Stomach | 13,873.0 ± 598 | 10,064.0 ± 497 | 9785.7 ± 389 | 4233.1 ± 187 | 4298.8 ± 194 |
| Lungs (left) | not defined | not defined | 10,038 ± 450 | 3624.5 ± 143 | 2911.0 ± 157 |
| Structures | Visualized Volume, mm3 | ||||
|---|---|---|---|---|---|
| 4 Days, (HH22–HH24) | 5 Days, (HH25–HH27) | 6 Days, (HH28–HH29) | 7 Days, (HH30–HH32) | 8 Days, (HH33–HH34) | |
| The chick embryo | 17.7 ± 1.6 | 105.3 ± 5.1 | 137.6 ± 8.9 | 367.6 ± 23.2 | 571.3 ± 38.0 |
| Brain | 3.7 ± 0.2 | 22.3 ± 1.9 | 27.9 ± 1.7 | 54.5 ± 3.4 | 80.8 ± 4.7 |
| Eye (left) | 0.31 ± 0.04 | 3.94 ± 0.5 | 6.54 ± 0.5 | 33.3 ± 1.9 | 66.0 ± 3.2 |
| Heart | 0.65 ± 0.05 | 2.33 ± 0.1 | 2.91 ± 0.2 | 6.6 ± 0.5 | 10.5 ± 0.9 |
| Liver | 0.08 ± 0.006 | 0.74 ± 0.04 | 1.1 ± 0.05 | 5.1 ± 0.3 | 10.1 ± 0.8 |
| Mesonephros (left) | 0.06 ± 0.004 | 0.44 ± 0.02 | 0.74 ± 0.03 | 2.3 ± 0.1 | 3.27 ± 0.2 |
| Stomach | 0.06 ± 0.005 | 0.44 ± 0.01 | 0.75 ± 0.03 | 2.24 ± 0.1 | 6.23 ± 0.3 |
| Lungs (left) | - | - | 0.09 ± 0.006 | 0.36 ± 0.03 | 1.46 ± 0.06 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Rzhepakovsky, I.; Piskov, S.; Avanesyan, S.; Shakhbanov, M.; Sizonenko, M.; Timchenko, L.; Shariati, M.A.; Rebezov, M.; Nagdalian, A. High-Performance Microcomputing Tomography of Chick Embryo in the Early Stages of Embryogenesis. Appl. Sci. 2023, 13, 10642. https://doi.org/10.3390/app131910642
Rzhepakovsky I, Piskov S, Avanesyan S, Shakhbanov M, Sizonenko M, Timchenko L, Shariati MA, Rebezov M, Nagdalian A. High-Performance Microcomputing Tomography of Chick Embryo in the Early Stages of Embryogenesis. Applied Sciences. 2023; 13(19):10642. https://doi.org/10.3390/app131910642
Chicago/Turabian StyleRzhepakovsky, Igor, Sergei Piskov, Svetlana Avanesyan, Magomed Shakhbanov, Marina Sizonenko, Lyudmila Timchenko, Mohammad Ali Shariati, Maksim Rebezov, and Andrey Nagdalian. 2023. "High-Performance Microcomputing Tomography of Chick Embryo in the Early Stages of Embryogenesis" Applied Sciences 13, no. 19: 10642. https://doi.org/10.3390/app131910642
APA StyleRzhepakovsky, I., Piskov, S., Avanesyan, S., Shakhbanov, M., Sizonenko, M., Timchenko, L., Shariati, M. A., Rebezov, M., & Nagdalian, A. (2023). High-Performance Microcomputing Tomography of Chick Embryo in the Early Stages of Embryogenesis. Applied Sciences, 13(19), 10642. https://doi.org/10.3390/app131910642