

A Spatial Location Representation Method Incorporating Boundary Information

Abstract
1. Introduction
- (1)
- Inspired by the mammalian spatial cognitive mechanism, a new boundary cell model is proposed to establish boundary cell activity states in multiple scenarios by the mutual excitation and the inhibition of the direction-aware and distance-aware information that is acquired by mobile robots. The boundary cell model proposed in this paper can encode the boundary information in the environment and supplement the lack of environmental boundary perceptual information with path integration.
- (2)
- The physiological phenomena indicate that the environmental boundary information can be used as the supplementary information of grid cells. The method in this paper maps the boundary cell response values to the input layer of LAHN, generates grid cells by LAHN learning rules, and uses the boundary cell response values to correct the grid cell distribution pattern, such that the grid cell firing response and distribution that is activated by the method are more consistent with the physiological characteristics.
- (3)
- According to the problem that the mobile robot runs for a long time in an unknown environment, when the mobile robot reaches the boundary cell excitation zone, the accumulated error caused by the long running time of the position cell is corrected by the activated boundary cells, such that only one place cell responds to the current position at each time in order to improve the location accuracy of the system.
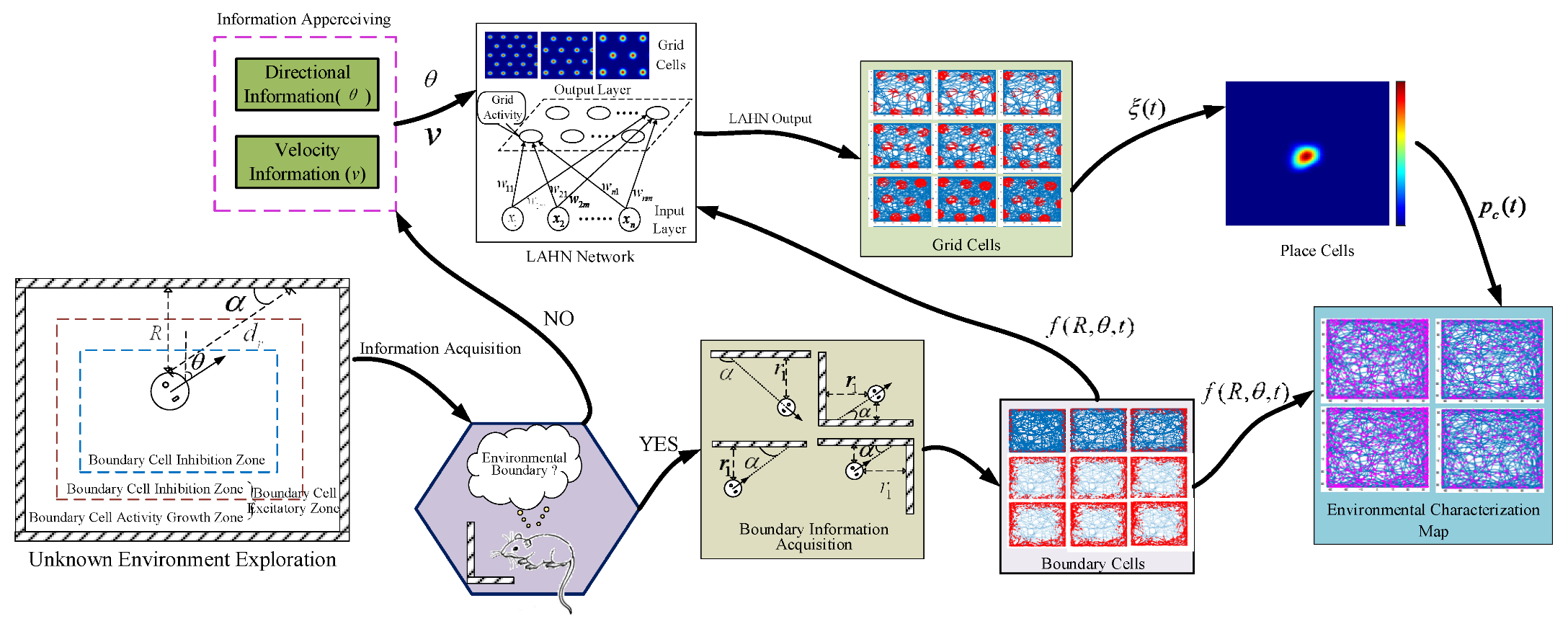
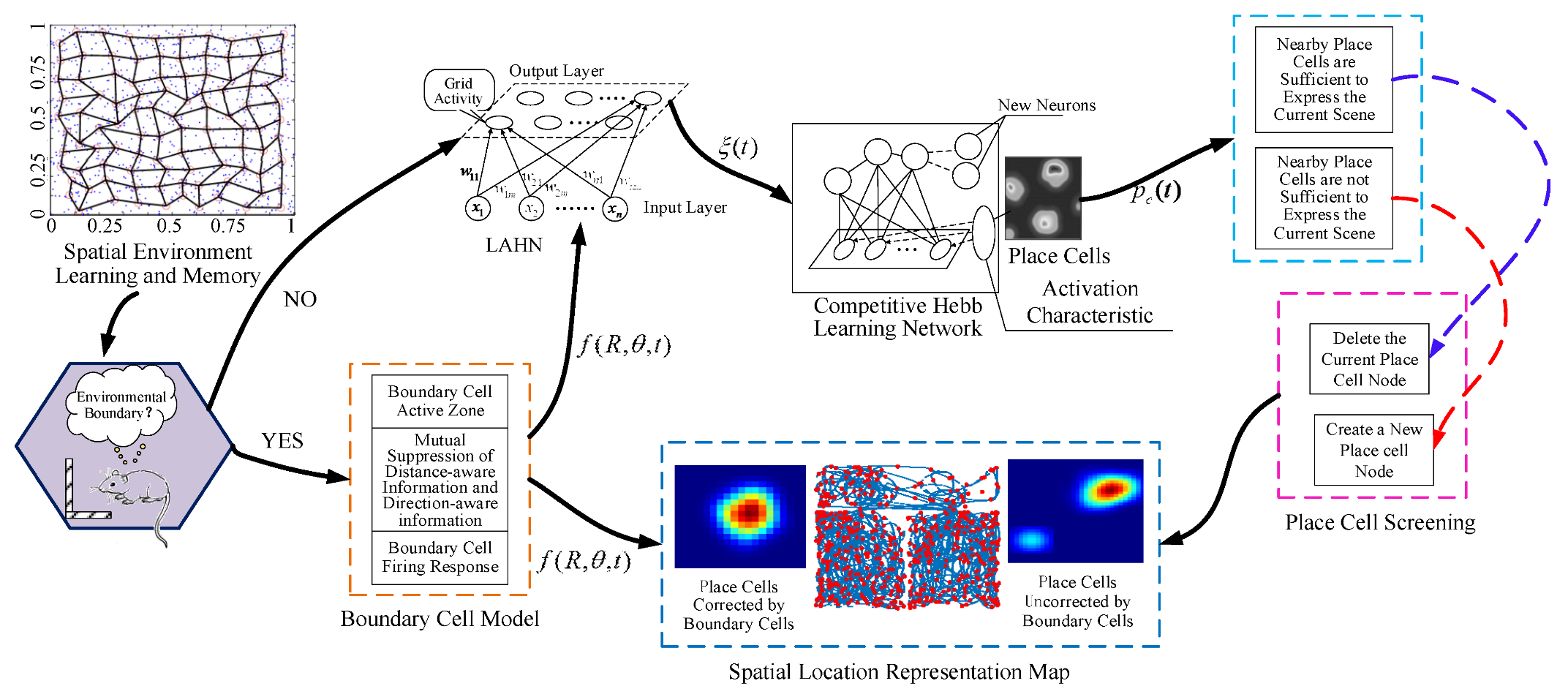
2. Spatial Navigation Cell Model
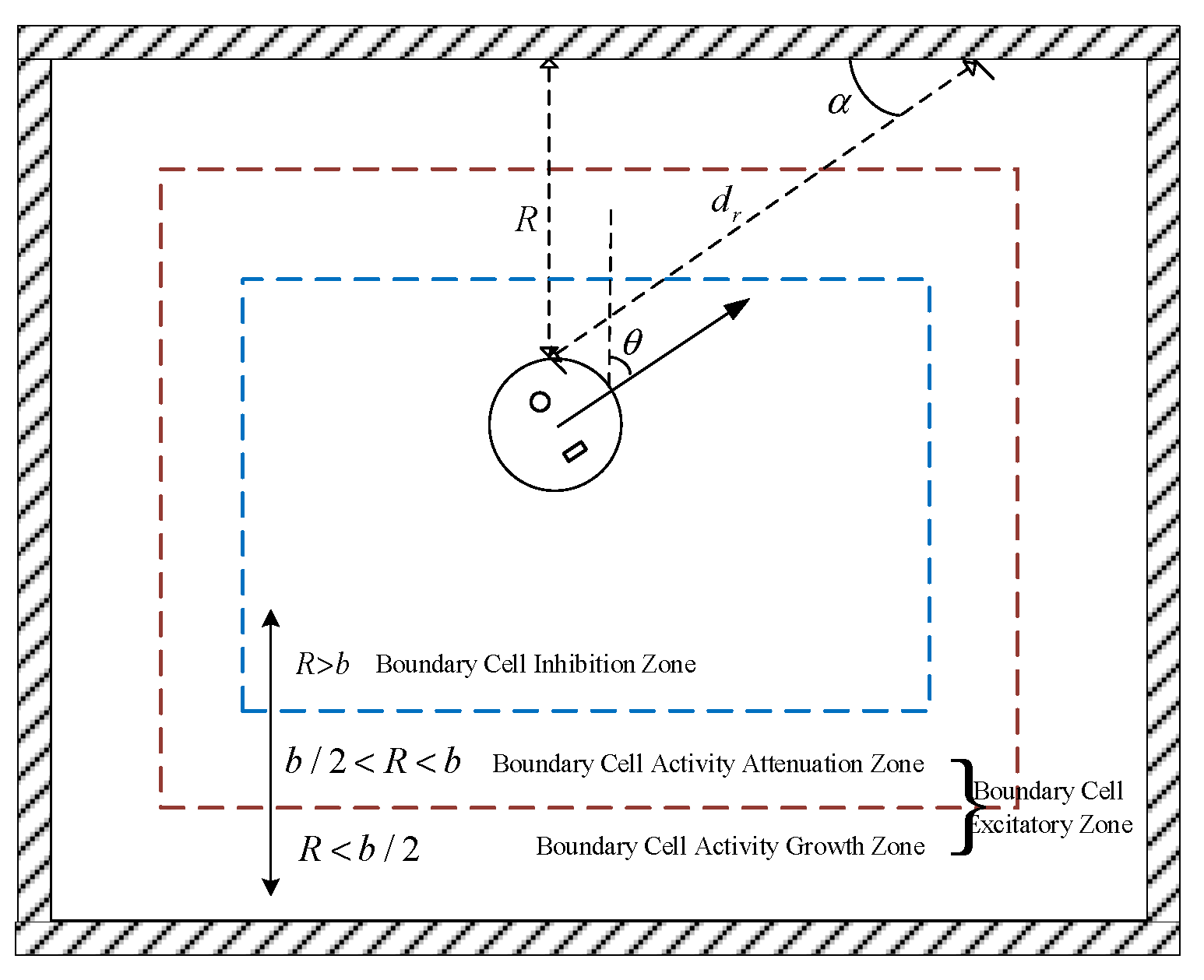
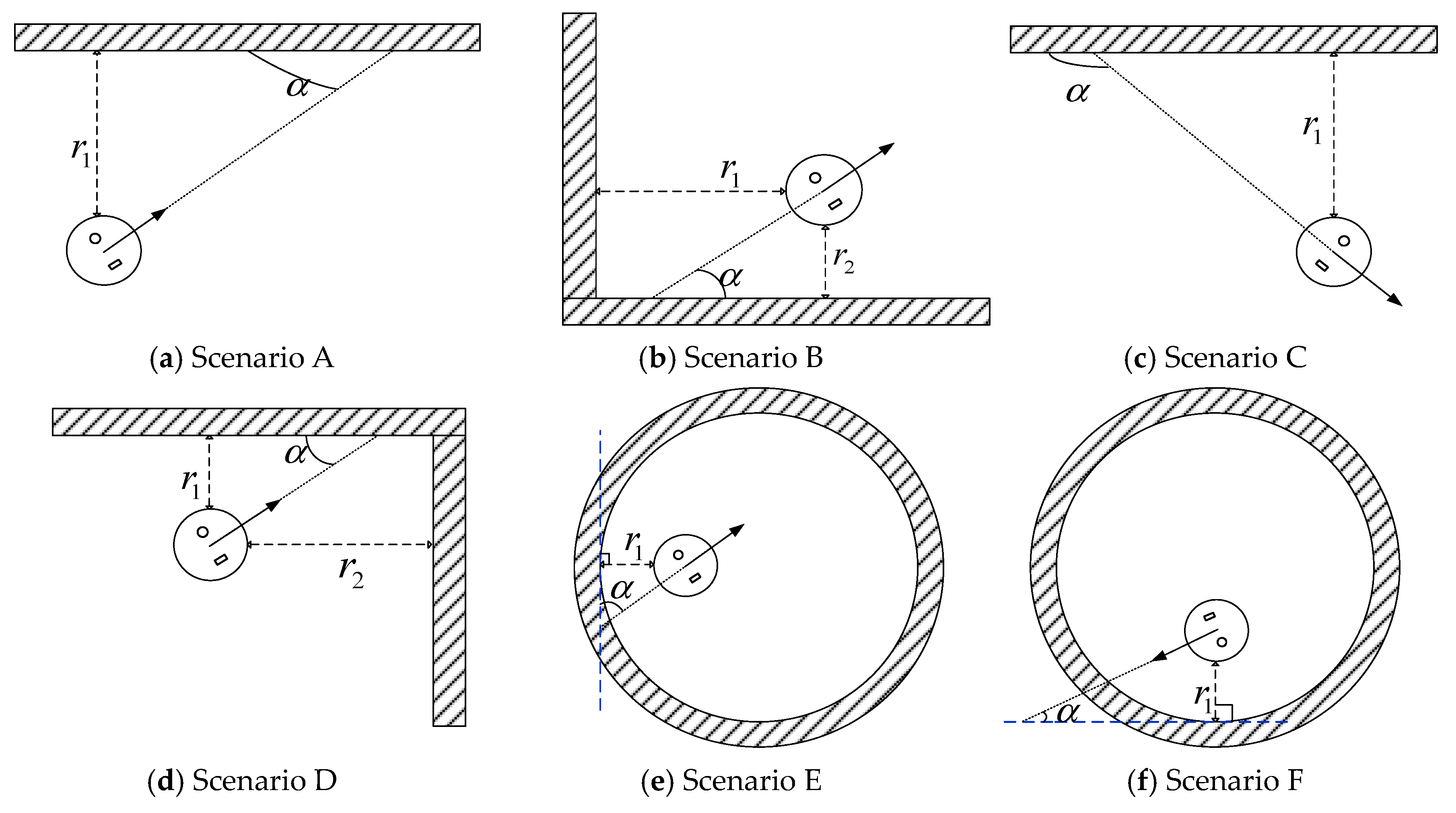
2.1. Boundary Cell Modeling
2.2. Grid Cell Update Model Based on Boundary Information
3. Spatial Location Representation Map Construction
| Algorithm 1: Spatial location representation map construction algorithm |
| Input: Grid cell response value, place cell distance threshold Output: Spatial Location Representation Map BEGIN: FOR Get grid cell response values Updating winning place cells through competitive Hebb learning network |
| Calculate the Euclidean distance between Current place cell and nearby place cell |
| IF < The previous place cell can represent the current scene, continue run forward ELSE The previous palace cell is not enough to represent the current scene and construct a new place cell END IF IF the movement is not over Continue forward motion and update grid cell response value information ELSE Output spatial location representation map END IF ND FOR |
4. Experimental Results and Analysis
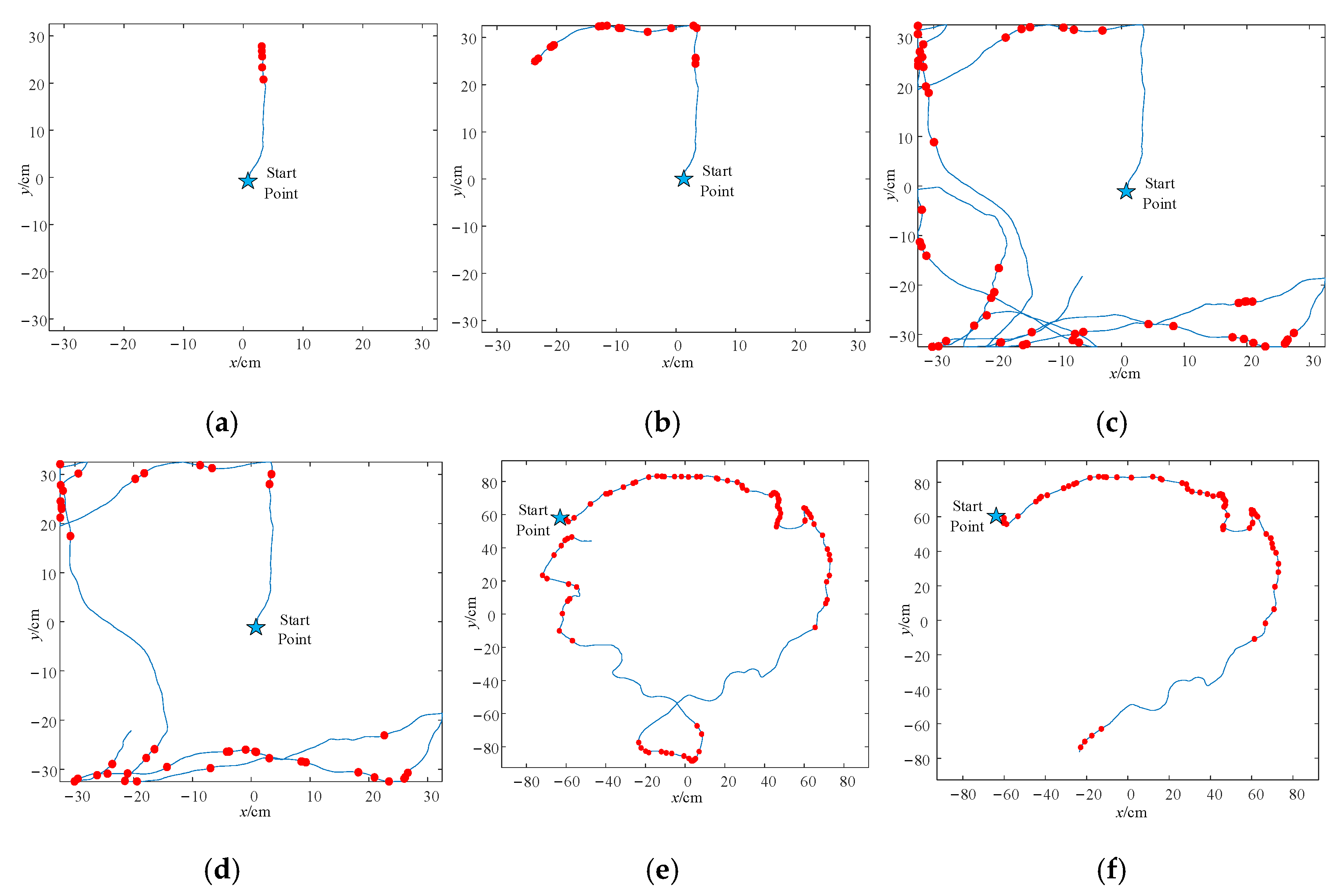
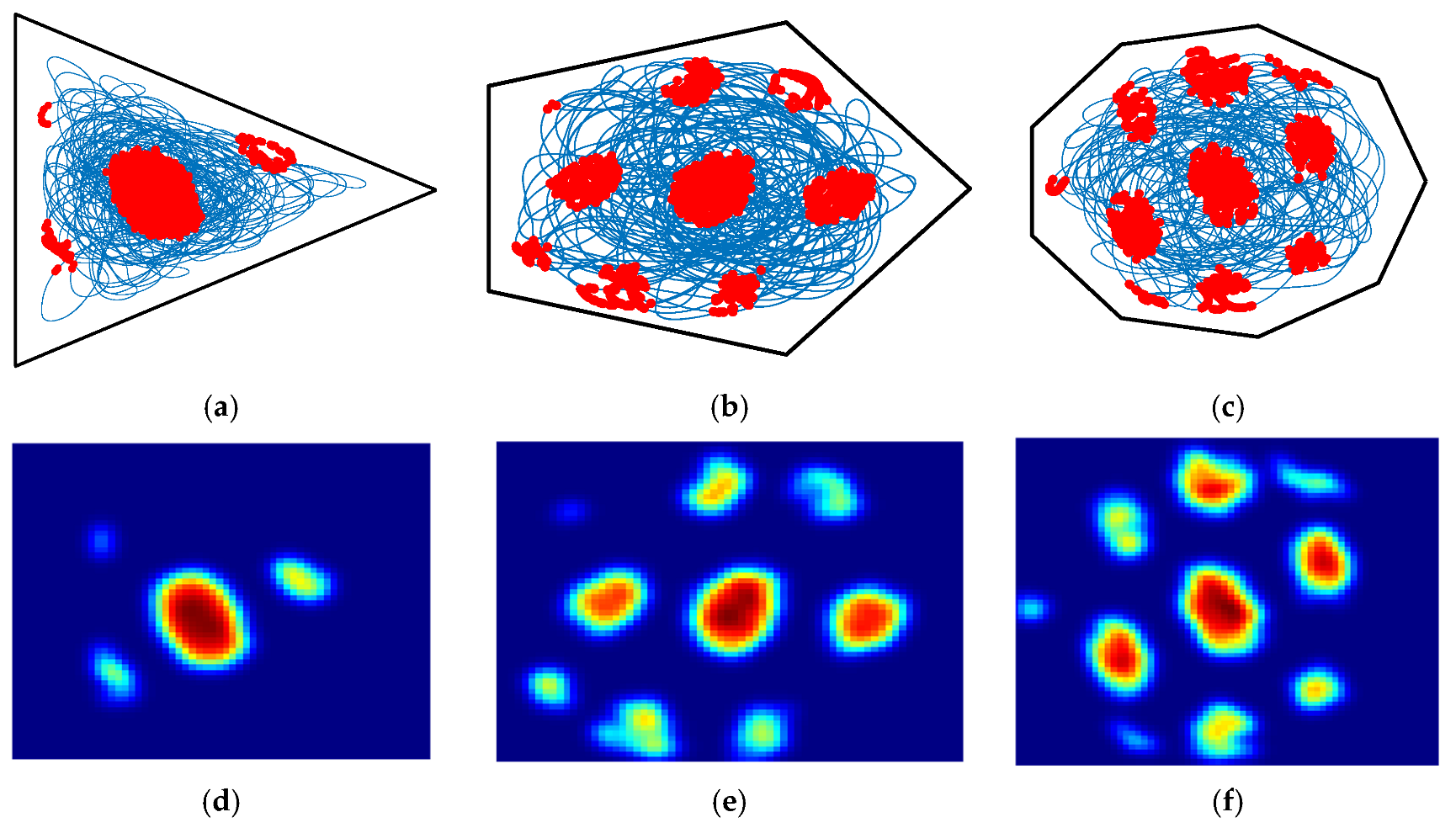
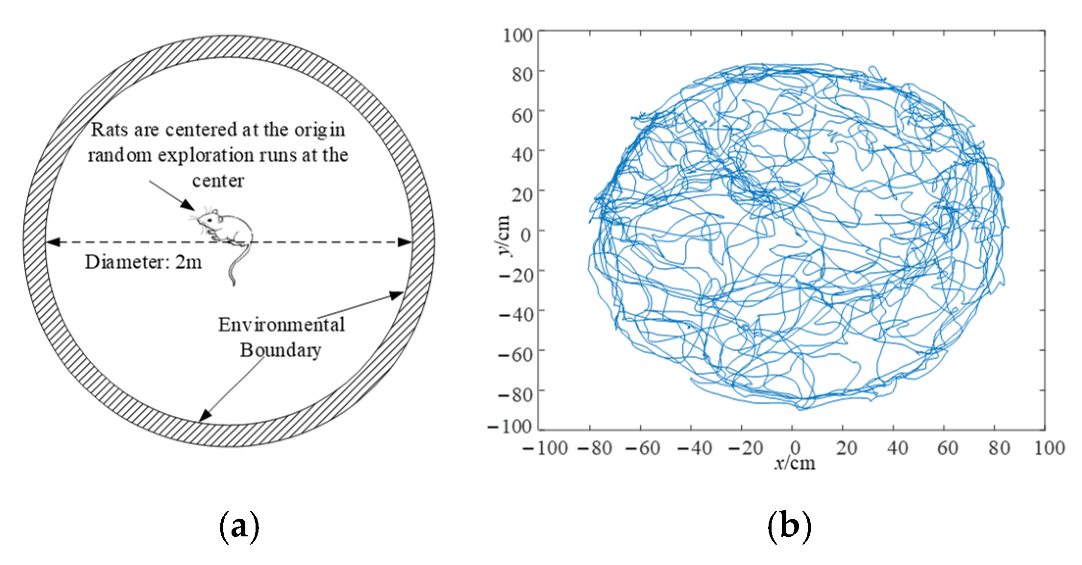
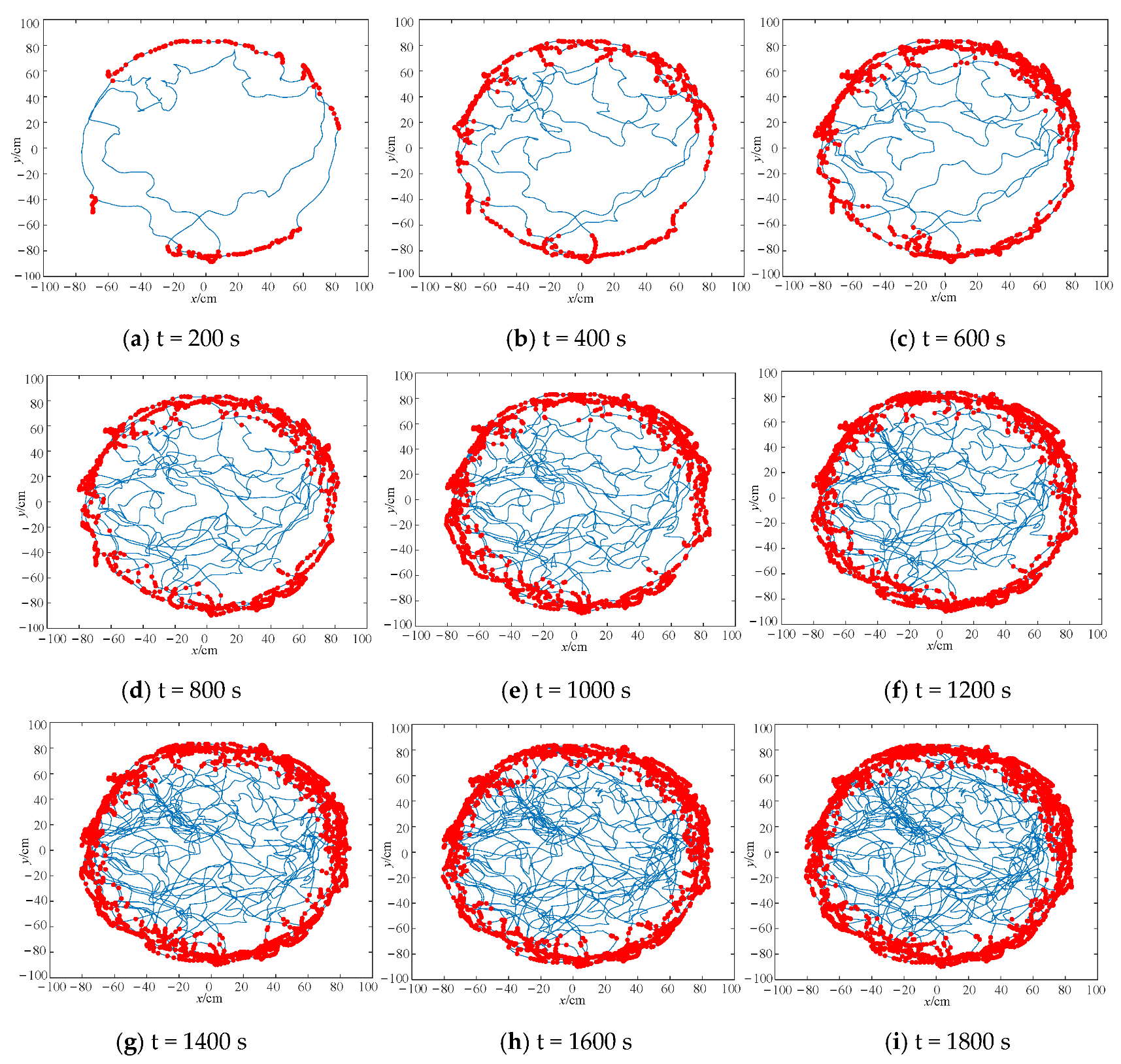
4.1. Boundary Cell Simulation Experiments
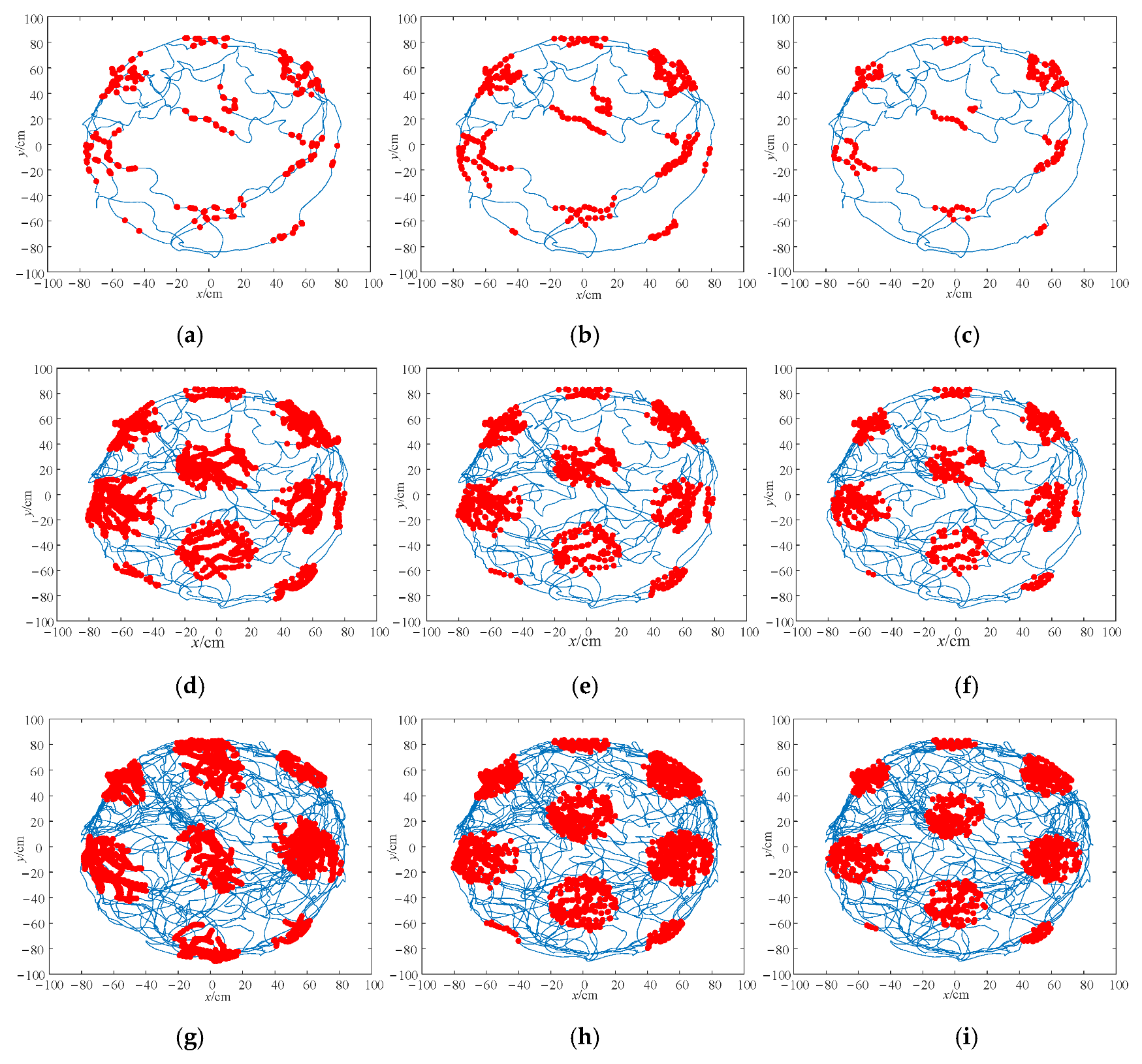
4.2. Grid Cell Simulation Experiment
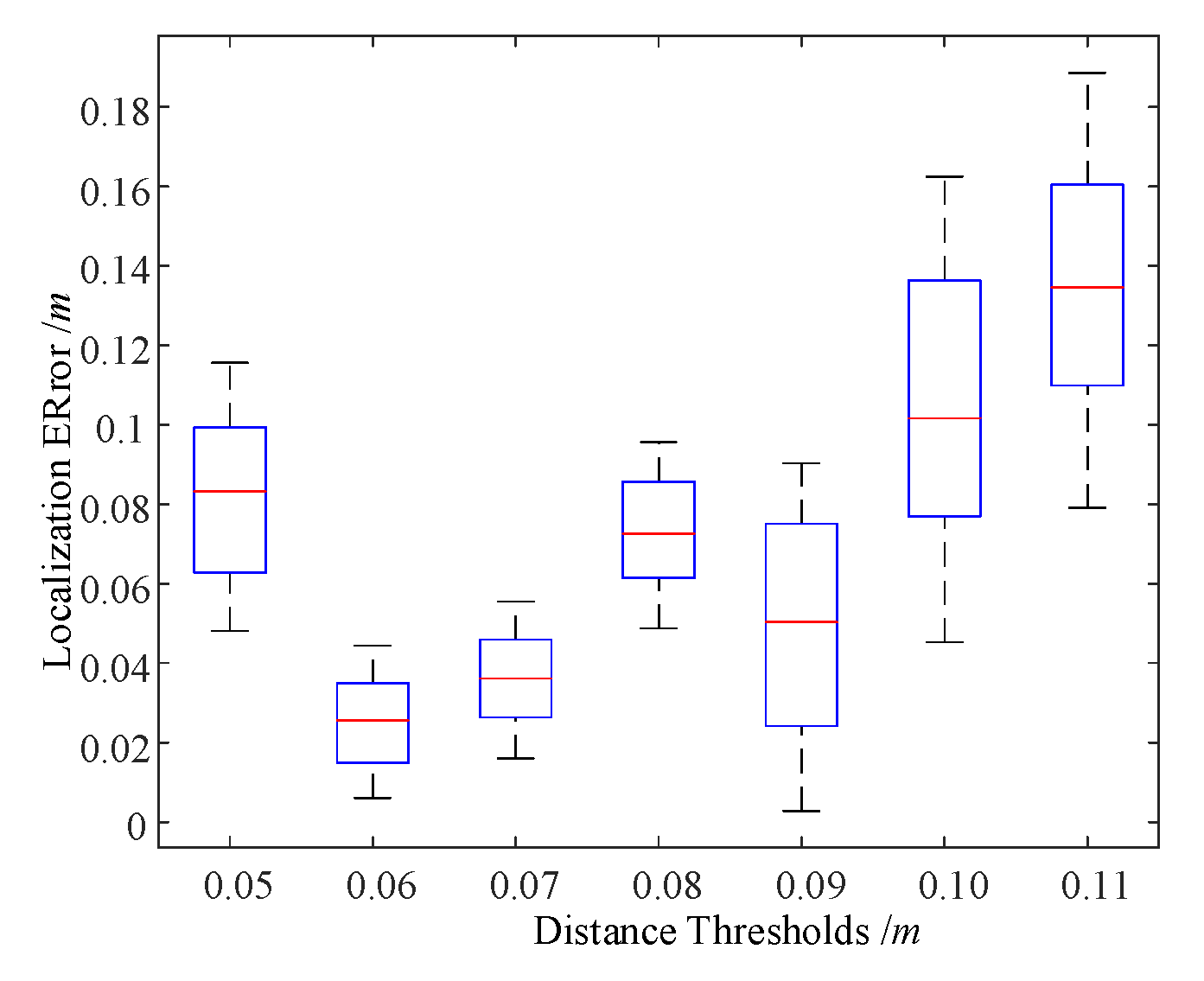
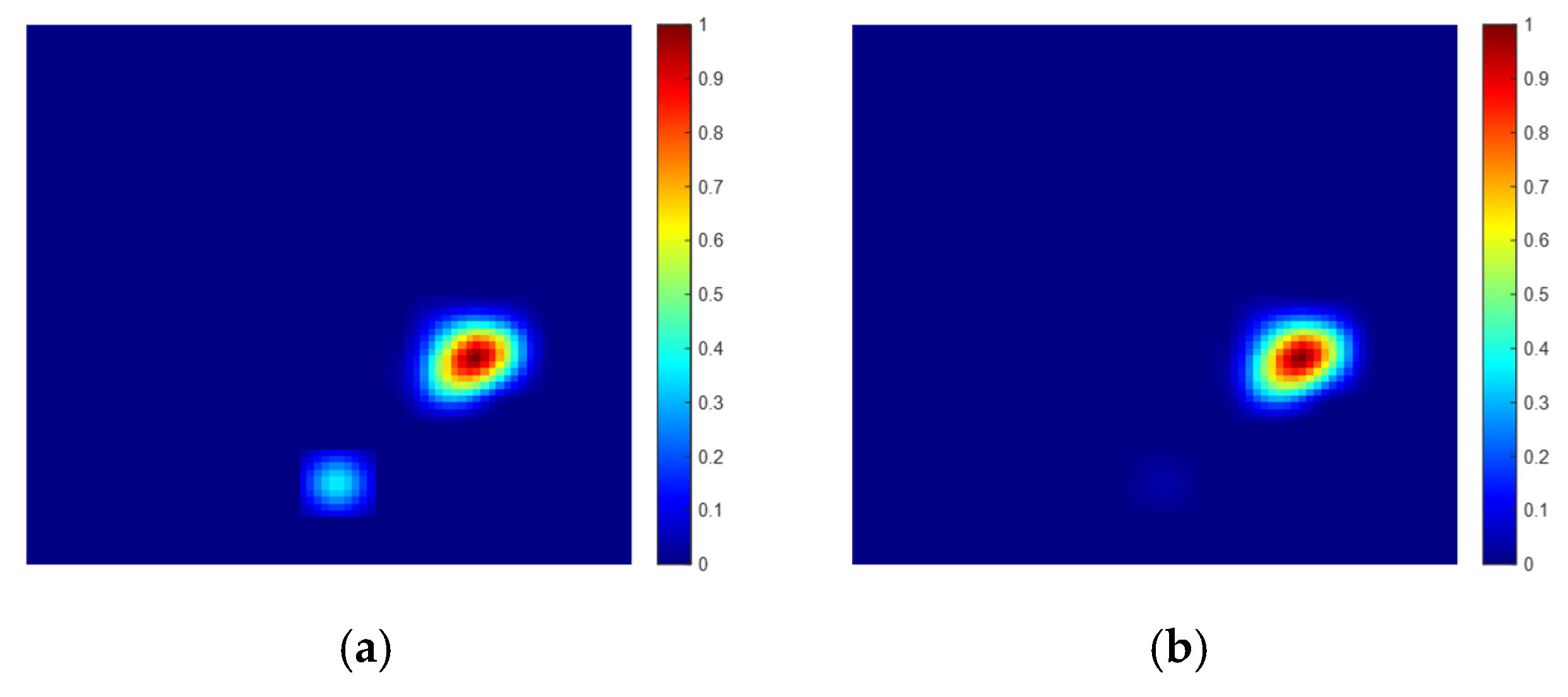
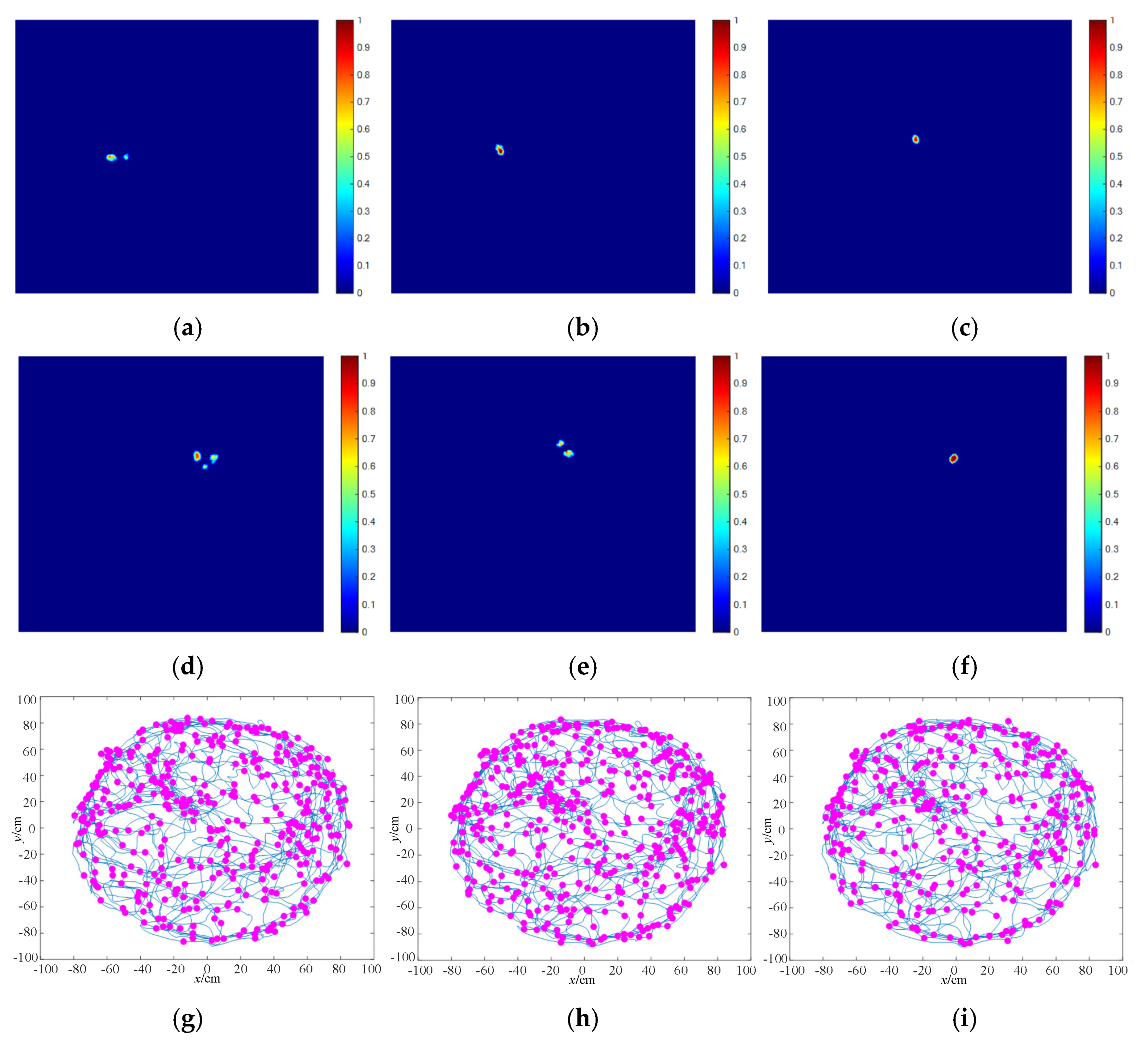
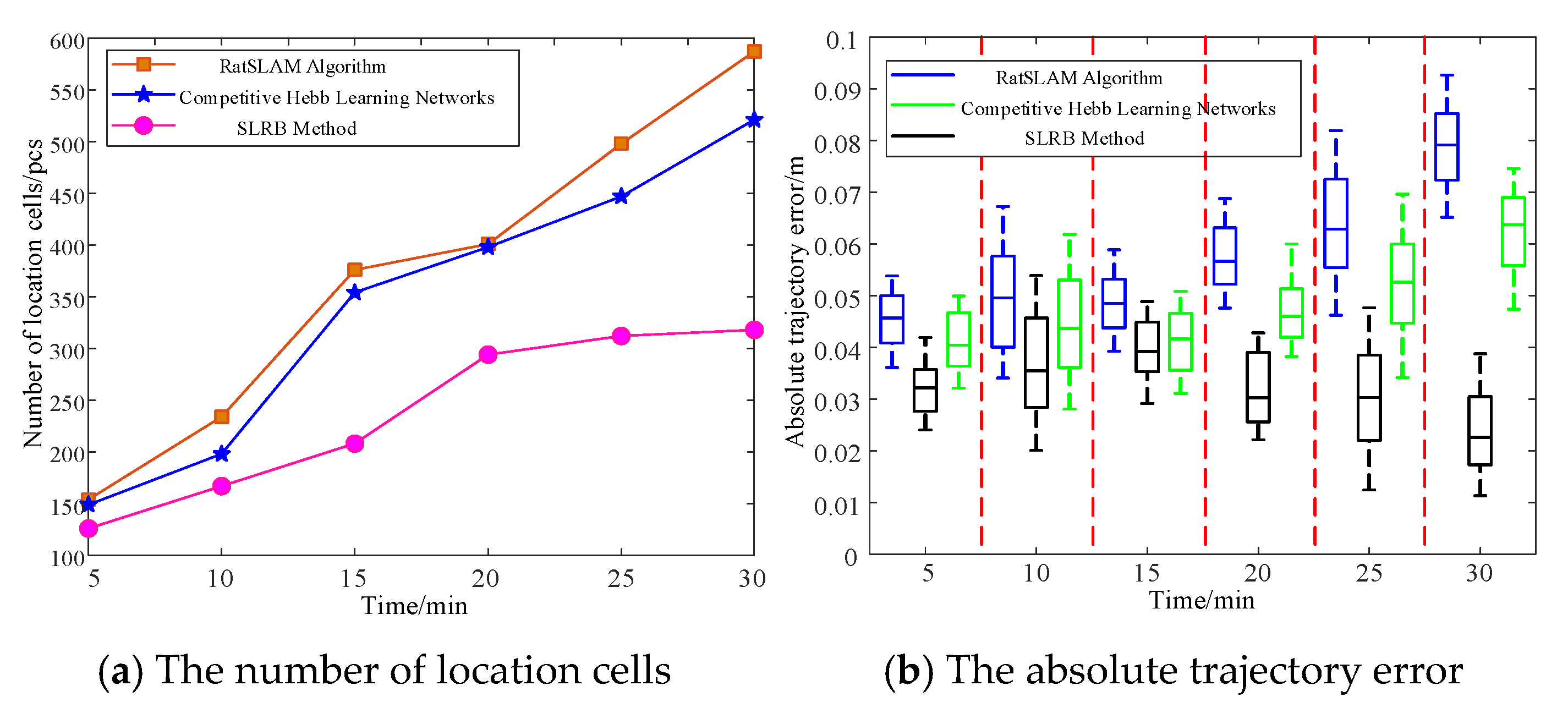
4.3. Spatial Location Representation Map Construction Experiment
5. Analysis and Discussion
6. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Yu, H.-J.; Fang, Y.-C. A robot scene recognition method based on improved autonomous developmental network. Acta Autom. Sin. 2021, 47, 1530–1538. [Google Scholar]
- Yu, F.; Wu, Y.; Ma, S.; Xu, M.; Li, H.; Qu, H.; Song, C.; Wang, T.; Zhao, R.; Shi, L. Brain-inspired multimodal hybrid neural network for robot place recognition. Sci. Robot. 2023, 8, 69–96. [Google Scholar] [CrossRef]
- Sorscher, B.; Mel, G.C.; Ocko, S.A.; Giocomo, L.M.; Ganguli, S. A unified theory for the computational and mechanistic origins of grid cells. Neuron 2023, 111, 121–137. [Google Scholar] [CrossRef]
- Zheng, J.; Huang, G. A novel grid cell–based urban flood resilience metric considering water velocity and duration of system performance being impacted. J. Hydrol. 2023, 617-628, 128911. [Google Scholar] [CrossRef]
- Wu, Y.; Ruan, X.-G.; Huang, J.; Chai, J. An improved cortical network model for environment cognition. Acta Autom. Sin. 2021, 47, 1401–1411. [Google Scholar]
- Ruan, X.-G.; Chai, J.; Wu, Y.; Zhang, X.-P. Huang Jing. Cognitive map construction and navigation based on hippocampal place cells. Acta Autom. Sin. 2021, 47, 666–677. [Google Scholar]
- Tian, Y.; Chang, Y.; Arias, F.H.; Nieto-Granda, C.; How, J.P.; Carlone, L. Kimera-multi: Robust, distributed, dense metric-semantic slam for multi-robot systems. IEEE Trans. Robot. 2022, 38, 2022–2038. [Google Scholar] [CrossRef]
- O’Keefe, J.; Doslrovskv, J. The hippocampus as a spatial map. preliminary evidence from unit activity in the freely-moving rat. Brain Res. 1971, 34, 171–175. [Google Scholar] [CrossRef] [PubMed]
- Moser, E.I.; Kropff, E.; Moser, M.B. Place cells, grid cells, and the brain’s spatial representation system. Annu. Rev. Neurosci. 2008, 31, 69–89. [Google Scholar] [CrossRef] [PubMed]
- Jeffery, K.J.; Burgess, N. A metric for the cognitive map: Found at last? Trends Cogn. Sci. 2006, 10, 1–3. [Google Scholar] [CrossRef] [PubMed]
- Hafting, T.; Fyhn, M.; Molden, S.; Moser, M.B.; Moser, E.I. Microstructure of a spatial map in the entorhinal cortex. Nature 2005, 436, 801–806. [Google Scholar] [CrossRef]
- Krishna, A.; Mittal, D.; Virupaksha, S.G.; Nair, A.R.; Narayanan, R.; Thakur, C.S. Biomimetic FPGA-based spatial navigation model with grid cells and place cells. Neural Netw. 2021, 139, 45–63. [Google Scholar] [CrossRef] [PubMed]
- Campbell, M.G.; Ocko, S.A.; Mallory, C.S.; Low, I.I.; Ganguli, S.; Giocomo, L.M. Principles governing the integration of landmark and self-motion cues in entorhinal cortical codes for navigation. Nat. Neurosci. 2018, 21, 1096–1106. [Google Scholar] [CrossRef] [PubMed]
- Moser, E.I.; Roudi, Y.; Witter, M.P.; Kentros, C.; Bonhoeffer, T.; Moser, M.B. Grid cells and cortical representation. Nuture Rev. Neuro Sci. 2014, 15, 466–481. [Google Scholar] [CrossRef]
- Barry, C.; Hayman, R.; Burgess, N.; Jeffery, K.J. Experience-dependent rescaling of entorhinal grids. Nat Neurosci. 2007, 10, 682–684. [Google Scholar] [CrossRef]
- Milford, M.J.; Wyeth, G.F. Mapping a suburb with a single camera using a biologically inspired SLAM system. IEEE Trans. Robot. 2008, 24, 1038–1053. [Google Scholar] [CrossRef]
- Yu, S.; Xu, H.; Wu, C.; Jiang, X.; Sun, R.; Sun, L. Bionic Path Planning Fusing Episodic Memory Based on RatSLAM. Biomimetics 2023, 8, 59–68. [Google Scholar] [CrossRef]
- Shipston-Sharman, O.; Solanka, L.; Nolan, M.F. Continuous attractor network models of grid cell firing based on excitatory-inhibitory interactions. J. Physiol. 2016, 594, 6547–6557. [Google Scholar] [CrossRef]
- Yu, N.G.; Yuan, Y.H.; Li, T.; Jiang, X.J.; Luo, Z.W. A cognitive map building algorithm by means of cognitive mechanism of hippocampus. Acta Autom. Sin. 2018, 44, 52–73. [Google Scholar]
- O’Keefe, J.; Burgess, N. Geometric determinants of the place fields of hippocampal neurons. Nature 1996, 381, 425–428. [Google Scholar] [CrossRef]
- Grieves, R.M.; Duvelle, E.; Dudchenko, P.A. A boundary vector cell model of place field repetition. Spatial Cogn. Comput. 2018, 18, 1–40. [Google Scholar] [CrossRef]
- Bjerknes, T.; Moser, E.; Moser, M.B. Representation of geometric borders in the developing rat. Neuron 2014, 82, 71–78. [Google Scholar] [CrossRef] [PubMed]
- Hardcastle, K.; Ganguli, S.; Giocomo, L.M. Environmental boundaries as an error correction mechanism for grid cells. Neuron 2015, 86, 827–839. [Google Scholar] [CrossRef] [PubMed]
- Weber, S.N.; Sprekeler, H. Learning place cells, grid cells and invariances with excitatory and inhibitory plasticity. Elife 2018, 7, e34560. [Google Scholar] [CrossRef]
- Gonzalez, M.; Marchand, E.; Kacete, A.; Royan, J. Twistslam: Constrained slam in dynamic environment. IEEE Robot. Autom. Lett. 2022, 7, 6846–6853. [Google Scholar] [CrossRef]
- Kim, H.; Granström, K.; Svensson, L.; Kim, S.; Wymeersch, H. PMBM-based SLAM filters in 5G mmwave vehicular networks. IEEE Trans. Veh. Technol. 2022, 71, 8646–8661. [Google Scholar] [CrossRef]
- Jayakumar, S.; Narayanamurthy, R.; Ramesh, R.; Soman, K.; Muralidharan, V.; Chakravarthy, V.S. Modeling the Effect of Environmental Geometries on Grid Cell Representations. Front. Neural Circuits 2019, 12, 120. [Google Scholar] [CrossRef] [PubMed]
- Fldiák, P. Forming sparse representations by local anti-Hebbian learning. Biol. Cybern. 1990, 64, 165–170. [Google Scholar] [CrossRef] [PubMed]
- Sargolini, F.; Fyhn, M.; Hafting, T.; McNaughton, B.L.; Witter, M.P.; Moser, M.B.; Moser, E.I. Conjunctive representative of position, direction, and velocity in entorchinal cortex. Science 2006, 312, 758–762. [Google Scholar] [CrossRef] [PubMed]
- Soman, K.; Muralidharan, V.; Chakravarthy, V.S. A unified hierarchical oscillatory network model of head direction cells, spatially periodic cells and place cells. Eur. J. Neurosci. 2018, 47, 1266–1281. [Google Scholar] [CrossRef]
- Jeewajee, A.; Barry, C.; O’Keefe, J.; Burgess, N. Grid cells and theta as oscillatory interference: Electrophysiological data from freely moving rats. Hippocampus 2010, 18, 1175–1185. [Google Scholar] [CrossRef] [PubMed]
- Cong, M.; Zou, Q.; Liu, D.; Du, Y. Review of robot navigation inspired by the localization cells cognitive mechanism. J. Mech. Eng. 2019, 55, 1–12. [Google Scholar]
- Knudsen, E.B.; Wallis, J.D. Hippocampal neurons construct a map of an abstract value space. Cell 2021, 184, 4640–4650. [Google Scholar] [CrossRef] [PubMed]
- Guo, J.; Cheng, M.; Ren, J.; Ren, Q. A Bio-inspired SLAM System for a Legged Robot. In Proceedings of the 2022 IEEE 17th Conference on Industrial Electronics and Applications (ICIEA), Chengdu, China, 16–19 December 2022; pp. 1074–1079. [Google Scholar]














{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Time (s) | 30 | 60 | 90 | 120 | 150 | 180 | 210 | 240 | 270 | 300 | 330 | 360 |
|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Trilateral environment | 0.25 | 0.32 | 0.38 | 0.45 | 0.55 | 0.56 | 0.68 | 0.69 | 0.74 | 0.73 | 0.74 | 0.74 |
| Pentagonal environment | 0.24 | 0.35 | 0.41 | 0.51 | 0.62 | 0.68 | 0.72 | 0.75 | 0.78 | 0.78 | 0.78 | 0.78 |
| Nine-sided environment | 0.20 | 0.34 | 0.40 | 0.49 | 0.58 | 0.71 | 0.80 | 0.84 | 0.86 | 0.84 | 0.85 | 0.85 |
| Time (s) | 200 | 400 | 600 | 800 | 1000 | 1200 | 1400 | 1600 | 1800 |
|---|---|---|---|---|---|---|---|---|---|
| Number of boundary cells (pcs) | 103 | 195 | 274 | 318 | 378 | 421 | 472 | 503 | 498 |
| Mean localization error (m) | 0.67 | 0.71 | 0.83 | 0.86 | 0.74 | 0.61 | 0.50 | 0.39 | 0.37 |
| Exploration Time (min) | Number of Activated Grid Cells (pcs) | Grid Cell Fraction | ||||
|---|---|---|---|---|---|---|
| OI | CAN | SLRB | OI | CAN | SLRB | |
| 5 | 171 | 160 | 128 | 0.71 | 0.75 | 0.74 |
| 10 | 382 | 171 | 165 | 0.73 | 0.74 | 0.69 |
| 15 | 410 | 290 | 195 | 0.68 | 0.79 | 0.79 |
| 20 | 472 | 353 | 287 | 0.54 | 0.71 | 0.84 |
| 25 | 524 | 427 | 354 | 0.51 | 0.72 | 0.88 |
| 30 | 614 | 541 | 478 | 0.43 | 0.65 | 0.86 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Jiang, H.; Zhang, Y. A Spatial Location Representation Method Incorporating Boundary Information. Appl. Sci. 2023, 13, 7929. https://doi.org/10.3390/app13137929
Jiang H, Zhang Y. A Spatial Location Representation Method Incorporating Boundary Information. Applied Sciences. 2023; 13(13):7929. https://doi.org/10.3390/app13137929
Chicago/Turabian StyleJiang, Hui, and Yukun Zhang. 2023. "A Spatial Location Representation Method Incorporating Boundary Information" Applied Sciences 13, no. 13: 7929. https://doi.org/10.3390/app13137929
APA StyleJiang, H., & Zhang, Y. (2023). A Spatial Location Representation Method Incorporating Boundary Information. Applied Sciences, 13(13), 7929. https://doi.org/10.3390/app13137929