Featured Application
The β-galactosidase enzyme identified in this study can be used to convert isoflavone glycosides to aglycones in wild soybeans.
Abstract
Isoflavone glycosides are commonly biotransformed into isoflavone aglycones due to the superior biological activities of the latter. Wild soybeans contain a higher isoflavone content than domesticated soybeans due to their high level of genetic diversity. In this study, we cloned and characterized a thermostable β-galactosidase from the extreme thermophile Thermoproteus uzoniensis for potential application in isoflavone conversion in Korean wild soybeans. The purified recombinant enzyme exhibited a maximum specific activity of 1103 μmol/min/mg at pH 5.0 and 90 °C with a half-life of 46 h and exists as a homodimer of 113 kDa. The enzyme exhibited the highest activity for p-nitrophenyl (pNP)-β-D-galactopyranoside among aryl glycosides and it hydrolyzed isoflavone glycosides in the order genistin > daidzin > ononin > glycitin. The enzyme completely hydrolyzed 2.77 mM daidzin and 3.85 mM genistin in the seed extract of wild soybean after 80 and 70 min with productivities of 1.86 and 3.30 mM/h, respectively, and 9.89 mM daidzin and 1.67 mM genistin in the root extract after 180 and 30 min, with the highest productivities of 3.30 and 3.36 mM/h, respectively, compared to other glycosidases. Our results will contribute to the industrial production of isoflavone aglycone using wild soybean and this is the first report on the enzymatic production of isoflavone aglycones from isoflavone glycosides in wild soybeans.
1. Introduction
Isoflavones, one of the most abundant phytochemicals in leguminous plants, have attracted attention because of their activity against cancer [1], cardiovascular disease [2], osteoporosis [3], and menopausal symptoms [4]. The biological activity of isoflavones is improved by their deglycosylation into aglycones, which are more quickly absorbed in the gut because of their lower molecular weight and better hydrophobicity [5,6]. However, isoflavone aglycones exist at low concentrations in leguminous plants. Therefore, many studies have focused on the biotransformation of isoflavone glycosides such as daidzin, genistin, and glycitin into their corresponding isoflavone aglycones; daidzein, genistein, and glycitein. These isoflavone aglycones have been obtained through the hydrolytic activity of microorganisms, including Bacillus subtilis, Enterococcus durans, Lactobacillus paraplantarum, Rhizopus oligosporus, Streptococcus salivarius, and Weissella confusa [7,8,9]. However, microbial hydrolysis has several disadvantages, including low productivity, poor yield, and the production of by-products. Therefore, enzymes have been commonly used for the hydrolysis of isoflavone glycosides because of their high specificity, productivity, and yield. Although various microbial and plant-derived glycosidases have been used, thermostable glycosidases from thermophilic microorganisms, including Dictyoglomus turgidum [10], Microbulbifer thermotolerans [11], Neosartorya fischeri [12], Pyrococcus furiosus [13], Sulfolobus solfataricus [14], Talaromyces leycettanus [15], Thermofilum pendens [16], and Thermotoga maritima [17] have exhibited high hydrolytic activity toward isoflavone glycosides. In order to use thermostable glycosidases from thermophilic microorganisms, it is more efficient to use recombinant enzymes expressed in E. coli through cloning, because the growth conditions of thermophilic microorganisms are generally harsh.
The extreme thermophile Thermoproteus uzoniensis is a rod-shaped archaebacterium found in hot springs and soil of the Uzon caldera (SW of Kamchatka pen.) [18]. Glycosidase from this strain is a powerful candidate for application in the hydrolysis of isoflavone glycosides because it showed high hydrolytic activity on ginsenosides (ginseng saponins) in a previous study [19]. However, this enzyme has not yet been characterized.
Soybean (Glycine max (L.) Merr.), a representative leguminous plant, contains a high abundance of isoflavones. However, significant differences have been noted in the total isoflavone content of soybean depending on the country or region of cultivation [20]. Wild soybean (Glycine soja Siebold & Zucc.), the ancestor of the domesticated soybean, was found to harbor a high level of genetic diversity that was abolished during the process of domestication [21]. Therefore, it has been previously suggested that the isoflavone content of domesticated soybean is not as high as that of wild soybean due to unintentional selection [22] and Korean wild soybeans have been reported to contain a high amount of isoflavones [23,24,25].
The global Crop Wild Relatives (CWR) project, launched in 2011, aims to adapt agriculture to climate change. The wild relatives of crops, which have evolved to survive tough conditions, can often contain useful traits, the analysis of which could help develop crops that are more resilient to climate change. Soybeans are one of the main target species for this project and wild soybean is an important CWR of soybean. Thus, considering the global importance of the soybean crop, the enzymatic conversion of isoflavones in wild soybeans to their aglycones is of great interest.
In this study, we cloned a thermostable β-galactosidase from T. uzoniensis and characterized its properties, such as molecular weight, pH, temperature, thermostability, and substrate specificity. The enzyme was used to produce daizein and genistein from daidzin and genistin in the seed and root extracts of Korean wild soybeans.
2. Materials and Methods
2.1. Materials
The aryl-glycosides, p-nitrophenyl (pNP)-β-D-glucopyranoside, pNP-β-D-galactopyranoside, pNP-β-D-xylopyranoside, pNP-α-L-arabinopyranoside, pNP-α-L-rhamnopyranoside, oNP-β-D-glucopyranoside, and oNP-β-D-galactopyranoside; and isoflavones, including daidzin, daidzein, genistin, genistein, glycitin, and glycitein were purchased from Sigma-Aldrich (St. Louis, MO, USA). Korean wild soybeans were provided by the Baekdudagan Wild Plant Seed Bank. All enzymes used for gene cloning were purchased from Thermo Fisher Scientific (Waltham, MA, USA) or New England Biolabs (Hertfordshire, UK).
2.2. Bacterial Strains, Plasmids, and Gene Cloning
The DNA template and expression vector for the β-galactosidase gene (GenBank accession no. AEA12778) cloning and host strain were T. uzoniensis DSM 5263 (DSMZ, Braunschweig, Germany), pET-29b (+) vector (Novagen, Darmstadt, Germany), and Escherichia coli BL21 (DE3), respectively. The β-galactosidase gene was cloned using the one-step isothermal DNA assembly method [26]. For the amplification of DNA fragments, the following primers were used: forward primer for β-galactosidase gene, AGC AGC GAA AAC CTG TAT TTT CAG GGA CAT ATG CGG AAG TTC CCA AGC GGA TTC AGA TGG; reverse primer for β-galactosidase gene, ATC TCA GTG GTG GTG GTG GTG GTG CTC GAG TTA AAG CGA TCT GAG GCC CCT CAA GAC CTC; forward primer for vector, GAG GTC TTG AGG GGC CTC AGA TCG CTT TAA CTC GAG CAC CAC CAC CAC CAC CAC TGA GAT; and reverse primer for vector, CCA TCT GAA TCC GCT TGG GAA CTT CCG CAT ATG TCC CTG AAA ATA CAG GTT TTC GCT GCT. The DNA fragments of the β-galactosidase gene and vector were synthesized at an annealing temperature of 55 °C using the Phusion High-Fidelity DNA polymerase (Thermo Fisher Scientific) by PCR. The synthesized DNA fragments, which consisted of amplicons with 60 bp of overhangs for the gene (1539 bp) and vector (5299 bp), were ligated using Master Mix for Gibson Assembly (New England Biolabs, Ipswich, MA, USA), and the ligated DNA was transformed into E. coli BL21.
2.3. Preparation of β-Galactosidase
E. coli cells containing β-galactosidase from T. uzoniensis were cultivated in a 2 L flask containing 500 mL of Luria-Bertani medium containing 50 µg/mL kanamycin at 37 °C, with 200 rpm shaking. When the optical density at 600 nm of the recombinant E. coli cells reached 0.6, expression of β-galactosidase from T. uzoniensis was induced by adding 100 mM IPTG and the cells were further incubated at 16 °C with 150 rpm shaking for 14 h.
The cells were harvested by centrifugation at 10,000 × g, suspended in a Ni-NTA lysis buffer containing 1 mg/mL lysozyme and disrupted by sonication on ice for 20 min. Cell debris and undisrupted cells were removed by centrifugation at 13,000× g for 20 min at 4 °C and the supernatant was used as a crude extract. The crude enzyme solution was applied to a His-Trap HP affinity column (Amersham Biosciences, Uppsala, Sweden) on a fast protein liquid chromatography system (Bio-Rad, Hercules, CA, USA) [27]. The bound protein was subsequently eluted with 20 mM phosphate buffer containing 250 mM imidazole at a flow rate of 1 mL/min. The collected fraction with hydrolytic activity was dialyzed against 50 mM citrate/phosphate buffer (CPB) at pH 5.0 for 12 h at 4 °C. The resulting solution was used as the purified β-galactosidase.
2.4. SDS-PAGE and Gel Filtration Chromatography
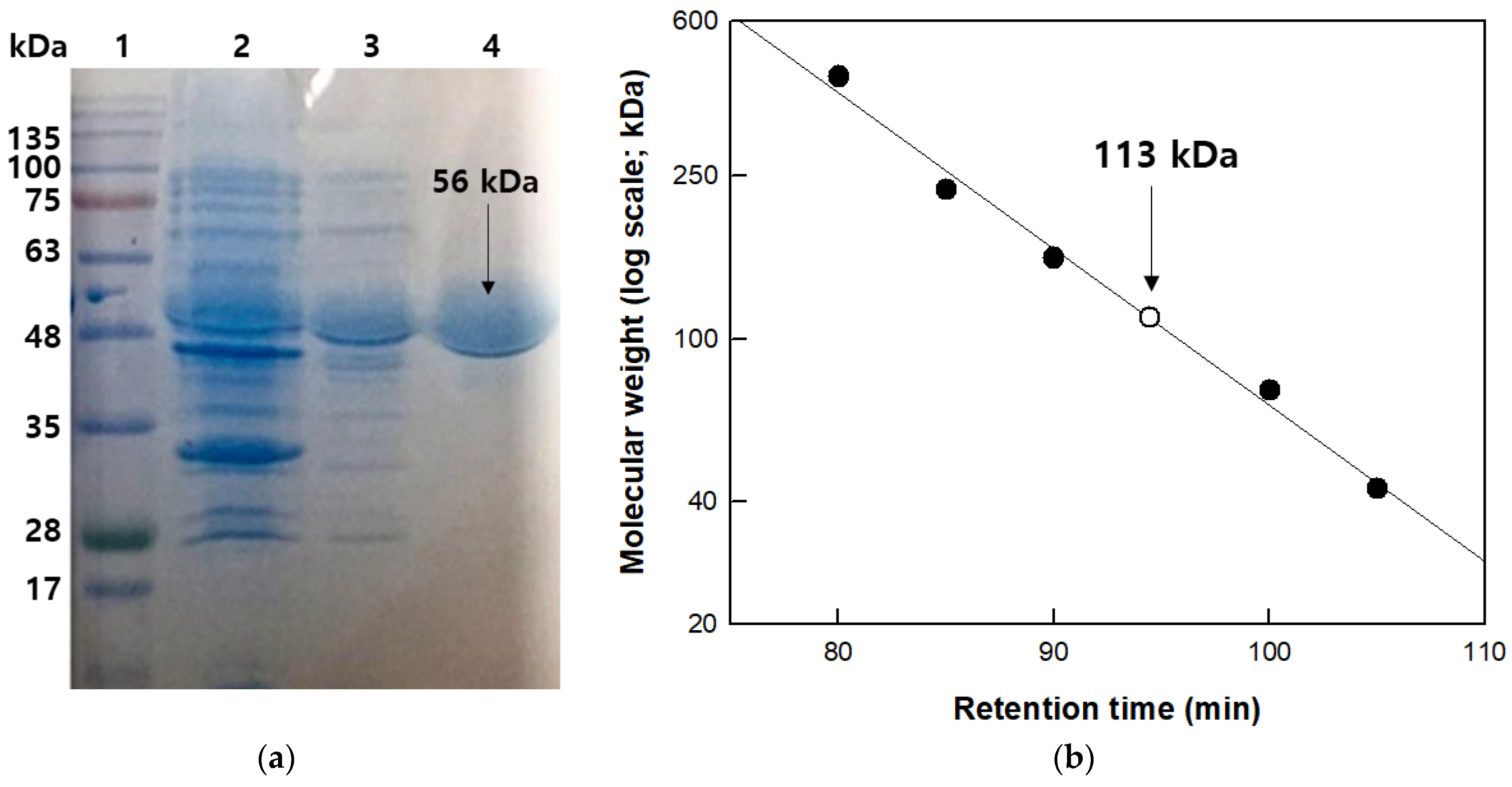
The molecular mass of the β-galactosidase subunit was examined by SDS-PAGE using a prestained protein ladder (MBI Fermentas, Hanover, MD, Canada) as a reference. Cell debris, crude extract, and purified enzyme were used at a concentration of 1 mg/mL to evaluate the degree of purification. All protein bands were stained with Coomassie blue dye for visualization. Gel-filtration chromatography was performed using a HiPrepTM 16/60 Sephacryl S-300 HR column (GE Healthcare, Chicago, IL, USA) to determine the total molecular mass of the recombinant β-galactosidase. The recombinant β-galactosidase was transferred through the column and the enzyme-bound column was eluted with 50 mM CPB (pH 5.0) containing 150 mM NaCl at a flow rate of 0.5 mL/min. The retention time (migration length) of β-galactosidase from T. uzoniensis was measured during the elution process using reference proteins (Amersham Biosciences) such as ferritin (440 kDa), catalase (232 kDa), aldolase (158 kDa), conalbumin (75 kDa), and ovalbumin (43 kDa). The total molecular mass of the β-galactosidase from T. uzoniensis was calculated by comparing its retention time (migration length) with those of the reference proteins.
2.5. Preparation of Wild Soybean Extracts
Korean wild soybean cultivated in the Baekdudaegan National Arboretum was used to prepare its seed, stem, and root extracts. The extract method was modified on the basis of the extraction method for soy flour [17]. Each part of the Korean wild soybean was ground using an electric grinder (BioloMix, Guangzhou, China). A fine powder (100 g) was extracted using 1 L of methanol at 40 °C for 24 h. Each extract was filtered, the methanol was completely removed using a rotary evaporator, and the residue was dissolved in 2 L of distilled water. To prevent the Maillard reaction of the β-galactosidase from T. uzoniensis with free sugars at temperatures above 70 °C, the sugars in the extract were removed using a column packed with Diaion HP20 resin (Sigma-Aldrich). Each extract was loaded onto the column and the isoflavone-adsorbed resin was washed with distilled water. The absorbed isoflavone was then eluted with 2 L of methanol at a flow rate of 0.5 mL/min. The methanol in the eluent was completely evaporated, and the residue was redissolved in 1 L of distilled water. The resulting extracts were used for the analysis of isoflavone content and the production of daidzein and genistein.
2.6. Hydrolytic Activity
Unless otherwise mentioned, the enzyme reaction was performed with 0.005 mg/mL of β-galactosidase from T. uzoniensis and 1 mM pNP-β-D-glucopyranoside for 10 min. The activity was determined by the increase in absorbance at 405 nm due to the release of nitrophenol. To assess the effects of pH and temperature on the hydrolytic activity of β-galactosidase from T. uzoniensis, the pH was varied from 4.0 to 7.0 using 50 mM CPB at 70 °C and the temperature was varied from 65 to 95 °C at pH 5.0. The effect of temperature on the stability of β-galactosidase from T. uzoniensis was monitored as a function of incubation time by incubating the enzyme solution in 50 mM CPB (pH 5.0) at different temperatures (75, 80, 85, 90, and 95 °C). After incubation, the enzyme solutions were assayed in 50 mM CPB (pH 5.0) containing 1 mM pNP-β-D-glucopyranoside at 90 °C for 10 min.
The specific activity of β-galactosidase from T. uzoniensis was determined using aryl-glycosides, including pNP-β-D-glucopyranoside, pNP-β-D-galactopyranoside, pNP-β-D-xylopyranoside, pNP-α-L-arabinopyranoside, pNP-α-L-rhamnopyranoside, oNP-β-D-glucopyranoside, and oNP-β-D-galactopyranoside, and isoflavones such as daidzin, genistin, and glycitin. The reactions were performed in 50 mM CPB (pH 5.0) containing 0.005−0.05 mg/mL β-galactosidase from T. uzoniensis and 1 mM aryl-glycoside or 0.2 mM isoflavone at 90 °C for 10 min.
2.7. Biotransformation of Isoflavones
Time-course reactions for the biotransformation of isoflavones by β-galactosidase from T. uzoniensis were assessed using reagent grade daidzin and genistin, and seed and root extracts of Korean wild soybean as substrates. The biotransformation of reagent grade daidzin or genistin into daidzein or genistein, respectively, was performed in 50 mM CPB (pH 5.0) containing 0.2 mg/mL enzyme and 3 mM isoflavone at 90 °C for 100 or 90 min, respectively. Daidzein and genistein were produced from daidzin and genistin, respectively, in seed or root extracts of Korean wild soybean, in 50 mM CPB (pH 5.0) containing 0.4 mg/mL enzyme and 20% (w/v) seed or root extract at 90 °C for 100 or 200 min, respectively.
2.8. HPLC Analysis
Isoflavones were analyzed using an Agilent 1100 HPLC system (Agilent Technologies, Santa Clara, CA, USA) equipped with a UV detector at 254 nm and a C18 column with dimensions of 4.6 × 150 mm (YMC, Kyoto, Japan). The column was eluted at 35 °C for 40 min at a flow rate of 1.0 mL/min with the following gradient of solvent A (3% acetic acid in water)/solvent B (3% acetic acid in acetonitrile) (v/v): 90/10 to 85/15 for 5 min; 85/15 to 80/20 for 5 min; 80/20 to 75/25 for 5 min; 75/25 to 70/30 for 5 min; 70/30 to 65/35 for 5 min; 65/35 to 10/90 for 5 min; and 10/90 to 90/10 for 10 min.
The retention times of daidzin, daidzein, genistin, genistein, glycitin, and glycitein were 6.2, 14.0, 9.3, 18.5, 7.5, and 15.8 min, respectively. The isoflavones in the reaction samples were confirmed to have the same retention times as those of the isoflavone standards. The quantitative analysis of isoflavones in Korean wild soybean extracts and reaction samples was performed using linear calibration curves relating the peak areas to the concentrations of isoflavone standards [28].
3. Results
3.1. Cloning, Purification, and Molecular Mass Determination of β-Galactosidase from T. uzoniensis
A glycoside hydrolase family 1 (GH1) protein from T. uzoniensis showed high hydrolytic activity on glucosides such as ginsenosides [19], but it has not yet been characterized and applied to hydrolyzed isoflavone. Therefore, the 1482 bp gene encoding the protein from T. uzoniensis DSM 5263 with the same sequence in GenBank (Accession No., AEA12778; gene name, TUZN_RS06265) was cloned and expressed in E. coli BL21 (DE3) in a soluble form. The amino acid sequence of the expressed enzyme showed a 62.1%, 61.9%, 56.9%, and 55.2% identity with those of the GH1 glycosidases from Sulfolobus shibatae (AAA79030) [29], S. solfataricus (AAA72843) [30], Sulfolobus acidocaldarius (AAY81155) [31], and P. furiosus (AAL80197) [32], respectively.
β-Galactosidase from T. uzoniensis was purified using HisTrap affinity chromatography into a soluble protein with a 10.5-fold purification, a final yield of 21%, and a specific activity for pNP-β-D-galactopyranoside of 1103 μmol/min/mg. The molecular mass of the purified enzyme was approximately 57 kDa on SDS-PAGE (Figure 1a), which was consistent with the calculated value of 57,131 Da based on 493 amino acids combined with six histidine residues. The native enzyme was determined to be a homodimer with a molecular mass of 113 kDa by gel-filtration chromatography (Figure 1b). The GH1 glycosidases from S. shibatae, S. solfataricus, S. acidocaldarius, and P. furiosus with high sequence identity (>50%) to the T. uzoniensis β-galactosidase are tetramers, whereas those from Caldicellulosiruptor owensensis [27], Caldicellulosiruptor saccharolyticus [33], and Thermotoga petrophila [34] with low sequence identity (<30%) exist as dimers. These results indicate that within the GH1 family, the level of sequence identity is not critical in determining the association form.
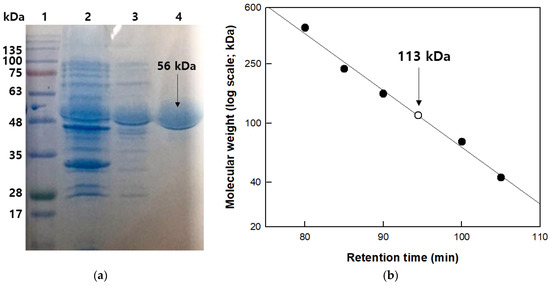
Figure 1.
Molecular mass determination of β-galactosidase from T. uzoniensis. (a) SDS-PAGE analysis. Lane 1, molecular weight size marker; lane 2, cell debris; lane 3, crude extract; and lane 4, purified enzyme. (b) Determination of native molecular mass using gel-filtration chromatography. The reference proteins ferritin (440 kDa), catalase (232 kDa), aldolase (158 kDa), conalbumin (75 kDa), and ovalbumin (43 kDa), and purified β-galactosidase from T. uzoniensis are represented by filled circles and open circles, respectively.
3.2. Effects of pH and Temperature on the Hydrolytic Activity of β-Galactosidase from T. uzoniensis
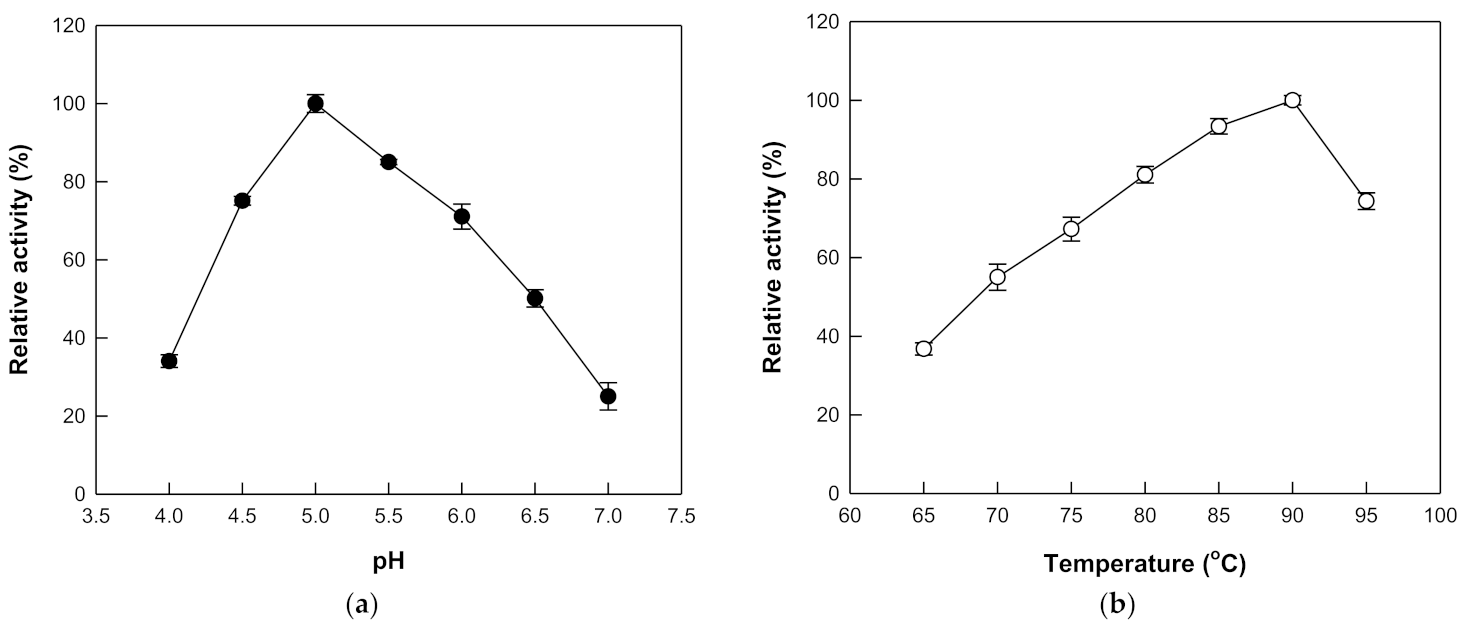
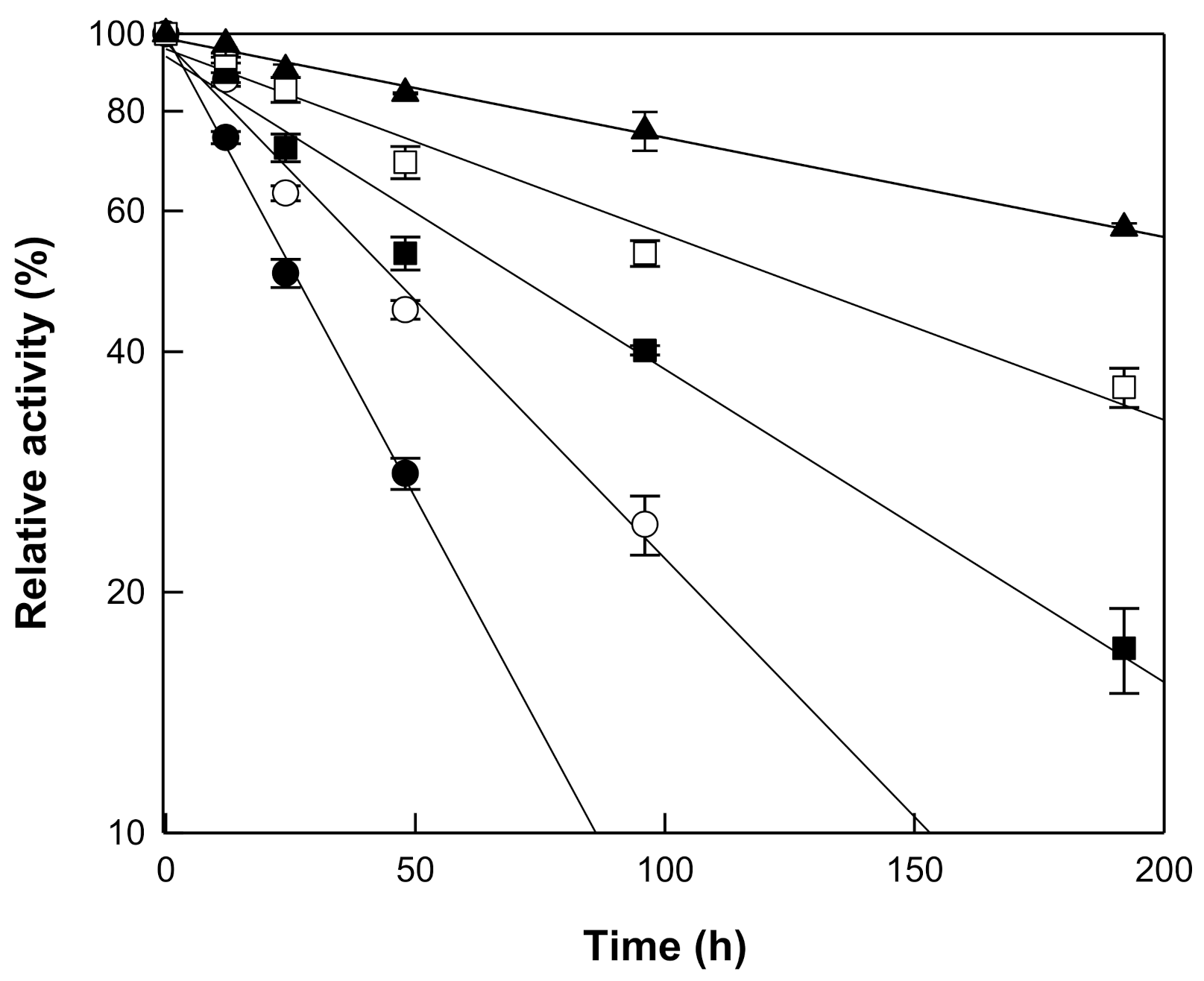
The hydrolytic activity of β-galactosidase from T. uzoniensis was examined in the pH range 4.0 to 7.0 and the maximal activity was observed at pH 5.0 (Figure 2a). At pH 4.5 and 6.0, the hydrolytic activity was approximately 70% of the maximum. The temperature was varied from 65 to 95 °C at pH 5.0, where the maximal activity was observed, to investigate its effect on the hydrolytic activity of β-galactosidase from T. uzoniensis, and the activity was maximal at 90 °C (Figure 2b). Thermophilic β-glucosidases from M. thermotolerans [11], T. leycettanus [15], N. fischeri [12], D. turgidum [10], T. pendens [16], S. solfataricus [14], and P. furiosus [13], which were applied to the hydrolysis of isoflavone glycosides, showed maximal activities at 50, 65, 80, 85, 90, 90, and 95 °C, respectively. The thermal stability of the enzyme was examined in the temperature range of 75–95 °C. First-order kinetics were displayed for thermal inactivation and the half-lives of β-galactosidase from T. uzoniensis were 235, 122, 68, 46, and 26 h at 75, 80, 85, 90, and 95 °C, respectively (Figure 3). Among the enzymes applied to the hydrolysis of isoflavone glycosides, β-glucosidases from P. furiosus [13] and S. solfataricus [14] had the highest thermal stabilities, respectively, displaying half-lives of 476, 307, 181, 96, and 59 h and 80, 51, 31, 18, and 10 h, at 75, 80, 85, 90, and 95 °C, respectively. At 90 °C, the half-lives of β-galactosidase from T. uzoniensis were 2.1-fold lower and 2.6-fold higher than those of β-glucosidases from P. furiosus and S. solfataricus, respectively.
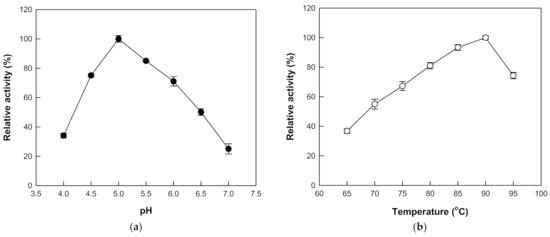
Figure 2.
Effects of pH and temperature on the activity of β-galactosidase from T. uzoniensis. (a) Effect of pH. The reactions were performed at the indicated pH values with 1 mM pNP-β-D-glucopyranoside and 0.005 mg/mL of enzyme at 70 °C for 10 min. (b) Effect of temperature. The reactions were performed at different temperatures in 50 mM CPB (pH 5.0) containing 1 mM pNP-β-D-glucopyranoside and 0.005 mg/mL of enzyme for 10 min. Data are represented as the means of triplicate experiments and error bars represent the standard deviation.
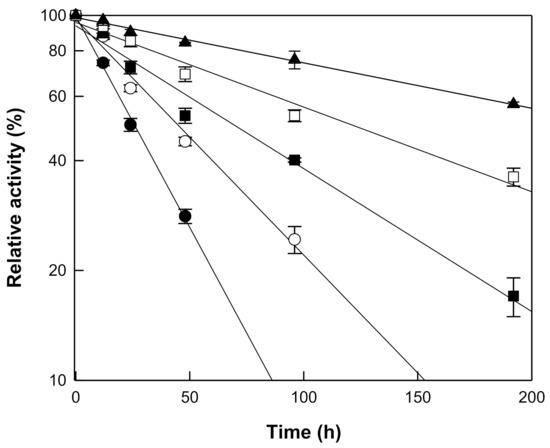
Figure 3.
Thermal inactivation of β-galactosidase from T. uzoniensis. The enzyme was incubated at 75 (closed triangle), 80 (open square), 85 (closed square), 90 (open circle), and 95 °C (closed circle) in 50 mM CPB (pH 5.0) for different time periods and the residual activity was determined. Data are represented as the means of thriplicate experiments and error bars represent the standard deviation.
3.3. Substrate Specificity of β-Galactosidase from T. uzoniensis for Aryl Glycosides and Isoflavone Glycosides
The specific activity of β-galactosidase from T. uzoniensis was investigated using aryl glycosides and isoflavone glycosides. The enzyme exhibited specific activities on aryl-glycosides as substrates in the order oNP-β-D-galactopyranoside > pNP-β-D-galactopyranoside > pNP-β-D-glucopyranoside > oNP-β-D-glucopyranoside > pNP-β-D-xylopyranoside (Table 1). However, no activity was observed on pNP-α-L-arabinopyranoside and pNP-α-L-rhamnopyranoside. These results indicated that this enzyme is a β-galactosidase with higher specificity for the β-1,2 linkage than for the β-1,4 linkage. The substrate specificity of β-galactosidase from T. uzoniensis for isoflavone glycosides followed the order: genistin > daidzin > ononin > glycitin (Table 1). Most other glycosidases from Dalbergia cohinchinensis [35], D. turgidum [10], P. furiosus [13], Paecilomyces thermophila [36], T. leycettanus [15], and T. maritima [17] also showed a higher specific activity for genistin than for daidzin as a substrate. Conversely, the substrate specificity of β-glucosidase from N. fischeri [12] was higher for daidzin than for genistin.

Table 1.
Substrate specificity of β-galactosidase from T. Uuzoniensis a.
Among the enzymes used for the hydrolysis of isoflavone glycosides, β-glucosidase from N. fischeri showed the highest specific activity (2189 μmol/min/mg) for pNP-β-D-glucopyranoside [12], followed by β-glucosidase from P. furiosus with a specific activity of 912 μmol/min/mg [13], which was 2.14-fold higher and 1.12-fold lower than that of β-galactosidase from T. uzoniensis, respectively. Nevertheless, the specific activities of β-galactosidase from T. uzoniensis were highest for daidzin and genistin and were 1.69- and 1.23-fold higher, respectively, than those of β-glucosidase from P. furiosus, with the highest activity so far [13]. These results indicate that β-galactosidase from T. uzoniensis is an enzyme suitable for the hydrolysis of isoflavone glycosides.
3.4. Conversion of Reagent Grade Isoflavone Glycosides into Isoflavone Aglycones by β-Galactosidase from T. uzoniensis
The time-course reactions for the conversion of daidzin and genistin, isoflavone glycosides abundantly expressed in soybeans, into daidzein and genistein were carried out under the conditions of pH 5.0, 90 °C, 2 mM substrate, and 0.2 mg/mL β-galactosidase from T. uzoniensis. Conversions were investigated by varying the enzyme concentration from 0.05 to 1 mg/mL for 2 mM reagent grade daidzin or genistin for 60 min. The production of daidzein or genistein increased with increasing enzyme concentrations up to 0.2 mg/mL and reached a plateau above that concentration (data not shown). Therefore, the optimal enzyme concentration for 2 mM reagent grade daidzein or genistein was 0.2 mg/mL.
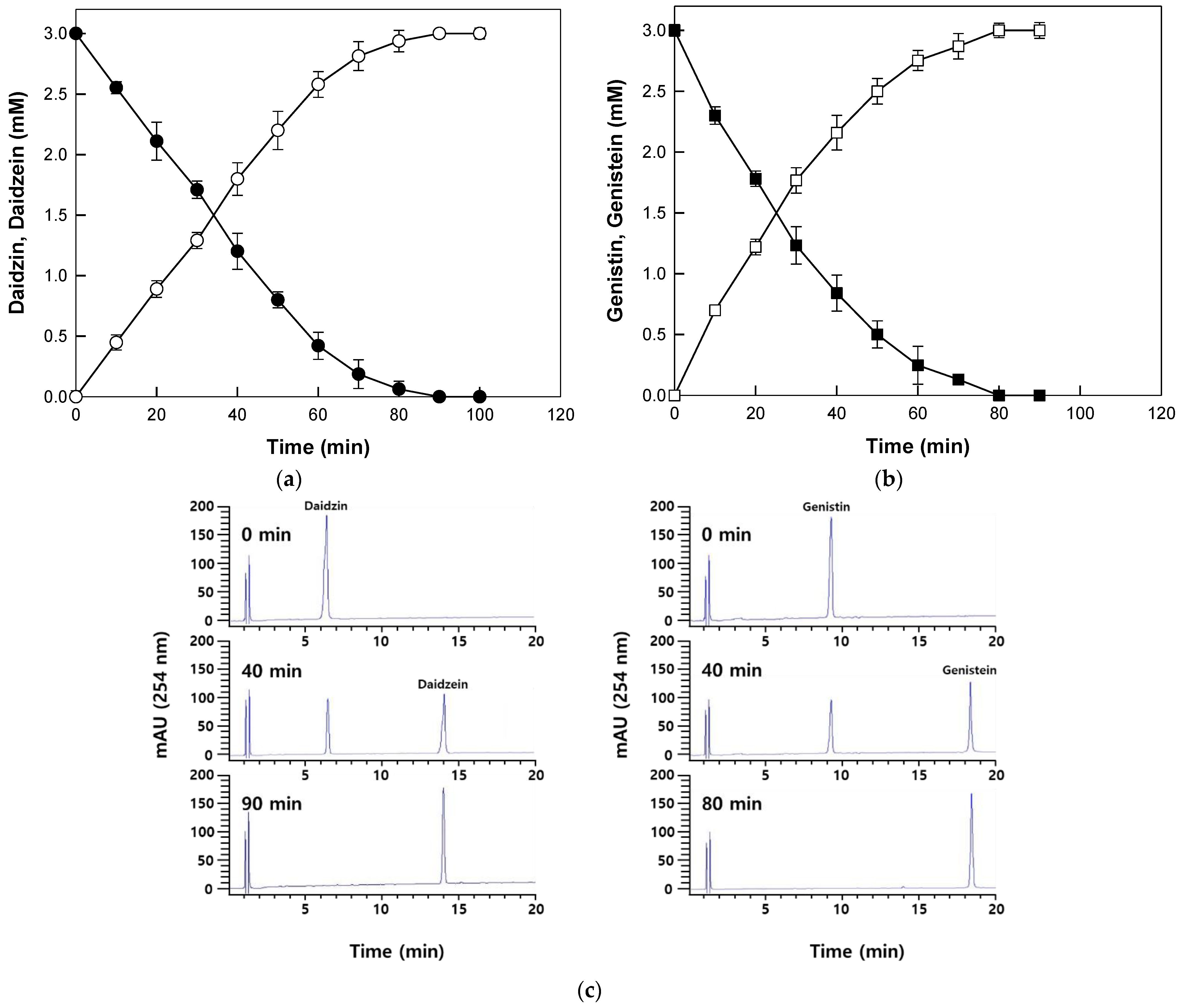
The enzyme completely hydrolyzed 2 mM reagent grade daidzin and genistin to daidzein and genistein within 90 min and 80 min, respectively (Figure 4a,b). The HPLC profiles for the conversion of daidzin and genistin into daidzein and genistein are shown in Figure 4c. The productivities for daidzein and genistein corresponded to 1.33 and 1.50 mM/h, which were 3.1- and 2.5-fold higher than those achieved with β-glucosidase from P. furiosus for isoflavone aglycones from soybean flour extract [13]. Therefore, β-galactosidase from T. uzoniensis was further applied to the production of isoflavone aglycones from extracts of Korean wild soybean.
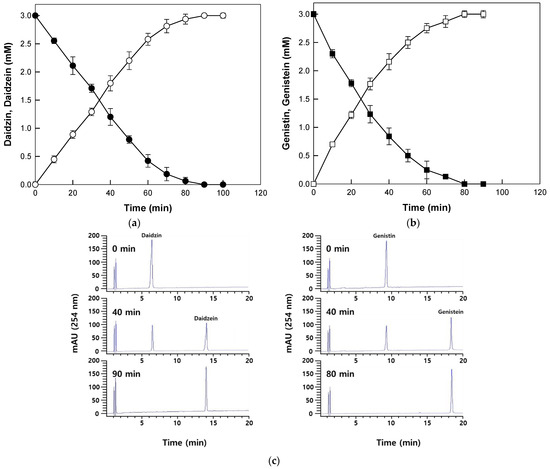
Figure 4.
Time-course reactions for the biotransformation of reagent grade daidzin and genistin into daidzein and genistein by β-galactosidase from T. Uzoniensis. (a) Biotransformation of daidzin (open circle) into daidzein (closed circle). (b) Biotransformation of genistin (open square) into genistein (closed square). (c) HPLC profiles for the biotransformation of daidzin and genistin into daidzein and genistein. The reactions were performed at 90 °C in 50 mM CPB (pH 5.0) containing 3 mM isoflavone substrate and 0.2 mg/mL enzyme. Data are represented as the means of thriplicate experiments and error bars represent the standard deviation.
3.5. Production of Ioflavone Aglycones from Isoflavone Glycosides in Seed and Root Extracts of Korean Wild Soybeans by β-Galactosidase from T. uzoniensis
Isoflavones were extracted from the seeds, stems, and roots of Korean wild soybeans and the amount of isoflavones was measured (Table 2). In Table 2, statistical steps through parametric or non-parametric tests were not performed because it was only to indicate the content of isoflavones required for biotransformation. Large amounts of daidzin (20,580 mg/g) and genistin (3600 mg/g) were found in the root and seed extracts, whereas no glycitin was found in any of the extracts. The root extract not only contained a 1.7-fold higher isoflavone glycoside content than that of the seed extract but also contained a high content of isoflavone aglycones, including 7560 mg/g of daidzein and 1770 mg/g of genistein. In contrast, the stem extract contained only genistin as an isoflavone glycoside without daidzin, which was only about 16% of the genistin contained in the seed extract. Therefore, we applied β-galactosidase from T. uzoniensis to the production of daidzein and genistein from daidzin and genistin in the seed and root extracts of Korean wild soybeans. The production was investigated by varying the enzyme concentration from 0.1 to 2 mg/mL for 20% (w/v) seed or root extracts for 60 or 90 min. The daidzein or genistein yield increased with increasing enzyme concentrations up to 0.4 mg/mL and decreased considerably at higher enzyme concentrations (data not shown), indicating that the optimal enzyme concentration for 20% (w/v) seed or root extracts was 0.4 mg/mL.
Table 2.
Isoflavone contents in 10% (w/v) seed, stem, and root extracts of Korean wild soybean a.
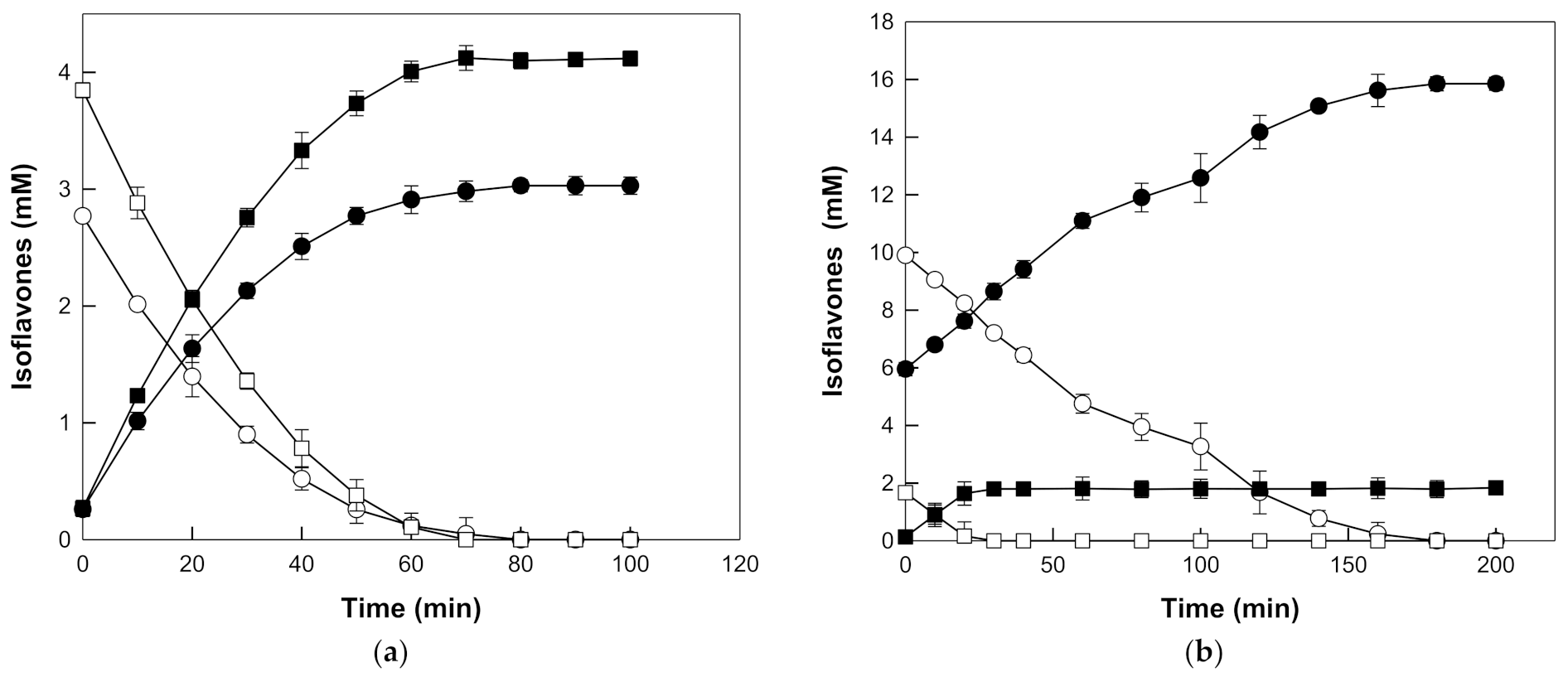
The time-course reactions for the production of daidzein and genistein from daidzin and genistin in the extracts were carried out at pH 5.0 and 90 °C with 20% (w/v) seed or root extract, and 0.4 mg/mL β-galactosidase from T. uzoniensis. The seed and root extracts at 20% (w/v) contained 6.62 mM and 11.56 mM isoflavone glycosides (2.77 mM daidzin and 3.85 mM genistin, and 9.89 mM daidzin and 1.67 mM genistin) as substrates and 0.53 mM and 6.08 mM of the corresponding aglycones (0.26 mM daidzein and 0.27 mM genistein, and 5.95 mM daidzein and 0.13 mM genistein), respectively. Daidzin and genistin in the seed extract were completely hydrolyzed to daidzein and genistein within 80 and 70 min, corresponding to the productivities of 1.86 and 3.30 mM/h, respectively. Daidzein and genistein were produced from daidzin and genistin in the root extract with higher productivities of 3.30 and 3.36 mM/h, respectively, and molar yields of 100%, after 180 min and 30 min, respectively (Figure 5).
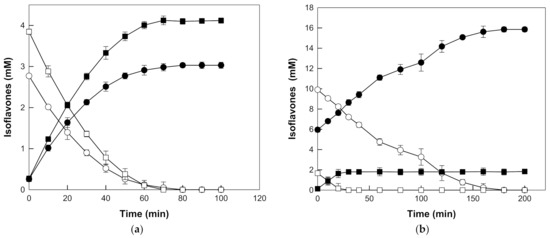
Figure 5.
Time-course reactions for the production of daidzein (closed circle) and genistein (closed square) from daidzin (open circle) and genistin (open square), respectively, in the (a) seed and (b) root extracts of Korean wild soybean by β-galactosidase from T. uzoniensis. The reactions were performed at 90 °C in 50 mM CPB (pH 5.0) containing 0.4 mg/mL enzyme and 20% (v/v) seed or root extracts of Korean wild soybean. Data are represented as the means of thriplicate experiments and error bars represent the standard deviation.
The production of daidzein and genistein by glycosidases from complex substrates is summarized in Table 3. The previous highest concentrations of the produced daidzein and genistein were 8.40 and 4.09 mM, respectively, using β-glucosidase from Aspergillus oryzae with soybean flour extract [37]. These values were 1.18-fold lower and 1.07-fold higher than those of daidzein and genistein from root and seed extracts of Korean wild soybean, respectively. However, the levels of daidzein and genistein produced using β-glucosidase from A. oryzae were 19- and 42-fold lower, respectively, than those produced using β-galactosidase from T. uzoniensis with the root extract of Korean wild soybean. The previous highest productivities obtained for daidzein and genistein were 1.50 and 1.23 mM/h, respectively, using β-glucosidase from Gongronella sp. with soybean flour extract [38]. These values are 2.2- and 2.7-fold lower, respectively, than those obtained using β-galactosidase from T. uzoniensis with the root extract of Korean wild soybean in this study. β-Galactosidase from T. uzoniensis completely hydrolyzed daidzin and genistin in seed and root extracts of Korean wild soybean to daidzein and genistein and the productivities were the highest reported to date. Thus, β-galactosidase from T. uzoniensis is a potent enzyme for the production of isoflavone aglycones.
Table 3.
Production of daidzein and genistein from complex substrates by glycosidases.
4. Conclusions
In conclusion, a thermostable β-galactosidase from T. uzoniensis was cloned and characterized as a purified recombinant enzyme, which exhibited a maximum specific activity of 1103 μmol/min/mg for pNP-β-D-galactopyranoside at pH 5.0 and 90 °C with a half-life of 46 h and exists as a homodimer of 113 kDa. The enzyme produced daidzein and genistein from daidzin and genistin in the seed and root extracts of Korean wild soybean with productivities of 1.86 and 3.30 mM/h, respectively. To the best of our knowledge, this enzyme exhibits the highest productivity for both daidzein and genistein of any enzyme reported to date. These results indicate that β-galactosidase from T. uzoniensis may be industrially useful for the hydrolysis of isoflavone glycosides and that wild soybeans are valuable materials for isoflavone aglycone production.
5. Patents
This section is not mandatory but may be added if there are patents resulting from the work reported in this manuscript.
Author Contributions
Conceptualization, K.-C.S., C.S.N. and Y.-S.K.; methodology, K.-C.S. and D.-K.O.; validation, D.W.K.; formal analysis, S.-H.K. and D.W.K.; investigation, K.-C.S. and S.-H.K.; resources, S.-H.K.; data curation, D.-K.O. and S.H.K.; writing—original draft preparation, K.-C.S.; writing—review and editing, C.S.N. and Y.-S.K.; visualization, K.-C.S.; supervision, C.S.N. and Y.-S.K.; project administration, C.S.N. and Y.-S.K.; funding acquisition, K.-C.S. and C.S.N. All authors have read and agreed to the published version of the manuscript.
Funding
This work was supported by the Basic Science Research Program through the National Research Foundation of Korea (NRF) funded by the Ministry of Education (NRF-2020R1I1A1A01063145) and the R&D Program for Forest Science Technology, Korea Forest Service (Korea Forestry Promotion Institute; 2021400A00-2125-CA02).
Institutional Review Board Statement
Not applicable.
Informed Consent Statement
Not applicable.
Data Availability Statement
Not applicable.
Acknowledgments
We would like to express our gratitude for the support of the KU Research Professor Program of Konkuk University.
Conflicts of Interest
The authors declare no conflict of interest. The funders had no role in the design of the study; in the collection, analyses, or interpretation of data; in the writing of the manuscript, or in the decision to publish the results.
References
- Zhao, T.T.; Jin, F.; Li, J.G.; Xu, Y.Y.; Dong, H.T.; Liu, Q.; Xing, P.; Zhu, G.L.; Xu, H.; Miao, Z.F. Dietary isoflavones or isoflavone-rich food intake and breast cancer risk: A meta-analysis of prospective cohort studies. Clin. Nutr. 2019, 38, 136–145. [Google Scholar] [CrossRef] [PubMed]
- Barańska, A.; Błaszczuk, A.; Kanadys, W.; Baczewska, B.; Jędrych, M.; Wawryk-Gawda, E.; Polz-Dacewicz, M. Effects of soy protein containing of isoflavones and isoflavones extract on plasma lipid profile in postmenopausal women as a potential prevention factor in cardiovascular diseases: Systematic review and meta-analysis of randomized controlled trials. Nutrients 2021, 13, 2531. [Google Scholar] [CrossRef] [PubMed]
- Jagga, S.; Sharma, A.R.; Kim, E.J.; Nam, J.S. Isoflavone-enriched whole soy milk powder stimulates osteoblast differentiation. J. Food Sci. Technol. 2021, 58, 595–603. [Google Scholar] [CrossRef] [PubMed]
- Chen, L.R.; Chen, K.H. Utilization of isoflavones in soybeans for women with menopausal syndrome: An overview. Int. J. Mol. Sci. 2021, 22, 3212. [Google Scholar] [CrossRef] [PubMed]
- Kawakami, Y.; Tsurugasaki, W.; Nakamura, S.; Osada, K. Comparison of regulative functions between dietary soy isoflavones aglycone and glucoside on lipid metabolism in rats fed cholesterol. J. Nutr. Biochem. 2005, 16, 205–212. [Google Scholar] [CrossRef]
- Izumi, T.; Piskula, M.K.; Osawa, S.; Obata, A.; Tobe, K.; Saito, M.; Kataoka, S.; Kubota, Y.; Kikuchi, M. Soy isoflavone aglycones are absorbed faster and in higher amounts than their glucosides in humans. J. Nutr. 2000, 130, 1695–1699. [Google Scholar] [CrossRef] [Green Version]
- Chun, J.; Kim, G.M.; Lee, K.W.; Choi, I.D.; Kwon, G.H.; Park, J.Y.; Jeong, S.J.; Kim, J.S.; Kim, J.H. Conversion of isoflavone glucosides to aglycones in soymilk by fermentation with lactic acid bacteria. J. Food Sci. 2007, 72, M39–M44. [Google Scholar] [CrossRef]
- Kuo, L.C.; Cheng, W.Y.; Wu, R.Y.; Huang, C.J.; Lee, K.T. Hydrolysis of black soybean isoflavone glycosides by Bacillus subtilis natto. Appl. Microbiol. Biotechnol. 2006, 73, 314–320. [Google Scholar] [CrossRef]
- Ibe, S.; Kumada, K.; Yoshiba, M.; Onga, T. Production of natto which contains a high level of isoflavone aglycons. J. Jpn. Soc. Food Sci. Technol.-Nippon Shokuhin Kagaku Kogaku Kaishi 2001, 48, 27–34. [Google Scholar] [CrossRef]
- Kim, Y.S.; Yeom, S.J.; Oh, D.K. Characterization of a GH3 family β-glucosidase from Dictyoglomus turgidum and its application to the hydrolysis of isoflavone glycosides in spent coffee grounds. J. Agric. Food Chem. 2011, 59, 11812–11818. [Google Scholar] [CrossRef]
- Pyeon, H.M.; Lee, Y.S.; Choi, Y.L. Cloning, purification, and characterization of GH3 β-glucosidase, MtBgl85, from Microbulbifer thermotolerans DAU221. PeerJ 2019, 7, e7106. [Google Scholar] [CrossRef] [Green Version]
- Yang, X.; Ma, R.; Shi, P.; Huang, H.; Bai, Y.; Wang, Y.; Yang, P.; Fan, Y.; Yao, B. Molecular characterization of a highly-active thermophilic β-glucosidase from Neosartorya fischeri P1 and its application in the hydrolysis of soybean isoflavone glycosides. PLoS ONE 2014, 9, e106785. [Google Scholar] [CrossRef]
- Yeom, S.J.; Kim, B.N.; Kim, Y.S.; Oh, D.K. Hydrolysis of isoflavone glycosides by a thermostable β-glucosidase from Pyrococcus furiosus. J. Agric. Food Chem. 2012, 60, 1535–1541. [Google Scholar] [CrossRef]
- Kim, B.N.; Yeom, S.J.; Kim, Y.S.; Oh, D.K. Characterization of a β-glucosidase from Sulfolobus solfataricus for isoflavone glycosides. Biotechnol. Lett. 2012, 34, 125–129. [Google Scholar] [CrossRef]
- Li, X.; Xia, W.; Bai, Y.; Ma, R.; Yang, H.; Luo, H.; Shi, P. A novel thermostable GH3 β-glucosidase from Talaromyce leycettanus with broad substrate specificity and significant soybean isoflavone glycosides-hydrolyzing capability. Biomed Res. Int. 2018, 2018, 4794690. [Google Scholar] [CrossRef] [Green Version]
- Li, D.; Li, X.; Dang, W.; Tran, P.L.; Park, S.H.; Oh, B.C.; Hong, W.S.; Lee, J.S.; Park, K.H. Characterization and application of an acidophilic and thermostable β-glucosidase from Thermofilum pendens. J. Biosci. Bioeng. 2013, 115, 490–496. [Google Scholar] [CrossRef]
- Xue, Y.; Yu, J.; Song, X. Hydrolysis of soy isoflavone glycosides by recombinant β-glucosidase from hyperthermophile Thermotoga maritima. J. Ind. Microbiol. Biotechnol. 2009, 36, 1401–1408. [Google Scholar] [CrossRef]
- Bonch-Osmolovskaya, E.A.; Miroshnichenko, M.L.; Kostrikina, N.A.; Chernych, N.A.; Zavarzin, G.A. Thermoproteus uzoniensis sp. nov., a new extremely thermophilic archaebacterium from Kamchatka continental hot springs. Arch. Microbiol. 1990, 154, 556–559. [Google Scholar] [CrossRef]
- Aguirre, A.; Peiru, S.; Eberhardt, F.; Vetcher, L.; Cabrera, R.; Menzella, H.G. Enzymatic hydrolysis of steryl glucosides, major contaminants of vegetable oil-derived biodiesel. Appl. Microbiol. Biotechnol. 2014, 98, 4033–4040. [Google Scholar] [CrossRef]
- Tepavcevic, V.; Cvejic, J.; Posa, M.; Popovic, J. Isoflavone Content and Composition in Soybean; IntechOpen: London, UK, 2011. [Google Scholar]
- Hyten, D.L.; Song, Q.; Zhu, Y.; Choi, I.Y.; Nelson, R.L.; Costa, J.M.; Specht, J.E.; Shoemaker, R.C.; Cregan, P.B. Impacts of genetic bottlenecks on soybean genome diversity. Proc. Natl. Acad. Sci. USA 2006, 103, 16666–16671. [Google Scholar] [CrossRef] [Green Version]
- Nawaz, M.A.; Golokhvast, K.S.; Rehman, H.M.; Tsukamoto, C.; Kim, H.S.; Yang, S.H.; Chung, G. Soyisoflavone diversity in wild soybeans (Glycine soja Sieb. & Zucc.) from the main centres of diversity. Biochem. Syst. Ecol. 2018, 77, 16–21. [Google Scholar]
- Lee, S.J.; Ahn, J.K.; Kim, S.H.; Kim, J.T.; Han, S.J.; Jung, M.Y.; Chung, I.M. Variation in isoflavone of soybean cultivars with location and storage duration. J. Agric. Food Chem. 2003, 51, 3382–3389. [Google Scholar] [CrossRef]
- Lee, Y.W.; Kim, J.D.; Zheng, J.; Row, K. Comparisons of isoflavones from Korean and Chinese soybean and processed product. Biochem. Eng. J. 2007, 36, 49–53. [Google Scholar] [CrossRef]
- Lee, J.S.; Kim, H.S.; Hwang, T.Y. Variation in protein and isoflavone contents of collected domestic and foreign soybean (Glycine max (L.) Merrill) germplasms in Korea. Agriculture 2021, 11, 735. [Google Scholar] [CrossRef]
- Gibson, D.G.; Young, L.; Chuang, R.Y.; Venter, J.C.; Hutchison, C.A., 3rd; Smith, H.O. Enzymatic assembly of DNA molecules up to several hundred kilobases. Nat. Methods 2009, 6, 343–345. [Google Scholar] [CrossRef]
- Shin, K.C.; Seo, M.J.; Kim, D.W.; Yeom, S.J.; Kim, Y.S. Characterization of β-glycosidase from Caldicellulosiruptor owensensis and its application in the production of platycodin D from balloon flower leaf. Catalysts 2019, 9, 1025. [Google Scholar] [CrossRef] [Green Version]
- Song, T.; Barua, K.; Buseman, G.; Murphy, P.A. Soy isoflavone analysis: Quality control and a new internal standard. Am. J. Clin. Nutr. 1998, 68, 1474s–1479s. [Google Scholar] [CrossRef]
- Park, N.Y.; Cha, J.; Kim, D.O.; Park, C.S. Enzymatic characterization and substrate specificity of thermostable β-glycosidase from hyperthermophilic archaea, Sulfolobus shibatae, expressed in E. coli. J. Microbiol. Biotechnol. 2007, 17, 454–460. [Google Scholar] [PubMed]
- D’Auria, S.; Morana, A.; Febbraio, F.; Vaccaro, C.; De Rosa, M.; Nucci, R. Functional and structural properties of the homogeneous β-glycosidase from the extreme thermoacidophilic archaeon Sulfolobus solfataricus expressed in Saccharomyces cerevisiae. Protein Expr. Purif. 1996, 7, 299–308. [Google Scholar] [CrossRef] [PubMed]
- D’Auria, S.; Nucci, R.; Rossi, M.; Bertoli, E.; Tanfani, F.; Gryczynski, I.; Malak, H.; Lakowicz, J.R. β-Glycosidase from the hyperthermophilic archaeon Sulfolobus solfataricus: Structure and activity in the presence of alcohols. J. Biochem. 1999, 126, 545–552. [Google Scholar] [CrossRef] [PubMed]
- Nakabayashi, M.; Kataoka, M.; Watanabe, M.; Ishikawa, K. Monomer structure of a hyperthermophilic β-glucosidase mutant forming a dodecameric structure in the crystal form. Acta Crystallogr. F-Struct. Biol. Commun. 2014, 70, 854–859. [Google Scholar] [CrossRef] [Green Version]
- Hong, M.R.; Kim, Y.S.; Park, C.S.; Lee, J.K.; Kim, Y.S.; Oh, D.K. Characterization of a recombinant β-glucosidase from the thermophilic bacterium Caldicellulosiruptor saccharolyticus. J. Biosci. Bioeng. 2009, 108, 36–40. [Google Scholar] [CrossRef]
- Colussi, F.; da Silva, V.M.; Miller, I.; Cota, J.; de Oliveira, L.C.; de Oliveira Neto, M.; Squina, F.M.; Garcia, W. Oligomeric state and structural stability of two hyperthermophilic β-glucosidases from Thermotoga petrophila. Amino Acids 2015, 47, 937–948. [Google Scholar] [CrossRef]
- Chuankhayan, P.; Rimlumduan, T.; Svasti, J.; Cairns, J.R. Hydrolysis of soybean isoflavonoid glycosides by Dalbergia β-glucosidases. J. Agric. Food Chem. 2007, 55, 2407–2412. [Google Scholar] [CrossRef]
- Yang, S.; Wang, L.; Yan, Q.J.; Jiang, Z.; Li, L. Hydrolysis of soybean isoflavone glycosides by a thermostable β-glucosidase from Paecilomyces thermophila. Food Chem. 2009, 115, 1247–1252. [Google Scholar] [CrossRef]
- Kaya, M.; Ito, J.; Kotaka, A.; Matsumura, K.; Bando, H.; Sahara, H.; Ogino, C.; Shibasaki, S.; Kuroda, K.; Ueda, M.; et al. Isoflavone aglycones production from isoflavone glycosides by display of β-glucosidase from Aspergillus oryzae on yeast cell surface. Appl. Microbiol. Biotechnol. 2008, 79, 51–60. [Google Scholar] [CrossRef]
- Fang, W.; Song, R.; Zhang, X.; Zhang, X.; Zhang, X.; Wang, X.; Fang, Z.; Xiao, Y. Characterization of a novel β-glucosidase from Gongronella sp. W5 and its application in the hydrolysis of soybean isoflavone glycosides. J. Agric. Food Chem. 2014, 62, 11688–11695. [Google Scholar] [CrossRef]
- Cao, H.; Zhang, Y.; Shi, P.; Ma, R.; Yang, H.; Xia, W.; Cui, Y.; Luo, H.; Bai, Y.; Yao, B. A highly glucose-tolerant GH1 β-glucosidase with greater conversion rate of soybean isoflavones in monogastric animals. J. Ind. Microbiol. Biotechnol. 2018, 45, 369–378. [Google Scholar] [CrossRef] [Green Version]
- Matsuura, M.; Obata, A. β-Glucosidases from soybeans hydrolyze daidzin and genistin. J. Food Sci. 1993, 58, 144–147. [Google Scholar] [CrossRef]
- Angelotti, J.A.F.; Dias, F.F.G.; Sato, H.H.; Fernandes, P.; Nakajima, V.M.; Macedo, J. Improvement of aglycone content in soy isoflavones extract by free and immobilized β-glucosidase and their effects in lipid accumulation. Appl. Biochem. Biotechnol. 2020, 192, 734–750. [Google Scholar] [CrossRef]
- Wu, J.; Muir, A.D. Isoflavone during protease hydrolysis of defatted soybean meal. Food Chem. 2010, 118, 328–332. [Google Scholar] [CrossRef]
- Abdella, A.; El-Baz, A.F.; Ibrahim, I.A.; Mahrous, E.E.; Yang, S.T. Biotransformation of soy flour isoflavones by Aspergillus niger NRRL 3122 β-glucosidase enzyme. Nat. Prod. Res. 2018, 32, 2382–2391. [Google Scholar] [CrossRef]
- Horii, K.; Adachi, T.; Matsuda, T.; Tanaka, T.; Sahara, H.; Shibasaki, S.; Ogino, C.; Hata, Y.; Ueda, M.; Kondo, A. Improvement of isoflavone aglycones production using β-glucosidase secretory produced in recombinant Aspergillus oryzae. J. Mol. Catal. B-Enzym. 2009, 59, 297–301. [Google Scholar] [CrossRef]
- Yan, F.Y.; Xia, W.; Zhang, X.X.; Chen, S.; Nie, X.Z.; Qian, L.C. Characterization of β-glucosidase from Aspergillus terreus and its application in the hydrolysis of soybean isoflavones. J. Zhejiang Univ. Sci. B 2016, 17, 455–464. [Google Scholar] [CrossRef] [Green Version]
- Mai, Z.; Wang, L.; Zeng, Q. Characterization of a novel isoflavone glycoside-hydrolyzing β-glucosidase from mangrove soil metagenomic library. Biochem. Biophys. Res. Commun. 2021, 569, 61–65. [Google Scholar] [CrossRef]
- Maitan-Alfenas, G.P.; Lorena, G.A.D.; de Almeida, M.N.; Visser, E.M.; de Rezende, S.T.; Guimarães, V.M. Hydrolysis of soybean isoflavones by Debaryomyces hansenii UFV-1 immobilised cells and free β-glucosidase. Food Chem. 2014, 146, 429–436. [Google Scholar] [CrossRef] [Green Version]
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).